

Molecular Characterization of Prunus lusitanica L. Fruit Extracts and Their Health-Promoting Potential in Inflammation, Diabetes, and Neurodegenerative Diseases

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction Yields, Phytochemical Characterization, and Antioxidant Activity
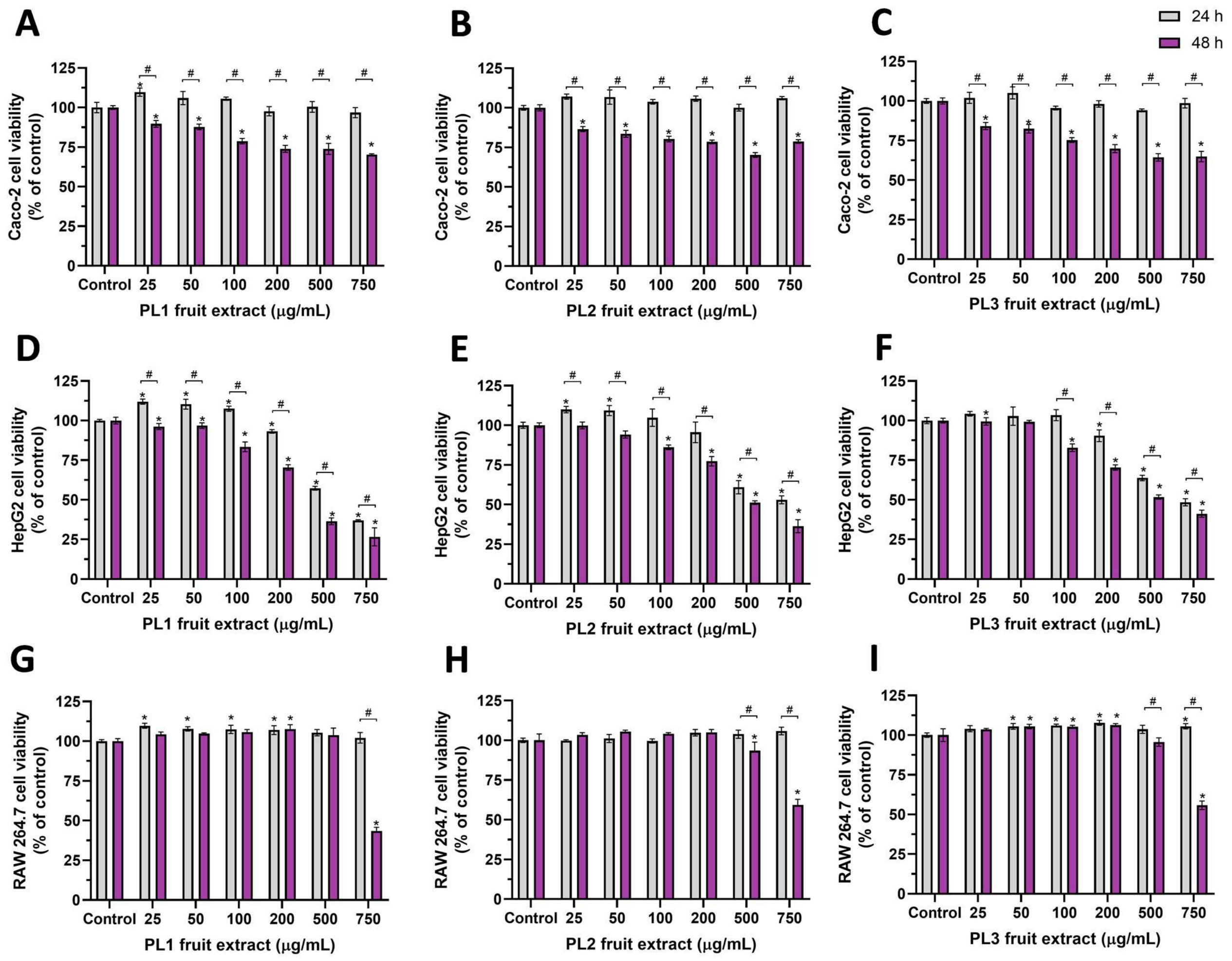
2.2. Antiproliferative/Cytotoxic Activity of P. lusitanica Fruit Extracts
2.3. Anti-Inflammatory Activity of Prunus lusitanica Fruit Extracts
2.4. Assessment of the Inhibitory Effect of P. lusitanica Fruit Extracts on the Activity of Metabolically Relevant Enzymes
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Preparation of Extracts from Prunus lusitanica L. Fruits
3.4. Determination of the Antioxidant Capacity
3.5. HPLC-PAD–ESI-MS/MS Analysis of the Quantitative (Poly)phenolic Profile of Prunus lusitanica L. Fruit Extracts
3.6. In Vitro Cell-Based Assays
3.6.1. Cell Culture Maintenance and Handling
3.6.2. Cell Viability/Cytotoxicity Assay
3.6.3. Assessment of Anti-Inflammatory Activity
3.7. In Vitro Bioassay for the Inhibition of Enzymes Involved in Several Relevant Metabolic Processes
3.7.1. Acetylcholinesterase (AChE) Inhibition Assay
3.7.2. Tyrosinase Inhibition Assay
3.7.3. Elastase Inhibition Assay
3.7.4. α-Amylase Inhibition Assay
3.7.5. α-Glucosidase Inhibition Assay
3.8. Data and Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vekic, J.; Zeljkovic, A.; Stefanovic, A.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V. Obesity and dyslipidemia. Metabolism 2019, 92, 71–81. [Google Scholar] [CrossRef]
- Tabatabaei-Malazy, O.; Larijani, B.; Abdollahi, M. Targeting metabolic disorders by natural products. J. Diabetes Metab. Disord. 2015, 14, 57. [Google Scholar] [CrossRef]
- Martins-Gomes, C.; Souto, E.B.; Silva, A.M. Chapter 15—Nanophytosomes: A novel approach for the delivery of herbal drugs. In Systems of Nanovesicular Drug Delivery; Nayak, A.K., Hasnain, M.S., Aminabhavi, T.M., Torchilin, V.P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 239–257. [Google Scholar]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 8267234. [Google Scholar] [CrossRef]
- Khutami, C.; Sumiwi, S.A.; Khairul Ikram, N.K.; Muchtaridi, M. The Effects of Antioxidants from Natural Products on Obesity, Dyslipidemia, Diabetes and Their Molecular Signaling Mechanism. Int. J. Mol. Sci. 2022, 23, 2056. [Google Scholar] [CrossRef]
- Baek, S.J.; Hammock, B.D.; Hwang, I.-K.; Li, Q.X.; Moustaid-Moussa, N.; Park, Y.; Safe, S.; Suh, N.; Yi, S.-S.; Zeldin, D.C.; et al. Natural Products in the Prevention of Metabolic Diseases: Lessons Learned from the 20th KAST Frontier Scientists Workshop. Nutrients 2021, 13, 1881. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Halpin, C.M.; Reilly, C.; Walsh, J.J. Nature’s Anti-Alzheimer’s Drug: Isolation and Structure Elucidation of Galantamine from Leucojum aestivum. J. Chem. Educ. 2010, 87, 1242–1243. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Yang, Z.; Liu, J.J.; Ueno, H.; Nantermet, P.G.; Guy, R.K.; Claiborne, C.F.; Renaud, J.; Couladouros, E.A.; Paulvannan, K.; et al. Total synthesis of taxol. Nature 1994, 367, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Elfawal, M.A.; Towler, M.J.; Reich, N.G.; Golenbock, D.; Weathers, P.J.; Rich, S.M. Dried whole plant Artemisia annua as an antimalarial therapy. PLoS ONE 2012, 7, e52746. [Google Scholar] [CrossRef]
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural Products for the Treatment of Type 2 Diabetes Mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef]
- Jugran, A.K.; Rawat, S.; Devkota, H.P.; Bhatt, I.D.; Rawal, R.S. Diabetes and plant-derived natural products: From ethnopharmacological approaches to their potential for modern drug discovery and development. Phytother. Res. 2021, 35, 223–245. [Google Scholar] [CrossRef]
- Vlachojannis, J.; Magora, F.; Chrubasik, S. Willow Species and Aspirin: Different Mechanism of Actions. Phytother. Res. 2011, 25, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Martins-Gomes, C.; Nunes, F.M.; Silva, A.M. Modulation of Cell Death Pathways for Cellular Protection and Anti-Tumoral Activity: The Role of Thymus spp. Extracts and Their Bioactive Molecules. Int. J. Mol. Sci. 2023, 24, 1691. [Google Scholar] [CrossRef] [PubMed]
- Gouvinhas, I.; Machado, N.; Sobreira, C.; Domínguez-Perles, R.; Gomes, S.; Rosa, E.; Barros, A.I.R.N.A. Critical Review on the Significance of Olive Phytochemicals in Plant Physiology and Human Health. Molecules 2017, 22, 1986. [Google Scholar] [CrossRef] [PubMed]
- Souto, E.B.; Sampaio, A.C.; Campos, J.R.; Martins-Gomes, C.; Aires, A.; Silva, A.M. Chapter 2—Polyphenols for skin cancer: Chemical properties, structure-related mechanisms of action and new delivery systems. In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 63, pp. 21–42. [Google Scholar]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Sánchez-López, E.; Ettcheto, M.; Egea, M.A.; Espina, M.; Cano, A.; Calpena, A.C.; Camins, A.; Carmona, N.; Silva, A.M.; Souto, E.B.; et al. Memantine loaded PLGA PEGylated nanoparticles for Alzheimer’s disease: In Vitro and in vivo characterization. J. Nanobiotechnology 2018, 16, 32. [Google Scholar] [CrossRef]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepulveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Félix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef] [PubMed]
- Abraão, A.; Yu, M.; Gouvinhas, I.; Ferreira, L.; Silva, A.M.; Domínguez-Perles, R.; Barros, A. Prunus lusitanica L. Fruits: A Promising Underexploited Source of Nutrients with Potential Economic Value. Foods 2023, 12, 973. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural products as alpha-amylase and alpha-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef]
- Pardo, A.; Cáceres, Y.; Pulido, F. Rangewide determinants of population performance in Prunus lusitanica: Lessons for the contemporary conservation of a Tertiary relict tree. Acta Oecologica 2018, 86, 42–48. [Google Scholar] [CrossRef]
- Costa, M.D.C.; Duarte, P.; Neng, N.R.; Nogueira, J.M.F.; Costa, F.; Rosado, C. Novel insights for permeant lead structures through in vitro skin diffusion assays of Prunus lusitanica L., the Portugal Laurel. J. Mol. Struct. 2015, 1079, 327–336. [Google Scholar] [CrossRef]
- Press, B. Green Guide to Trees of Britain and Europe; Bloomsbury Publishing: London, UK, 2016. [Google Scholar]
- Biessels, H.W.A.; van der Kerk-van Hoof, A.C.; Kettenes-van den Bosch, J.J.; Salemink, C.A. Triterpenes of Prunus serotina and P. lusitanica. Phytochemistry 1974, 13, 203–207. [Google Scholar] [CrossRef]
- Abraão, A.S.; Fernandes, N.; Silva, A.M.; Domínguez-Perles, R.; Barros, A. Prunus lusitanica L. Fruits as a Novel Source of Bioactive Compounds with Antioxidant Potential: Exploring the Unknown. Antioxidants 2022, 11, 1738. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Sommella, E.; Santarcangelo, C.; D’Avino, D.; Rossi, A.; Dacrema, M.; Minno, A.D.; Di Matteo, G.; Mannina, L.; Campiglia, P.; et al. Hydroethanolic Extract of Prunus domestica L.: Metabolite Profiling and In Vitro Modulation of Molecular Mechanisms Associated to Cardiometabolic Diseases. Nutrients 2022, 14, 340. [Google Scholar] [CrossRef]
- Holton, T.A.; Cornish, E.C. Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Ertani, A.; Serio, G.; Bertea, C.M. Anthocyanins: Biosynthesis, Distribution, Ecological Role, and Use of Biostimulants to Increase Their Content in Plant Foods—A Review. Agriculture 2021, 11, 212. [Google Scholar] [CrossRef]
- Geng, D.; Shen, X.; Xie, Y.; Yang, Y.; Bian, R.; Gao, Y.; Li, P.; Sun, L.; Feng, H.; Ma, F.; et al. Regulation of phenylpropanoid biosynthesis by MdMYB88 and MdMYB124 contributes to pathogen and drought resistance in apple. Hortic. Res. 2020, 7, 102. [Google Scholar] [CrossRef]
- Macoy, D.M.; Kim, W.-Y.; Lee, S.Y.; Kim, M.G. Biosynthesis, physiology, and functions of hydroxycinnamic acid amides in plants. Plant Biotechnol. Rep. 2015, 9, 269–278. [Google Scholar] [CrossRef]
- Sunil, L.; Shetty, N.P. Biosynthesis and regulation of anthocyanin pathway genes. Appl. Microbiol. Biotechnol. 2022, 106, 1783–1798. [Google Scholar] [CrossRef] [PubMed]
- Hernández, R.; Eguchi, T.; Deveci, M.; Kubota, C. Tomato seedling physiological responses under different percentages of blue and red photon flux ratios using LEDs and cool white fluorescent lamps. Sci. Hortic. 2016, 213, 270–280. [Google Scholar] [CrossRef]
- Costa, R.; Fraga, H.; Fernandes, P.M.; Santos, J.A. Implications of future bioclimatic shifts on Portuguese forests. Reg. Environ. Chang. 2017, 17, 117–127. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Jáuregui, O.; Codina, C.; Tiburcio, A.F.; Bastida, J.; Viladomat, F. Analysis of phenolic compounds by high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry in senescent and water-stressed tobacco. Plant Sci. 2012, 182, 71–78. [Google Scholar] [CrossRef]
- Marčetić, M.; Samardžić, S.; Ilić, T.; Božić, D.D.; Vidović, B. Phenolic Composition, Antioxidant, Anti-Enzymatic, Antimicrobial and Prebiotic Properties of Prunus spinosa L. Fruits. Foods 2022, 11, 3289. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Murakami, T.; Ishiwada, T.; Morikawa, T.; Kagawa, M.; Higashi, Y.; Matsuda, H. New Flavonol Oligoglycosides and Polyacylated Sucroses with Inhibitory Effects on Aldose Reductase and Platelet Aggregation from the Flowers of Prunus mume. J. Nat. Prod. 2002, 65, 1151–1155. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In Vitro Antioxidant versus Metal Ion Chelating Properties of Flavonoids: A Structure-Activity Investigation. PLoS ONE 2016, 11, e0165575. [Google Scholar] [CrossRef]
- Braicu, C.; Pilecki, V.; Balacescu, O.; Irimie, A.; Berindan Neagoe, I. The Relationships Between Biological Activities and Structure of Flavan-3-Ols. Int. J. Mol. Sci. 2011, 12, 9342–9353. [Google Scholar] [CrossRef]
- Silva, A.M.; Martins-Gomes, C.; Fangueiro, J.F.; Andreani, T.; Souto, E.B. Comparison of antiproliferative effect of epigallocatechin gallate when loaded into cationic solid lipid nanoparticles against different cell lines. Pharm. Dev. Technol. 2019, 24, 1243–1249. [Google Scholar] [CrossRef]
- Salamon, I.; Şimşek Sezer, E.N.; Kryvtsova, M.; Labun, P. Antiproliferative and Antimicrobial Activity of Anthocyanins from Berry Fruits after Their Isolation and Freeze-Drying. Appl. Sci. 2021, 11, 2096. [Google Scholar] [CrossRef]
- Geran, R.I.; Greenberg, N.H.; Donald, M.M.M. Protocols for screening chemical agents and natural products against animal tumors and other biological systems. Cancer Chemother. Rep. 1972, 3, 1–103. [Google Scholar]
- Diaconeasa, Z.; Leopold, L.; Rugină, D.; Ayvaz, H.; Socaciu, C. Antiproliferative and Antioxidant Properties of Anthocyanin Rich Extracts from Blueberry and Blackcurrant Juice. Int. J. Mol. Sci. 2015, 16, 2352–2365. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Martins-Gomes, C.; Nunes, F.M.; Silva, A.M. Elderberry (Sambucus nigra L.) extracts promote anti-inflammatory and cellular antioxidant activity. Food Chem. X 2022, 15, 100437. [Google Scholar] [CrossRef]
- Jung, H.; Kwak, H.-K.; Hwang, K.T. Antioxidant and antiinflammatory activities of cyanidin-3-glucoside and cyanidin-3-rutinoside in hydrogen peroxide and lipopolysaccharide-treated RAW264.7 cells. Food Sci. Biotechnol. 2014, 23, 2053–2062. [Google Scholar] [CrossRef]
- Serra, D.; Paixão, J.; Nunes, C.; Dinis, T.C.P.; Almeida, L.M. Cyanidin-3-Glucoside Suppresses Cytokine-Induced Inflammatory Response in Human Intestinal Cells: Comparison with 5-Aminosalicylic Acid. PLoS ONE 2013, 8, e73001. [Google Scholar]
- Moore, K.; Howard, L.; Brownmiller, C.; Gu, I.; Lee, S.-O.; Mauromoustakos, A. Inhibitory effects of cranberry polyphenol and volatile extracts on nitric oxide production in LPS activated RAW 264.7 macrophages. Food Funct. 2019, 10, 7091–7102. [Google Scholar] [CrossRef]
- Grace, M.H.; Esposito, D.; Dunlap, K.L.; Lila, M.A. Comparative Analysis of Phenolic Content and Profile, Antioxidant Capacity, and Anti-inflammatory Bioactivity in Wild Alaskan and Commercial Vaccinium Berries. J. Agric. Food Chem. 2014, 62, 4007–4017. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J.; Liao, H.; Li, C.; Chen, M. Anti-inflammatory effect of lipophilic grape seed proanthocyanidin in RAW 264.7 cells and a zebrafish model. J. Funct. Foods 2020, 75, 104217. [Google Scholar] [CrossRef]
- Sandjo, L.P.; dos Santos Nascimento, M.V.P.; de H. Moraes, M.; Rodrigues, L.M.; Dalmarco, E.M.; Biavatti, M.W.; Steindel, M. NOx-, IL-1β-, TNF-α-, and IL-6-Inhibiting Effects and Trypanocidal Activity of Banana (Musa acuminata) Bracts and Flowers: UPLC-HRESI-MS Detection of Phenylpropanoid Sucrose Esters. Molecules 2019, 24, 4564. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P. Profile of Phenolic Compounds of Prunus armeniaca L. Leaf Extract Determined by LC-ESI-QTOF-MS/MS and Their Antioxidant, Anti-Diabetic, Anti-Cholinesterase, and Anti-Inflammatory Potency. Antioxidants 2021, 10, 1869. [Google Scholar] [CrossRef]
- Lee, J.; Yang, G.; Lee, K.; Lee, M.-H.; Eom, J.-W.; Ham, I.; Choi, H.-Y. Anti-inflammatory effect of Prunus yedoensis through inhibition of nuclear factor-κB in macrophages. BMC Complement. Altern. Med. 2013, 13, 92. [Google Scholar] [CrossRef]
- Nunes, A.R.; Flores-Félix, J.D.; Gonçalves, A.C.; Falcão, A.; Alves, G.; Silva, L.R. Anti-Inflammatory and Antimicrobial Activities of Portuguese Prunus avium L. (Sweet Cherry) By-Products Extracts. Nutrients 2022, 14, 4576. [Google Scholar] [CrossRef]
- Uddin, M.S.; Al Mamun, A.; Kabir, M.T.; Ashraf, G.M.; Bin-Jumah, M.N.; Abdel-Daim, M.M. Multi-Target Drug Candidates for Multifactorial Alzheimer’s Disease: AChE and NMDAR as Molecular Targets. Mol. Neurobiol. 2021, 58, 281–303. [Google Scholar] [CrossRef]
- Hasegawa, T. Tyrosinase-Expressing Neuronal Cell Line as in Vitro Model of Parkinson’s Disease. Int. J. Mol. Sci. 2010, 11, 1082–1089. [Google Scholar] [CrossRef]
- Li, X.; Bai, Y.; Jin, Z.; Svensson, B. Food-derived non-phenolic α-amylase and α-glucosidase inhibitors for controlling starch digestion rate and guiding diabetes-friendly recipes. LWT 2022, 153, 112455. [Google Scholar] [CrossRef]
- Ghosh, S.; More, P.; Derle, A.; Patil, A.B.; Markad, P.; Asok, A.; Kumbhar, N.; Shaikh, M.L.; Ramanamurthy, B.; Shinde, V.S.; et al. Diosgenin from Dioscorea bulbifera: Novel hit for treatment of type II diabetes mellitus with inhibitory activity against α-amylase and α-glucosidase. PLoS ONE 2014, 9, e106039. [Google Scholar]
- Frazão, D.F.; Martins-Gomes, C.; Steck, J.L.; Keller, J.; Delgado, F.; Gonçalves, J.C.; Bunzel, M.; Pintado, C.M.B.S.; Díaz, T.S.; Silva, A.M. Labdanum Resin from Cistus ladanifer L.: A Natural and Sustainable Ingredient for Skin Care Cosmetics with Relevant Cosmeceutical Bioactivities. Plants 2022, 11, 1477. [Google Scholar] [CrossRef]
- Azmi, N.; Hashim, P.; Hashim, D.M.; Halimoon, N.; Majid, N.M.N. Anti–elastase, anti–tyrosinase and matrix metalloproteinase–1 inhibitory activity of earthworm extracts as potential new anti–aging agent. Asian Pac. J. Trop. Biomed. 2014, 4, S348–S352. [Google Scholar] [CrossRef]
- Daroi, P.A.; Dhage, S.N.; Juvekar, A.R. p-Coumaric acid mitigates lipopolysaccharide induced brain damage via alleviating oxidative stress, inflammation and apoptosis. J. Pharm. Pharmacol. 2021, 74, 556–564. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K. Phenolic acids from malt are efficient acetylcholinesterase and butyrylcholinesterase inhibitors. J. Inst. Brew. 2012, 118, 40–48. [Google Scholar] [CrossRef]
- Cásedas, G.; Les, F.; Gómez-Serranillos, M.P.; Smith, C.; López, V. Bioactive and functional properties of sour cherry juice (Prunus cerasus). Food Funct. 2016, 7, 4675–4682. [Google Scholar] [CrossRef]
- Bonesi, M.; Tenuta, M.C.; Loizzo, M.R.; Sicari, V.; Tundis, R. Potential Application of Prunus armeniaca L. and P. domestica L. Leaf Essential Oils as Antioxidant and of Cholinesterases Inhibitors. Antioxidants 2018, 8, 2. [Google Scholar] [CrossRef]
- Park, J.W.; Yuk, H.G.; Lee, S.C. Antioxidant and tyrosinase inhibitory activities of different parts of oriental cherry (Prunus serrulata var. spontanea). Food Sci. Biotechnol. 2012, 21, 339–343. [Google Scholar] [CrossRef]
- Murata, K.; Suzuki, S.; Miyamoto, A.; Horimoto, M.; Nanko, S.; Mori, D.; Kanamaru, H.; Endo, Y. Tyrosinase Inhibitory Activity of Extracts from Prunus persica. Separations 2022, 9, 107. [Google Scholar] [CrossRef]
- Plainfossé, H.; Burger, P.; Verger-Dubois, G.; Azoulay, S.; Fernandez, X. Design Methodology for the Development of a New Cosmetic Active Based on Prunus domestica L. Leaves Extract. Cosmetics 2019, 6, 8. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Silva, P.; Silva, A.M.; Nunes, F.M. Effect of harvesting year and elderberry cultivar on the chemical composition and potential bioactivity: A three-year study. Food Chem. 2020, 302, 125366. [Google Scholar] [CrossRef]
- Stierlin, E.; Azoulay, S.; Massi, L.; Fernandez, X.; Michel, T. Cosmetic potentials of Prunus domestica L. leaves. J. Sci. Food Agric. 2018, 98, 726–736. [Google Scholar] [CrossRef]
- Leal, C.; Gouvinhas, I.; Santos, R.A.; Rosa, E.; Silva, A.M.; Saavedra, M.J.; Barros, A.I.R.N.A. Potential application of grape (Vitis vinifera L.) stem extracts in the cosmetic and pharmaceutical industries: Valorization of a by-product. Ind. Crops Prod. 2020, 154, 112675. [Google Scholar] [CrossRef]
- Silva, A.M.; Félix, L.M.; Teixeira, I.; Martins-Gomes, C.; Schäfer, J.; Souto, E.B.; Santos, D.J.; Bunzel, M.; Nunes, F.M. Orange thyme: Phytochemical profiling, in vitro bioactivities of extracts and potential health benefits. Food Chem. X 2021, 12, 100171. [Google Scholar] [CrossRef] [PubMed]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding phenolic acids inhibition of α-amylase and α-glucosidase and influence of reaction conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative Evaluation of Quercetin, Isoquercetin and Rutin as Inhibitors of α-Glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef] [PubMed]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-anun, S.; Adisakwattana, S. Inhibitory Activities of Cyanidin and Its Glycosides and Synergistic Effect with Acarbose against Intestinal α-Glucosidase and Pancreatic α-Amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef]
- Nan, X.; Jia, W.; Zhang, Y.; Wang, H.; Lin, Z.; Chen, S. An on-line detection system for screening small molecule inhibitors of α-Amylase and α-Glucosidase in Prunus mume. J. Chromatogr. A 2022, 1663, 462754. [Google Scholar] [CrossRef]
- Mendes Lemos, A.; Machado, N.; Egea-Cortines, M.; Barros, A.I. Assessment of quality parameters and phytochemical content of thirty ‘Tempranillo’ grape clones for varietal improvement in two distinct sub-regions of Douro. Sci. Hortic. 2020, 262, 109096. [Google Scholar] [CrossRef]
- Beltrão Martins, R.; Gouvinhas, I.; Nunes, M.C.; Alcides Peres, J.; Raymundo, A.; Barros, A.I.R.N.A. Acorn Flour as a Source of Bioactive Compounds in Gluten-Free Bread. Molecules 2020, 25, 3568. [Google Scholar] [CrossRef]
- Abellán, Á.; Domínguez-Perles, R.; García-Viguera, C.; Moreno, D.A. In Vitro Evidence on Bioaccessibility of Flavonols and Cinnamoyl Derivatives of Cruciferous Sprouts. Nutrients 2021, 13, 4140. [Google Scholar] [CrossRef]
- Silva, A.M.; Martins-Gomes, C.; Souto, E.B.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M. Thymus zygis subsp. zygis an Endemic Portuguese Plant: Phytochemical Profiling, Antioxidant, Anti-Proliferative and Anti-Inflammatory Activities. Antioxidants 2020, 9, 482. [Google Scholar] [CrossRef]
- Andreani, T.; Kiill, C.P.; de Souza, A.L.; Fangueiro, J.F.; Fernandes, L.; Doktorovova, S.; Santos, D.L.; Garcia, M.L.; Gremiao, M.P.; Souto, E.B.; et al. Surface engineering of silica nanoparticles for oral insulin delivery: Characterization and cell toxicity studies. Colloids Surf B Biointerfaces 2014, 123, 916–923. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Martins-Gomes, C.; Coutinho, T.E.; Silva, T.L.; Andreani, T.; Silva, A.M. Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays. Toxics 2022, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Apraj, V.D.; Pandita, N.S. Evaluation of Skin Anti-aging Potential of Citrus reticulata Blanco Peel. Pharmacogn. Res. 2016, 8, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Torres-Naranjo, M.; Suárez, A.; Gilardoni, G.; Cartuche, L.; Flores, P.; Morocho, V. Chemical Constituents of Muehlenbeckia tamnifolia (Kunth) Meisn (Polygonaceae) and Its In Vitro α-Amilase and α-Glucosidase Inhibitory Activities. Molecules 2016, 21, 1461. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Extraction Yield (%, w/w) | Antioxidant Activity (mmol TE/g dw) | |||
|---|---|---|---|---|
| FRAP | DPPH | ABTS | ||
| PL1 | 55.28 ± 1.53 ab | 0.36 ± 0.01 a | 0.36 ± 0.01 b | 0.45 ± 0.01 a |
| PL2 | 59.70 ± 1.12 b | 0.33 ± 0.02 a | 0.32 ± 0.01 a | 0.42 ± 0.02 a |
| PL3 | 54.05 ± 3.12 a | 0.43 ± 0.01 b | 0.40 ± 0.01 c | 0.52 ± 0.02 b |
| R.T. (min) | λ Max (nm) | [M − H]−/[M + H]+ (m/z) | MS n [M – H]−/[M + H]+ (m/z) | Identification | Location | |||
|---|---|---|---|---|---|---|---|---|
| PL1 | PL2 | PL3 | ||||||
| Hydroxycinnamic acids | ||||||||
| 1 | 14.4 | 324 | 353/- | 191,179/- | 3-O-Caffeoylquinic acid | 0.19 ± 0.01 b | 0.15 ± 0.01 a | 0.13 ± 0.00 a |
| 2 | 15.2 | 328 | 503/- | 341,323,179,161,143,135/- | Caffeoyl di-hexoside | 0.67 ± 0.03 c | 0.32 ± 0.02 b | 0.15 ± 0.01 a |
| 3 | 17.3 | 306 | 487/- | 341,307,173,163,145/- | p-Coumaroyl-3-O-sucrose | 1.55 ± 0.09 a | 1.74 ± 0.14 a | 2.26 ± 0.23 b |
| 4 | 18.0 | 310 | 337/- | 191,163, 119/- | 3-p-Coumaroylquinic acid | 1.55 ± 0.14 c | 1.16 ± 0.04 b | 0.61 ± 0.07 a |
| 5 | 18.9 | 312 | 487/- | 341,307,179,163,145/- | Caffeic acid-O-(coumaroyl)hexoside | 4.82 ± 0.41 c | 3.35 ± 0.12 b | 1.64 ± 0.20 a |
| 7 | 21.9 | 326 | 353/- | 191,179,173,155,135,127,111/- | Caffeoyl-isocitrate | 1.70 ± 0.19 b | 2.03 ± 0.25 a | 2.36 ± 0.12 a |
| 10 | 26.4 | 312 | 337/- | 173,163,155,137,127/- | 4-p-Coumaroylquinic acid | 6.28 ± 0.47 c | 3.77 ± 0.38 b | 2.16 ± 0.24 a |
| 11 | 27.0 | 312 | 529/- | 487,349,307,173,162,145/- | Mono-O-acetyl-3-O-p-coumaroylsucrose isomer | 17.97 ± 1.27 a | 19.96 ± 1.82 a | 27.24 ± 2.71 b |
| 12 | 27.4 | 314 | 529/- | 487,349,307,173,162,145/- | Mono-O-acetyl-3-O-p-coumaroylsucrose isomer | 2.35 ± 0.30 b | 2.01 ± 0.14 b | 1.26 ± 0.04 a |
| 13 | 32.4 | 314 | 571/- | 529,511,487,469,393,383,341,307/- | Di-O-acetyl-3-O-p-coumaroyl sucrose isomer | 20.15 ± 1.50 b | 17.85 ± 0.95 b | 13.93 ± 1.03 a |
| 14 | 32.7 | 308 | 571/- | 529,511,487,469,425,383,367,349,341,307,217,173,171,163/- | Di-O-acetyl-3-O-p-coumaroyl sucrose isomer | 1.51 ± 0.20 b | 0.87 ± 0.06 a | 0.74 ± 0.01 a |
| 15 | 33.2 | 314 | 571/- | 529,511,487,469,425,383,367,341,307,289/- | Di-O-acetyl-3-O-p-coumaroyl sucrose isomer | 1.52 ± 0.11 b | 1.41 ± 0.06 b | 1.05 ± 0.08 a |
| 16 | 33.8 | 314 | 613/- | 571,559,553,451,449,425,407,289,273/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 1.05 ± 0.12 b | 0.76 ± 0.11 a | 0.59 ± 0.05 a |
| 17 | 34.9 | 308 | 613/- | 571,553,529,511,425,383,349,289,217,163/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 2.54 ± 0.20 b | 1.26 ± 0.01 a | 1.37 ± 0.15 a |
| 18 | 35.9 | 306 | 571/- | 529,511,487,469,451,422,349,331,307,289,271,259,231,214,173,145/- | Di-O-acetyl-3-O-p-coumaroyl sucrose isomer | 1.23 ± 0.11 a | 1.14 ± 0.11 a | 1.15 ± 0.16 a |
| 19 | 36.2 | 310 | 613/- | 571,553,529,511,487,469,467,425,407,383,349,307,277,228,219,201,163/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 19.65 ± 1.25 b | 16.78 ± 0.66 a | 14.64 ± 1.25 a |
| 22 | 37.6 | 316 | 613/- | 571,553,511,493,469,425,365,349,331,307,289,271,269,245,214,187,163/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 6.39 ± 0.63 b | 5.19 ± 0.01 a | 5.69 ± 0.30 ab |
| 23 | 38.1 | 316 | 613/- | 571,553,511,493,469,451,425,407,391,331,303,287,271,214,197/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 3.39 ± 0.33 b | 2.39 ± 0.14 a | 3.27 ± 0.36 b |
| 24 | 38.2 | 318 | 613/- | 571,553,511,493,451,407,391,389,349,331,318,245,163/- | Tri-O-acetyl-3-O-p-coumaroyl sucrose isomer | 5.78 ± 0.51 b | 4.38 ± 0.38 a | 4.86 ± 0.31 ab |
| 25 | 38.6 | 316 | 655/- | 613,595,553,535,493,391,349,330,313,287,270/- | Tetra-O-acetyl-3-O-p-coumaroylsucrose isomer | 6.81 ± 0.41 b | 3.66 ± 0.07 a | 3.33 ± 0.28 a |
| 26 | 39.3 | 39398, sh334 | 655/- | 613,595,553,535,511,493,393,331/- | Tetra-O-acetyl-3-O-p-coumaroylsucrose isomer | 25.53 ± 0.49 b | 22.17 ± 0.17 a | 23.64 ± 1.19 a |
| Secoiridoids | ||||||||
| 6 | 21.6 | 328 | 581/- | 545,503,341,323,235,161/- | 6-O-β-D-Glucosyl swertiamarin (tentative) | 1.70 ± 0.19 b | 1.46 ± 0.13 ab | 1.25 ± 0.03 a |
| Flavan-3-ols | ||||||||
| 8 | 25.3 | 278 | 289/- | 245,205,203,179,165/- | Catechin | 1.19 ± 0.15 a | 2.93 ± 0.19 c | 2.48 ± 0.09 b |
| 9 | 25.9 | 280 | 865/- | 739,713,695,577,575,451,407,363,289,287,173,163/- | B-type proanthocyanidin trimer | 4.73 ± 0.05a | 8.68 ± 0.58 b | 10.92 ± 0.81 c |
| Flavonols | ||||||||
| 20 | 36.9 | 340 | 463/- | 301,271,255,229,213,193,179,151,121/- | Quercetin-3-O-glucoside | 0.18 ± 0.01 b | 0.11 ± 0.01 a | 0.20 ± 0.02 b |
| 21 | 37.1 | 356 | 609/- | 301,271,255,229,211,193,179,151,121,107/- | Quercetin-3-O-rutinoside | 2.40 ± 0.19 b | 1.19 ± 0.11 a | 2.33 ± 0.08 b |
| Anthocyanins | ||||||||
| 27 | 28.6 | 518 | -/449 | -/366,307,287,227,213,203,187,160 | Cyanidin-3-O-glucoside | 2.00 ± 0.01 b | 0.67 ± 0.01 a | 2.90 ± 0.06 c |
| 28 | 30.4 | 520 | -/595 | -/467,329,287,269,259,252,219,127 | Cyanidin-3-(6-trans-p-coumaroyl)glucoside | 17.08 ± 0.28 b | 10.01 ± 0.27 a | 20.94 ± 2.21 c |
| Total phenolic compounds | 162.29 ± 4.55 b | 137.38 ±1.76 a | 153.11± 8.58 b | |||||
| Total hydroxycinnamic acids | 133.02 ± 4.38 b | 112.34 ± 1.34 a | 112.07 ± 5.47 a | |||||
| Total secoiridoids | 1.70 ± 0.19 b | 1.46 ± 0.13 ab | 1.25 ± 0.03 a | |||||
| Total Flavan-3-ols | 5.92 ± 0.16 a | 11.61 ± 0.42 b | 13.41 ± 0.77 c | |||||
| Total Flavonols | 2.58 ± 0.19 b | 1.30 ± 0.11 a | 2.53 ± 0.10 b | |||||
| Total Anthocyanins | 19.08 ± 0.29 b | 10.68 ± 0.28 a | 23.85 ± 2.27 c | |||||
| IC50 (µg/mL) | ||||
|---|---|---|---|---|
| Caco-2 | HepG2 | RAW 264.7 | ||
| PL1 | 24 h | - | 584.10 ± 25.51 c | - |
| 48 h | - | 352.60 ± 10.00 a | 745.20 ± 36.01 | |
| PL2 | 24 h | - | - | - |
| 48 h | - | 501.10 ± 13.45 b | - | |
| PL3 | 24 h | - | 701.30 ± 26.42 d | - |
| 48 h | - | 517.30 ± 22.66 b | - | |
| Enzymatic Inhibition (% of the Control) | ||||||
|---|---|---|---|---|---|---|
| Extract | Concentration | AChE | Tyrosinase | Elastase | α-Amylase | α-Glucosidase |
| PL1 | 0.5 mg/mL | 28.33 ± 1.04 c | 2.92 ± 0.79 a | n.d. | 1.81 ± 0.16 a | 14.59 ± 1.92 c |
| 1 mg/mL | 39.41 ± 4.18 d | 15.33 ± 3.38 c | n.d. | 2.58 ± 0.29 b | 15.10 ± 0.88 c | |
| PL2 | 0.5 mg/mL | 23.15 ± 1.04 b | 2.00 ± 0.17 a | n.d. | 1.55 ± 0.92 ab | 12.67 ± 1.28 bc |
| 1 mg/mL | 37.19 ± 0.85 d | 11.67 ± 3.6 bc | n.d. | 2.03 ± 0.66 ab | 15.21 ± 0.96 c | |
| PL3 | 0.5 mg/mL | 17.98 ± 1.04 a | 4.75 ± 1.80 a | n.d. | 1.31 ± 0.39 a | 1.98 ± 2.40 a |
| 1 mg/mL | 34.24 ± 3.13 d | 9.48 ± 0.06 b | n.d. | 2.14 ± 0.47 ab | 10.63 ± 0.80 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abraão, A.; Martins-Gomes, C.; Domínguez-Perles, R.; Barros, A.; Silva, A.M. Molecular Characterization of Prunus lusitanica L. Fruit Extracts and Their Health-Promoting Potential in Inflammation, Diabetes, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 8830. https://doi.org/10.3390/ijms24108830
Abraão A, Martins-Gomes C, Domínguez-Perles R, Barros A, Silva AM. Molecular Characterization of Prunus lusitanica L. Fruit Extracts and Their Health-Promoting Potential in Inflammation, Diabetes, and Neurodegenerative Diseases. International Journal of Molecular Sciences. 2023; 24(10):8830. https://doi.org/10.3390/ijms24108830
Chicago/Turabian StyleAbraão, Ana, Carlos Martins-Gomes, Raúl Domínguez-Perles, Ana Barros, and Amélia M. Silva. 2023. "Molecular Characterization of Prunus lusitanica L. Fruit Extracts and Their Health-Promoting Potential in Inflammation, Diabetes, and Neurodegenerative Diseases" International Journal of Molecular Sciences 24, no. 10: 8830. https://doi.org/10.3390/ijms24108830