
Hepatic Global Transcriptomic Profiles of Holstein Cows According to Parity Reveal Age-Related Changes in Early Lactation
 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Blood Metabolites and Animal Performance Traits between the Three Lactation Groups
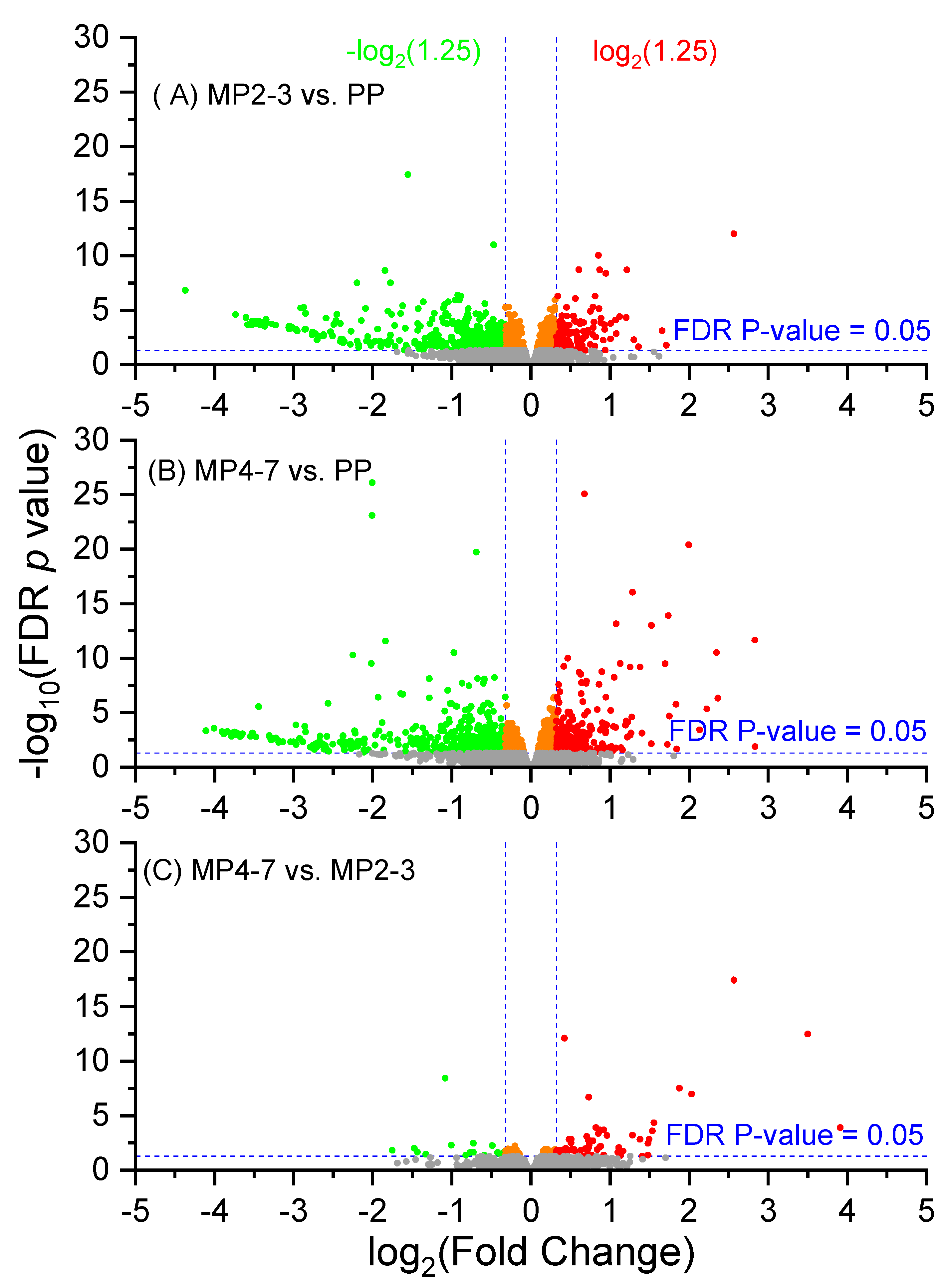
2.2. Hepatic Gene Expression Profiles between the Three Lactation Groups
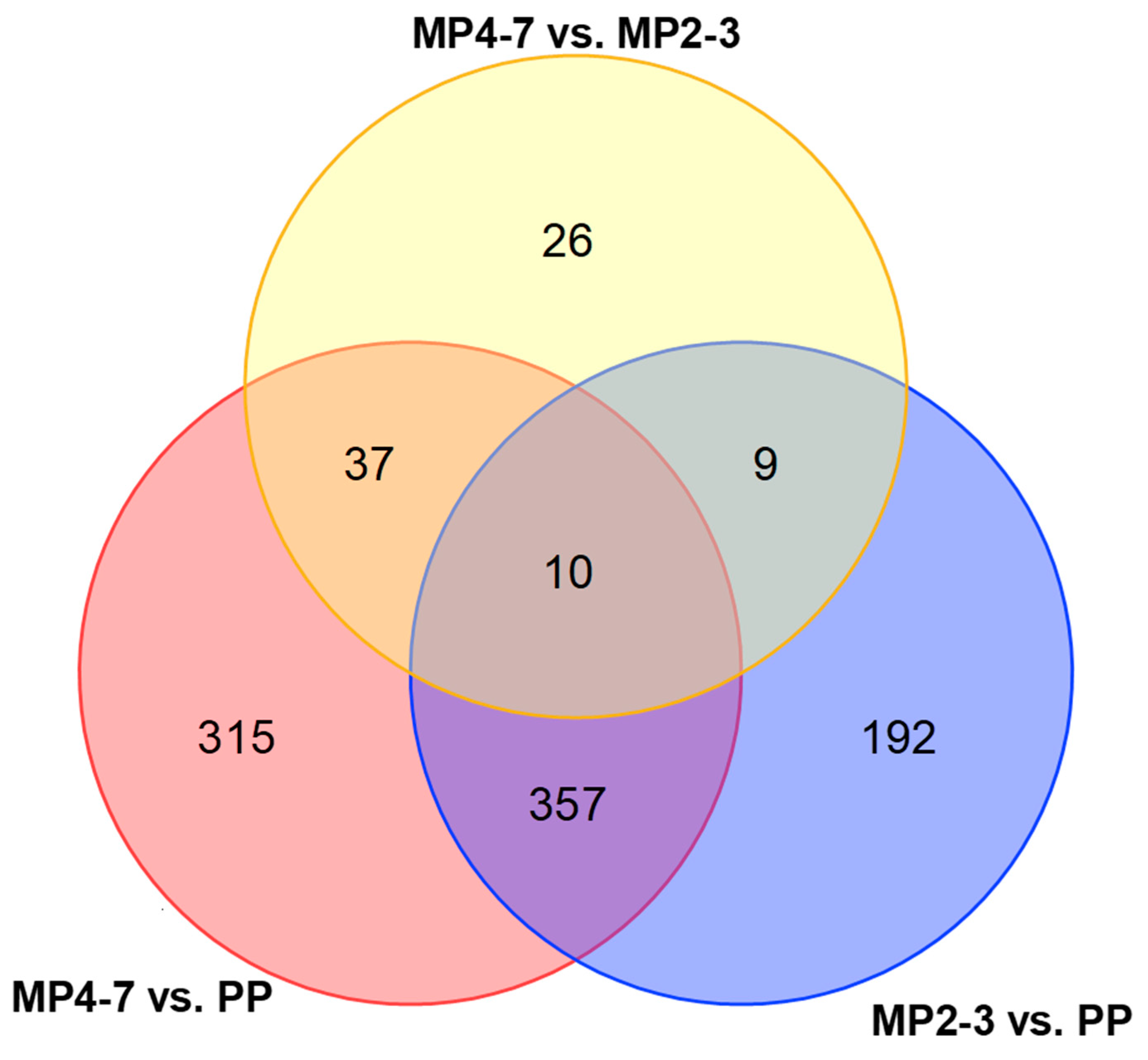
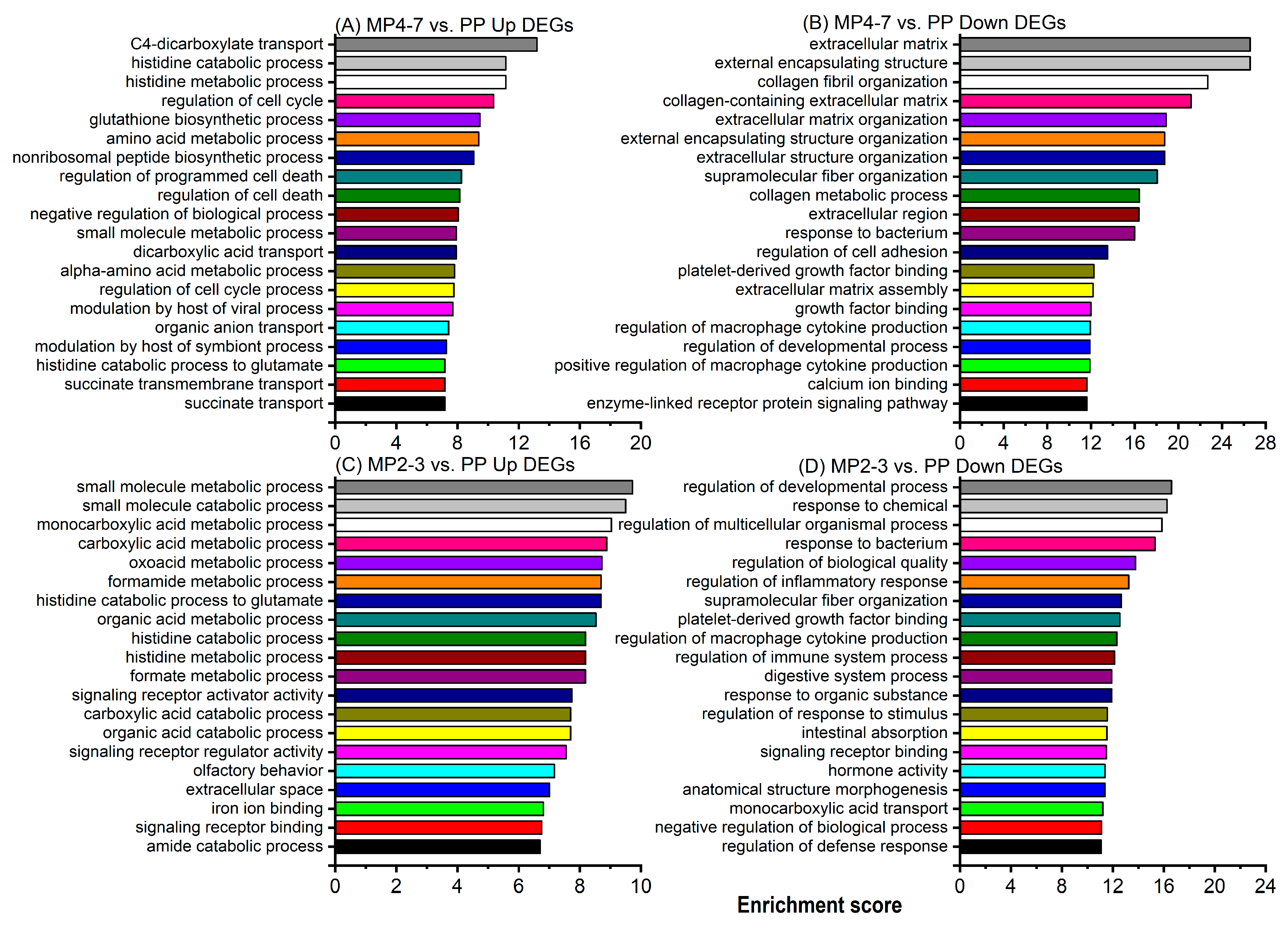
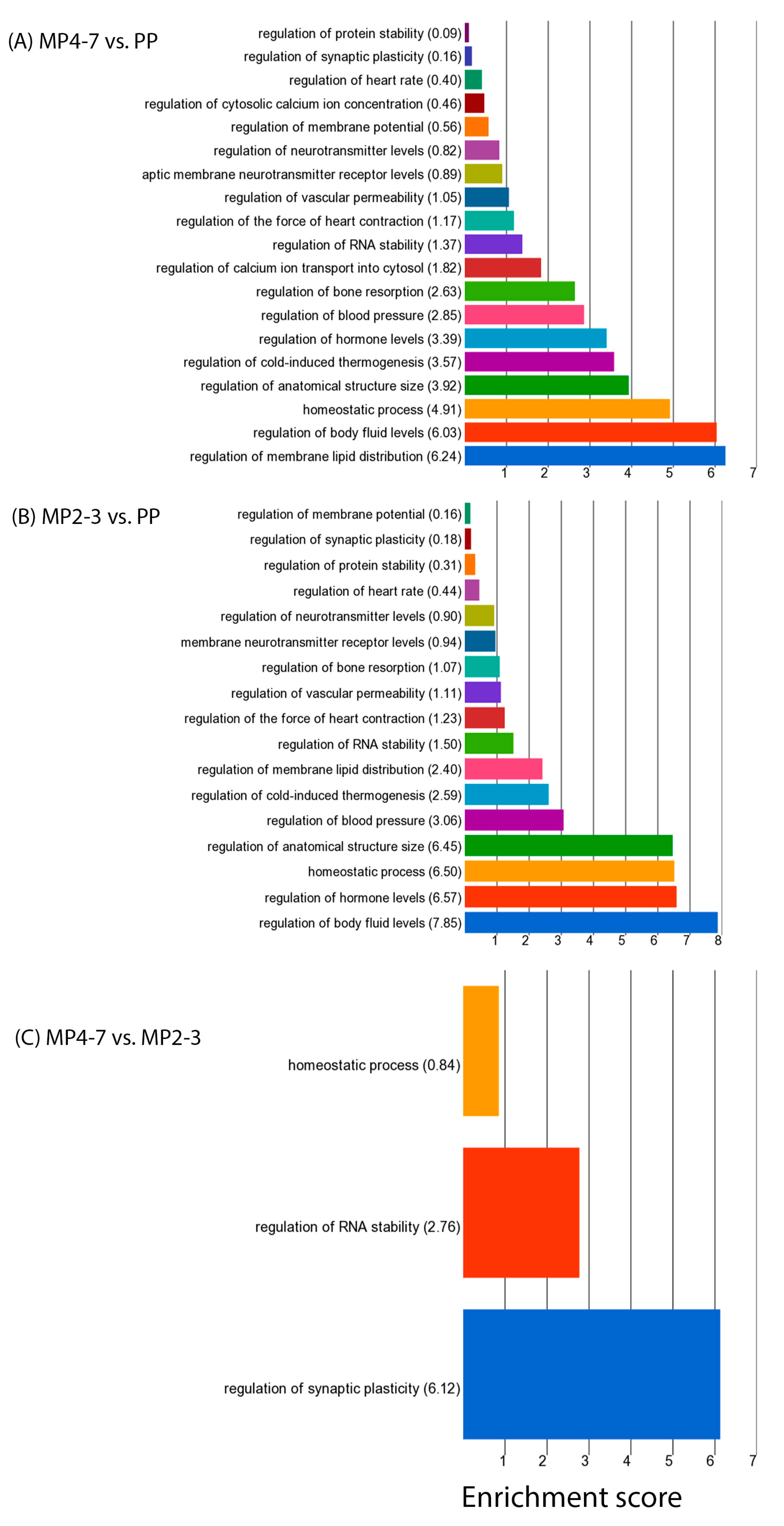
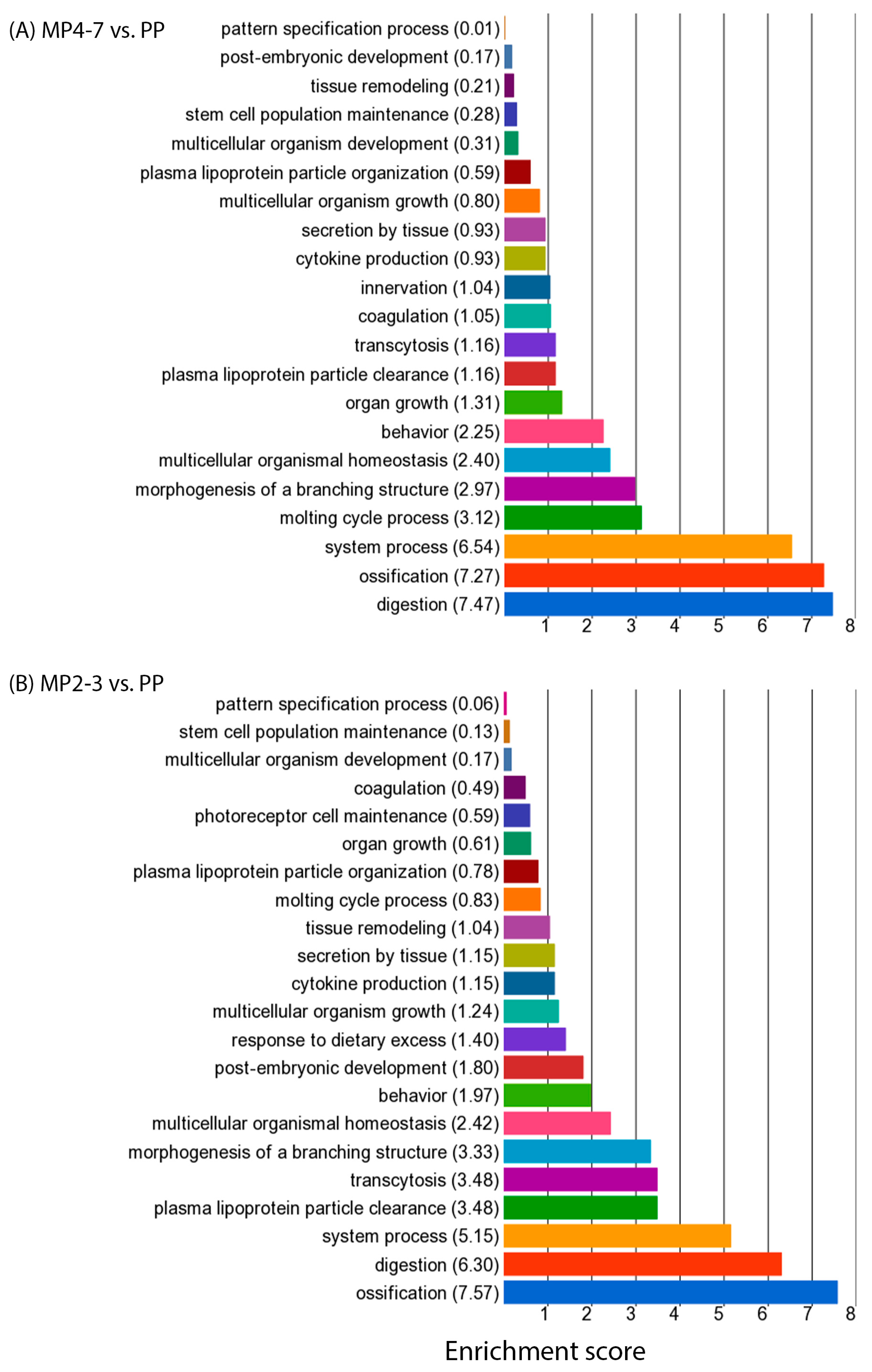
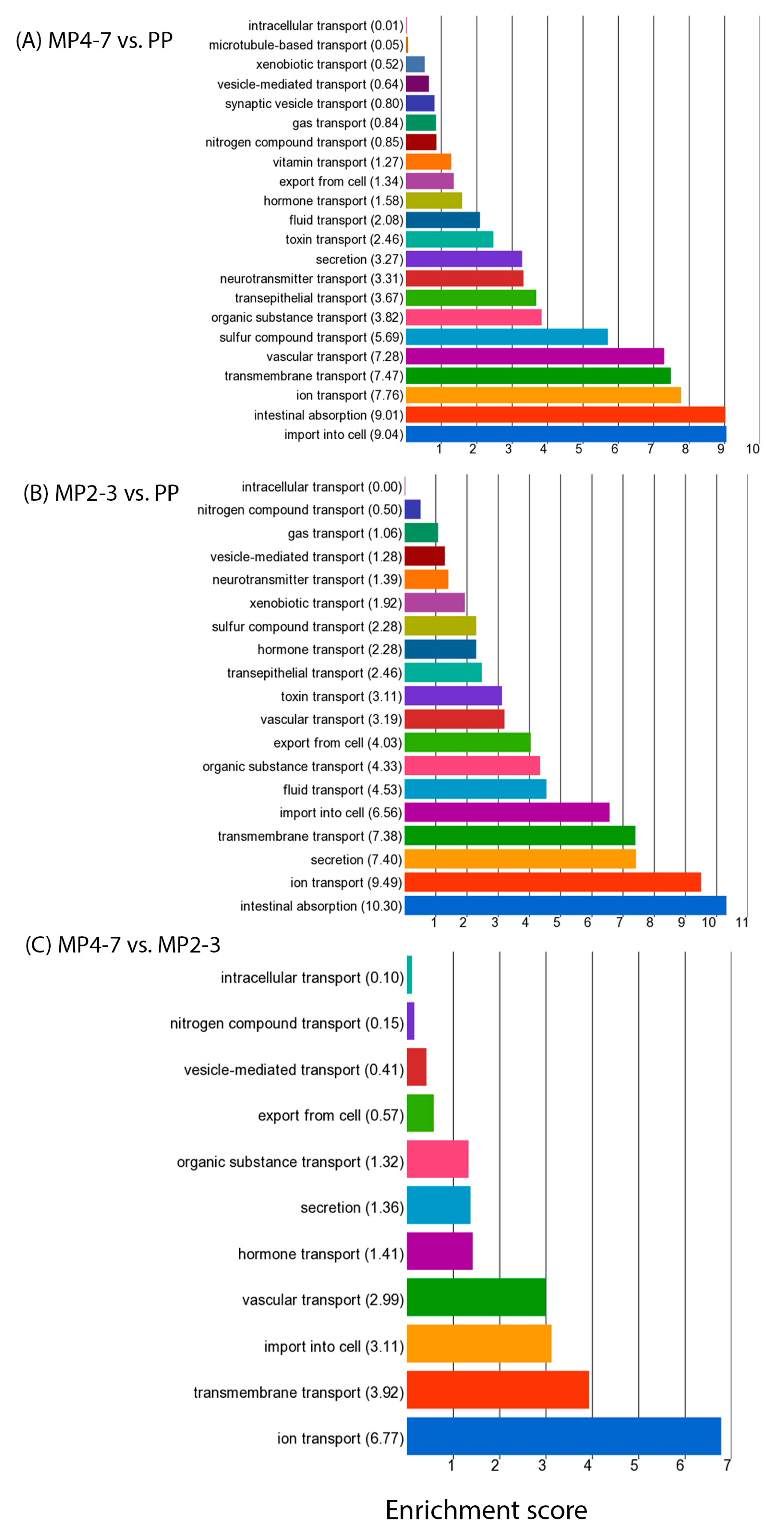
2.3. Comparison of Hepatic Gene Expression between MP Cows with 4–7 Lactations and the PP Cows
2.4. Comparison of Hepatic Gene Expression between the MP Cows with 2–3 Lactations and the PP Cows
2.5. Comparison of Hepatic Gene Expression between the MP Cows with 4–7 and 2–3 Lactations
3. Discussion
3.1. Cow Longevity and Performance
3.2. Mechanisms Underlying the Ageing Process
3.2.1. Digestion and Absorption Processes
3.2.2. Protein Synthesis
3.2.3. Metabolism
3.2.4. Immune Function
3.2.5. Growth Factor Signalling
3.2.6. Responses to DNA Damage, Gene Expression, and mRNA Processing (Transcription and Translation)
3.2.7. Hepatic Morphology
3.2.8. Nutrient Transport
3.2.9. Oestrogen Signalling
4. Materials and Methods
4.1. Animals and Blood Sampling
4.2. Animal Grouping Based on the Lactation Number
4.3. Analysis of Circulating Metabolites and IGF-1
4.4. Cow Phenotype Data Collection
4.5. Liver Biopsy Sampling and RNA Extraction
4.6. RNA-Sequencing, Mapping, and Quantification
4.7. Analysis of Differential Gene Expression between the Dietary Groups
4.8. Gene Ontology Enrichment Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dallago, G.M.; Wade, K.M.; Cue, R.I.; McClure, J.T.; Lacroix, R.; Pellerin, D.; Vasseur, E. Keeping dairy cows for longer: A critical literature review on dairy cow longevity in high milk-producing countries. Animals 2021, 11, 808. [Google Scholar] [CrossRef]
- Boulton, A.C.; Rushton, J.; Wathes, D.C. An empirical analysis of the cost of rearing dairy heifers from birth to first calving and the time taken to repay these costs. Animal 2017, 11, 1372–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.W.; Williams, E.J.; Evans, A.C. A review of the causes of poor fertility in high milk producing dairy cows. Anim. Reprod. Sci. 2011, 123, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, T.; Coffey, M.; Mrode, R.; Wall, E. Understanding the genetics of survival in dairy cows. J. Dairy Sci. 2013, 96, 3296–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, A.; Marcondes, M.I. Review: Overview of factors affecting productive lifespan of dairy cows. Animal 2020, 14, s155–s164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, A. Economic trade-offs between genetic improvement and longevity in dairy cattle. J. Dairy Sci. 2017, 100, 4184–4192. [Google Scholar] [CrossRef]
- Grandl, F.; Furger, M.; Kreuzer, M.; Zehetmeier, M. Impact of longevity on greenhouse gas emissions and profitability of individual dairy cows analysed with different system boundaries. Animal 2019, 13, 198–208. [Google Scholar] [CrossRef]
- Bell, M.J.; Wall, E.; Russell, G.; Roberts, D.J.; Simm, G. Risk factors for culling in Holstein-Friesian dairy cows. Vet. Rec. 2010, 167, 238–240. [Google Scholar] [CrossRef]
- De Vries, A. Symposium review: Why revisit dairy cattle productive lifespan? J. Dairy Sci. 2020, 103, 3838–3845. [Google Scholar] [CrossRef]
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic stress and inflammatory response in high-yielding, periparturient dairy cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef]
- Chagas, L.M.; Bass, J.J.; Blache, D.; Burke, C.R.; Kay, J.K.; Lindsay, D.R.; Lucy, M.C.; Martin, G.B.; Meier, S.; Rhodes, F.M.; et al. Invited review: New perspectives on the roles of nutrition and metabolic priorities in the subfertility of high-producing dairy cows. J. Dairy Sci. 2007, 90, 4022–4032. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Fazio, F.; Levanti, M.; Rizzo, M.; Di Pietro, S.; Giudice, E.; Piccione, G. Lipid and lipoprotein profile changes in dairy cows in response to late pregnancy and the early postpartum period. Arch. Anim. Breed. 2016, 59, 429–434. [Google Scholar] [CrossRef]
- Taylor, V.J.; Cheng, Z.; Pushpakumara, P.G.; Beever, D.E.; Wathes, D.C. Relationships between the plasma concentrations of insulin-like growth factor-I in dairy cows and their fertility and milk yield. Vet. Rec. 2004, 155, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7 (Suppl. S1), 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loor, J.J.; Dann, H.M.; Guretzky, N.A.; Everts, R.E.; Oliveira, R.; Green, C.A.; Litherland, N.B.; Rodriguez-Zas, S.L.; Lewin, H.A.; Drackley, J.K. Plane of nutrition prepartum alters hepatic gene expression and function in dairy cows as assessed by longitudinal transcript and metabolic profiling. Physiol. Genom. 2006, 27, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.; Sundrum, A. Mismatch of glucose allocation between different life functions in the transition period of dairy cows. Animals 2020, 10, 1028. [Google Scholar] [CrossRef]
- Lacasse, P.; Vanacker, N.; Ollier, S.; Ster, C. Innovative dairy cow management to improve resistance to metabolic and infectious diseases during the transition period. Res. Vet. Sci. 2018, 116, 40–46. [Google Scholar] [CrossRef]
- Moyes, K.M.; Larsen, T.; Friggens, N.C.; Drackley, J.K.; Ingvartsen, K.L. Identification of potential markers in blood for the development of subclinical and clinical mastitis in dairy cattle at parturition and during early lactation. J. Dairy Sci. 2009, 92, 5419–5428. [Google Scholar] [CrossRef] [Green Version]
- Kessler, E.C.; Gross, J.J.; Bruckmaier, R.M.; Albrecht, C. Cholesterol metabolism, transport, and hepatic regulation in dairy cows during transition and early lactation. J. Dairy Sci. 2014, 97, 5481–5490. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Wylie, A.; Ferris, C.; Ingvartsen, K.L.; Wathes, D.C.; Gplus, E.C. Effect of diet and nonesterified fatty acid levels on global transcriptomic profiles in circulating peripheral blood mononuclear cells in early lactation dairy cows. J. Dairy Sci. 2021, 104, 10059–10075. [Google Scholar] [CrossRef]
- Wathes, D.C.; Fenwick, M.; Cheng, Z.; Bourne, N.; Llewellyn, S.; Morris, D.G.; Kenny, D.; Murphy, J.; Fitzpatrick, R. Influence of negative energy balance on cyclicity and fertility in the high producing dairy cow. Theriogenology 2007, 68 (Suppl. S1), S232–S241. [Google Scholar] [CrossRef]
- Wathes, D.C.; Cheng, Z.; Chowdhury, W.; Fenwick, M.A.; Fitzpatrick, R.; Morris, D.G.; Patton, J.; Murphy, J.J. Negative energy balance alters global gene expression and immune responses in the uterus of postpartum dairy cows. Physiol. Genom. 2009, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.; Cheng, Z.; Mareiy, W.; Fouladi-Nasht, A. Polyunsaturated fatty acids and fertility in female mammals: An update. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2013, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vels, L.; Rontved, C.M.; Bjerring, M.; Ingvartsen, K.L. Cytokine and acute phase protein gene expression in repeated liver biopsies of dairy cows with a lipopolysaccharide-induced mastitis. J. Dairy Sci. 2009, 92, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, H.H.; Nielsen, J.P.; Heegaard, P.M. Application of acute phase protein measurements in veterinary clinical chemistry. Vet. Res. 2004, 35, 163–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyes, K.M.; Sorensen, P.; Bionaz, M. The Impact of intramammary Escherichia coli challenge on liver and mammary transcriptome and cross-talk in dairy cows during early lactation using RNAseq. PLoS ONE 2016, 11, e0157480. [Google Scholar] [CrossRef] [Green Version]
- Fenwick, M.A.; Fitzpatrick, R.; Kenny, D.A.; Diskin, M.G.; Patton, J.; Murphy, J.J.; Wathes, D.C. Interrelationships between negative energy balance (NEB) and IGF regulation in liver of lactating dairy cows. Domest. Anim. Endocrinol. 2008, 34, 31–44. [Google Scholar] [CrossRef]
- Wathes, D.C.; Becker, F.; Buggiotti, L.; Crowe, M.A.; Ferris, C.; Foldager, L.; Grelet, C.; Hostens, M.; Ingvartsen, K.L.; Marchitelli, C.; et al. Associations between circulating IGF-1 concentrations, disease status and the leukocyte transcriptome in early lactation dairy cows. Ruminants 2021, 1, 147–177. [Google Scholar] [CrossRef]
- Grossmann, M.; Wierman, M.E.; Angus, P.; Handelsman, D.J. Reproductive endocrinology of nonalcoholic fatty liver disease. Endocr. Rev. 2019, 40, 417–446. [Google Scholar] [CrossRef]
- Blankley, S.; Berry, M.P.; Graham, C.M.; Bloom, C.I.; Lipman, M.; O’Garra, A. The application of transcriptional blood signatures to enhance our understanding of the host response to infection: The example of tuberculosis. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2014, 369, 20130427. [Google Scholar] [CrossRef]
- Frenk, S.; Houseley, J. Gene expression hallmarks of cellular ageing. Biogerontology 2018, 19, 547–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.J.; Kang, S.W.S.; Lockwood, G.P.; Le Couteur, D.G.; Cogger, V.C. Hallmarks of aging in the liver. Comput. Struct. Biotechnol. J. 2019, 17, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Buggiotti, L.; Cheng, Z.; Salavati, M.; Wathes, C.D.; Consortium, G.p.E. Comparison of the transcriptome in circulating leukocytes in early lactation between primiparous and multiparous cows provides evidence for age-related changes. BMC Genom. 2021, 22, 693. [Google Scholar] [CrossRef]
- Miglior, F.; Fleming, A.; Malchiodi, F.; Brito, L.F.; Martin, P.; Baes, C.F. A 100-Year Review: Identification and genetic selection of economically important traits in dairy cattle. J. Dairy Sci. 2017, 100, 10251–10271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, M.P.; Hickey, J.; Brotherstone, S. Genetic aspects of growth of Holstein-Friesian dairy cows from birth to maturity. J. Dairy Sci. 2006, 89, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wathes, D.C.; Cheng, Z.; Bourne, N.; Taylor, V.J.; Coffey, M.P.; Brotherstone, S. Differences between primiparous and multiparous dairy cows in the inter-relationships between metabolic traits, milk yield and body condition score in the periparturient period. Domest. Anim. Endocrinol. 2007, 33, 203–225. [Google Scholar] [CrossRef] [Green Version]
- Morales Pineyrua, J.T.; Farina, S.R.; Mendoza, A. Effects of parity on productive, reproductive, metabolic and hormonal responses of Holstein cows. Anim. Reprod. Sci. 2018, 191, 9–21. [Google Scholar] [CrossRef]
- Wiley, C.D.; Campisi, J. The metabolic roots of senescence: Mechanisms and opportunities for intervention. Nat. Metab. 2021, 3, 1290–1301. [Google Scholar] [CrossRef]
- Crispe, I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009, 27, 147–163. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkwood, T.B. Understanding the odd science of aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.W.; Lu, R.; Danthi, P.S.; Bravo, J.I.; Goumba, A.; Sampathkumar, N.K.; Benayoun, B.A. Multi-level remodeling of transcriptional landscapes in aging and longevity. BMB Rep. 2019, 52, 86–108. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, A.A.; Shaposhnikov, M.V.; Zemskaya, N.V.; Koval Lcapital, A.C.; Schegoleva, E.V.; Guvatova, Z.G.; Krasnov, G.S.; Solovev, I.A.; Sheptyakov, M.A.; Zhavoronkov, A.; et al. Transcriptome analysis of long-lived drosophila melanogaster E(z) mutants sheds light on the molecular mechanisms of longevity. Sci. Rep. 2019, 9, 9151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavlakadze, T.; Morris, M.; Fang, J.; Wang, S.X.; Zhu, J.; Zhou, W.; Tse, H.W.; Mondragon-Gonzalez, R.; Roma, G.; Glass, D.J. Age-related Gene expression signature in rats demonstrate early, late, and linear transcriptional changes from multiple tissues. Cell. Rep. 2019, 28, 3263–3273 e3263. [Google Scholar] [CrossRef] [Green Version]
- Tarkhov, A.E.; Alla, R.; Ayyadevara, S.; Pyatnitskiy, M.; Menshikov, L.I.; Shmookler Reis, R.J.; Fedichev, P.O. A universal transcriptomic signature of age reveals the temporal scaling of Caenorhabditis elegans aging trajectories. Sci. Rep. 2019, 9, 7368. [Google Scholar] [CrossRef] [Green Version]
- de Magalhaes, J.P.; Curado, J.; Church, G.M. Meta-analysis of age-related gene expression profiles identifies common signatures of aging. Bioinformatics 2009, 25, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Sebastiani, P.; Montano, M.; Puca, A.; Solovieff, N.; Kojima, T.; Wang, M.C.; Melista, E.; Meltzer, M.; Fischer, S.E.; Andersen, S.; et al. RNA editing genes associated with extreme old age in humans and with lifespan in C. elegans. PLoS ONE 2009, 4, e8210. [Google Scholar] [CrossRef]
- Cieslak, K.P.; Baur, O.; Verheij, J.; Bennink, R.J.; van Gulik, T.M. Liver function declines with increased age. HPB 2016, 18, 691–696. [Google Scholar] [CrossRef] [Green Version]
- Kono, M.; Dreier, J.L.; Ellis, J.M.; Allende, M.L.; Kalkofen, D.N.; Sanders, K.M.; Bielawski, J.; Bielawska, A.; Hannun, Y.A.; Proia, R.L. Neutral ceramidase encoded by the Asah2 gene is essential for the intestinal degradation of sphingolipids. J. Biol. Chem. 2006, 281, 7324–7331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, D.; Fan, C.; Zhou, X.; Liu, Z.; Zheng, B.; Zhu, L.; Jin, Y. Novel compound heterozygous TMPRSS15 gene variants cause enterokinase deficiency. Front. Genet. 2020, 11, 538778. [Google Scholar] [CrossRef] [PubMed]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of lysozyme, an innate immune defense factor, as an alternative antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, R.I.; Cuervo, A.M. Proteostasis and the aging proteome in health and disease. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. S1), S33–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Sojka, D.R.; Gogler-Piglowska, A.; Vydra, N.; Cortez, A.J.; Filipczak, P.T.; Krawczyk, Z.; Scieglinska, D. Functional redundancy of HSPA1, HSPA2 and other HSPA proteins in non-small cell lung carcinoma (NSCLC); an implication for NSCLC treatment. Sci. Rep. 2019, 9, 14394. [Google Scholar] [CrossRef] [Green Version]
- Govindan, J.A.; Jayamani, E.; Ruvkun, G. ROS-based lethality of Caenorhabditis elegans mitochondrial electron transport mutants grown on Escherichia coli siderophore iron release mutants. Proc. Natl. Acad. Sci. USA 2019, 116, 21651–21658. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.T.; Ma, L.; Zhang, Y.D.; Wang, J.Q.; Loor, J.J.; Bu, D.P. Hepatic transcriptome perturbations in dairy cows fed different forage resources. BMC Genom. 2021, 22, 35. [Google Scholar] [CrossRef]
- Magalhaes, S.; Goodfellow, B.J.; Nunes, A. Aging and proteins: What does proteostasis have to do with age? Curr. Mol. Med. 2018, 18, 178–189. [Google Scholar] [CrossRef]
- Hishikawa, D.; Hashidate, T.; Shimizu, T.; Shindou, H. Diversity and function of membrane glycerophospholipids generated by the remodeling pathway in mammalian cells. J. Lipid Res. 2014, 55, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizardo, D.Y.; Lin, Y.L.; Gokcumen, O.; Atilla-Gokcumen, G.E. Regulation of lipids is central to replicative senescence. Mol. Biosyst. 2017, 13, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Hamsanathan, S.; Gurkar, A.U. Lipids as regulators of cellular senescence. Front. Physiol. 2022, 13, 796850. [Google Scholar] [CrossRef] [PubMed]
- McFadden, J.W. Review: Lipid biology in the periparturient dairy cow: Contemporary perspectives. Animal 2020, 14, s165–s175. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.G.; Crookenden, M.A.; Henty, K.M.; Handley, R.R.; Kuhn-Sherlock, B.; White, H.M.; Donkin, S.S.; Snell, R.G.; Meier, S.; Heiser, A.; et al. Epigenetic regulation of pyruvate carboxylase gene expression in the postpartum liver. J. Dairy Sci. 2016, 99, 5820–5827. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, W.M.; Read, M.L.; Gilligan, M.; Bujalska, I.; Docherty, K. Glucose modulates the binding activity of the beta-cell transcription factor IUF1 in a phosphorylation-dependent manner. Biochem. J. 1994, 303, 625–631. [Google Scholar] [CrossRef]
- Peterson, J.M.; Wei, Z.; Wong, G.W. C1q/TNF-related protein-3 (CTRP3), a novel adipokine that regulates hepatic glucose output. J. Biol. Chem. 2010, 285, 39691–39701. [Google Scholar] [CrossRef] [Green Version]
- Weiskirchen, R.; Meurer, S.K.; Liedtke, C.; Huber, M. Mast cells in liver fibrogenesis. Cells 2019, 8, 1429. [Google Scholar] [CrossRef] [Green Version]
- Capelletti, M.M.; Manceau, H.; Puy, H.; Peoc’h, K. Ferroptosis in liver diseases: An Overview. Int. J. Mol. Sci. 2020, 21, 4908. [Google Scholar] [CrossRef]
- Prysyazhnyuk, V.; Sydorchuk, L.; Sydorchuk, R.; Prysiazhniuk, I.; Bobkovych, K.; Buzdugan, I.; Dzuryak, V.; Prysyazhnyuk, P. Glutathione-S-transferases genes-promising predictors of hepatic dysfunction. World J. Hepatol. 2021, 13, 620–633. [Google Scholar] [CrossRef]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Doherty, D.G.; Norris, S.; Madrigal-Estebas, L.; McEntee, G.; Traynor, O.; Hegarty, J.E.; O’Farrelly, C. The human liver contains multiple populations of NK cells, T cells, and CD3+CD56+ natural T cells with distinct cytotoxic activities and Th1, Th2, and Th0 cytokine secretion patterns. J. Immunol. 1999, 163, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- Kenna, T.; Golden-Mason, L.; Norris, S.; Hegarty, J.E.; O’Farrelly, C.; Doherty, D.G. Distinct subpopulations of gamma delta T cells are present in normal and tumor-bearing human liver. Clin. Immunol. 2004, 113, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kenna, T.; Golden-Mason, L.; Porcelli, S.A.; Koezuka, Y.; Hegarty, J.E.; O’Farrelly, C.; Doherty, D.G. NKT cells from normal and tumor-bearing human livers are phenotypically and functionally distinct from murine NKT cells. J. Immunol. 2003, 171, 1775–1779. [Google Scholar] [CrossRef] [Green Version]
- Knolle, P.A.; Wohlleber, D. Immunological functions of liver sinusoidal endothelial cells. Cell. Mol. Immunol. 2016, 13, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell. Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Nikolich-Zugich, J. The twilight of immunity: Emerging concepts in aging of the immune system. Nat. Immunol. 2018, 19, 10–19. [Google Scholar] [CrossRef]
- Salminen, A. Feed-forward regulation between cellular senescence and immunosuppression promotes the aging process and age-related diseases. Ageing Res. Rev. 2021, 67, 101280. [Google Scholar] [CrossRef]
- Loftus, R.M.; Finlay, D.K. Immunometabolism: Cellular metabolism turns immune regulator. J. Biol. Chem. 2016, 291, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Alegre, F.; Pelegrin, P.; Feldstein, A.E. Inflammasomes in Liver Fibrosis. Semin. Liver Dis. 2017, 37, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Uetsuka, K.; Nishikawa, S.; Isobe, K.; Nakayama, H. Histopathological characteristics of Kupffer cells and ito cells in the porcine and bovine liver. J. Vet. Med. Sci. 2007, 69, 767–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, T.S.; Shaffer, A.F.; Currie, J.L.; Isakson, P.C. Role of mast cells in calcium ionophore (A23187)-induced peritoneal inflammation in mice. Inflammation 1994, 18, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Fierbinteanu-Braticevici, C.; Dina, I.; Petrisor, A.; Tribus, L.; Negreanu, L.; Carstoiu, C. Noninvasive investigations for non alcoholic fatty liver disease and liver fibrosis. World J. Gastroenterol. 2010, 16, 4784–4791. [Google Scholar] [CrossRef] [PubMed]
- Kieler, M.; Hofmann, M.; Schabbauer, G. More than just protein building blocks: How amino acids and related metabolic pathways fuel macrophage polarization. FEBS J. 2021, 288, 3694–3714. [Google Scholar] [CrossRef] [PubMed]
- Halaby, M.J.; McGaha, T.L. Amino acid transport and metabolism in myeloid function. Front. Immunol. 2021, 12, 695238. [Google Scholar] [CrossRef]
- Cheng, Z.; Buggiotti, L.; Salavati, M.; Marchitelli, C.; Palma-Vera, S.; Wylie, A.; Takeda, H.; Tang, L.; Crowe, M.A.; Wathes, D.C.; et al. Global transcriptomic profiles of circulating leucocytes in early lactation cows with clinical or subclinical mastitis. Mol. Biol. Rep. 2021, 48, 4611–4623. [Google Scholar] [CrossRef]
- Cheng, Z.; Palma-Vera, S.; Buggiotti, L.; Salavati, M.; Becker, F.; Werling, D.; Wathes, D.C.; Gplus, E.C. Transcriptomic analysis of circulating leukocytes obtained during the recovery from clinical mastitis caused by Escherichia coli in Holstein dairy cows. Animals 2022, 12, 2146. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, S.J. Reproductive tract inflammatory disease in postpartum dairy cows. Animal 2014, 8 (Suppl. S1), 54–63. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef]
- Esslemont, R.J.; Kossaibati, M.A. Culling in 50 dairy herds in England. Vet. Rec. 1997, 140, 36–39. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Boyd, C.K.; Bracken, C.J.; Lamberson, W.R.; Keisler, D.H.; Lucy, M.C. Reduced growth hormone receptor (GHR) messenger ribonucleic acid in liver of periparturient cattle is caused by a specific down-regulation of GHR 1A that is associated with decreased insulin-like growth factor I. Endocrinology 1999, 140, 3947–3954. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K. ADSA foundation scholar award. biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, E.; Franchitto, A.; Pannarale, L.; Carpino, G.; Alpini, G.; Francis, H.; Glaser, S.; Alvaro, D.; Onori, P. Cholangiocytes and blood supply. World J. Gastroenterol. 2006, 12, 3546–3552. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.C.; Farias, J.O.; Forti, F.L. DUSP3 maintains genomic stability and cell proliferation by modulating NER pathway and cell cycle regulatory proteins. Cell. Cycle 2020, 19, 1545–1561. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yue, T.T.; Yang, C.L.; Wang, F.X.; Luo, J.H.; Rong, S.J.; Zhang, M.; Guo, Y.; Xiong, F.; Wang, C.Y. The MAPK dual specific phosphatase (DUSP) proteins: A versatile wrestler in T cell functionality. Int. Immunopharmacol. 2021, 98, 107906. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Jat, P. Mechanisms of cellular senescence: Cell cycle arrest and senescence associated secretory phenotype. Front. Cell. Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The roles of cyclin-dependent kinases in cell-cycle progression and therapeutic strategies in human breast cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef] [Green Version]
- Idda, M.L.; McClusky, W.G.; Lodde, V.; Munk, R.; Abdelmohsen, K.; Rossi, M.; Gorospe, M. Survey of senescent cell markers with age in human tissues. Aging 2020, 12, 4052–4066. [Google Scholar] [CrossRef]
- Gonzalez-Gualda, E.; Baker, A.G.; Fruk, L.; Munoz-Espin, D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2021, 288, 56–80. [Google Scholar] [CrossRef]
- Dai, N. The diverse functions of IMP2/IGF2BP2 in metabolism. Trends Endocrinol. Metab. 2020, 31, 670–679. [Google Scholar] [CrossRef]
- Wang, S.; Jung, Y.; Hyun, J.; Friedersdorf, M.; Oh, S.H.; Kim, J.; Premont, R.T.; Keene, J.D.; Diehl, A.M. RNA binding proteins control transdifferentiation of hepatic stellate cells into myofibroblasts. Cell. Physiol. Biochem. 2018, 48, 1215–1229. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, T.; Wu, D.; Min, Z.; Tan, J.; Yu, B. RNA N6-methyladenosine reader IGF2BP3 regulates cell cycle and angiogenesis in colon cancer. J. Exp. Clin. Cancer Res. 2020, 39, 203. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E.; Arendt, T.; Bennett, M.; Narita, M. Impact of cellular senescence signature on ageing research. Ageing Res. Rev. 2011, 10, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Stahl, E.C.; Haschak, M.J.; Popovic, B.; Brown, B.N. Macrophages in the aging liver and age-related liver disease. Front. Immunol. 2018, 9, 2795. [Google Scholar] [CrossRef] [Green Version]
- Hohn, A.; Grune, T. Lipofuscin: Formation, effects and role of macroautophagy. Redox Biol. 2013, 1, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Kisseleva, T.; Brenner, D.A. Aging and liver disease. Curr. Opin. Gastroenterol. 2015, 31, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Little, M.W.; Ferris, C.; Takeda, H.; Ingvartsen, K.L.; Crowe, M.; Wathes, D.C. Influence of the concentrate inclusion level in a grass silage-based diet on hepatic transcriptomic profiles in Holstein-Friesian dairy cows in early lactation. J. Dairy Sci. 2023, in press. [Google Scholar]
- Ezhilarasan, D. Mitochondria: A critical hub for hepatic stellate cells activation during chronic liver diseases. Hepatobiliary Pancreat. Dis. Int. 2021, 20, 315–322. [Google Scholar] [CrossRef]
- Ortiz, C.; Schierwagen, R.; Schaefer, L.; Klein, S.; Trepat, X.; Trebicka, J. Extracellular matrix remodeling in chronic liver disease. Curr. Tissue Microenviron. Rep. 2021, 2, 41–52. [Google Scholar] [CrossRef]
- Conroy, K.P.; Kitto, L.J.; Henderson, N.C. alphav integrins: Key regulators of tissue fibrosis. Cell. Tissue Res. 2016, 365, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Lomax, M.A.; Baird, G.D. Blood flow and nutrient exchange across the liver and gut of the dairy cow. Effects of lactation and fasting. Br. J. Nutr. 1983, 49, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, C.K.; Aikman, P.C.; Lupoli, B.; Humphries, D.J.; Beever, D.E. Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. J. Dairy Sci. 2003, 86, 1201–1217. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K.; Durst, B.; Lupoli, B.; Humphries, D.J.; Beever, D.E. Visceral tissue mass and rumen volume in dairy cows during the transition from late gestation to early lactation. J. Dairy Sci. 2004, 87, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Lynn, A.K.; Yannas, I.V.; Bonfield, W. Antigenicity and immunogenicity of collagen. J. Biomed. Mater. Res. B Appl. Biomater. 2004, 71, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Barchetta, I.; Cimini, F.A.; Cavallo, M.G. Vitamin D and metabolic dysfunction-associated fatty liver disease (MAFLD): An update. Nutrients 2020, 12, 3302. [Google Scholar] [CrossRef]
- Fiore, E.; Arfuso, F.; Colitti, M.; Gianesella, M.; Giudice, E.; Piccione, G.; Morgante, M. Expression of selected genes related to energy mobilisation and insulin resistance in dairy cows. Anim. Prod. Sci. 2017, 57, 1007–1013. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Vinnakota, R.R. Regulation of intestinal nutrient transport is impaired in aged mice. J. Nutr. 1993, 123, 502–511. [Google Scholar] [CrossRef]
- Bernard, D.; Wiel, C. Transport and senescence. Oncoscience 2015, 2, 741–742. [Google Scholar] [CrossRef] [Green Version]
- Carroll, B.; Korolchuk, V.I. Nutrient sensing, growth and senescence. FEBS J. 2018, 285, 1948–1958. [Google Scholar] [CrossRef] [Green Version]
- Lallet-Daher, H.; Wiel, C.; Gitenay, D.; Navaratnam, N.; Augert, A.; Le Calve, B.; Verbeke, S.; Carling, D.; Aubert, S.; Vindrieux, D.; et al. Potassium channel KCNA1 modulates oncogene-induced senescence and transformation. Cancer Res. 2013, 73, 5253–5265. [Google Scholar] [CrossRef] [Green Version]
- Wiel, C.; Lallet-Daher, H.; Gitenay, D.; Gras, B.; Le Calve, B.; Augert, A.; Ferrand, M.; Prevarskaya, N.; Simonnet, H.; Vindrieux, D.; et al. Endoplasmic reticulum calcium release through ITPR2 channels leads to mitochondrial calcium accumulation and senescence. Nat. Commun. 2014, 5, 3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, M.A.D.; Khadangi, F.; Mersaoui, S.Y.; Naik, D.; Masson, J.Y.; Bilodeau, S. HSP70 mediates a crosstalk between the estrogen and the heat shock response pathways. J. Biol. Chem. 2023, 299, 102872. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of estrogens in the regulation of liverlipid metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [CrossRef] [Green Version]
- Geisler, C.E.; Ghimire, S.; Hepler, C.; Miller, K.E.; Bruggink, S.M.; Kentch, K.P.; Higgins, M.R.; Banek, C.T.; Yoshino, J.; Klein, S.; et al. Hepatocyte membrane potential regulates serum insulin and insulin sensitivity by altering hepatic GABA release. Cell. Rep. 2021, 35, 109298. [Google Scholar] [CrossRef]
- Rezazadeh, H.; Sharifi, M.R.; Sharifi, M.; Soltani, N. Gamma-aminobutyric acid attenuates insulin resistance in type 2 diabetic patients and reduces the risk of insulin resistance in their offspring. Biomed. Pharmacother. 2021, 138, 111440. [Google Scholar] [CrossRef]
- Krogh, M.A.; Hostens, M.; Salavati, M.; Grelet, C.; Sorensen, M.T.; Wathes, D.C.; Ferris, C.P.; Marchitelli, C.; Signorelli, F.; Napolitano, F.; et al. Between- and within-herd variation in blood and milk biomarkers in Holstein cows in early lactation. Animal 2020, 14, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Little, M.W.; Wylie, A.R.G.; O’Connell, N.E.; Welsh, M.D.; Grelet, C.; Bell, M.J.; Gordon, A.; Ferris, C.P. Immunological effects of altering the concentrate inclusion level in a grass silage-based diet for early lactation Holstein Friesian cows. Animal 2019, 13, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Beltman, M.E.; Forde, N.; Furney, P.; Carter, F.; Roche, J.F.; Lonergan, P.; Crowe, M.A. Characterisation of endometrial gene expression and metabolic parameters in beef heifers yielding viable or non-viable embryos on Day 7 after insemination. Reprod. Fertil. Dev. 2010, 22, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmonson, A.J.; Lean, I.; Weaver, L.D.; Farver, T.; Webster, G. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 1989, 72, 18. [Google Scholar] [CrossRef]
- Wathes, D.C.; Cheng, Z.; Salavati, M.; Buggiotti, L.; Takeda, H.; Tang, L.; Becker, F.; Ingvartsen, K.I.; Ferris, C.; Hostens, M.; et al. Relationships between metabolic profiles and gene expression in liver and leukocytes of dairy cows in early lactation. J. Dairy Sci. 2021, 104, 3596–3616. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters 2 | PP Cows | MP2–3 Cows | MP4–7 Cows |
|---|---|---|---|
| N | 41 | 87 | 40 |
| Glucose (mmol/L) | 3.81 ± 0.06 a | 3.44 ± 0.05 b | 3.28 ± 0.07 c |
| Urea (mmol/L) | 3.04 ± 0.14 | 3.10 ± 0.15 | 2.71 ± 0.17 |
| BHB mmol/L) | 0.59 ± 0.07 | 0.70 ± 0.06 | 0.82 ± 0.11 |
| NEFA (mmol/L) | 623.9 ± 74.4 b | 697.7 ± 48.7 b | 881.2 ± 73.1 a |
| IGF1 (ng/mL) | 162.5 ± 13.0 a | 92.3 ± 5.7 b | 61.4 ± 7.0 c |
| Cholesterol (mmol/L) | 2.65 ± 0.08 b | 3.04 ± 0.07 a | 3.02 ± 0.11 a |
| Total DMI (kg/d) | 14.90 ± 0.39 b | 19.20 ± 0.43 a | 19.80 ± 0.67 a |
| BW (kg) | 534.7 ± 6.9 c | 634.5 ± 7.5 b | 686.6 ± 11.4 a |
| BCS | 3.10 ± 0.05 a | 2.83 ± 0.04 b | 2.86 ± 0.08 b |
| MY (kg/d) 3 | 23.2 ± 0.9 c | 35.2 ± 0.7 b | 37.4 ± 0.9 a |
| Milk fat (%) 3 | 4.33 ± 0.13 | 4.37 ± 0.08 | 4.53 ± 0.12 |
| Milk protein (%) 3 | 3.28 ± 0.05 | 3.25 ± 0.03 | 3.23 ± 0.05 |
| ECM (kg/d) 3 | 24.0 ± 0.8 c | 36.0 ± 0.8 b | 39.5 ± 0.9 a |
| EBAL (MJ/d) 3 | −2.6 ± 0.9 a | −5.5 ± 0.8 a | −8.4 ± 1.0 b |
| Functions | Enrichment Score | DEGs in the Function |
|---|---|---|
| Interspecies interaction between organisms | 8.6 | Downregulated DEGs (27): ADIPOQ, ANPEP, CASP1, CCDC80, CHGA, CHMP4A, CUBN, FCER2, GSDME, IFI44, IFI44L, IL22RA1, IRF4, JCHAIN, LEAP2, NLRP6, NOD2, P2RX7, PLA2G1B, PNLIPRP2, PRKCD, PYCARD, REG4, RIPK3, S100A14, SARM1, VIL1 Upregulated DEGs (12): ARG1, ARG2, HAMP, LRRC19, LTF, NOD1, OASL, PC, PRF1, PRLR, PTX3, ZDHHC8 |
| Developmental process | 7.2 | Downregulated DEGs (84): ADA, ADAM19, ADAMTS12, ADAMTS15, ADAMTS2, ADIPOQ, AFF3, ANGPT1, ANPEP, ANXA2, ASB2, C1QTNF3, CA9, CCL11, CCR9, CD109, CDHR2, CDKN1A, CDX2, CHI3L1, CNTNAP1, COL12A1, COL1A1, COL1A2, COL3A1, COL5A1, COL5A2, CRISPLD2, DAPL1, EFEMP1, ELN, EPCAM, FKBP10, FUT1, GAL3ST1, GCNT3, GIP, GLUL, GSDME, INHBA, IRF4, ITGB4, KL, LGR5, LOX, LOXL2, MMP2, MYB, NANOS1, NFATC4, NKX2–3, NPR3, NPY1R, ONECUT1, P2RX7, PDX1, PKDCC, PPARGC1B, PSPH, RFLNB, RHOJ, RIPK3, RNF112, ROS1, RTN4RL1, SEMA3E, SERPINH1, SGCD, SLC27A4, SLC7A11, SLC9A1, SRC, SULF2, TGFA, TGFB3, TGM2, TMEM176B, TNC, TPPP3, VDR, VEGFC, VIL1, WNT5A, ZNF385A Upregulated DEGs (38): ANGPTL4, ARG2, ATF3, ATP6V1B1, BARX2, BHLHA15, CCND1, CDK1, CFTR, CLCF1, CYP1A2, DRGX, DYRK3, ECE2, ECT2, EOMES, GGT1, HSPA2, INSIG1, JPH1, LTF, MEIS1, MFSD2A, MLLT3, NEK2, PCDH19, PRLR, RFX2, SIK1, SLC1A2, SOX9, SP5, SPOCK1, TANC2, TGFB2, TPBG, ZBTB16, ZCWPW1 |
| Multicellular organismal process | 7.0 | Downregulated DEGs (60): ADA, ADAMTS12, ADIPOQ, AHSG, ANGPT1, ASAH2, ASB2, CCL11, CD109, CDX2, CEL, CEMIP, CLDN4, CNTNAP1, COL1A1, COL3A1, CUX2, EDN3, EVC, FKBP10, FMO2, GCLC, GCNT3, GIP, HTR1B, INHBA, JCHAIN, LGR5, LYZ2, LYZ3, MMP2, MMRN1, NFATC4, NOD2, NPR3, NPY1R, OTOG, OTOGL, P2RX7, PI3, PKDCC, PNLIP, PPARGC1B, PROCR, PSPH, SEMA3E, SLC1A4, SLC27A4, SLC5A1, SLC7A11, SRC, SULF2, TGFB3, TGM2, TMPRSS15, TUSC3, VDR, VIL1, WNT5A, ZNF385A Upregulated DEGs (21): ARG2, ATP6V0A4, ATP6V1B1, CFTR, CYP1A2, DRGX, GRID1, HAMP, LTF, MEIS1, MFSD2A, MLLT3, PRLR, PRSS1, SLC1A2, SOX9, TANC2, TGFB2, TPBG, ZBTB16, ZDHHC8 |
| Metabolic process | 6.6 | Downregulated DEGs (133): See Supplementary file Table S3C Upregulated DEGs (72): See Supplementary file Table S3D |
| Biomineralisation | 5.3 | Downregulated DEGs (6): ADA, ANXA2, COL1A1, COL1A2, LOX, PKDCC Upregulated DEGs (1): SOX9 |
| Biological regulation | 4.7 | Downregulated DEGs (195): See Supplementary file Table S3E. Upregulated DEGs (114): See Supplementary file Table S3F. |
| Localisation | 4.7 | Downregulated DEGs (76): See Supplementary file Table S3G. Upregulated DEGs (36): See Supplementary file Table S3H. |
| Pathways | Enrichment FDR p-Value | Number of DEGs |
|---|---|---|
| Protein digestion and absorption | 2.85 × 10−6 | 19 |
| Metabolic pathways | 1.95 × 10−4 | 82 |
| Pancreatic secretion | 6.74 × 10−4 | 13 |
| Glutathione metabolism | 3.38 × 10−3 | 9 |
| Vitamin digestion and absorption | 4.25 × 10−3 | 6 |
| Amoebiasis | 7.01 × 10−3 | 12 |
| Arachidonic acid metabolism | 7.01 × 10−3 | 10 |
| Prostate cancer | 1.60 × 10−2 | 10 |
| Oestrogen signalling pathway | 1.85 × 10−2 | 12 |
| AGE-RAGE signalling pathway in diabetic complications | 1.85 × 10−2 | 10 |
| Ether lipid metabolism | 1.85 × 10−2 | 7 |
| Rheumatoid arthritis | 2.00 × 10−2 | 10 |
| ECM–receptor interaction | 2.00 × 10−2 | 9 |
| Relaxin signalling pathway | 2.59 × 10−2 | 11 |
| Viral protein interaction with cytokine and cytokine receptor | 2.59 × 10−2 | 9 |
| Taurine and hypotaurine metabolism | 2.59 × 10−2 | 4 |
| Arginine and proline metabolism | 3.28 × 10−2 | 6 |
| MAPK signalling pathway | 3.43 × 10−2 | 18 |
| Proteoglycans in cancer | 3.43 × 10−2 | 14 |
| Glycerolipid metabolism | 3.43 × 10−2 | 7 |
| Fat digestion and absorption | 3.43 × 10−2 | 6 |
| AMPK signalling pathway | 3.56 × 10−2 | 10 |
| Biosynthesis of amino acids | 4.64 × 10−2 | 7 |
| PI3K-Akt signalling pathway | 4.91 × 10−2 | 21 |
| Linoleic acid metabolism | 4.91 × 10−2 | 5 |
| Functions | Enrichment Score | DEGs in the Function |
|---|---|---|
| Biomineralisation | 8.6 | Downregulated DEGs (7): ADA, ANKH, COL1A1, COL1A2, LOX, PKDCC, SLC24A3 Upregulated DEGs (1): SPP1 |
| Interspecies interaction between organisms | 7.8 | Downregulated DEGs (26): ADIPOQ, ANPEP, CASP1, CCDC80, CHGA, gzmA, IL22RA1, IRF4, JCHAIN, LCN2, LEAP2, LYPD8, MST1R, NLRP1, NLRP6, NOD2, PLA2G1B, PNLIPRP2, POU2AF1, PRKCD, PYCARD, REG4, RIPK3, SARM1, SLC7A1, VIL1 Upregulated DEGs (5): ARG1, FER1L6, LTF, PRLR, VNN1 |
| Multicellular organismal process | 6.8 | Downregulated DEGs (55): ADA, ADAMTS12, ADIPOQ, AHSG, ANGPT1, ANKS6, AQP3, ASAH2, CCL11, CCND2, CDX2, CEL, CELA1, CLDN4, COL1A1, COL3A1, CUX2, FABP2, FOXS1, GCLC, GIP, HTR1B, INHBA, IQCB1, JCHAIN, LYZ2, LYZ3, MYO1A, NFATC4, NOD2, NPR3, PI3, PIGR, PKDCC, PNLIP, PPARGC1B, PROCR, PSPH, SEMA3E, SGPL1, SLC1A4, SLC5A1, SLC7A11, SRC, STK39, TGFB3, TGM2, TIFAB, TMEM79, TMPRSS15, TUSC3, VDR, VIL1, VLDLR, WNT5A Upregulated DEGs (9): ATP6V1B1, CFTR, CYP1A2, FGF12, LTF, MFSD2A, PRLR, SPP1, TPBG |
| Developmental process | 6.6 | Downregulated DEGs (79): See Supplementary file Table S5B. Upregulated DEGs (16): See Supplementary file Table S5C. |
| Biological regulation | 6.3 | Downregulated DEGs (192): See Supplementary file Table S5D. Upregulated DEGs (52): See Supplementary file Table S5E. |
| Localisation | 5.1 | Downregulated DEGs (77): See Supplementary file Table S5F. Upregulated DEGs (12): See Supplementary file Table S5G. |
| Immune system process | 4.7 | Downregulated DEGs (28): ADA, CCL26, CCR9, CDH17, CHGA, CTSL, DPEP1, ENPP3, FRK, IRAK3, IRF4, JCHAIN, LOC100139916, LOC504295, MCOLN2, NLRP6, NOD2, PLA2G1B, POU2AF1, PRKCD, PSMB8, PSMB9, PYCARD, RFTN1, RIPK3, SARM1, SRC, STK39 Upregulated DEGs (6): CD70, CLCF1, LTF, NLRP1, TNFSF9, VNN1 |
| Locomotion | 4.4 | Downregulated DEGs (10): ANGPT1, ARHGEF16, CCL26, CHGA, DEFB13, DPEP1, LOX, PRKCD, VEGFC, WNT5A Upregulated DEGs (1): TPBG |
| Response to stimulus | 4.1 | Downregulated DEGs (91): See Supplementary file Table S5H. Upregulated DEGs (21): See Supplementary file Table S5I. |
| Pathways | Enrichment FDR p-Value | Number of DEGs |
|---|---|---|
| Protein digestion and absorption | 4.19 × 10−4 | 14 |
| Pancreatic secretion | 4.19 × 10−4 | 12 |
| Metabolic pathways | 1.09 × 10−3 | 66 |
| Relaxin signalling pathway | 1.25 × 10−2 | 11 |
| AMPK signalling pathway | 2.15 × 10−2 | 10 |
| Glycerolipid metabolism | 2.15 × 10−2 | 7 |
| Fat digestion and absorption | 2.15 × 10−2 | 6 |
| Cytokine–cytokine receptor interaction | 2.43 × 10−2 | 18 |
| Ether lipid metabolism | 2.43 × 10−2 | 6 |
| Oestrogen signalling pathway | 2.49 × 10−2 | 10 |
| Viral protein interaction with cytokine and cytokine receptor | 2.49 × 10−2 | 8 |
| Prostate cancer | 2.59 × 10−2 | 8 |
| Linoleic acid metabolism | 2.61 × 10−2 | 5 |
| PI3K-Akt signalling pathway | 2.75 × 10−2 | 19 |
| Metabolism of xenobiotics by cytochrome P450 | 3.79 × 10−2 | 6 |
| Functions | Enrichment Score | DEGs in the Function |
|---|---|---|
| Immune system process | 7.4 | Downregulated DEGs (3): AGER, LGALS2, MCOLN2 Upregulated DEGs (8): BCL6, CASP4, CD247, IFI16, LBP, NOD1, PTX3, TRAT1 |
| Biological regulation | 4.4 | Downregulated DEGs (9): AGER, IGF2BP3, LGALS2, LGR5, MCOLN2, NFATC4, NKD1, WFDC2, ZFPM1 Upregulated DEGs (34): ATF3, BCL6, BHLHA15, CASP4, CCL24, CCL5, CCL8, CCNL1, CD247, CDKN2A, CHAC1, CHRM1, DNAJB1, FBXW5, HOPX, HSPA1A, IFI16, LBP, MECOM, MICAL2, MYOM1, NOD1, PTX3, RAB20, RELL1, RFX2, S100A1, SLCO4A1, SMAP2, SOCS3, SPIDR, STEAP4, TAGLN3, TRAT1 |
| Cellular process | 3.5 | Downregulated DEGs (10): AGER, DNAAF3, DNER, LGALS2, LGR5, MCOLN2, NFATC4, NKD1, SLC15A1, ZFPM1 Upregulated DEGs (38): AEN, ATF3, ATP6V0D2, ATP6V1C2, BCL6, BHLHA15, CASP4, CCL24, CCL5, CCL8, CCNL1, CD247, CDKN2A, CHAC1, CHRM1, DNAJB1, FBXW5, HOPX, HSPA1A, HSPA6, HSPH1, KRBA2, LBP, MICAL2, MYOM1, NOD1, PTX3, RAB20, RFX2, SLC13A5, SLC5A8, SLCO4A1, SOCS3, SPIDR, SRM, STEAP4, TBATA, UCHL1 |
| Interspecies interaction between organisms | 3.2 | Downregulated DEGs (0): Upregulated DEGs (6): CASP4, CCL8, FKBP5, LBP, NOD1, PTX3 |
| Response to stimulus | 3.1 | Downregulated DEGs (2): AGER, NFATC4 Upregulated DEGs (19): AEN, ATF3, BCL6, BHLHA15, CASP4, CHAC1, CHRM1, DNAJB1, FKBP5, HSPA6, LBP, NOD1, PTX3, RAB20, SLC13A5, SOCS3, SPIDR, SRM, TRAT1 |
| Pathways | Enrichment FDR p-Value | Number of DEGs |
|---|---|---|
| Lipid and atherosclerosis | 3.43 × 10−3 | 5 |
| Protein processing in endoplasmic reticulum | 3.43 × 10−3 | 4 |
| Collecting duct acid secretion | 3.43 × 10−3 | 2 |
| Viral protein interaction with cytokine and cytokine receptor | 3.43 × 10−3 | 3 |
| NOD-like receptor signalling pathway | 3.43 × 10−3 | 4 |
| Rheumatoid arthritis | 3.43 × 10−3 | 3 |
| Oestrogen signalling pathway | 5.93 × 10−3 | 3 |
| Legionellosis | 5.93 × 10−3 | 2 |
| Glutathione metabolism | 5.93 × 10−3 | 2 |
| Longevity regulating pathway—multiple species | 5.93 × 10−3 | 2 |
| Wnt signalling pathway | 6.41 × 10−3 | 3 |
| Chronic myeloid leukaemia | 6.41 × 10−3 | 2 |
| Influenza A | 6.41 × 10−3 | 3 |
| Synaptic vesicle cycle | 6.41 × 10−3 | 2 |
| Chemokine signalling pathway | 6.41 × 10−3 | 3 |
| Antigen processing and presentation | 6.88 × 10−3 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Ferris, C.; Crowe, M.A.; Ingvartsen, K.L.; Grelet, C.; Vanlierde, A.; Foldager, L.; Becker, F.; Wathes, D.C.; the GplusE Consortium. Hepatic Global Transcriptomic Profiles of Holstein Cows According to Parity Reveal Age-Related Changes in Early Lactation. Int. J. Mol. Sci. 2023, 24, 9906. https://doi.org/10.3390/ijms24129906
Cheng Z, Ferris C, Crowe MA, Ingvartsen KL, Grelet C, Vanlierde A, Foldager L, Becker F, Wathes DC, the GplusE Consortium. Hepatic Global Transcriptomic Profiles of Holstein Cows According to Parity Reveal Age-Related Changes in Early Lactation. International Journal of Molecular Sciences. 2023; 24(12):9906. https://doi.org/10.3390/ijms24129906
Chicago/Turabian StyleCheng, Zhangrui, Conrad Ferris, Mark A. Crowe, Klaus L. Ingvartsen, Clément Grelet, Amélie Vanlierde, Leslie Foldager, Frank Becker, D. Claire Wathes, and the GplusE Consortium. 2023. "Hepatic Global Transcriptomic Profiles of Holstein Cows According to Parity Reveal Age-Related Changes in Early Lactation" International Journal of Molecular Sciences 24, no. 12: 9906. https://doi.org/10.3390/ijms24129906