
Role of Mast-Cell-Derived RANKL in Ovariectomy-Induced Bone Loss in Mice
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Estrogen Decreased Mast Cell RANKL Secretion
2.2. Rankl Deletion in Mast Cells Did Not Affect Physiological Bone Turnover in Mice
2.3. Rankl Deletion in Mast Cells Did Not Protect from OVX-Induced Bone Loss in Mice
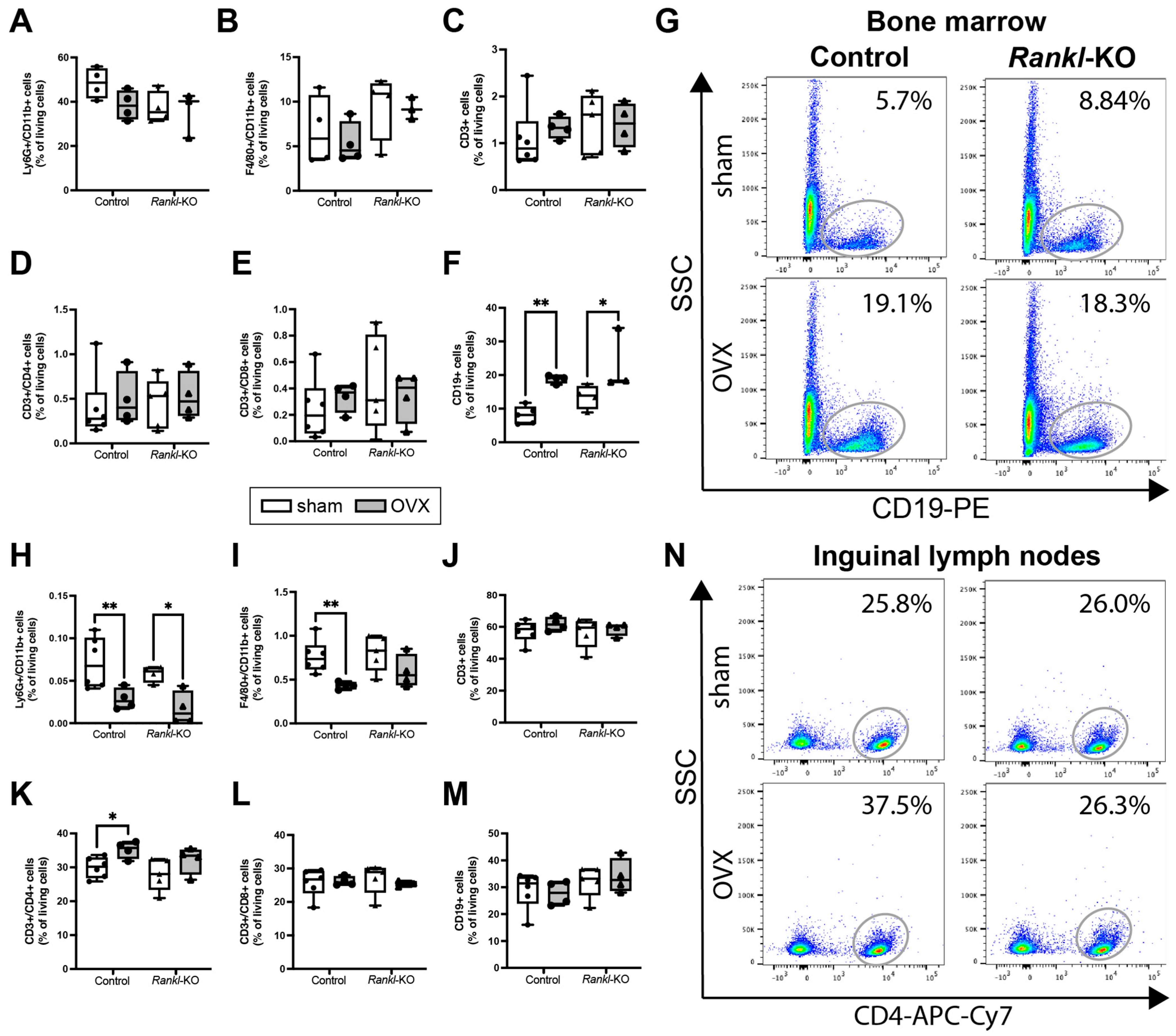
2.4. Rankl Deletion in Mast Cells Only Marginally Affected the Immune Phenotype after OVX
3. Discussion
4. Materials and Methods
4.1. In Vitro Experiments with HMC-1.2 Cells
4.2. In Vivo Experimental Mouse Model
4.3. Bone Phenotyping
4.4. Ovariectomy
4.5. Blood Analyses
4.6. Biomechanical Bone Testing
4.7. Micro-Computed Tomography
4.8. Histomorphometry
4.9. Flow Cytometric Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; Norton, N.; Harvey, N.C.; Jacobson, T.; Johansson, H.; Lorentzon, M.; McCloskey, E.V.; Willers, C.; Borgstrom, F. SCOPE 2021: A new scorecard for osteoporosis in Europe. Arch. Osteoporos. 2021, 16, 82. [Google Scholar] [CrossRef]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Piemontese, M.; Onal, M.; Campbell, J.; Goellner, J.J.; Dusevich, V.; Bonewald, L.; Manolagas, S.C.; O’Brien, C.A. Osteocytes, not Osteoblasts or Lining Cells, are the Main Source of the RANKL Required for Osteoclast Formation in Remodeling Bone. PLoS ONE 2015, 10, e0138189. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Tsuda, E.; Goto, M.; Mochizuki, S.; Yano, K.; Kobayashi, F.; Morinaga, T.; Higashio, K. Isolation of a novel cytokine from human fibroblasts that specifically inhibits osteoclastogenesis. Biochem. Biophys. Res. Commun. 1997, 234, 137–142. [Google Scholar] [CrossRef]
- Eghbali-Fatourechi, G.; Khosla, S.; Sanyal, A.; Boyle, W.J.; Lacey, D.L.; Riggs, B.L. Role of RANK ligand in mediating increased bone resorption in early postmenopausal women. J. Clin. Investig. 2003, 111, 1221–1230. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Piemontese, M.; Liu, Y.; Thostenson, J.D.; Xiong, J.; O’Brien, C.A. RANKL (Receptor Activator of NFkappaB Ligand) Produced by Osteocytes Is Required for the Increase in B Cells and Bone Loss Caused by Estrogen Deficiency in Mice. J. Biol. Chem. 2016, 291, 24838–24850. [Google Scholar] [CrossRef]
- Fischer, V.; Haffner-Luntzer, M. Interaction between bone and immune cells: Implications for postmenopausal osteoporosis. Semin. Cell Dev. Biol. 2022, 123, 14–21. [Google Scholar] [CrossRef]
- Anderson, D.M.; Maraskovsky, E.; Billingsley, W.L.; Dougall, W.C.; Tometsko, M.E.; Roux, E.R.; Teepe, M.C.; DuBose, R.F.; Cosman, D.; Galibert, L. A homologue of the TNF receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature 1997, 390, 175–179. [Google Scholar] [CrossRef]
- Wong, B.R.; Josien, R.; Lee, S.Y.; Sauter, B.; Li, H.L.; Steinman, R.M.; Choi, Y. TRANCE (tumor necrosis factor [TNF]-related activation-induced cytokine), a new TNF family member predominantly expressed in T cells, is a dendritic cell-specific survival factor. J. Exp. Med. 1997, 186, 2075–2080. [Google Scholar] [CrossRef]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef]
- Hayer, S.; Polzer, K.; Brandl, A.; Zwerina, J.; Kireva, T.; Smolen, J.S.; Schett, G. B-cell infiltrates induce endosteal bone formation in inflammatory arthritis. J. Bone Min. Res. 2008, 23, 1650–1660. [Google Scholar] [CrossRef]
- Kanematsu, M.; Sato, T.; Takai, H.; Watanabe, K.; Ikeda, K.; Yamada, Y. Prostaglandin E2 induces expression of receptor activator of nuclear factor-kappa B ligand/osteoprotegrin ligand on pre-B cells: Implications for accelerated osteoclastogenesis in estrogen deficiency. J. Bone Min. Res. 2000, 15, 1321–1329. [Google Scholar] [CrossRef]
- Onal, M.; Xiong, J.; Chen, X.; Thostenson, J.D.; Almeida, M.; Manolagas, S.C.; O’Brien, C.A. Receptor activator of nuclear factor kappaB ligand (RANKL) protein expression by B lymphocytes contributes to ovariectomy-induced bone loss. J. Biol. Chem. 2012, 287, 29851–29860. [Google Scholar] [CrossRef]
- Pacifici, R. Role of T cells in ovariectomy induced bone loss--revisited. J. Bone Min. Res. 2012, 27, 231–239. [Google Scholar] [CrossRef]
- Dahlin, J.S.; Hallgren, J. Mast cell progenitors: Origin, development and migration to tissues. Mol. Immunol. 2015, 63, 9–17. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.S.; Ginhoux, F. New perspectives on the origins and heterogeneity of mast cells. Nat. Rev. Immunol. 2023, 23, 55–68. [Google Scholar] [CrossRef]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2015, 6, 620. [Google Scholar] [CrossRef]
- Fallon, M.D.; Whyte, M.P.; Craig, R.B., Jr.; Teitelbaum, S.L. Mast-cell proliferation in postmenopausal osteoporosis. Calcif. Tissue Int. 1983, 35, 29–31. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.J. Histomorphometric study of mast cells in normal bone, osteoporosis and mastocytosis using a new stain. Calcif. Tissue Int. 1994, 55, 257–259. [Google Scholar] [CrossRef]
- Frame, B.; Nixon, R.K. Bone-marrow mast cells in osteoporosis of aging. N. Engl. J. Med. 1968, 279, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.; Barvencik, F.; Koehne, T.; Priemel, M.; Pogoda, P.; Semler, J.; Minne, H.; Pfeiffer, M.; Zustin, J.; Puschel, K.; et al. Increased osteoblast and osteoclast indices in individuals with systemic mastocytosis. Osteoporos Int. 2013, 24, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Chines, A.; Pacifici, R.; Avioli, L.V.; Teitelbaum, S.L.; Korenblat, P.E. Systemic mastocytosis presenting as osteoporosis: A clinical and histomorphometric study. J. Clin. Endocrinol. Metab. 1991, 72, 140–144. [Google Scholar] [CrossRef]
- Rossini, M.; Zanotti, R.; Bonadonna, P.; Artuso, A.; Caruso, B.; Schena, D.; Vecchiato, D.; Bonifacio, M.; Viapiana, O.; Gatti, D.; et al. Bone mineral density, bone turnover markers and fractures in patients with indolent systemic mastocytosis. Bone 2011, 49, 880–885. [Google Scholar] [CrossRef]
- Lesclous, P.; Saffar, J.L. Mast cells accumulate in rat bone marrow after ovariectomy. Cells Tissues Organs 1999, 164, 23–29. [Google Scholar] [CrossRef]
- Lesclous, P.; Guez, D.; Llorens, A.; Saffar, J.L. Time-course of mast cell accumulation in rat bone marrow after ovariectomy. Calcif. Tissue Int. 2001, 68, 297–303. [Google Scholar] [CrossRef]
- Kroner, J.; Kovtun, A.; Kemmler, J.; Messmann, J.J.; Strauss, G.; Seitz, S.; Schinke, T.; Amling, M.; Kotrba, J.; Froebel, J.; et al. Mast Cells Are Critical Regulators of Bone Fracture-Induced Inflammation and Osteoclast Formation and Activity. J. Bone Min. Res. 2017, 32, 2431–2444. [Google Scholar] [CrossRef]
- Fischer, V.; Ragipoglu, D.; Diedrich, J.; Steppe, L.; Dudeck, A.; Schutze, K.; Kalbitz, M.; Gebhard, F.; Haffner-Luntzer, M.; Ignatius, A. Mast Cells Trigger Disturbed Bone Healing in Osteoporotic Mice. J. Bone Min. Res. 2022, 37, 137–151. [Google Scholar] [CrossRef]
- Ali, A.S.; Lax, A.S.; Liljestrom, M.; Paakkari, I.; Ashammakhi, N.; Kovanen, P.T.; Konttinen, Y.T. Mast cells in atherosclerosis as a source of the cytokine RANKL. Clin. Chem. Lab. Med. 2006, 44, 672–674. [Google Scholar]
- Ng, C.W.; Chan, B.C.L.; Ko, C.H.; Tam, I.Y.S.; Sam, S.W.; Lau, C.B.S.; Leung, P.C.; Lau, H.Y.A. Human mast cells induce osteoclastogenesis through cell surface RANKL. Inflamm. Res. 2022, 71, 1261–1270. [Google Scholar] [CrossRef]
- Kim, K.W.; Kim, B.M.; Won, J.Y.; Min, H.K.; Lee, K.A.; Lee, S.H.; Kim, H.R. Regulation of osteoclastogenesis by mast cell in rheumatoid arthritis. Arthritis Res. 2021, 23, 124. [Google Scholar] [CrossRef]
- Nam, S.Y.; Kim, H.Y.; Min, J.Y.; Kim, H.M.; Jeong, H.J. An osteoclastogenesis system, the RANKL/RANK signalling pathway, contributes to aggravated allergic inflammation. Br. J. Pharm. 2019, 176, 1664–1679. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef]
- Erdei, A.; Andrasfalvy, M.; Peterfy, H.; Toth, G.; Pecht, I. Regulation of mast cell activation by complement-derived peptides. Immunol. Lett. 2004, 92, 39–42. [Google Scholar] [CrossRef]
- El-Lati, S.G.; Dahinden, C.A.; Church, M.K. Complement peptides C3a- and C5a-induced mediator release from dissociated human skin mast cells. J. Investig. Derm. 1994, 102, 803–806. [Google Scholar] [CrossRef]
- Kosa, J.P.; Balla, B.; Kiss, J.; Podani, J.; Takacs, I.; Lazary, A.; Nagy, Z.; Bacsi, K.; Karsai, A.; Speer, G.; et al. Postmenopausal expression changes of immune system-related genes in human bone tissue. J. Clin. Immunol. 2009, 29, 761–768. [Google Scholar] [CrossRef]
- El Khoudary, S.R.; Shields, K.J.; Chen, H.Y.; Matthews, K.A. Menopause, complement, and hemostatic markers in women at midlife: The Study of Women’s Health Across the Nation. Atherosclerosis 2013, 231, 54–58. [Google Scholar] [CrossRef]
- Bulow, J.M.; Renz, N.; Haffner-Luntzer, M.; Fischer, V.; Schoppa, A.; Tuckermann, J.; Kohl, J.; Huber-Lang, M.; Ignatius, A. Complement receptor C5aR1 on osteoblasts regulates osteoclastogenesis in experimental postmenopausal osteoporosis. Front. Endocrinol. 2022, 13, 1016057. [Google Scholar] [CrossRef] [PubMed]
- Bord, S.; Ireland, D.C.; Beavan, S.R.; Compston, J.E. The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 2003, 32, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Streicher, C.; Heyny, A.; Andrukhova, O.; Haigl, B.; Slavic, S.; Schuler, C.; Kollmann, K.; Kantner, I.; Sexl, V.; Kleiter, M.; et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci. Rep. 2017, 7, 6460. [Google Scholar] [CrossRef] [PubMed]
- Cindik, E.D.; Maurer, M.; Hannan, M.K.; Muller, R.; Hayes, W.C.; Hovy, L.; Kurth, A.A. Phenotypical characterization of c-kit receptor deficient mouse femora using non-destructive high-resolution imaging techniques and biomechanical testing. Technol. Health Care 2000, 8, 267–275. [Google Scholar] [CrossRef]
- Katz, H.R.; Austen, K.F. Mast cell deficiency, a game of kit and mouse. Immunity 2011, 35, 668–670. [Google Scholar] [CrossRef]
- Lotinun, S.; Krishnamra, N. Disruption of c-Kit Signaling in Kit(W-sh/W-sh) Growing Mice Increases Bone Turnover. Sci. Rep. 2016, 6, 31515. [Google Scholar] [CrossRef]
- Crotti, T.N.; Smith, M.D.; Weedon, H.; Ahern, M.J.; Findlay, D.M.; Kraan, M.; Tak, P.P.; Haynes, D.R. Receptor activator NF-kappaB ligand (RANKL) expression in synovial tissue from patients with rheumatoid arthritis, spondyloarthropathy, osteoarthritis, and from normal patients: Semiquantitative and quantitative analysis. Ann. Rheum. Dis. 2002, 61, 1047–1054. [Google Scholar] [CrossRef]
- Findlay, D.; Chehade, M.; Tsangari, H.; Neale, S.; Hay, S.; Hopwood, B.; Pannach, S.; O’Loughlin, P.; Fazzalari, N. Circulating RANKL is inversely related to RANKL mRNA levels in bone in osteoarthritic males. Arthritis Res. 2008, 10, R2. [Google Scholar] [CrossRef]
- Vyzoukaki, R.; Tsirakis, G.; Pappa, C.A.; Devetzoglou, M.; Tzardi, M.; Alexandrakis, M.G. The Impact of Mast Cell Density on the Progression of Bone Disease in Multiple Myeloma Patients. Int. Arch. Allergy Immunol. 2015, 168, 263–268. [Google Scholar] [CrossRef]
- Brito, V.G.B.; Patrocinio, M.S.; Sousa, M.C.L.; Barreto, A.E.A.; Frasnelli, S.C.T.; Lara, V.S.; Santos, C.F.; Oliveira, S.H.P. Mast cells contribute to alveolar bone loss in Spontaneously Hypertensive Rats with periodontal disease regulating cytokines production. PLoS ONE 2021, 16, e0247372. [Google Scholar] [CrossRef]
- McElroy, J.F.; Wade, G.N. Short- and long-term effects of ovariectomy on food intake, body weight, carcass composition, and brown adipose tissue in rats. Physiol. Behav. 1987, 39, 361–365. [Google Scholar] [CrossRef]
- Mason, J.B.; Cargill, S.L.; Anderson, G.B.; Carey, J.R. Ovarian status influenced the rate of body-weight change but not the total amount of body-weight gained or lost in female CBA/J mice. Exp. Gerontol. 2010, 45, 435–441. [Google Scholar] [CrossRef]
- Nishio, E.; Hayashi, T.; Nakatani, M.; Aida, N.; Suda, R.; Fujii, T.; Wakatsuki, T.; Honda, S.; Harada, N.; Shimono, Y. Lack of association of ovariectomy-induced obesity with overeating and the reduction of physical activities. Biochem. Biophys. Rep. 2019, 20, 100671. [Google Scholar] [CrossRef]
- Nordqvist, J.; Bernardi, A.; Islander, U.; Carlsten, H. Effects of a tissue-selective estrogen complex on B lymphopoiesis and B cell function. Immunobiology 2017, 222, 918–923. [Google Scholar] [CrossRef]
- Weitzmann, M.N. The Role of Inflammatory Cytokines, the RANKL/OPG Axis, and the Immunoskeletal Interface in Physiological Bone Turnover and Osteoporosis. Science 2013, 2013, 125705. [Google Scholar] [CrossRef]
- Li, Y.; Terauchi, M.; Vikulina, T.; Roser-Page, S.; Weitzmann, M.N. B Cell Production of Both OPG and RANKL is Significantly Increased in Aged Mice. Open Bone J. 2014, 6, 8–17. [Google Scholar]
- Huang, C.; Li, S. Association of blood neutrophil lymphocyte ratio in the patients with postmenopausal osteoporosis. Pak. J. Med. Sci. 2016, 32, 762–765. [Google Scholar] [CrossRef]
- Stubelius, A.; Andersson, A.; Islander, U.; Carlsten, H. Ovarian hormones in innate inflammation. Immunobiology 2017, 222, 878–883. [Google Scholar] [CrossRef]
- Rigoni, T.S.; Vellozo, N.S.; Cabral-Piccin, M.; Fabiano-Coelho, L.; Lopes, U.G.; Filardy, A.A.; DosReis, G.A.; Lopes, M.F. RANK Ligand Helps Immunity to Leishmania major by Skewing M2-Like Into M1 Macrophages. Front. Immunol. 2020, 11, 886. [Google Scholar] [CrossRef]
- Dou, C.; Ding, N.; Zhao, C.; Hou, T.; Kang, F.; Cao, Z.; Liu, C.; Bai, Y.; Dai, Q.; Ma, Q.; et al. Estrogen Deficiency-Mediated M2 Macrophage Osteoclastogenesis Contributes to M1/M2 Ratio Alteration in Ovariectomized Osteoporotic Mice. J. Bone Min. Res. 2018, 33, 899–908. [Google Scholar] [CrossRef]
- Abildgaard, J.; Tingstedt, J.; Zhao, Y.; Hartling, H.J.; Pedersen, A.T.; Lindegaard, B.; Dam Nielsen, S. Increased systemic inflammation and altered distribution of T-cell subsets in postmenopausal women. PLoS ONE 2020, 15, e0235174. [Google Scholar] [CrossRef] [PubMed]
- Haffner-Luntzer, M.; Fischer, V.; Prystaz, K.; Liedert, A.; Ignatius, A. The inflammatory phase of fracture healing is influenced by oestrogen status in mice. Eur. J. Med. Res. 2017, 22, 23. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.S.; Sun, L.J.; Qu, H.L.; Chen, F.; Tian, B.M.; Chen, F.M. The relationship between T-helper cell polarization and the RANKL/OPG ratio in gingival tissues from chronic periodontitis patients. Clin. Exp. Dent. Res. 2019, 5, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Fischer, V.; Kalbitz, M.; Muller-Graf, F.; Gebhard, F.; Ignatius, A.; Liedert, A.; Haffner-Luntzer, M. Influence of Menopause on Inflammatory Cytokines during Murine and Human Bone Fracture Healing. Int. J. Mol. Sci. 2018, 19, 2070. [Google Scholar] [CrossRef]
- Wehrle, E.; Liedert, A.; Heilmann, A.; Wehner, T.; Bindl, R.; Fischer, L.; Haffner-Luntzer, M.; Jakob, F.; Schinke, T.; Amling, M.; et al. The impact of low-magnitude high-frequency vibration on fracture healing is profoundly influenced by the oestrogen status in mice. Dis. Model. Mech. 2015, 8, 93–104. [Google Scholar] [CrossRef]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Min. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef]
- Fischer, V.; Haffner-Luntzer, M.; Prystaz, K.; Vom Scheidt, A.; Busse, B.; Schinke, T.; Amling, M.; Ignatius, A. Calcium and vitamin-D deficiency marginally impairs fracture healing but aggravates posttraumatic bone loss in osteoporotic mice. Sci. Rep. 2017, 7, 7223. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Min. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Rankl-KO |
|---|---|---|
| CtTMD (mgHA/cm3) | 1176 ± 68 | 1230 ± 34 |
| CtTh (mm) | 0.166 ± 0.002 | 0.158 ± 0.006 * |
| TMD (mgHA/cm3) | 612 ± 49 | 645 ± 44 |
| BV/TV (%) | 7.4 ± 2.1 | 8.0 ± 2.3 |
| TbTh (mm) | 0.048 ± 0.001 | 0.046 ± 0.003 |
| TbN (1/mm) | 1.59 ± 0.51 | 1.72 ± 0.44 |
| TbSp (mm) | 0.192 ± 0.042 | 0.184 ± 0.03 |
| NOb/BPm (1/mm) | 13.1 ± 4.0 | 14.4 ± 2.6 |
| ObS/BS (%) | 8.9 ± 2.1 | 9.4 ± 1.2 |
| NOc/BPm (1/mm) | 6.5 ± 1.2 | 5.6 ± 0.5 |
| OcS/BS (%) | 11.6 ± 2.6 | 9.8 ± 1.5 |
| Parameters | Control | Rankl-KO | ||
|---|---|---|---|---|
| Sham | OVX | Sham | OVX | |
| TMD (mgHA/cm3) | 739 ± 21 | 693 ± 14 * | 714 ± 33 | 684 ± 21 |
| BV/TV (%) | 24.5 ± 2.6 | 18.4 ± 1.3 * | 24.7 ± 2.5 | 18.6 ± 1.6 * |
| TbTh (mm) | 0.055 ± 0.003 | 0.050 ± 0.002 * | 0.054 ± 0.004 | 0.050 ± 0.002 * |
| TbN (1/mm) | 4.42 ± 0.33 | 3.69 ± 0.19 * | 4.57 ± 0.43 | 3.75 ± 0.24 * |
| TbSp (mm) | 0.157 ± 0.017 | 0.196 ± 0.024 * | 0.158 ± 0.035 | 0.186 ± 0.035 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, V.; Bülow, J.M.; Krüger, B.T.; Ragipoglu, D.; Vikman, A.; Haffner-Luntzer, M.; Katsoulis-Dimitriou, K.; Dudeck, A.; Ignatius, A. Role of Mast-Cell-Derived RANKL in Ovariectomy-Induced Bone Loss in Mice. Int. J. Mol. Sci. 2023, 24, 9135. https://doi.org/10.3390/ijms24119135
Fischer V, Bülow JM, Krüger BT, Ragipoglu D, Vikman A, Haffner-Luntzer M, Katsoulis-Dimitriou K, Dudeck A, Ignatius A. Role of Mast-Cell-Derived RANKL in Ovariectomy-Induced Bone Loss in Mice. International Journal of Molecular Sciences. 2023; 24(11):9135. https://doi.org/10.3390/ijms24119135
Chicago/Turabian StyleFischer, Verena, Jasmin Maria Bülow, Benjamin Thilo Krüger, Deniz Ragipoglu, Anna Vikman, Melanie Haffner-Luntzer, Konstantinos Katsoulis-Dimitriou, Anne Dudeck, and Anita Ignatius. 2023. "Role of Mast-Cell-Derived RANKL in Ovariectomy-Induced Bone Loss in Mice" International Journal of Molecular Sciences 24, no. 11: 9135. https://doi.org/10.3390/ijms24119135