
Plants and Mushrooms as Possible New Sources of H2S Releasing Sulfur Compounds

1
Department of Pharmacy, University of Pisa, Via Bonanno, 56120 Pisa, Italy
2
Golden Wave srl, Via Don G. Lago, 35013 Padova, Italy
3
Interdepartmental Center of Nutrafood, University of Pisa, Via Del Borghetto 80, 56124 Pisa, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(15), 11886; https://doi.org/10.3390/ijms241511886
Submission received: 30 June 2023
/
Revised: 20 July 2023
/
Accepted: 22 July 2023
/
Published: 25 July 2023
(This article belongs to the Special Issue Hydrogen Sulfide (H2S)-Donor Molecules: Chemical, Biological, and Therapeutical Tools)
Abstract
:Hydrogen sulfide (H2S), known for many decades exclusively for its toxicity and the smell of rotten eggs, has been re-discovered for its pleiotropic effects at the cardiovascular and non-cardiovascular level. Therefore, great attention is being paid to the discovery of molecules able to release H2S in a smart manner, i.e., slowly and for a long time, thus ensuring the maintenance of its physiological levels and preventing “H2S-poor” diseases. Despite the development of numerous synthetically derived molecules, the observation that plants containing sulfur compounds share the same pharmacological properties as H2S led to the characterization of naturally derived compounds as H2S donors. In this regard, polysulfuric compounds occurring in plants belonging to the Alliaceae family were the first characterized as H2S donors, followed by isothiocyanates derived from vegetables belonging to the Brassicaceae family, and this led us to consider these plants as nutraceutical tools and their daily consumption has been demonstrated to prevent the onset of several diseases. Interestingly, sulfur compounds are also contained in many fungi. In this review, we speculate about the possibility that they may be novel sources of H2S-donors, furnishing new data on the release of H2S from several selected extracts from fungi.
1. Introduction
Hydrogen sulfide (H2S) has been known for many decades for its toxicity and the smell of rotten eggs. Hydrogen sulfide poisoning usually occurs by inhalation; local irritant effects result in direct irritation to the eyes, causing conjunctival injection initially and corneal injury eventually. When inhaled, it leads to pulmonary injury presenting as hemorrhagic pulmonary edema. Hydrogen sulfide inhibits mitochondrial cytochrome oxidase by making a complex bond to the ferric moiety of the protein, therefore arresting aerobic metabolism. Once it enters the bloodstream and passes the blood–brain barrier, neurotoxic effects can be seen, namely dizziness, seizure, coma, and ultimately death. In brief, hydrogen sulfide is a known pulmonary irritant and asphyxiant that primarily causes respiratory and neurological clinical manifestations when inhaled. High concentrations (more than 700 ppm) have the potential to cause sudden death [1,2].
However, in the last twenty years, it has been discovered as the third endogenously produced gasotransmitter, in addition to nitric oxide and carbon monoxide [3].
The importance of characterizing and discovering new sources to be used as supplements of H2S or developing new chemical entities endowed with H2S-donor properties is due to the pleiotropic role of this gaseous molecule. Indeed, H2S is involved in the regulation of numerous pathophysiological functions: it intervenes in the anti-inflammatory process [4], in oxidative stress [5], neuro-modulation [6], vaso-regulation [7], protection from ischaemia/reperfusion damage after myocardial infarction [8] and insulin resistance [9]. Furthermore, recent studies have shown that an abnormal metabolism and altered pathways of H2S are related to the development of various cardiovascular and non-cardiovascular diseases (Figure 1) [10,11,12].
Numerous synthetically derived molecules have been developed, and the discovery of new interesting scaffolds able to release H2S in biological environments, including thioureas [13], isothiocyanates [14,15,16], thiazolydindione [17], arylthioamide [18], and imminothioether derivatives [19], have highlighted the concrete usefulness of using these molecules for the treatment of several pathological conditions. Moreover, H2S-donor moieties have also been used for developing hybrid drugs upon merging the chemical portion able to release H2S to drugs already used in the clinic [20,21,22,23,24,25,26,27]. This strategy allows us to exploit the beneficial effect of H2S and the well characterized pharmacological effect of the native drug.
In addition to synthetic molecules, the observation that plants containing sulfur compounds share the same pharmacological properties with H2S led to characterize naturally derived compounds as H2S donors. Polysulfuric compounds occurring in plants belonging to the Alliaceae family were the first to be characterized as H2S donors, followed by isothiocyanates derived from vegetables belonging to the Brassicaceae family [28,29]. This important discovery paved the way to consider these plants as nutraceutical tools and their daily consumption has been demonstrated to prevent the onset of several diseases [30]. Sulfur compounds are also contained in many fungi. However, to date no data about their possible H2S releasing properties have been reported.
This review aims to describe the pharmacological effects of sulfur compounds present in plants belonging to the Alliaceae and Brassicaceae families and provide new data on the H2S-releasing properties of fungi extracts.
2. Hydrogen Sulfide Endogenous Production
In mammals, the endogenous production of this gasotransmitter is ensured by several enzymes, that are differently expressed in tissues, suggesting its crucial role in the control of homeostasis in numerous systems. Recently, H2S has been reported to modulate target proteins through S-sulfhydration reactions, which are a post-translational modification of the hydrosulfuryl groups of cysteine residues responsible for the activation/inhibition of specific proteins [31,32].
H2S is mainly bio-synthetized using two cytosolic enzymes: cystathionine γ-lyase (CSE) and cystathionine β-synthase (CBS), and one mitochondrial enzyme: 3-mercaptopyruvate sulfur-transferase (MST), using a shared substrate, the amino acid L-cysteine [33]. Interestingly, CBS is mainly but not exclusively expressed in the central nervous system (CNS), while CSE is recognized as the enzyme responsible for the endogenous production of H2S in the cardiovascular system. More recently, additional/alternative biosynthetic pathways have been described; indeed, H2S can be generated by cysteinyl-tRNA-synthetase enzyme and by selenium-binding protein 1, although their contribution is still unclear [34]. The wide distribution of biosynthetic pathways for the endogenous production of H2S accounts for its ubiquitous and pleiotropic behavior.
With regards to metabolic processes, H2S can follow different pathways: firstly, H2S is a reducing molecule, and it is involved in reducing reactive oxygen species (ROS). Oxidation of H2S represents the main catabolic function at the mitochondrial level, where H2S is quickly metabolized to sulfite and sulfate species due to the involvement of quinone oxidoreductase, rhodanese, and sulfur dioxygenase [32].
3. Overview of Hydrogen Sulfide and Its Therapeutic Potential in Oxidative Stress-Based Pathologies
3.1. Oxidative Stress
It is well-known that oxygen is an indispensable molecule, but because of its structure, it can lead to the generation of highly unstable intermediates known as ROS [35]. When the ROS level exceeds the neutralizing capability of endogenous anti-oxidant systems, the tissue is exposed to oxidative stress. The accumulation of ROS is implicated in mitochondrial dysfunction of the cells due to oxidation of mitochondrial lipids leading to a decrease in the membrane potential and finally apoptosis [36]. In this regard, the nuclear transcription factor erythroid 2 (Nrf-2) is the main regulator of the anti-oxidant defense mechanisms within the cell. Nrf-2 modulates the gene expression of anti-oxidant and cytoprotective enzymes and is inhibited by Kelch-like ECH-associated protein 1 (Keap1). For this reason, Nrf-2 represents a possible promising pharmacological target to decrease cellular oxidative stress [37].
Although many studies have demonstrated direct H2S scavenging action, the physiological concentration of H2S is in the low nanomolar range, significantly lower than that of other endogenous anti-oxidant molecules [38]. Consequently, the anti-oxidant effects described for H2S comprise the up-regulation of anti-oxidant defense systems [39].
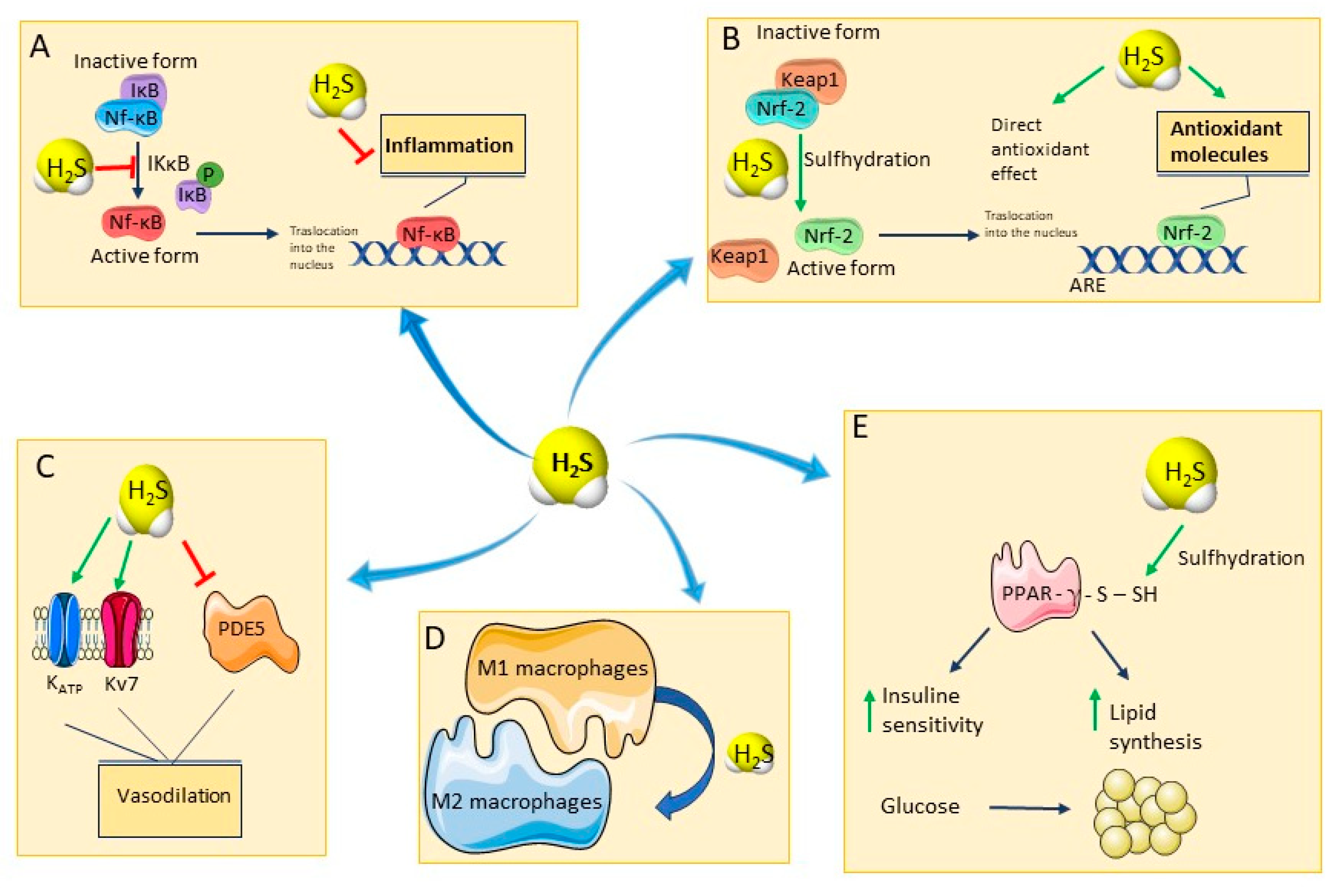
H2S is known for its ability to protect from oxidative stress by inducing an S-sulfhydration reaction of the Keap1 protein, in cysteine-151 residue (Figure 2B). This reaction induces a conformational change in Keap1 and its consequent dissociation from Nrf-2, which translocates into the nucleus. Here, Nrf-2 binds to promoters containing the anti-oxidant response element (ARE) gene sequence, favoring the expression of ARE-dependent genes encoding for major anti-oxidant enzymes, such as glutathione reductase, Catalase (CAT), Superoxide dismutase (SOD), and for non-enzymatic anti-oxidants, such as glutathione (GSH) and Thioredoxin 1 (Trx-1) [40].
3.2. Inflammation
H2S promotes a significant anti-inflammatory effect mainly by inhibiting the pro-inflammatory transcription factor Nuclear factor kappaB (NF-kB) and preventing mast cell degranulation [41,42]. In a model of induced inflammatory response in pulmonary artery endothelial cells, H2S inhibits the inhibitor of κβ kinase (IKκβ) enzyme activity through S-sulfhydration of the Cysteine 179 residue, thereby preventing NF-kB translocation into the nucleus [43,44] (Figure 2A).
H2S also plays a regulatory role in macrophages, which are traditionally classified into two subgroups: M1, which are classically activated macrophages, are differentiated by the action of Th1 cytokines, and M2, which are alternatively activated macrophages, are differentiated by the action of Th2 cytokines, such as IL-4 or IL-13 [45].
M1s produce pro-inflammatory cytokines, and are important for antibacterial defense, while M2 macrophages mainly produce anti-inflammatory mediators and are involved in the anti-inflammatory and repair response [46]. H2S promotes the polarization of macrophages toward M2, thus involving an anti-inflammatory effect (Figure 2D) [47].
3.3. Immune System
H2S is also involved in the regulation of the immune system, playing a pivotal role in the regulation of neutrophils. Interestingly, neutrophils express all three major enzymes responsible for H2S production, although recent studies have failed to measure detectable activity of CBS in the homogenate of human neutrophils [48]. Even if the mechanisms are complex, research data suggest that H2S inhibits the adhesion of neutrophils [48]. The molecular mechanisms by which H2S reduces leukocyte migration and adhesion in various experiments are different: activation of different classes of potassium channels [49], induction of heme oxygenase-1 [50], and downregulation of CD11b expression [51]. However, in the presence of a strong pro-inflammatory stimulus, H2S stimulates neutrophil tissue adhesion and infiltration [52] through the upregulation of adhesion receptors, such as intercellular adhesion molecule 1 (ICAM-1) and P-selectin [53]. It is not contradictory with data reported earlier: under a septic condition, an increase in leukocyte infiltration is desirable since the organism is compromised.
Studies performed under microbial infection conditions showed that inhibition of H2S biosynthesis increased the capacity for bacterial migration (and thus reduced the rate of killing) as fewer mycobacteria were internalized in the acidic vesicles of the macrophages [54]. This suggests that the biosynthesis of H2S by the host organism is necessary for proper macrophage activity. In fact, when neutrophils were co-cultured with E. coli HB101 in the presence of H2S for 24 h, the elimination of bacteria was more efficient than in the absence of H2S [32].
3.4. Cardiovascular and Metabolic Systems
At the vascular level, H2S induces most of its effects by S-sulfhydration of proteins, such as ion channels, enzymes, and receptors, which thereby undergo a conformational change responsible for their activation or inhibition [55,56].
One of the first reported mechanisms of action underlying H2S-induced vasodilation consists of the activation of ATP-sensitive potassium (KATP) channels with the subsequent hyperpolarization of vascular smooth muscle cells [57]. Subsequently, it was shown that another important mechanism of action responsible for H2S-induced vasodilation is the activation of voltage-dependent potassium channels belonging to the Kv7 family [58]. H2S is also able to inhibit 5-phosphodiesterase (5-PDE) enzyme and has also been described as an endothelium-derived hyperpolarizing factor (EDHF) (Figure 2C) [59]. Several studies shed light on the cardioprotection played by this gasotransmitter against several kinds of damage, including the ischemia-reperfusion, diabetic cardiomyopathy, myocardial infarction, and drug-induced cardiotoxicity [60,61]. Although, multiple targets seem to be implicated, among which the regulation of nitric oxide (NO) levels [62], the up-regulation of 5’ adenosine monophosphate-activated protein kinase (AMPK) and Nrf-2 [63], the stimulation of mitochondrial ATP-sensitive potassium (mitoKATP) channels has been probably the most investigated target [64].
Moreover, H2S is considered a master regulator of systemic metabolism, indeed through the S-sulfhydration of the insulin receptor substrate 1 (IRS-1), it may maintain insulin sensitivity and, upon direct S-sulfhydration at the cysteine139 site, it may increase peroxisome proliferator-activated receptor γ (PPARγ) activity, thereby changing glucose into triglyceride storage in adipocytes (Figure 2E) [65,66].
Based on the numerous beneficial effects that can be mediated by this gasotransmitter, great attention is being paid to the discovery of molecules able to release H2S in a smart manner, i.e., slowly and for a long time, thus ensuring the maintenance of its physiological levels and preventing “H2S-poor” diseases.
4. Naturally Derived H2S-Donors
4.1. Polysulfides from the Alliaceae Family
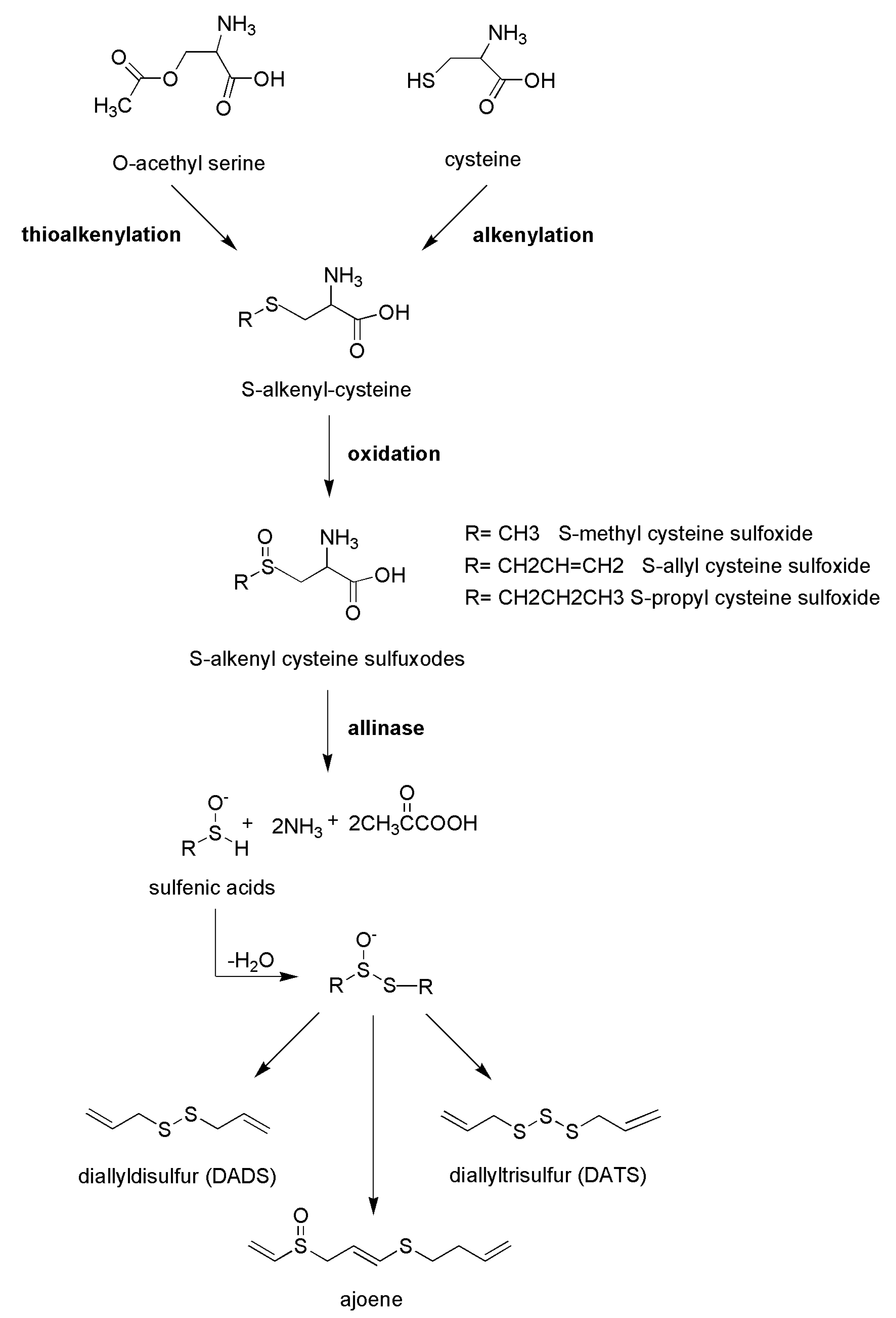
Traditional medicine suggests daily consumption of garlic (Allium sativum L.) to maintain the physiological values of systolic and diastolic pressure, and several studies support the effects of garlic extracts and its polysulfides as cardiovascular-protective compounds [67,68,69]. The most abundant polysulfides present in garlic and in other edible species from the Alliaceae family, such as chives, onion, and shallot, are allyl sulfide derivatives, including diallyl disulfide and diallyl trisulfide, produced by the cleavage of odorless molecules, including S-alk(en)yl cysteine sulphoxide precursors due to the activity of alliinase enzyme. These secondary metabolites are S-methyl cysteine sulphoxide, S-allyl cysteine sulphoxide (known as alliin), S-propyl cysteine sulphoxide, and S-transprop-1-enyl cysteine sulphoxide [70]. Consequently, after the disruption of cells (upon cutting or cooking), alliinase enzyme, present in vacuoles, it is released and the reaction responsible for the formation of typical polysulfides occurs (Figure 3). The enrichment in organosulfur compounds is closely correlated with the aging of the vegetable; interestingly, it has been reported that reactive organosulfur compounds, such as allicin, are converted to their stable isoforms, such as S-allyl cysteine [71].
H2S release is the main mechanism accounting for the vascular effects of polysulfides. The vasorelaxant effects following the administration of garlic extract in rat aortic rings were associated with a release of H2S in the isolated organ bath [72]. Finally, the exact mechanism through which H2S is released from polysulfides has also been described. They demonstrated that the functional group present in organic thiols, such as L-cysteine or glutathione, can promote a nucleophilic attack on the carbon atom in α-position to the allyl group and then trigger the rupture of the molecule, ultimately favoring the release of H2S (Figure 4) [72].
In addition, other beneficial effects of garlic are related to the release of H2S, including metabolic ones. Garlic shows potential beneficial effects in T2D [73,74,75]; this vegetable, and its derivatives are endowed with antihyperglycemic effects in genetic animal models of diabetes and in humans, thus preventing cardiovascular complications [76]. Consistently, garlic improves insulin sensitivity and the associated metabolic syndrome in animal models, and its derivatives reduce both insulin resistance and blood glucose in streptozotocin- and alloxan-induced diabetes [77].
Based on the putative targets identified for H2S, it has been hypothesized that the contribution of garlic in the maintenance of glucose homeostasis or lipid profile is due—at least in part—to the release of H2S, and several putative targets have been identified. Interestingly, polysulfides upregulated and S-sulfhydrated PPARγ and sirtuin 3 (SIRT-3) in cardiomyocytes and such a mechanism was relevant for assuring a prevention of diabetic cardiomyopathy in murine model [78].
4.2. Isothiocyanates from the Brassicaceae Family
The discovery about the H2S releasing properties of the Alliaceae family paved the way for a great breakthrough in the research on plant species containing sulfur compounds, as a nutraceutical approach to be employed at early stages of several diseases. In this scenario, the Brassicaceae family is rich in glucosinolates and sulfur compounds that are metabolized into isothiocyanates because of the activation of myrosinase when these plants are cut or shredded [79]. Since there is an overlap between physio/biological effects evoked by isothiocyanates and those exerted by H2S (as reported above), the H2S-releasing properties of some natural isothiocyanates were evaluated [29]. Most of the compounds generated appreciable release of H2S following incubation with cysteine. This behavior is due to the nucleophilic reaction of the thiol group of L-cysteine with the isothiocyanate moiety. This characteristic makes these molecules “smart H2S donors” since they can donate the gaseous transmitter only in a biological environment. Recently, the chemical mechanism explaining the generation of H2S from isothiocyanates has been clarified: the reaction between L-cysteine and isothiocyanates resulted in the generation of H2S due to the generation of dithiocarbamic derivatives between L-cysteine and the isothiocyanate moiety. Intramolecular cyclization leads to the generation of dihydrothiazole derivatives and the formation of H2S (Figure 5) [80].
Currently, isothiocyanates are attracting increasing interest mainly for their pharmacological effects at the cardiovascular level. In this regard, a recent study evaluated the vascular effects of erucin related to its H2S-donating properties. In this study, the authors demonstrated that erucin releases H2S in human aortic smooth muscle cells (HASMCs) in a concentration-dependent manner. Furthermore, a significant antihypertensive effect was reported in spontaneously hypertensive rats after erucin intraperitoneal administration, recording systolic pressure values similar to normotensive rats [81].
Moreover, erucin also promotes protective effects in oxidative stress and LPS-induced damage in endothelial and vascular cells, and at cardiac level [82,83,84].
Finally, an emerging role of H2S is the regulation of glucose metabolism: several studies have been performed on Brassicaceae as a possible nutraceutical approach in an early stage of diabetes or combination with standard therapy. Consumption of Alliaceae and Brassicaceae can significantly improve blood glucose control in patients with diabetes, concluding that edible plants that contain sulfur compounds could be useful for treating type 2 diabetes (T2D). Furthermore, supplementation with edible plants containing sulfur compounds could significantly enhance the effect of standard therapy on glucose control [85].
Male balb/c mice fed for 10 weeks with a high-fat diet and supplemented with an extract obtained from the seeds of Eruca sativa Mill., rich in glucoerucin, showed a reduction in the body weight gain and improvement of glucose homeostasis. Interestingly, a decrease in white adipose tissue and the size of adipocytes was also observed. In addition, the extract improved adipocyte metabolism by enhancing the activity of citrate synthase and reducing triglyceride levels in mice fed a high-fat diet. Worthy of note, a diet rich in Brassicaceae has no adverse effects: the most common adverse events are mild gastrointestinal issues (heartburn, flatulence, stomach discomfort, increased defecation) and hot flashes [85].
Another isothiocyanate capable of releasing H2S is moringin. This molecule derives from Moringa oleifera Lam., a plant belonging to the Moringaceae family that is widely used in the traditional medicine for the treatment of stomach pain, ulcers, vision defects, joint pain and as a digestive [86]. Specifically, the authors analyzed changes in the content of glucosinolates and isothiocyanates in different moringa tissues and measured the H2S-releasing properties with the lead acetate assay, a method that takes advantage of the high affinity of divalent lead with H2S to form a black precipitate of lead sulfide (PbS). The authors showed that moringa seeds and leaves had a significant amount of total glucosinolates that can be converted to isothiocyanates (mainly benzyl isothiocyanate) by the action of myrosinase [82].
4.3. Organosulfur Compounds Present in Mushrooms
Edible mushrooms have been widely used in cooking, but in recent years, their consumption for their medical benefits has increased. In this regard, similar to Alliaceae and Brassicaceae vegetables, the main areas of use are cardio-metabolic disorders and the regulation of the immune system. A recent systematic review highlights that the consumption of mushrooms—as part of a healthy dietary pattern—is associated with a reduced risk of mortality for cardiovascular diseases, suggesting benefits on cardiovascular and metabolic systems. It has been proposed that these benefits are correlated to hypolipidemic and hypoglycemic effects and anti-oxidants and anti-inflammatory activities [87,88,89,90]. More recently, a putative role in slowing down aging has also been considered and associated with the ability to influence the expression of hallmarks of senescence, including Nrf-2 and FOXOs [91]. Moreover, some authors supposed the role of supplementation with mushrooms to support the treatment of the early stages of neurodegenerative diseases or anti-cancer because of the high anti-oxidant and anti-inflammatory capacities [92,93,94]. Finally, with regard to the regulation of immune system, mushrooms are reported to stimulate cell surface receptor activity, enhancing the activity of natural killer cells, neutrophils and macrophages, responsible for anti-viral and anti-tumor responses [94].
Undoubtedly in these properties, an important contribution is due to the presence of polysaccharides (including β-glucans), enzymes, nucleic acids and other bioactive compounds, including ergosterol and monacolin k [92,93,94]; indeed, usually, the phyto-complex is considered to be responsible for a wide spectrum of beneficial effects; nevertheless today investigations on the mechanisms of action are poor, and a deeper exploration seems to be necessary.
Exploring further bioactive compounds endowed with putative healthy properties, a peculiar characteristic of these vegetables is their distinctive flavor, and the scent is critical in the determination of their quality and popularity. Indeed, a wide variety of volatile organic compounds has been described, and their content, along with non-volatile organic compounds, renders each edible mushroom unique, but at the same time, it can influence medicinal properties. Interestingly, among the volatile organic compounds, organic sulfur compounds have been described [95]. Their recognized precursors are L-cysteine and methionine, which can be converted into thioheterocycles, thioethers, thiols and thiophenes in enzymatic and non-enzymatic ways [96]. A peculiar organic sulfur compound typical of mushrooms and not present in other kinds of vegetable and nonvegetable cells is ergothioneine, an amino acid bearing a cyclized thioureidic group that participates in the health-promoting activity of mushrooms [97].
One of the most studied mushrooms is Lentinula edodes (well-known as shiitake). It is rich in sulfurs; indeed, the unique aroma of the mushroom is due to the presence of lenthionine (1,2,3,5,6-pentathiepane), a cyclic sulfur compound [98,99]. In addition to its sensorial properties, lenthionine is endowed with biological effects, including antibiotic and anti-aggregation properties [100]; therefore, great attention has been paid to it.
From a biosynthetic perspective, lenthionine derives from lentinic acid in a two-step enzymatic reaction. In particular, lentinic acid is activated upon the elimination of its γ-glutamyl moiety due to the action of γ-glutamyl transpeptidase, producing an L-cysteine sulfoxide derivative, which then undergoes α,β-elimination, catalyzed by cysteine sulfoxide lyase (better known as alliinase, present in Alliaceae vegetables), resulting in a highly reactive sulfenic acid intermediate. The sulfenic acid is then rapidly condensed to form thiosulfinate, which is often further transformed into other sulfur compounds, including lenthionine [101]. Drying conditions, probably due to the Maillard reaction, may deeply influence the organic volatile composition, particularly the content of organic sulfur compounds. The concentration of organic sulfur compounds in shiitake increased in the early stages of drying (about 0.5–1.5 h), and the most representative compounds were dimethyl trisulfide, thioanisole, and lenthionine [102]. In the middle stages of drying the organic sulfur compounds increased with the esters; while in the final stages (4–12 h) their concentration decreased. Interestingly, the sulfur perception as a negative impression of food was hypothesized; nevertheless, it could be interesting from a health point of view [103].
Currently, to the best of our knowledge, no organic sulfur compounds described in shiitake have been described as H2S donors; on the other hand, poor information is currently available on the content of organic sulfur compounds in other edible mushrooms. Interestingly, our preliminary studies suggest it is a potentially new source of sulfur compounds endowed with H2S-donor properties.
5. Analysis of H2S Release from Mushroom Extracts
Using an amperometric approach [13,104], we screened a few extracts of mushrooms (listed in Table 1) to explore their ability to release H2S.
Surprisingly, the extracts of Lentinula edodes Berk., Ganoderma lucidum Curtis and Polyporus umbellatus, tested using an amperometric method at a concentration of 1 mg/mL in the presence and absence of L-cysteine (4 mM), showed a cysteine-dependent release of H2S. In detail, in the presence of this amino acid, Lentinula edodes was able to release about 2.5 μM of H2S, while Ganoderma lucidum released 2.1 μM of H2S after 30 min incubation and Polyporus umbellatus 2 μM. In the absence of cysteine, there was no increase in the concentration of H2S (Table 1 and Figure 6).
Three other extracts showed an H2S cysteine-mediated release but reached lower H2S levels: Inonotus obliquus P. Karst., which showed a release of 1.8 μM, Agaricus subrufescens Peck., with a release of 1.6 μM and Cordyceps sinensis Berk. with a release of 1.2 μM. For these extracts, the release of H2S in the absence of cysteine did not reach the instrument detection threshold (Table 1).
A further group could be identified using extracts with thiol-independent H2S-donor properties: Grifola frondosa Dicks. which, both in the presence and absence of cysteine, reached values of approximately 1.1 μM and Hericum erinaceus Bull. which reached values of approximately 1.2 μM (Table 1).
Finally, the last group could be identified by including those extracts devoid of H2S-donor properties, either in the presence or absence of cysteine: the extracts of Pleurotus ostreatus Jacq., Coprinus comatus O.F. Müll., Auricularia auricula Hook. f., Poria cocos F.A. Wolf and Phellinus igniarius L. did not reach the threshold of instrumental detectability for the release of H2S.
Based on quantitative analysis, all mushroom extracts contained significant levels of ergothioneine. Agaricus subrufescens Peck., Pleurotus ostreatus Jacq., Coprinus comatus O.F. Müll., Lentinula edodes Berk., and Grifola frondosa Dicks. appear to be the extracts with the highest ergothioneine content, far exceeding 200 μg per gram of extract. Nevertheless, the levels of the amino acid detected did not correlate with the release of H2S; therefore, we supposed that it did not significantly contribute to this property. Furthermore, using amperometric analysis, commercial ergothioneine incubated at a concentration of 1 mM in the presence and absence of cysteine showed a very modest release of H2S, amounting to 0.38 μM. This result suggests that despite the molecule’s chemical–physical potential, it is not capable of donating H2S, probably because cyclic thiourea does not exhibit the same reactivity as the previously tested thioureas [13].
The other organosulfur compound previously described in mushrooms is lenthionine. Therefore, we evaluated the release of H2S by testing the isolated compound at a concentration of 1 mM. In the presence of cysteine, the release of H2S from lenthionine reached very high concentrations (approximately 30 μM after 30 min of incubation) (Table 1). In contrast, in the absence of cysteine, lenthionine showed no release of H2S, suggesting that the cysteine-derived thiols can interact with lenthionine, thus leading to the hydrolysis of the molecule and release of H2S. Although this compound has already been described in the literature, this is the first time it has been identified as an H2S donor. Based on these results, it is possible to hypothesize that Lentinula edodes owes H2S-donor properties—at least in part—due to the presence of lenthionine. However, to our knowledge, this sulfur compound has not been identified in the other mushroom extracts that emerged from our screening as potential novel sources of H2S; therefore, it might be challenging to further characterize the content of organosulfur compounds in mushroom extracts that have been tested.
6. Conclusions
Alliaceae and Brassicaceae families were confirmed to be the main natural sources of organosulfur compounds, endowed with pleiotropic beneficial effects that reflect—under numerous aspects—those exerted by H2S.
In addition, among the vegetables in the Alliaceae and Brassicaceae families, mushrooms emerged as a new putative source of organosulfur compounds endowed with H2S-donor properties, and this hypothesis paves the way for a re-analysis of the health benefits of these vegetables and for the exploration of a new field of study in which the gasotransmitter is implicated.
These original data presented in this review represent a novelty in the scenario of research on naturally derived H2S donors. In this regard, a qualitative and quantitative analysis of the organosulfur compounds contained in the selected mushroom extracts will have to be carried out in order to associate the release of H2S with specific chemical entity/ies. It will be the first step in discovering new natural sources of H2S and studying further medicinal or nutraceutical applications of these vegetables.
Author Contributions
Conceptualization, L.T. and V.C. (Valentina Citi); methodology, V.C. (Valentina Citi); data curation, V.C. (Valentina Citi); writing—original draft preparation, L.T. and V.C. (Valentina Citi); writing—review and editing, V.C. (Vincenzo Calderone) and M.P.; supervision, V.C. (Vincenzo Calderone) and L.T.; funding acquisition, L.T. and V.C. (Vincenzo Calderone). All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by a grant entitled “Valorization of mushroom-based extracts as H2S-donors” founded by Golden Wave srl, Italy, grant number 449999_23_CPR.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Coburn, R.F. Carbon Monoxide (CO), Nitric Oxide, and Hydrogen Sulfide Signaling During Acute CO Poisoning. Front. Pharmacol. 2021, 12, 830241. [Google Scholar] [CrossRef]
- Ng, P.C.; Hendry-Hofer, T.B.; Witeof, A.E.; Brenner, M.; Mahon, S.B.; Boss, G.R.; Haouzi, P.; Bebarta, V.S. Hydrogen Sulfide Toxicity: Mechanism of Action, Clinical Presentation, and Countermeasure Development. J. Med. Toxicol. 2019, 15, 287–294. [Google Scholar] [CrossRef]
- Szabo, C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Bhatia, M.; Gaddam, R.R. Hydrogen Sulfide in Inflammation: A Novel Mediator and Therapeutic Target. Antioxid. Redox Signal. 2021, 34, 1368–1377. [Google Scholar] [CrossRef]
- Chen, T.; Tian, M.; Han, Y. Hydrogen sulfide: A multi-tasking signal molecule in the regulation of oxidative stress responses. J. Exp. Bot. 2020, 71, 2862–2869. [Google Scholar] [CrossRef]
- Kimura, H.; Shibuya, N.; Kimura, Y. Hydrogen sulfide is a signaling molecule and a cytoprotectant. Antioxid. Redox Signal. 2012, 17, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Dombkowski, R.A.; Russell, M.J.; Olson, K.R. Hydrogen sulfide as an endogenous regulator of vascular smooth muscle tone in trout. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R678–R685. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Gu, Y.; Zhu, D. Cardioprotective effects of hydrogen sulfide in attenuating myocardial ischemia-reperfusion injury (Review). Mol. Med. Rep. 2021, 24, 875. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, Y.; Chen, S.; Tang, C.; Wang, G.; Du, J.; Jin, H. Hydrogen sulfide regulates insulin secretion and insulin resistance in diabetes mellitus, a new promising target for diabetes mellitus treatment? A review. J. Adv. Res. 2021, 27, 19–30. [Google Scholar] [CrossRef]
- Pan, L.L.; Qin, M.; Liu, X.H.; Zhu, Y.Z. The Role of Hydrogen Sulfide on Cardiovascular Homeostasis: An Overview with Update on Immunomodulation. Front. Pharmacol. 2017, 8, 686. [Google Scholar] [CrossRef] [Green Version]
- Munteanu, C.; Rotariu, M.; Turnea, M.; Dogaru, G.; Popescu, C.; Spinu, A.; Andone, I.; Postoiu, R.; Ionescu, E.V.; Oprea, C.; et al. Recent Advances in Molecular Research on Hydrogen Sulfide (H2S) Role in Diabetes Mellitus (DM)-A Systematic Review. Int. J. Mol. Sci. 2022, 23, 6720. [Google Scholar] [CrossRef]
- Lv, B.; Chen, S.; Tang, C.; Jin, H.; Du, J.; Huang, Y. Hydrogen sulfide and vascular regulation—An update. J. Adv. Res. 2021, 27, 85–97. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Bucci, M.; Piragine, E.; Testai, L.; Vellecco, V.; Cirino, G.; Calderone, V. Searching for novel hydrogen sulfide donors: The vascular effects of two thiourea derivatives. Pharmacol. Res. 2020, 159, 105039. [Google Scholar] [CrossRef]
- Citi, V.; Corvino, A.; Fiorino, F.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Brogi, S.; Flori, L.; Gorica, E.; et al. Structure-activity relationships study of isothiocyanates for H2S releasing properties: 3-Pyridyl-isothiocyanate as a new promising cardioprotective agent. J. Adv. Res. 2021, 27, 41–53. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.G.; Ghimenti, S.; Breschi, M.C.; Calderone, V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef]
- Martelli, A.; Citi, V.; Testai, L.; Brogi, S.; Calderone, V. Organic Isothiocyanates as Hydrogen Sulfide Donors. Antioxid. Redox Signal. 2020, 32, 110–144. [Google Scholar] [CrossRef]
- Severino, B.; Corvino, A.; Fiorino, F.; Luciano, P.; Frecentese, F.; Magli, E.; Saccone, I.; Di Vaio, P.; Citi, V.; Calderone, V.; et al. 1,2,4-Thiadiazolidin-3,5-diones as novel hydrogen sulfide donors. Eur. J. Med. Chem. 2018, 143, 1677–1686. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Pugliesi, I.; Barresi, E.; Nesi, G.; Rapposelli, S.; Taliani, S.; Da Settimo, F.; et al. Arylthioamides as H2S Donors: l-Cysteine-Activated Releasing Properties and Vascular Effects in Vitro and in Vivo. ACS Med. Chem. Lett. 2013, 4, 904–908. [Google Scholar] [CrossRef] [Green Version]
- Barresi, E.; Nesi, G.; Citi, V.; Piragine, E.; Piano, I.; Taliani, S.; Da Settimo, F.; Rapposelli, S.; Testai, L.; Breschi, M.C.; et al. Iminothioethers as Hydrogen Sulfide Donors: From the Gasotransmitter Release to the Vascular Effects. J. Med. Chem. 2017, 60, 7512–7523. [Google Scholar] [CrossRef]
- Calderone, V.; Martelli, A.; Testai, L.; Citi, V.; Breschi, M.C. Using hydrogen sulfide to design and develop drugs. Expert. Opin. Drug Discov. 2016, 11, 163–175. [Google Scholar] [CrossRef]
- Sestito, S.; Daniele, S.; Pietrobono, D.; Citi, V.; Bellusci, L.; Chiellini, G.; Calderone, V.; Martini, C.; Rapposelli, S. Memantine prodrug as a new agent for Alzheimer’s Disease. Sci. Rep. 2019, 9, 4612. [Google Scholar] [CrossRef] [Green Version]
- Sestito, S.; Pruccoli, L.; Runfola, M.; Citi, V.; Martelli, A.; Saccomanni, G.; Calderone, V.; Tarozzi, A.; Rapposelli, S. Design and synthesis of H2S-donor hybrids: A new treatment for Alzheimer’s disease? Eur. J. Med. Chem. 2019, 184, 111745. [Google Scholar] [CrossRef]
- Rapposelli, S.; Gambari, L.; Digiacomo, M.; Citi, V.; Lisignoli, G.; Manferdini, C.; Calderone, V.; Grassi, F. A Novel H2S-releasing Amino-Bisphosphonate which combines bone anti-catabolic and anabolic functions. Sci. Rep. 2017, 7, 11940. [Google Scholar] [CrossRef] [Green Version]
- Ercolano, G.; De Cicco, P.; Frecentese, F.; Saccone, I.; Corvino, A.; Giordano, F.; Magli, E.; Fiorino, F.; Severino, B.; Calderone, V.; et al. Anti-metastatic Properties of Naproxen-HBTA in a Murine Model of Cutaneous Melanoma. Front. Pharmacol. 2019, 10, 66. [Google Scholar] [CrossRef]
- Corvino, A.; Citi, V.; Fiorino, F.; Frecentese, F.; Magli, E.; Perissutti, E.; Santagada, V.; Calderone, V.; Martelli, A.; Gorica, E.; et al. H2S donating corticosteroids: Design, synthesis and biological evaluation in a murine model of asthma. J. Adv. Res. 2022, 35, 267–277. [Google Scholar] [CrossRef]
- Sparaco, R.; Citi, V.; Magli, E.; Martelli, A.; Piragine, E.; Calderone, V.; Andreozzi, G.; Perissutti, E.; Frecentese, F.; Santagada, V.; et al. Design, Synthesis and Evaluation of Novel Molecular Hybrids between Antiglaucoma Drugs and H2S Donors. Int. J. Mol. Sci. 2022, 23, 3804. [Google Scholar] [CrossRef]
- Giordano, F.; Corvino, A.; Scognamiglio, A.; Citi, V.; Gorica, E.; Fattorusso, C.; Persico, M.; Caliendo, G.; Fiorino, F.; Magli, E.; et al. Hybrids between H2S-donors and betamethasone 17-valerate or triamcinolone acetonide inhibit mast cell degranulation and promote hyperpolarization of bronchial smooth muscle cells. Eur. J. Med. Chem. 2021, 221, 113517. [Google Scholar] [CrossRef]
- Gu, X.; Zhu, Y.Z. Therapeutic applications of organosulfur compounds as novel hydrogen sulfide donors and/or mediators. Expert Rev. Clin. Pharmacol. 2011, 4, 123–133. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Testai, L.; Marino, A.; Breschi, M.C.; Calderone, V. Hydrogen sulfide releasing capacity of natural isothiocyanates: Is it a reliable explanation for the multiple biological effects of Brassicaceae? Planta Med. 2014, 80, 610–613. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Kopec, A.; Piatkowska, E.; Borczak, B.; Leszczynska, T. The beneficial effects of Brassica vegetables on human health. Rocz. Panstw. Zakl. Hig. 2012, 63, 389–395. [Google Scholar]
- Martelli, A.; Testai, L.; Breschi, M.C.; Blandizzi, C.; Virdis, A.; Taddei, S.; Calderone, V. Hydrogen sulphide: Novel opportunity for drug discovery. Med. Res. Rev. 2012, 32, 1093–1130. [Google Scholar] [CrossRef]
- Cirino, G.; Szabo, C.; Papapetropoulos, A. Physiological roles of hydrogen sulfide in mammalian cells, tissues, and organs. Physiol. Rev. 2023, 103, 31–276. [Google Scholar] [CrossRef]
- Testai, L.; Citi, V.; Martelli, A.; Brogi, S.; Calderone, V. Role of hydrogen sulfide in cardiovascular ageing. Pharmacol. Res. 2020, 160, 105125. [Google Scholar] [CrossRef]
- Randi, E.B.; Casili, G.; Jacquemai, S.; Szabo, C. Selenium-Binding Protein 1 (SELENBP1) Supports Hydrogen Sulfide Biosynthesis and Adipogenesis. Antioxidants 2021, 10, 361. [Google Scholar] [CrossRef]
- Juan, C.A.; Perez de la Lastra, J.M.; Plou, F.J.; Perez-Lebena, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Shefa, U.; Kim, M.S.; Jeong, N.Y.; Jung, J. Antioxidant and Cell-Signaling Functions of Hydrogen Sulfide in the Central Nervous System. Oxid. Med. Cell. Longev. 2018, 2018, 1873962. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.Z.; Liu, Y.; Bian, J.S. Hydrogen Sulfide and Cellular Redox Homeostasis. Oxid. Med. Cell. Longev. 2016, 2016, 6043038. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Gu, Y.; Wen, M.; Zhao, S.; Wang, W.; Ma, Y.; Meng, G.; Han, Y.; Wang, Y.; Liu, G.; et al. Hydrogen Sulfide Induces Keap1 S-sulfhydration and Suppresses Diabetes-Accelerated Atherosclerosis via Nrf2 Activation. Diabetes 2016, 65, 3171–3184. [Google Scholar] [CrossRef] [Green Version]
- Cornwell, A.; Badiei, A. From Gasotransmitter to Immunomodulator: The Emerging Role of Hydrogen Sulfide in Macrophage Biology. Antioxidants 2023, 12, 935. [Google Scholar] [CrossRef]
- Marino, A.; Martelli, A.; Citi, V.; Fu, M.; Wang, R.; Calderone, V.; Levi, R. The novel H2S donor 4-carboxy-phenyl isothiocyanate inhibits mast cell degranulation and renin release by decreasing intracellular calcium. Br. J. Pharmacol. 2016, 173, 3222–3234. [Google Scholar] [CrossRef] [Green Version]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-kappaB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Citi, V.; Martelli, A.; Brancaleone, V.; Brogi, S.; Gojon, G.; Montanaro, R.; Morales, G.; Testai, L.; Calderone, V. Anti-inflammatory and antiviral roles of hydrogen sulfide: Rationale for considering H2S donors in COVID-19 therapy. Br. J. Pharmacol. 2020, 177, 4931–4941. [Google Scholar] [CrossRef]
- Sun, F.; Luo, J.H.; Yue, T.T.; Wang, F.X.; Yang, C.L.; Zhang, S.; Wang, X.Q.; Wang, C.Y. The role of hydrogen sulphide signalling in macrophage activation. Immunology 2021, 162, 3–10. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Du, J.; Huang, Y.; Tang, C.; Jin, H. Hydrogen Sulfide Regulates Macrophage Function in Cardiovascular Diseases. Antioxid. Redox Signal. 2023, 38, 45–56. [Google Scholar] [CrossRef]
- Dilek, N.; Papapetropoulos, A.; Toliver-Kinsky, T.; Szabo, C. Hydrogen sulfide: An endogenous regulator of the immune system. Pharmacol. Res. 2020, 161, 105119. [Google Scholar] [CrossRef]
- Sunzini, F.; De Stefano, S.; Chimenti, M.S.; Melino, S. Hydrogen Sulfide as Potential Regulatory Gasotransmitter in Arthritic Diseases. Int. J. Mol. Sci. 2020, 21, 1180. [Google Scholar] [CrossRef] [Green Version]
- D’Araio, E.; Shaw, N.; Millward, A.; Demaine, A.; Whiteman, M.; Hodgkinson, A. Hydrogen sulfide induces heme oxygenase-1 in human kidney cells. Acta Diabetol. 2014, 51, 155–157. [Google Scholar] [CrossRef]
- Wu, T.; Li, H.; Wu, B.; Zhang, L.; Wu, S.W.; Wang, J.N.; Zhang, Y.E. Hydrogen Sulfide Reduces Recruitment of CD11b+Gr-1+ Cells in Mice With Myocardial Infarction. Cell Transplant. 2017, 26, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Farahat, S.; Kherkheulidze, S.; Nopp, S.; Kainz, A.; Borriello, M.; Perna, A.F.; Cohen, G. Effect of Hydrogen Sulfide on Essential Functions of Polymorphonuclear Leukocytes. Toxins 2023, 15, 198. [Google Scholar] [CrossRef]
- Kumar, A.; Bhatia, M. Role of Hydrogen Sulfide, Substance P and Adhesion Molecules in Acute Pancreatitis. Int. J. Mol. Sci. 2021, 22, 12136. [Google Scholar] [CrossRef]
- Toliver-Kinsky, T.; Cui, W.; Toro, G.; Lee, S.J.; Shatalin, K.; Nudler, E.; Szabo, C. H2S, a Bacterial Defense Mechanism against the Host Immune Response. Infect. Immun. 2019, 87, 10–1128. [Google Scholar] [CrossRef] [Green Version]
- Bibli, S.I.; Hu, J.; Looso, M.; Weigert, A.; Ratiu, C.; Wittig, J.; Drekolia, M.K.; Tombor, L.; Randriamboavonjy, V.; Leisegang, M.S.; et al. Mapping the Endothelial Cell S-Sulfhydrome Highlights the Crucial Role of Integrin Sulfhydration in Vascular Function. Circulation 2021, 143, 935–948. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Gorica, E.; Brogi, S.; Testai, L.; Calderone, V. Role of hydrogen sulfide in endothelial dysfunction: Pathophysiology and therapeutic approaches. J. Adv. Res. 2021, 27, 99–113. [Google Scholar] [CrossRef]
- Jiang, B.; Tang, G.; Cao, K.; Wu, L.; Wang, R. Molecular mechanism for H2S-induced activation of K(ATP) channels. Antioxid. Redox Signal. 2010, 12, 1167–1178. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Breschi, M.C.; Lawson, K.; McKay, N.G.; Miceli, F.; Taglialatela, M.; Calderone, V. Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol. Res. 2013, 70, 27–34. [Google Scholar] [CrossRef]
- Bucci, M.; Papapetropoulos, A.; Vellecco, V.; Zhou, Z.; Pyriochou, A.; Roussos, C.; Roviezzo, F.; Brancaleone, V.; Cirino, G. Hydrogen sulfide is an endogenous inhibitor of phosphodiesterase activity. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1998–2004. [Google Scholar] [CrossRef] [Green Version]
- Testai, L.; Marino, A.; Piano, I.; Brancaleone, V.; Tomita, K.; Di Cesare Mannelli, L.; Martelli, A.; Citi, V.; Breschi, M.C.; Levi, R.; et al. The novel H2S-donor 4-carboxyphenyl isothiocyanate promotes cardioprotective effects against ischemia/reperfusion injury through activation of mitoK(ATP) channels and reduction of oxidative stress. Pharmacol. Res. 2016, 113, 290–299. [Google Scholar] [CrossRef]
- Citi, V.; Piragine, E.; Testai, L.; Breschi, M.C.; Calderone, V.; Martelli, A. The Role of Hydrogen Sulfide and H2S-donors in Myocardial Protection Against Ischemia/Reperfusion Injury. Curr. Med. Chem. 2018, 25, 4380–4401. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; D’Antongiovanni, V.; Piano, I.; Martelli, A.; Citi, V.; Duranti, E.; Virdis, A.; Blandizzi, C.; Gargini, C.; Breschi, M.C.; et al. Different patterns of H2S/NO activity and cross-talk in the control of the coronary vascular bed under normotensive or hypertensive conditions. Nitric Oxide 2015, 47, 25–33. [Google Scholar] [CrossRef]
- Wang, M.; Tang, W.; Zhu, Y.Z. An Update on AMPK in Hydrogen Sulfide Pharmacology. Front. Pharmacol. 2017, 8, 810. [Google Scholar] [CrossRef] [Green Version]
- Walewska, A.; Szewczyk, A.; Koprowski, P. Gas Signaling Molecules and Mitochondrial Potassium Channels. Int. J. Mol. Sci. 2018, 19, 3227. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Ju, Y.; Fu, M.; Zhang, Y.; Pei, Y.; Racine, M.; Baath, S.; Merritt, T.J.S.; Wang, R.; Wu, L. Cystathionine gamma-lyase/hydrogen sulfide system is essential for adipogenesis and fat mass accumulation in mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 165–176. [Google Scholar] [CrossRef]
- Cai, J.; Shi, X.; Wang, H.; Fan, J.; Feng, Y.; Lin, X.; Yang, J.; Cui, Q.; Tang, C.; Xu, G.; et al. Cystathionine gamma lyase-hydrogen sulfide increases peroxisome proliferator-activated receptor gamma activity by sulfhydration at C139 site thereby promoting glucose uptake and lipid storage in adipocytes. Biochim. Biophys. Acta 2016, 1861, 419–429. [Google Scholar] [CrossRef]
- Recinella, L.; Libero, M.L.; Citi, V.; Chiavaroli, A.; Martelli, A.; Foligni, R.; Mannozzi, C.; Acquaviva, A.; Di Simone, S.; Calderone, V.; et al. Anti-Inflammatory and Vasorelaxant Effects Induced by an Aqueous Aged Black Garlic Extract Supplemented with Vitamins D, C, and B12 on Cardiovascular System. Foods 2023, 12, 1558. [Google Scholar] [CrossRef]
- Piragine, E.; Citi, V.; Lawson, K.; Calderone, V.; Martelli, A. Potential Effects of Natural H2S-Donors in Hypertension Management. Biomolecules 2022, 12, 581. [Google Scholar] [CrossRef]
- Piragine, E.; Citi, V.; Lawson, K.; Calderone, V.; Martelli, A. Regulation of blood pressure by natural sulfur compounds: Focus on their mechanisms of action. Biochem. Pharmacol. 2022, 206, 115302. [Google Scholar] [CrossRef]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the flavour precursors of onion and garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Rani, V. Therapeutic Potential of Stable Organosulfur Compounds of Aged Garlic. Cardiovasc. Hematol. Agents Med. Chem. 2023, 21, 84–95. [Google Scholar] [CrossRef]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef]
- Thomson, M.; Al-Qattan, K.K.; Js, D.; Ali, M. Anti-diabetic and anti-oxidant potential of aged garlic extract (AGE) in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2016, 16, 17. [Google Scholar] [CrossRef] [Green Version]
- Iciek, M.; Kwiecien, I.; Wlodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen. 2009, 50, 247–265. [Google Scholar] [CrossRef]
- Piragine, E.; Calderone, V. Pharmacological modulation of the hydrogen sulfide (H2S) system by dietary H2S-donors: A novel promising strategy in the prevention and treatment of type 2 diabetes mellitus. Phytother. Res. 2021, 35, 1817–1846. [Google Scholar] [CrossRef]
- Banerjee, S.K.; Maulik, S.K. Effect of garlic on cardiovascular disorders: A review. Nutr. J. 2002, 1, 4. [Google Scholar] [CrossRef]
- Melino, S.; Leo, S.; Toska Papajani, V. Natural Hydrogen Sulfide Donors from Allium sp. as a Nutraceutical Approach in Type 2 Diabetes Prevention and Therapy. Nutrients 2019, 11, 1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.P.; Sun, H.J.; Cao, X.; Wu, Z.Y.; Zhu, M.Y.; Cao, L.; Nie, X.W.; Bian, J.S. Polysulfide Protects Against Diabetic Cardiomyopathy Through Sulfhydration of Peroxisome Proliferator-Activated Receptor-gamma and Sirtuin 3. Antioxid. Redox Signal. 2023, 38, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Miekus, N.; Marszalek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Swiergiel, A.H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yang, X.; Lu, Y.; Liang, D.; Huang, D. Isothiocyanates as H2S Donors Triggered by Cysteine: Reaction Mechanism and Structure and Activity Relationship. Org. Lett. 2019, 21, 5977–5980. [Google Scholar] [CrossRef]
- Martelli, A.; Piragine, E.; Citi, V.; Testai, L.; Pagnotta, E.; Ugolini, L.; Lazzeri, L.; Di Cesare Mannelli, L.; Manzo, O.L.; Bucci, M.; et al. Erucin exhibits vasorelaxing effects and antihypertensive activity by H2S-releasing properties. Br. J. Pharmacol. 2020, 177, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, Y.; Liu, X.; Lin, Y.; Zheng, X.; Lu, Y. Hydrogen Sulfide (H2S) Releasing Capacity of Isothiocyanates from Moringa oleifera Lam. Molecules 2018, 23, 2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, A.; Piragine, E.; Gorica, E.; Citi, V.; Testai, L.; Pagnotta, E.; Lazzeri, L.; Pecchioni, N.; Ciccone, V.; Montanaro, R.; et al. The H2S-Donor Erucin Exhibits Protective Effects against Vascular Inflammation in Human Endothelial and Smooth Muscle Cells. Antioxidants 2021, 10, 961. [Google Scholar] [CrossRef]
- Testai, L.; Pagnotta, E.; Piragine, E.; Flori, L.; Citi, V.; Martelli, A.; Mannelli, L.D.C.; Ghelardini, C.; Matteo, R.; Suriano, S.; et al. Cardiovascular benefits of Eruca sativa mill. Defatted seed meal extract: Potential role of hydrogen sulfide. Phytother. Res. 2022, 36, 2616–2627. [Google Scholar] [CrossRef]
- Piragine, E.; Petri, D.; Giometto, S.; Martelli, A.; Lucenteforte, E.; Calderone, V. Potential effects of Alliaceae and Brassicaceae edible plants on blood glucose levels in patients with type 2 diabetes: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2022, 185, 106519. [Google Scholar] [CrossRef]
- Abd Rani, N.Z.; Husain, K.; Kumolosasi, E. Moringa Genus: A Review of Phytochemistry and Pharmacology. Front. Pharmacol. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, S.W.; Hoskin, R.T.; Komarnytsky, S.; Moncada, M. Mushrooms as Functional and Nutritious Food Ingredients for Multiple Applications. ACS Food Sci. Technol. 2022, 2, 1184–1195. [Google Scholar] [CrossRef]
- Krittanawong, C.; Isath, A.; Hahn, J.; Wang, Z.; Fogg, S.E.; Bandyopadhyay, D.; Jneid, H.; Virani, S.S.; Tang, W.H.W. Mushroom Consumption and Cardiovascular Health: A Systematic Review. Am. J. Med. 2021, 134, 637–642.e632. [Google Scholar] [CrossRef] [PubMed]
- Meneses, M.E.; Galicia-Castillo, M.; Perez-Herrera, A.; Martinez, R.; Leon, H.; Martinez-Carrera, D. Traditional Mushroom Consumption Associated to Lower Levels of Triglycerides and Blood Pressure in an Indigenous Peasant Community from Oaxaca, Mexico. Int. J. Med. Mushrooms 2020, 22, 953–966. [Google Scholar] [CrossRef]
- Uffelman, C.N.; Chan, N.I.; Davis, E.M.; Wang, Y.; McGowan, B.S.; Campbell, W.W. An Assessment of Mushroom Consumption on Cardiometabolic Disease Risk Factors and Morbidities in Humans: A Systematic Review. Nutrients 2023, 15, 1079. [Google Scholar] [CrossRef]
- Shevchuk, Y.; Kuypers, K.; Janssens, G.E. Fungi as a source of bioactive molecules for the development of longevity medicines. Ageing Res. Rev. 2023, 87, 101929. [Google Scholar] [CrossRef]
- Liuzzi, G.M.; Petraglia, T.; Latronico, T.; Crescenzi, A.; Rossano, R. Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases. Nutrients 2023, 15, 1913. [Google Scholar] [CrossRef]
- Sousa, A.S.; Araujo-Rodrigues, H.; Pintado, M.E. The Health-promoting Potential of Edible Mushroom Proteins. Curr. Pharm. Des. 2023, 29, 804–823. [Google Scholar] [CrossRef]
- Bell, V.; Silva, C.; Guina, J.; Fernandes, T.H. Mushrooms as future generation healthy foods. Front. Nutr. 2022, 9, 1050099. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, J.; Chen, W.; Yang, Y.; Zhang, J.; Feng, J.; Yu, H.; Li, Q. Analysis of volatile compounds of Lentinula edodes grown in different culture substrate formulations. Food Res. Int. 2019, 125, 108517. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.; Chen, X.; Han, D.; Han, J.; Wang, L.; Ren, A.; Yu, H.; Zhao, M. Lenthionine, a Key Flavor Substance in Lentinula edodes, Is Regulated by Cysteine under Drought Stress. J. Agric. Food Chem. 2021, 69, 12645–12653. [Google Scholar] [CrossRef]
- Liu, Q.; Mao, Y.; Liao, X.; Luo, J.; Ma, H.; Jiang, W. Recent progress in ergothioneine biosynthesis: A review. Sheng Wu Gong Cheng Xue Bao 2022, 38, 1408–1420. [Google Scholar] [CrossRef]
- Hiraide, M.; Miyazaki, Y.; Shibata, Y. The smell and odorous components of dried shiitake mushroom, Lentinula edodes I: Relationship between sensory evaluations and amounts of odorous components. J. Wood Sci. 2004, 50, 358–364. [Google Scholar] [CrossRef]
- Chen, C.-C.; Ho, C.-T. High-performance liquid chromatographic determination of cyclic sulfur compounds of Shiitake mushroom (Lentinus edodes Sing.). J. Chromatogr. A 1986, 356, 455–459. [Google Scholar] [CrossRef]
- Kupcova, K.; Stefanova, I.; Plavcova, Z.; Hosek, J.; Hrouzek, P.; Kubec, R. Antimicrobial, Cytotoxic, Anti-Inflammatory, and Antioxidant Activity of Culinary Processed Shiitake Medicinal Mushroom (Lentinus edodes, Agaricomycetes) and Its Major Sulfur Sensory-Active Compound-Lenthionine. Int. J. Med. Mushrooms 2018, 20, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lei, X.Y.; Chen, L.F.; Bian, Y.B.; Yang, H.; Ibrahim, S.A.; Huang, W. A novel cysteine desulfurase influencing organosulfur compounds in Lentinula edodes. Sci. Rep. 2015, 5, 10047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Gao, J.X.; Xue, J.; Chen, D.; Lin, S.Y.; Dong, X.P.; Zhu, B.W. Changes in Aroma Profile of Shiitake Mushroom (Lentinus edodes) during Different Stages of Hot Air Drying. Foods 2020, 9, 444. [Google Scholar] [CrossRef] [Green Version]
- Dermiki, M.; Phanphensophon, N.; Mottram, D.S.; Methven, L. Contributions of non-volatile and volatile compounds to the umami taste and overall flavour of shiitake mushroom extracts and their application as flavour enhancers in cooked minced meat. Food Chem. 2013, 141, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Brancaleone, V.; Esposito, I.; Gargiulo, A.; Vellecco, V.; Asimakopoulou, A.; Citi, V.; Calderone, V.; Gobbetti, T.; Perretti, M.; Papapetropoulos, A.; et al. D-Penicillamine modulates hydrogen sulfide (H2S) pathway through selective inhibition of cystathionine-gamma-lyase. Br. J. Pharmacol. 2016, 173, 1556–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
H2S-related disorders.

Figure 2.
The figure schematically shows the main pharmacological activities of hydrogen sulfide, (A) H2S and inflammation: through the sulfhydration of IKkB, H2S inhibits the dissociation between Nf-kB and IkB, impeding the translocation of Nf-kB into the nucleus and promoting anti-inflammatory effects. (B) H2S and oxidative stress: H2S sulfhydrates Keap1 allowing the dissociation of Nrf-2 which translocates into the nucleus and promotes the expression of anti-oxidant species. Furthermore, H2S acts as a direct scavenger of ROS. (C) H2S and vasodilation: H2S induces the opening of potassium channels and inhibits PDE5, leading to hyperpolarization of vascular cells and increase in NO levels. (D) H2S and immune system: H2S facilitates the differentiation in M2 polarized macrophages, which produce anti-inflammatory mediators. (E) H2S and metabolism: H2S sulfhydrates PPAR- γ, increasing insulin sensitivity and lipid synthesis. Abbreviations: Nf-kB: nuclear factor kappa-light-chain-enhancer of activated B cells; IkB: IkappaB kinase; IKkB: inhibitor of nuclear factor kappa-B kinase subunit beta; P: phosphate; Nrf-2: Nuclear factor erythroid 2-related factor 2; Keap1: Kelch-like ECH-associated protein 1; ARE: anti-oxidant responsive elements; KATP: ATP-sensitive potassium channel; Kv7: voltage-gated potassium channels; PDE5: Phosphodiesterase Type 5; PPAR—γ: Peroxisome proliferator-activated receptor gamma.
Figure 2.
The figure schematically shows the main pharmacological activities of hydrogen sulfide, (A) H2S and inflammation: through the sulfhydration of IKkB, H2S inhibits the dissociation between Nf-kB and IkB, impeding the translocation of Nf-kB into the nucleus and promoting anti-inflammatory effects. (B) H2S and oxidative stress: H2S sulfhydrates Keap1 allowing the dissociation of Nrf-2 which translocates into the nucleus and promotes the expression of anti-oxidant species. Furthermore, H2S acts as a direct scavenger of ROS. (C) H2S and vasodilation: H2S induces the opening of potassium channels and inhibits PDE5, leading to hyperpolarization of vascular cells and increase in NO levels. (D) H2S and immune system: H2S facilitates the differentiation in M2 polarized macrophages, which produce anti-inflammatory mediators. (E) H2S and metabolism: H2S sulfhydrates PPAR- γ, increasing insulin sensitivity and lipid synthesis. Abbreviations: Nf-kB: nuclear factor kappa-light-chain-enhancer of activated B cells; IkB: IkappaB kinase; IKkB: inhibitor of nuclear factor kappa-B kinase subunit beta; P: phosphate; Nrf-2: Nuclear factor erythroid 2-related factor 2; Keap1: Kelch-like ECH-associated protein 1; ARE: anti-oxidant responsive elements; KATP: ATP-sensitive potassium channel; Kv7: voltage-gated potassium channels; PDE5: Phosphodiesterase Type 5; PPAR—γ: Peroxisome proliferator-activated receptor gamma.

Figure 3.
Schematic representation of the biosyntethic reactions leading to the formation of allylsulfur compounds in Alliaceae vegetables.
Figure 3.
Schematic representation of the biosyntethic reactions leading to the formation of allylsulfur compounds in Alliaceae vegetables.

Figure 4.
Mechanism of the release of H2S from allylsulfur compounds present in Alliaceae vegetables. The reaction is triggered by a nucleophilic attack of organic thiols (here represented by glutathione (GSH)) that promotes the formation of H2S and perthiols, which generate another molecule of H2S and glutathione disulfide (GSSG).
Figure 4.
Mechanism of the release of H2S from allylsulfur compounds present in Alliaceae vegetables. The reaction is triggered by a nucleophilic attack of organic thiols (here represented by glutathione (GSH)) that promotes the formation of H2S and perthiols, which generate another molecule of H2S and glutathione disulfide (GSSG).

Figure 5.
Mechanism of the release of H2S from organic isothiocyanates (ITCs) compounds present in Brassicaceae vegetables. The reaction is triggered by the nucleophilic attack of thiols (here represented by cysteine) and leads to the formation of an instable intermediate, that spontaneously decomposes in H2S and aminic derivatives.
Figure 5.
Mechanism of the release of H2S from organic isothiocyanates (ITCs) compounds present in Brassicaceae vegetables. The reaction is triggered by the nucleophilic attack of thiols (here represented by cysteine) and leads to the formation of an instable intermediate, that spontaneously decomposes in H2S and aminic derivatives.

Figure 6.
Amperometric recordings of H2S release after the incubation of Lentinula edodes (a), Ganoderma lucidum (b) and Polyporus umbellatus (c) 1 mg/mL in the presence or absence of L-Cysteine. Data are expressed as mean ± SEM.
Figure 6.
Amperometric recordings of H2S release after the incubation of Lentinula edodes (a), Ganoderma lucidum (b) and Polyporus umbellatus (c) 1 mg/mL in the presence or absence of L-Cysteine. Data are expressed as mean ± SEM.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Maximum H2S release after the incubation of the mushroom extracts or ergothioneine and lenthionine.
Table 1.
Maximum H2S release after the incubation of the mushroom extracts or ergothioneine and lenthionine.
| Tested Item | Ergothioneine Content (µg/g) | H2S Release in the Presence of L-Cys 4 mM (μM) | H2S Release in the Absence of L-Cys (μM) |
|---|---|---|---|
| Ganoderma lucidum | 39.1 ± 0.2 | 2.1 ± 0.2 | n.d. |
| Hericum erinaceus | 184.6 ± 0.9 | 1.2 ± 0.2 | 1.2 ± 0.3 |
| Lentinula edodes | 348.3 ± 1.2 | 2.5 ± 0.1 | n.d. |
| Grifola frondose | 205.1 ± 3.4 | 1.1 ± 0.2 | 1.0 ± 0.2 |
| Polyporus umbellatus | 25.7 ± 0.8 | 2.0 ± 0.6 | n.d. |
| Auricularia auricola | 4.24 ± 0.1 | n.d. | n.d. |
| Inonotus obliquus | 0.3 ± 0.0 | 1.8 ± 0.1 | n.d. |
| Agaricus subrufescens | 479.3 ± 4.6 | 1.6 ± 0.4 | n.d. |
| Poria cocos | 5.4 ± 0.3 | n.d. | n.d. |
| Pleurotus ostreatus | 477.7 ± 2.7 | n.d. | n.d. |
| Coprinus comatus | 450.2 ± 4.8 | n.d. | n.d. |
| Cordyceps sinensis | 5.9 ± 0.1 | 1.2 ± 0.4 | n.d. |
| Ergothioneine | n.d. | n.d. | n.d. |
| Lenthionine | n.d. | 32 ± 4.3 | n.d. |
Abbreviations: L-Cys: L-Cysteine; n.d.: not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Citi, V.; Passerini, M.; Calderone, V.; Testai, L. Plants and Mushrooms as Possible New Sources of H2S Releasing Sulfur Compounds. Int. J. Mol. Sci. 2023, 24, 11886. https://doi.org/10.3390/ijms241511886
AMA Style
Citi V, Passerini M, Calderone V, Testai L. Plants and Mushrooms as Possible New Sources of H2S Releasing Sulfur Compounds. International Journal of Molecular Sciences. 2023; 24(15):11886. https://doi.org/10.3390/ijms241511886
Chicago/Turabian StyleCiti, Valentina, Marco Passerini, Vincenzo Calderone, and Lara Testai. 2023. "Plants and Mushrooms as Possible New Sources of H2S Releasing Sulfur Compounds" International Journal of Molecular Sciences 24, no. 15: 11886. https://doi.org/10.3390/ijms241511886
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.