
Highly-Sensitive In Vitro Bioassays for FSH, TSH, PTH, Kp, and OT in Addition to LH in Mouse Leydig Tumor Cell
 , and
, and 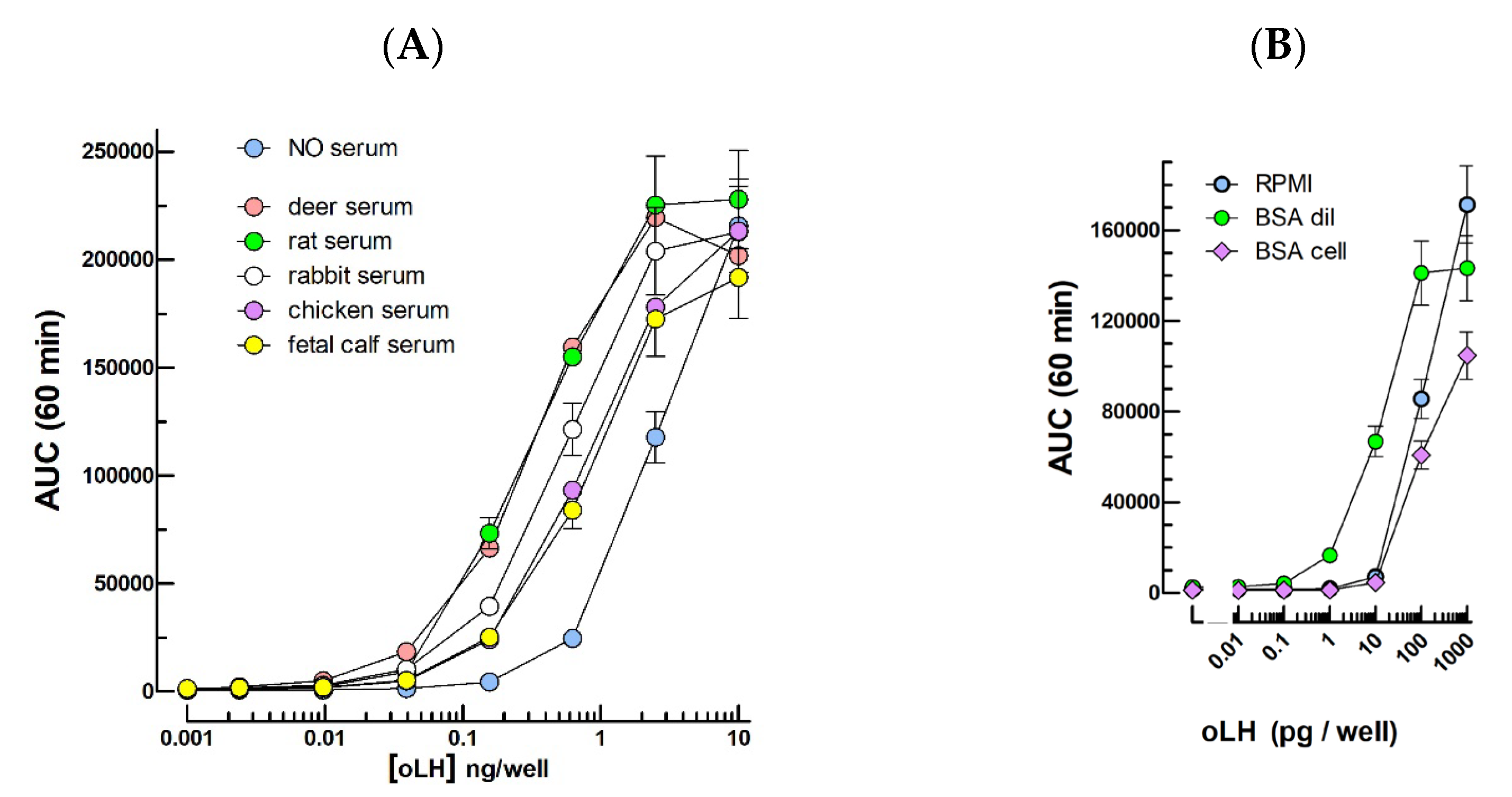{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LH Bioassays
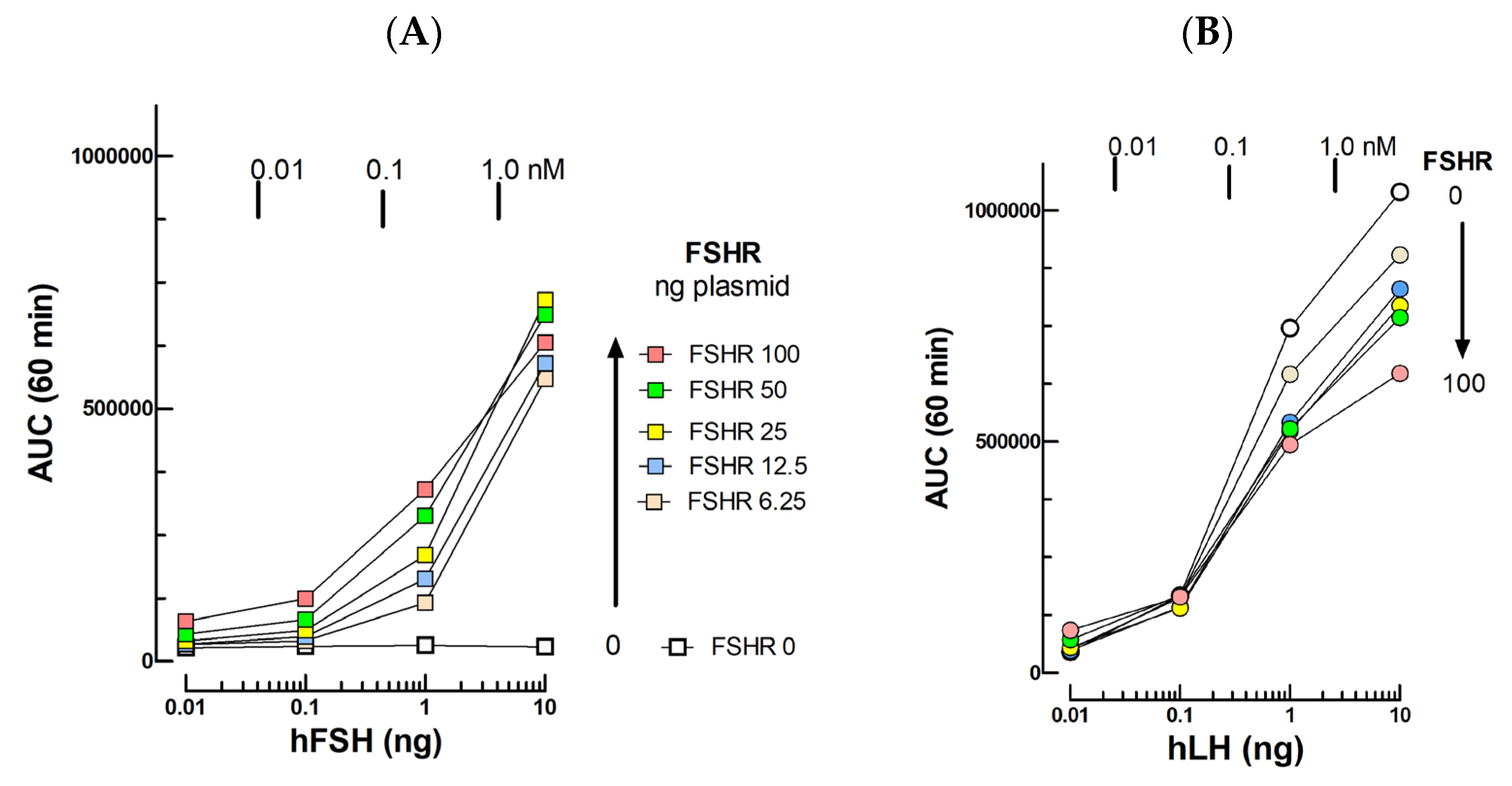
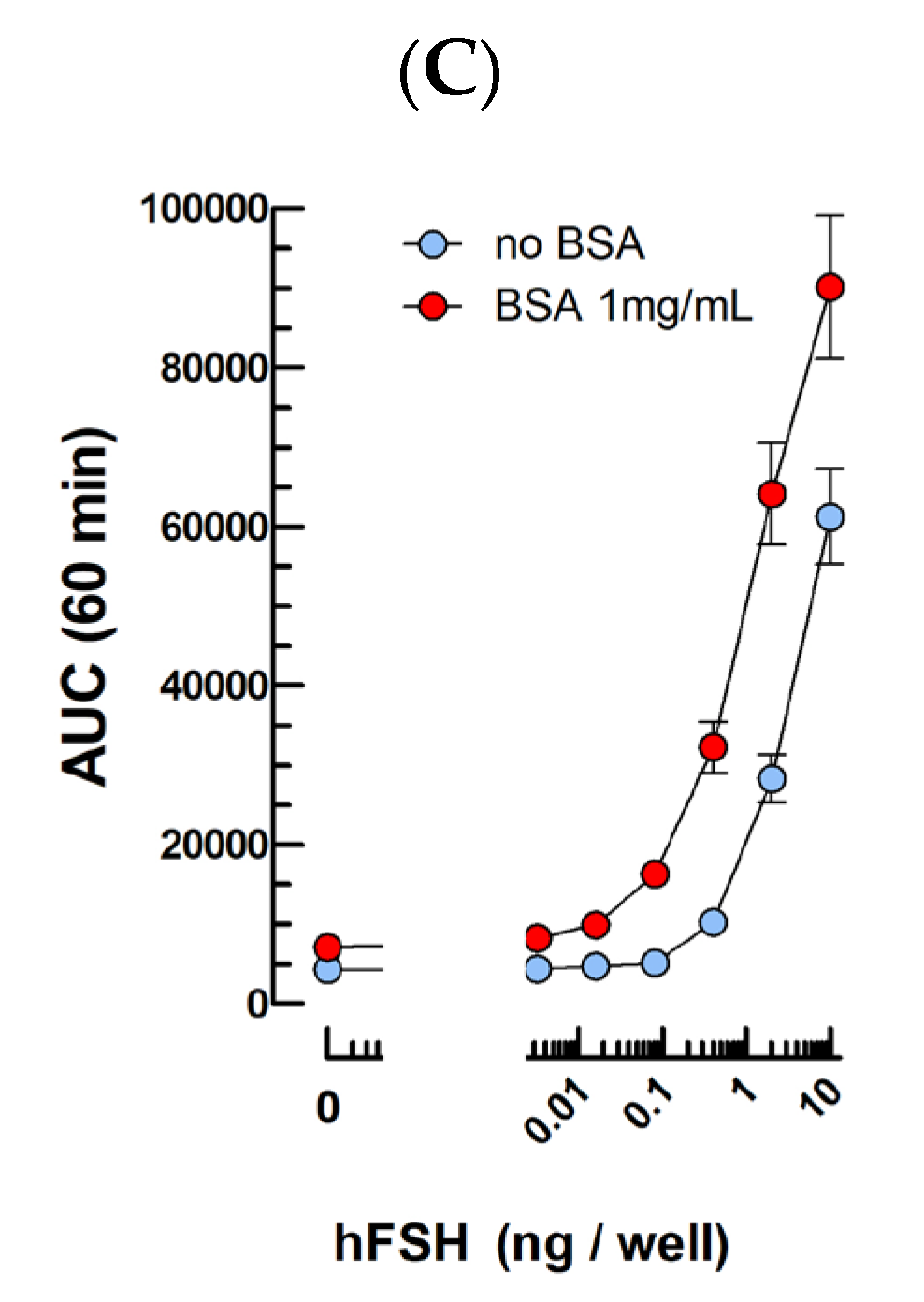
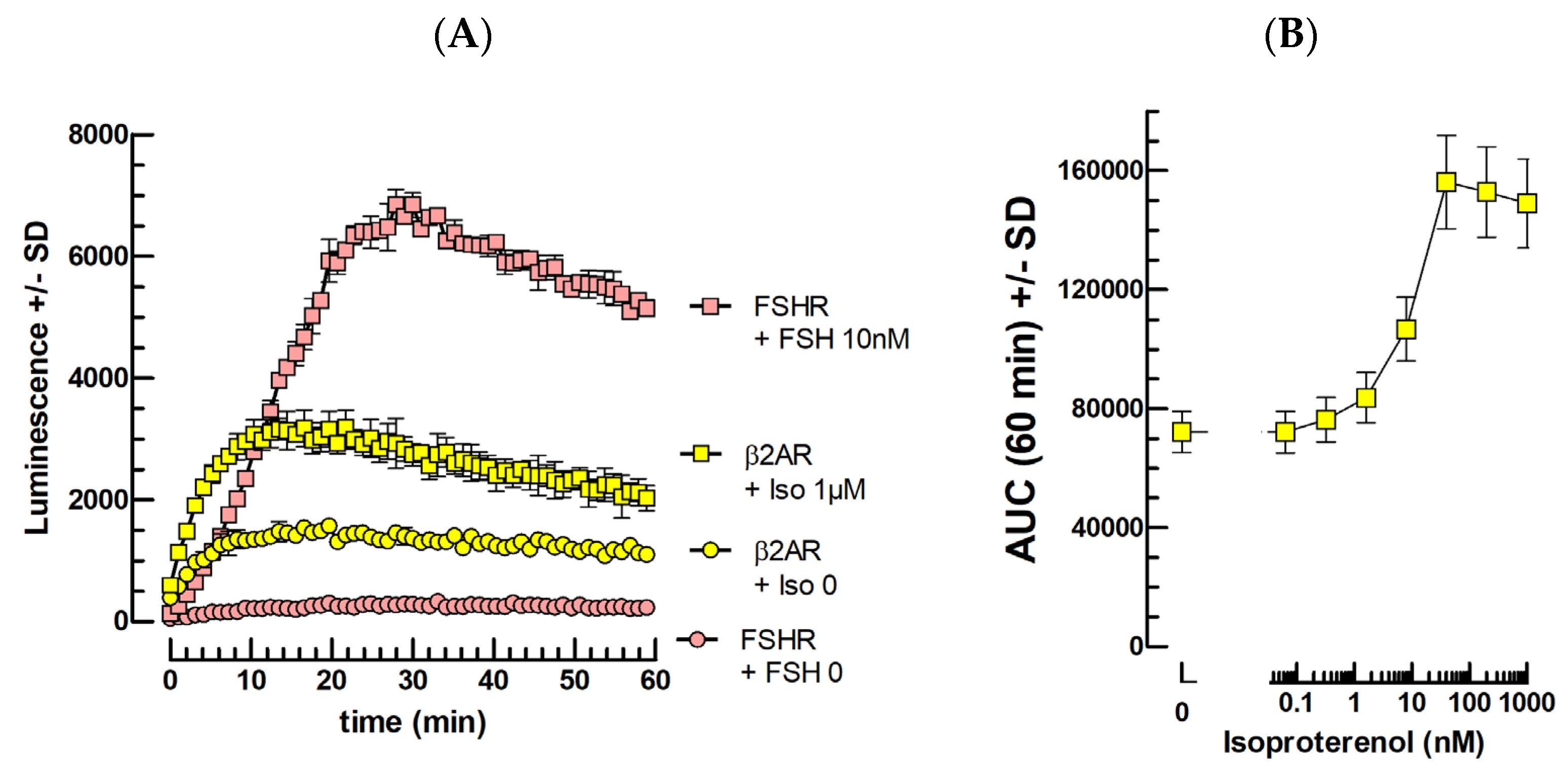
2.2. FSH Bioassays
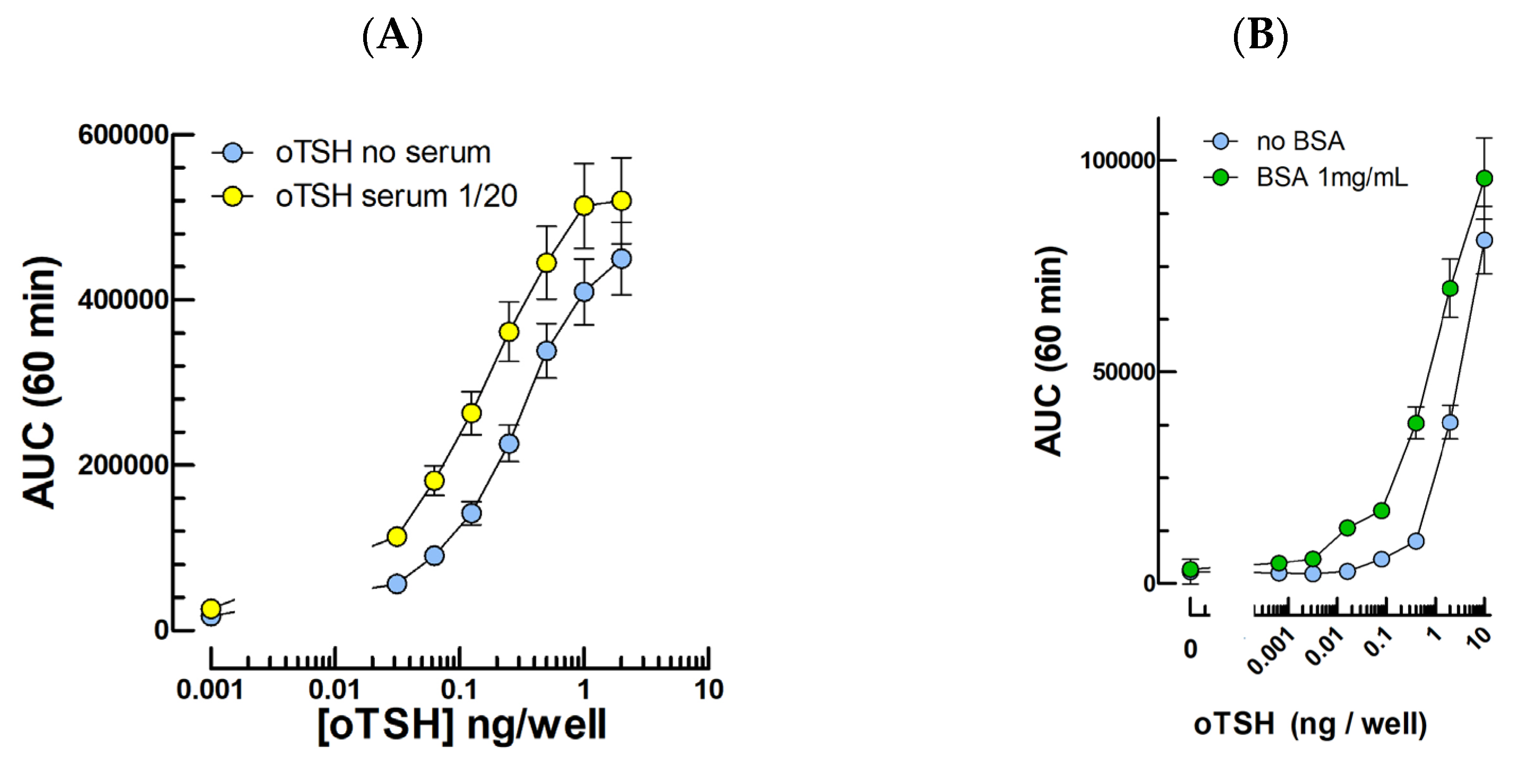
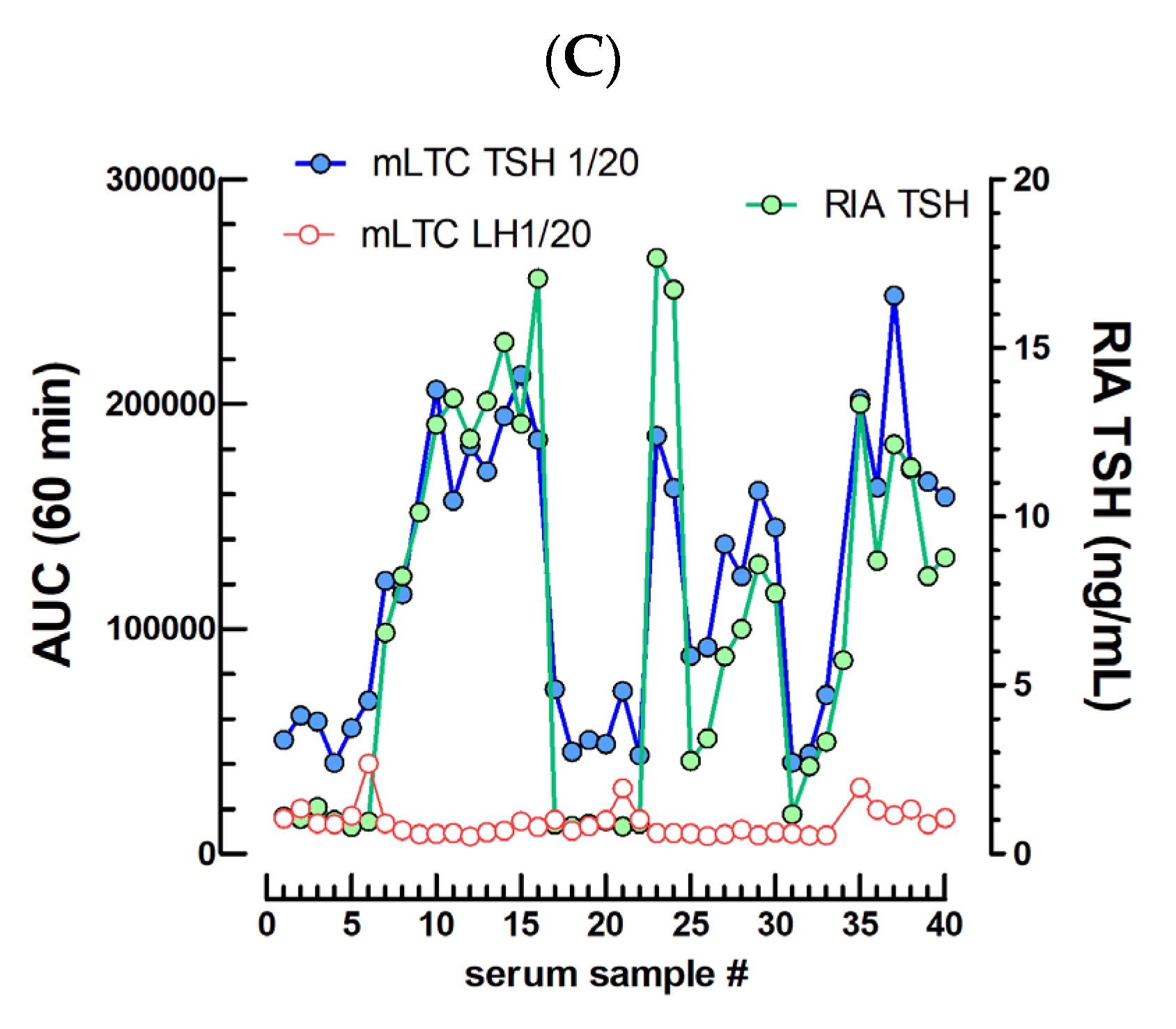
2.3. TSH Bioassays
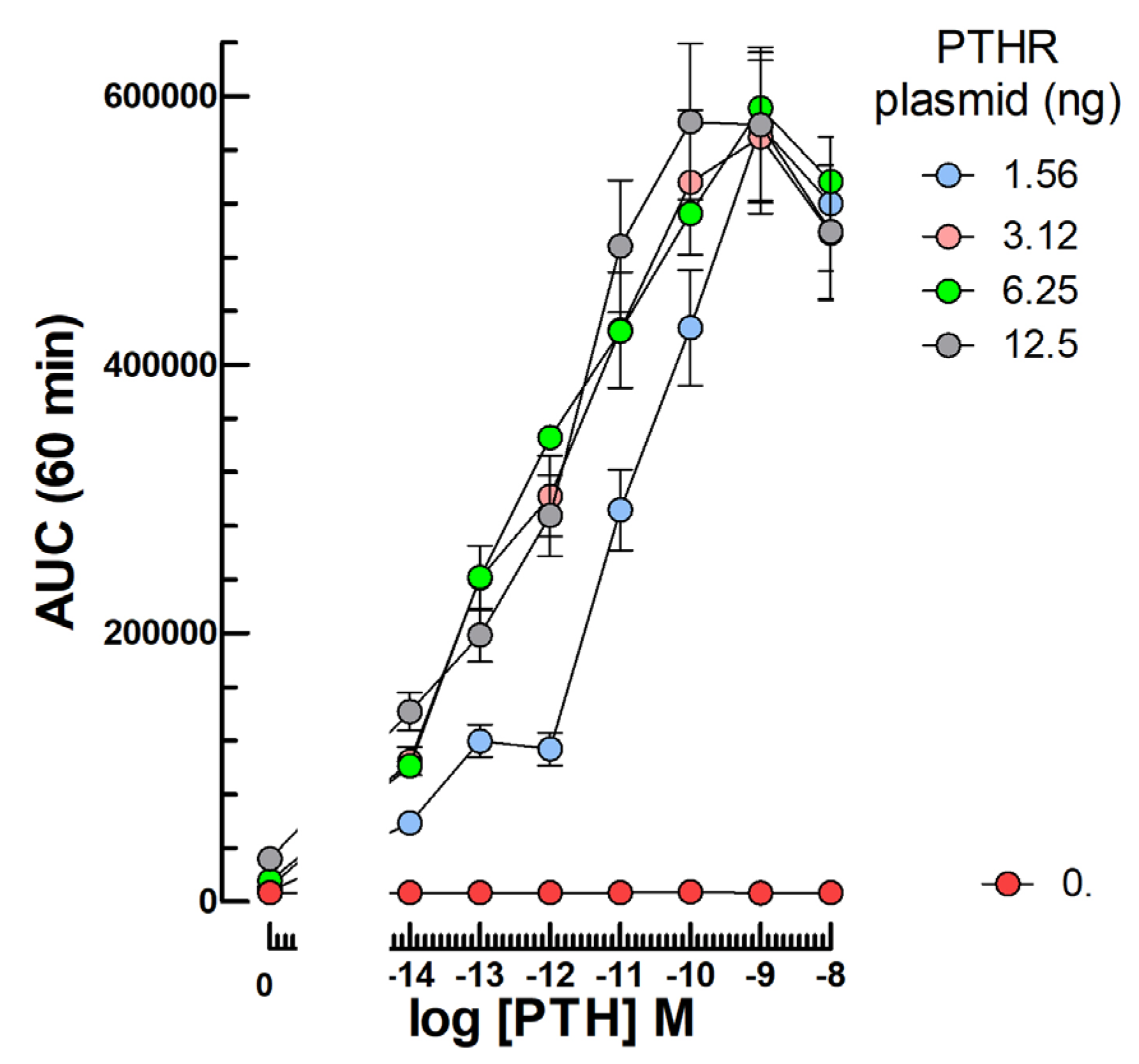
2.4. PTH Bioassays
2.5. Catecholamine Bioassays
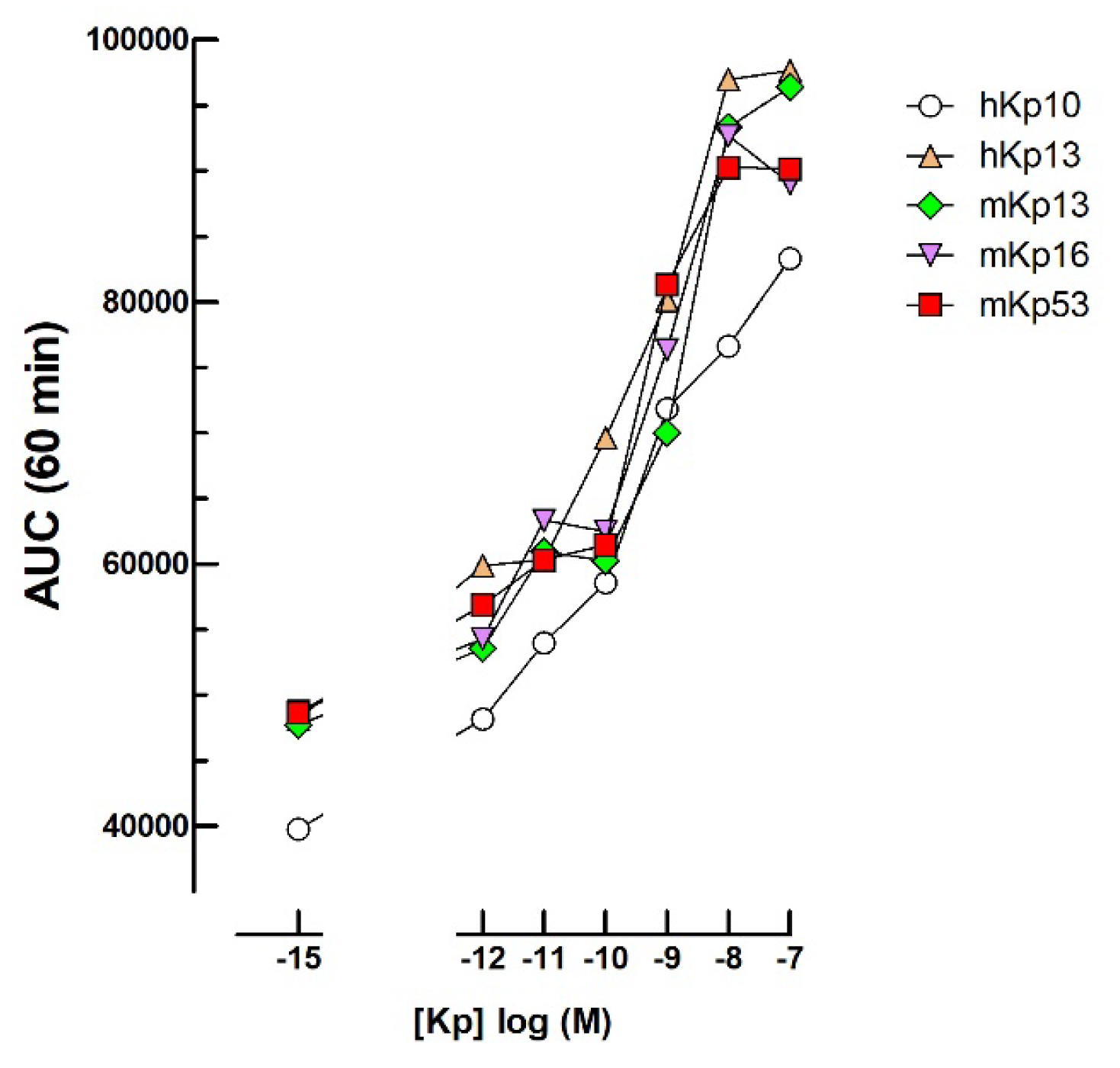
2.6. Kisspeptin Bioassays
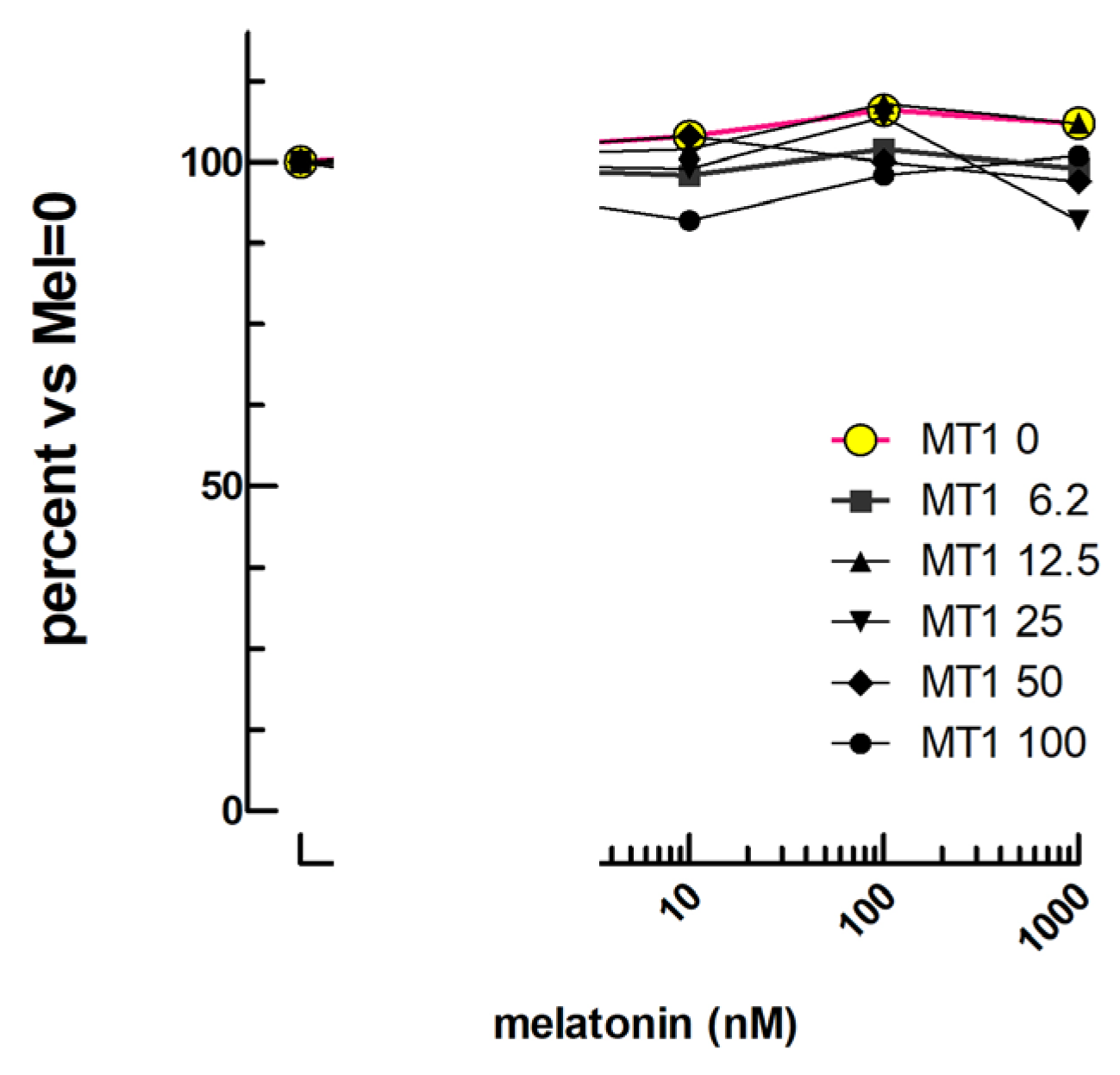
2.7. Melatonin Bioassay
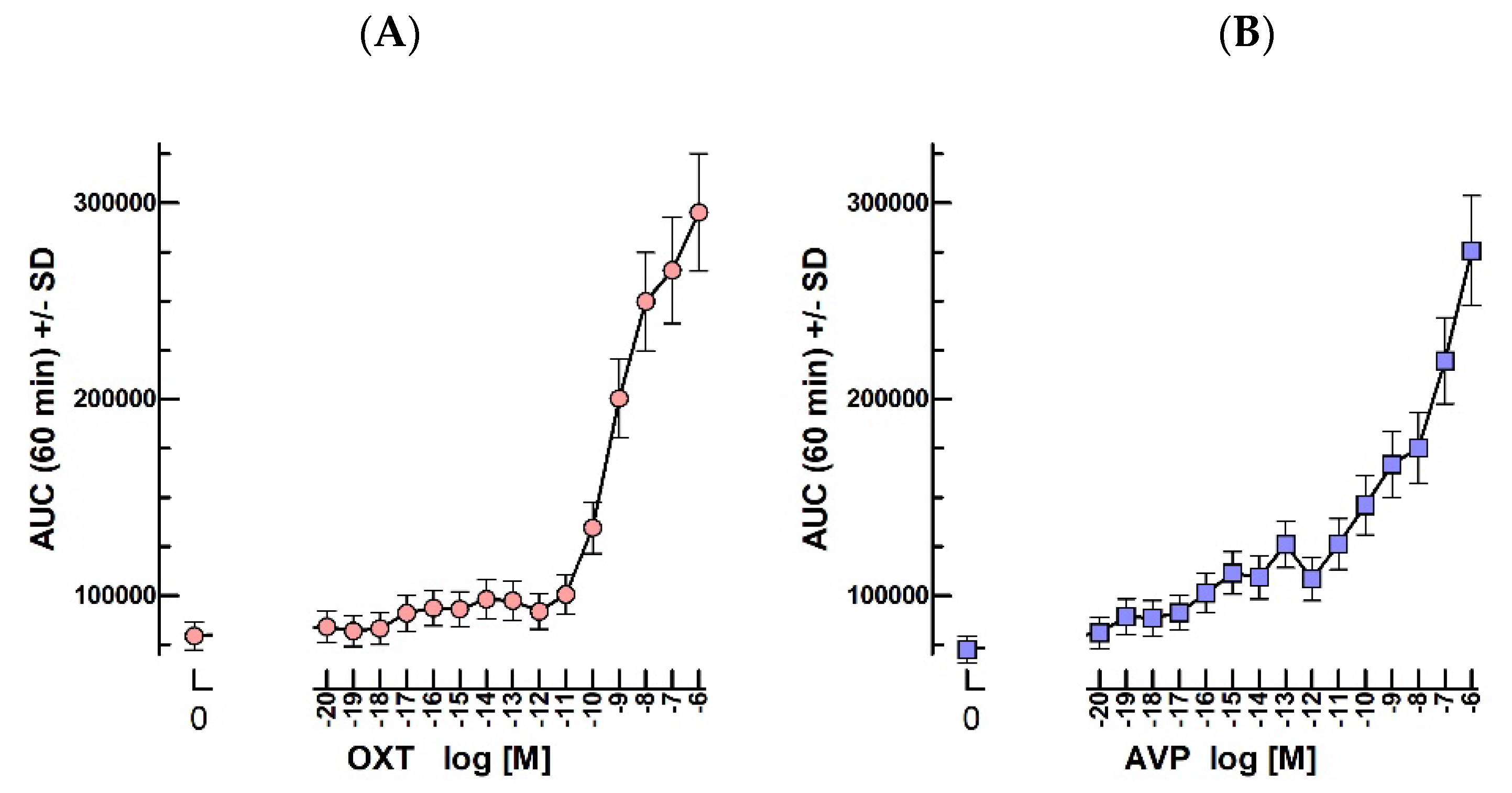
2.8. Oxytocin and Vasopressin Bioassays
3. Discussion
3.1. LH/CG
3.2. FSH
3.3. TSH
3.4. PTH
3.5. Kisspeptins
3.6. β2AR Ligands
3.7. Melatonin
3.8. OT
4. Materials and Methods
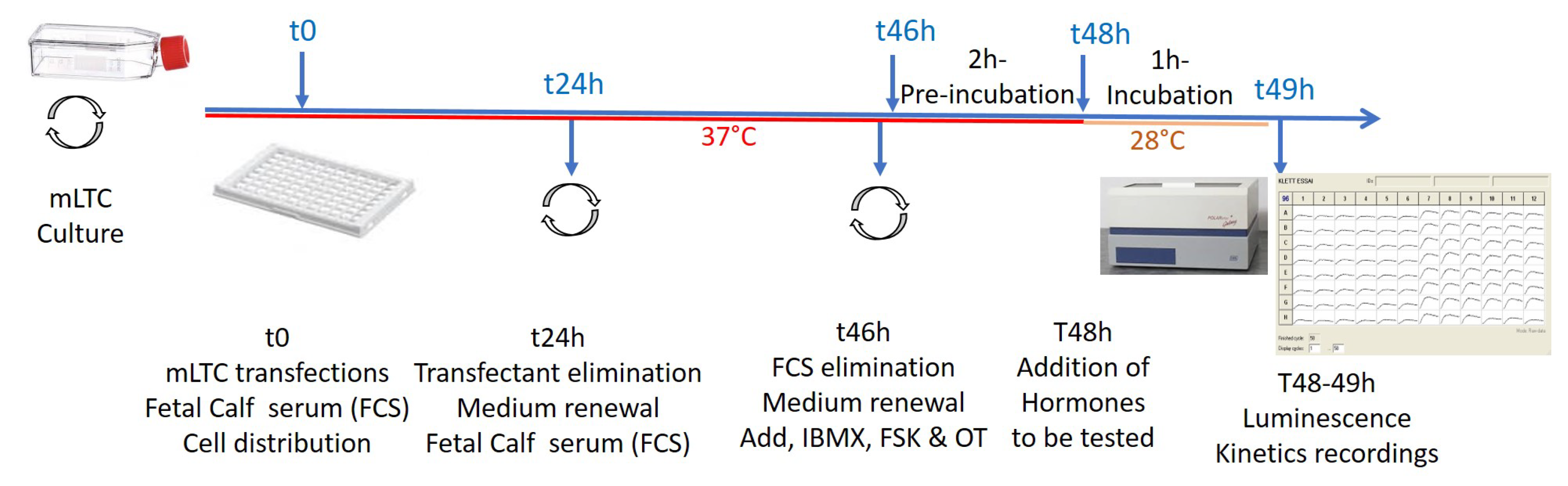
4.1. Protocols
4.2. Hormones
4.3. Receptors’ Expression Vectors
4.3.1. Human FSH Receptor
4.3.2. Ovine TSH Receptor
4.3.3. Human PTH Receptor
4.3.4. Human KISS Receptor
4.3.5. Human β2-Adrenergic Receptor
4.4. Biological Samples
4.5. Area Under Curve (AUC) Calculations and Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klett, D.; Combarnous, Y. Highly sensitive in vitro bioassay for luteinizing hormone and chorionic gonadotropin allowing their measurement in plasma. Reprod. Fertil. 2021, 2, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Rebois, R.V. Establishment of gonadotropin-responsive murine leydig tumor cell line. J. Cell Biol. 1982, 94, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.; Dighe, R.R. Critical involvement of the hinge region of the follicle-stimulating hormone receptor in the activation of the receptor. J. Biol. Chem. 2009, 284, 2636–2647. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Choi, S.H.; Hong, N.; Rhee, Y.; Kim, J.K.; Lee, C.R.; Kang, S.W.; Lee, J.; Jeong, J.J.; Nam, K.H.; et al. Comparisons Between Normocalcemic Primary Hyperparathyroidism and Typical Primary Hyperparathyroidism. J. Korean Med. Sci. 2022, 37, e99. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.; Houillier, P.; Gauci, C.; Haymann, J.P.; Flamant, M.; Thervet, E.; Boffa, J.J.; Vrtovsnik, F.; Froissart, M.; Stengel, B.; et al. Relation between circulating levels of 25(OH) vitamin D and parathyroid hormone in chronic kidney disease: Quest for a threshold. J. Clin. Endocrinol. Metab. 2013, 98, 2922–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzoni, A.; Zhang, X.; Naim, N.; Altschuler, D.L. Soluble cyclase-mediated nuclear cAMP synthesis is sufficient for cell proliferation. Proc. Natl. Acad. Sci. USA 2023, 120, e2208749120. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, M.; Guan, R.; Segaloff, D.L. Heterodimerization between the lutropin and follitropin receptors is associated with an attenuation of hormone-dependent signaling. Endocrinology 2013, 154, 3925–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazurkiewicz, J.E.; Herrick-Davis, K.; Barroso, M.; Ulloa-Aguirre, A.; Lindau-Shepard, B.; Thomas, R.M.; Dias, J.A. Single-molecule analyses of fully functional fluorescent protein-tagged follitropin receptor reveal homodimerization and specific heterodimerization with lutropin receptor. Biol. Reprod. 2015, 92, 100. [Google Scholar] [CrossRef] [Green Version]
- Kleinau, G.; Worth, C.L.; Kreuchwig, A.; Biebermann, H.; Marcinkowski, P.; Scheerer, P.; Krause, G. Structural-Functional Features of the Thyrotropin Receptor: A Class A G-Protein-Coupled Receptor at Work. Front. Endocrinol. 2017, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Zoenen, M.; Urizar, E.; Swillens, S.; Vassart, G.; Costagliola, S. Evidence for activity-regulated hormone-binding cooperativity across glycoprotein hormone receptor homomers. Nat. Commun. 2012, 3, 1007. [Google Scholar] [CrossRef] [Green Version]
- Vieira, J.G.H. PTH assays: Understanding what we have and forecasting what we will have. J. Osteoporos. 2012, 2012, 523246. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, J.; Pan, P.; Nguyen, T.; Cwik, M.; Morgan, L. Novel sequential immunocapture microflow LC/MS/MS approach to measuring PTH-Fc protein in human serum. Bioanalysis 2022, 14, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Valsecchi, F.; Konrad, C.; Manfredi, G. Role of soluble adenylyl cyclase in mitochondria. Biochim. Biophys. Acta 2014, 1842, 2555–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvin, T.N.; Kamenetsky, M.; Zarifyan, A.; Buck, J.; Levin, L.R. Kinetic properties of “soluble” adenylyl cyclase. Synergism between calcium and bicarbonate. J. Biol. Chem. 2003, 278, 15922–15926. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.M.D.; Filliatreau, L.; Klett, D.; Hai, N.V.; Duong, N.T.; Combarnous, Y. Effect of Soluble Adenylyl Cyclase (ADCY10) Inhibitors on the LH-Stimulated cAMP Synthesis in Mltc−1 Leydig Cell Line. Int. J. Mol. Sci. 2021, 22, 4641. [Google Scholar] [CrossRef]
- Parker, T.; Wang, K.-W.; Manning, D.; Dart, C. Soluble adenylyl cyclase links Ca2+ entry to Ca2+/cAMP-response element binding protein (CREB) activation in vascular smooth muscle. Sci. Rep. 2019, 9, 7317. [Google Scholar] [CrossRef] [Green Version]
- Sposini, S.; Jean-Alphonse, F.G.; Ayoub, M.A.; Oqua, A.; West, C.; Lavery, S.; Brosens, J.J.; Reiter, E.; Hanyaloglu, A.C. Integration of GPCR Signaling and Sorting from Very Early Endosomes via Opposing APPL1 Mechanisms. Cell Rep. 2017, 21, 2855–2867. [Google Scholar] [CrossRef] [Green Version]
- Xiao, R.-p. β-Adrenergic Signaling in the Heart: Dual Coupling of the β2-Adrenergic Receptor to Gs and Gi Proteins. Sci. STKE 2001, 2001, re15. [Google Scholar]
- Heng, J.; Hu, Y.; Perez-Hernandez, G.; Inoue, A.; Zhao, J.; Ma, X.; Sun, X.; Kawakami, K.; Ikuta, T.; Ding, J.; et al. Function and dynamics of the intrinsically disordered carboxyl terminus of beta2 adrenergic receptor. Nat. Commun. 2023, 14, 2005. [Google Scholar] [CrossRef]
- Logez, C.; Berger, S.; Legros, C.; Banères, J.-L.; Cohen, W.; Delagrange, P.; Nosjean, O.; Boutin, J.A.; Ferry, G.; Simonin, F.; et al. Recombinant Human Melatonin Receptor MT1 Isolated in Mixed Detergents Shows Pharmacology Similar to That in Mammalian Cell Membranes. PLoS ONE 2014, 9, e100616. [Google Scholar] [CrossRef]
- Lin, Y.T.; Huang, C.C.; Hsu, K.S. Oxytocin promotes long-term potentiation by enhancing epidermal growth factor receptor-mediated local translation of protein kinase Mzeta. J. Neurosci. 2012, 32, 15476–15488. [Google Scholar] [CrossRef]
- Devost, D.; Carrier, M.E.; Zingg, H.H. Oxytocin-induced activation of eukaryotic elongation factor 2 in myometrial cells is mediated by protein kinase C. Endocrinology 2008, 149, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinetz, S.; Meinung, C.P.; Jurek, B.; von Schack, D.; van den Burg, E.H.; Slattery, D.A.; Neumann, I.D. De Novo Protein Synthesis Mediated by the Eukaryotic Elongation Factor 2 Is Required for the Anxiolytic Effect of Oxytocin. Biol. Psychiatry 2019, 85, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Klett, D.; Meslin, P.; Relav, L.; Nguyen, T.M.; Mariot, J.; Jegot, G.; Cahoreau, C.; Combarnous, Y. Low reversibility of intracellular cAMP accumulation in mouse Leydig tumor cells (MLTC-1) stimulated by human Luteinizing Hormone (hLH) and Chorionic Gonadotropin (hCG). Mol. Cell. Endocrinol. 2016, 434, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Hennen, G.; Prusik, Z.; Maghuin-Rogister, G. Porcine luteinizing hormone and its subunits. Isolation and characterization. Eur. J. Biochem. 1971, 18, 376–383. [Google Scholar] [CrossRef]
- Tranchant, T.; Durand, G.; Gauthier, C.; Crepieux, P.; Ulloa-Aguirre, A.; Royere, D.; Reiter, E. Preferential beta-arrestin signalling at low receptor density revealed by functional characterization of the human FSH receptor A189 V mutation. Mol. Cell. Endocrinol. 2011, 331, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Dupre, S.M. Encoding and decoding photoperiod in the mammalian pars tuberalis. Neuroendocrinology 2011, 94, 101–112. [Google Scholar] [CrossRef]
- Dupre, S.M.; Dardente, H.; Birnie, M.J.; Loudon, A.S.; Lincoln, G.A.; Hazlerigg, D.G. Evidence for RGS4 modulation of melatonin and thyrotrophin signalling pathways in the pars tuberalis. J. Neuroendocrinol. 2011, 23, 725–732. [Google Scholar] [CrossRef]
- Shimada, M.; Chen, X.; Cvrk, T.; Hilfiker, H.; Parfenova, M.; Segre, G.V. Purification and Characterization of a Receptor for Human Parathyroid Hormone and Parathyroid Hormone-related Peptide. J. Biol. Chem. 2002, 277, 31774–31780. [Google Scholar] [CrossRef] [Green Version]
- Beltramo, M.; Robert, V.; Galibert, M.; Madinier, J.B.; Marceau, P.; Dardente, H.; Decourt, C.; De Roux, N.; Lomet, D.; Delmas, A.F.; et al. Rational design of triazololipopeptides analogs of kisspeptin inducing a long-lasting increase of gonadotropins. J. Med. Chem. 2015, 58, 3459–3470. [Google Scholar] [CrossRef]
- Decourt, C.; Robert, V.; Anger, K.; Galibert, M.; Madinier, J.B.; Liu, X.; Dardente, H.; Lomet, D.; Delmas, A.F.; Caraty, A.; et al. A synthetic kisspeptin analog that triggers ovulation and advances puberty. Sci. Rep. 2016, 6, 26908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomet, D.; Cognie, J.; Chesneau, D.; Dubois, E.; Hazlerigg, D.; Dardente, H. The impact of thyroid hormone in seasonal breeding has a restricted transcriptional signature. Cell. Mol. Life Sci. 2018, 75, 905–919. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klett, D.; Pellissier, L.; Lomet, D.; Derouin-Tochon, F.; Robert, V.; Nguyen, T.M.D.; Duittoz, A.; Reiter, E.; Locatelli, Y.; Dupont, J.; et al. Highly-Sensitive In Vitro Bioassays for FSH, TSH, PTH, Kp, and OT in Addition to LH in Mouse Leydig Tumor Cell. Int. J. Mol. Sci. 2023, 24, 12047. https://doi.org/10.3390/ijms241512047
Klett D, Pellissier L, Lomet D, Derouin-Tochon F, Robert V, Nguyen TMD, Duittoz A, Reiter E, Locatelli Y, Dupont J, et al. Highly-Sensitive In Vitro Bioassays for FSH, TSH, PTH, Kp, and OT in Addition to LH in Mouse Leydig Tumor Cell. International Journal of Molecular Sciences. 2023; 24(15):12047. https://doi.org/10.3390/ijms241512047
Chicago/Turabian StyleKlett, Danièle, Lucie Pellissier, Didier Lomet, Flavie Derouin-Tochon, Vincent Robert, Thi Mong Diep Nguyen, Anne Duittoz, Eric Reiter, Yann Locatelli, Joëlle Dupont, and et al. 2023. "Highly-Sensitive In Vitro Bioassays for FSH, TSH, PTH, Kp, and OT in Addition to LH in Mouse Leydig Tumor Cell" International Journal of Molecular Sciences 24, no. 15: 12047. https://doi.org/10.3390/ijms241512047