
Dental Pulp Stem Cells for Salivary Gland Regeneration—Where Are We Today?
 , ,
, ,
Abstract
:1. Introduction
2. Organogenesis of Salivary Glands
3. The Effect of DPSC on Primary Salivary Glands
4. The Immunomodulatory Effect of DPSC
5. Differentiation of DPSCs into Acinar-like Cells
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talha, B.; Swarnkar, S.A. Xerostomia. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Hey, J.; Setz, J.; Gerlach, R.; Janich, M.; Hildebrandt, G.; Vordermark, D.; Gernhardt, C.R.; Kuhnt, T. Parotid gland-recovery after radiotherapy in the head and neck region--36 months follow-up of a prospective clinical study. Radiat. Oncol. 2011, 6, 125. [Google Scholar] [CrossRef]
- Hedström, J.; Tuomi, L.; Finizia, C.; Olsson, C. Identifying organs at risk for radiation-induced late dysphagia in head and neck cancer patients. Clin. Transl. Radiat. Oncol. 2019, 19, 87–95. [Google Scholar] [CrossRef]
- Murdoch-Kinch, C.-A.; Kim, H.M.; Vineberg, K.A.; Ship, J.A.; Eisbruch, A. Dose-effect relationships for the submandibular salivary glands and implications for their sparing by intensity modulated radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2008, 72, 373–382. [Google Scholar] [CrossRef]
- Wu, V.W.C.; Leung, K.Y. A Review on the Assessment of Radiation Induced Salivary Gland Damage after Radiotherapy. Front. Oncol. 2019, 9, 1090. [Google Scholar] [CrossRef] [PubMed]
- Young, W.; Khan, F.; Brandt, R.; Savage, N.; Razek, A.A.; Huang, Q. Syndromes with salivary dysfunction predispose to tooth wear: Case reports of congenital dysfunction of major salivary glands, Prader-Willi, congenital rubella, and Sjögren’s syndromes. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2001, 92, 38–48. [Google Scholar] [CrossRef]
- Dutt, P.; Chaudhary, S.; Kumar, P. Oral health and menopause: A comprehensive review on current knowledge and associated dental management. Ann. Med. Health Sci. Res. 2013, 3, 320–323. [Google Scholar] [CrossRef]
- Kwon, H.-K.; Kim, J.M.; Shin, S.-C.; Sung, E.-S.; Kim, H.-S.; Park, G.C.; Cheon, Y.-I.; Lee, J.-C.; Lee, B.-J. The mechanism of submandibular gland dysfunction after menopause may be associated with the ferroptosis. Aging 2020, 12, 21376–21390. [Google Scholar] [CrossRef]
- Agha-Hosseini, F.; Shirzad, N.; Moosavi, M.-S. Evaluation of Xerostomia and salivary flow rate in Hashimoto’s Thyroiditis. Med. Oral Patol. Oral Cir. Bucal 2016, 21, e1–e5. [Google Scholar] [CrossRef]
- Sroussi, H.Y.; Epstein, J.B.; Bensadoun, R.-J.; Saunders, D.P.; Lalla, R.V.; Migliorati, C.A.; Heaivilin, N.; Zumsteg, Z.S. Common oral complications of head and neck cancer radiation therapy: Mucositis, infections, saliva change, fibrosis, sensory dysfunctions, dental caries, periodontal disease, and osteoradionecrosis. Cancer Med. 2017, 6, 2918–2931. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Pal, M.; Rawat, S.; Grewal, M.S.; Garg, H.; Chauhan, D.; Ahlawat, P.; Tandon, S.; Khurana, R.; Pahuja, A.K.; et al. Radiation-induced dental caries, prevention and treatment—A systematic review. Natl. J. Maxillofac. Surg. 2015, 6, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Connell, C.L.; Abati, S. Diagnosis and management of xerostomia and hyposalivation. Ther. Clin. Risk Manag. 2014, 11, 45–51. [Google Scholar] [CrossRef]
- Tanaka, J.; Ogawa, M.; Hojo, H.; Kawashima, Y.; Mabuchi, Y.; Hata, K.; Nakamura, S.; Yasuhara, R.; Takamatsu, K.; Irié, T.; et al. Generation of orthotopically functional salivary gland from embryonic stem cells. Nat. Commun. 2018, 9, 4216. [Google Scholar] [CrossRef]
- Adine, C.; Ng, K.K.; Rungarunlert, S.; Souza, G.R.; Ferreira, J.N. Engineering innervated secretory epithelial organoids by magnetic three-dimensional bioprinting for stimulating epithelial growth in salivary glands. Biomaterials 2018, 180, 52–66. [Google Scholar] [CrossRef]
- Yan, M.; Kluwe, L.; Friedrich, R.E.; Li, X.; Ren, G.; Smeets, R.; Gosau, M.; Liu, X. Inducing differentiation of human dental pulp cells toward acinar-lineage. Am. J. Transl. Res. 2020, 12, 5781–5788. [Google Scholar] [PubMed]
- Chansaenroj, A.; Yodmuang, S.; Ferreira, J.N. Trends in Salivary Gland Tissue Engineering: From Stem Cells to Secretome and Organoid Bioprinting. Tissue Eng. Part B Rev. 2021, 27, 155–165. [Google Scholar] [CrossRef]
- Zamproni, L.N.; Mundim, M.T.V.V.; Porcionatto, M.A. Neurorepair and Regeneration of the Brain: A Decade of Bioscaffolds and Engineered Microtissue. Front. Cell Dev. Biol. 2021, 9, 764. [Google Scholar] [CrossRef]
- Jacob, G.; Shimomura, K.; Nakamura, N. Osteochondral Injury, Management and Tissue Engineering Approaches. Front. Cell Dev. Biol. 2020, 8, 1118. [Google Scholar] [CrossRef] [PubMed]
- Leal, B.B.J.; Wakabayashi, N.; Oyama, K.; Kamiya, H.; Braghirolli, D.I.; Pranke, P. Vascular Tissue Engineering: Polymers and Methodologies for Small Caliber Vascular Grafts. Front. Cardiovasc. Med. 2021, 7, 376. [Google Scholar] [CrossRef]
- Montero, P.; Flandes-Iparraguirre, M.; Musquiz, S.; Pérez Araluce, M.; Plano, D.; Sanmartín, C.; Orive, G.; Gavira, J.J.; Prosper, F.; Mazo, M.M. Cells, Materials, and Fabrication Processes for Cardiac Tissue Engineering. Front. Bioeng. Biotechnol. 2020, 8, 955. [Google Scholar] [CrossRef] [PubMed]
- Mende, W.; Götzl, R.; Kubo, Y.; Pufe, T.; Ruhl, T.; Beier, J.P. The Role of Adipose Stem Cells in Bone Regeneration and Bone Tissue Engineering. Cells 2021, 10, 975. [Google Scholar] [CrossRef]
- Dai, T.-Q.; Zhang, L.-L.; An, Y.; Xu, F.-F.; An, R.; Xu, H.-Y.; Liu, Y.-P.; Liu, B. In vitro transdifferentiation of adipose tissue-derived stem cells into salivary gland acinar-like cells. Am. J. Transl. Res. 2019, 11, 2908–2924. [Google Scholar] [PubMed]
- Choi, J.-S.; An, H.-Y.; Shin, H.-S.; Kim, Y.-M.; Lim, J.-Y. Enhanced tissue remodelling efficacy of adipose-derived mesenchymal stem cells using injectable matrices in radiation-damaged salivary gland model. J. Tissue Eng. Regen. Med. 2018, 12, e695–e706. [Google Scholar] [CrossRef]
- Saylam, G.; Bayır, Ö.; Gültekin, S.S.; Pınarlı, F.A.; Han, Ü.; Korkmaz, M.H.; Sancaktar, M.E.; Tatar, İ.; Sargon, M.F.; Tatar, E.Ç. Protective/restorative Role of the Adipose Tissue-derived Mesenchymal Stem Cells on the Radioiodine-induced Salivary Gland Damage in Rats. Radiol. Oncol. 2017, 51, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-Y.; Ra, J.C.; Shin, I.S.; Jang, Y.H.; An, H.-Y.; Choi, J.-S.; Kim, W.C.; Kim, Y.-M. Systemic transplantation of human adipose tissue-derived mesenchymal stem cells for the regeneration of irradiation-induced salivary gland damage. PLoS ONE 2013, 8, e71167. [Google Scholar] [CrossRef] [PubMed]
- Bellini, E.; Grieco, M.P.; Raposio, E. A journey through liposuction and liposculture: Review. Ann. Med. Surg. 2017, 24, 53–60. [Google Scholar] [CrossRef]
- Mentz, J.A.; Mentz, H.A.; Nemir, S. Pneumothorax as a Complication of Liposuction. Aesthetic Surg. J. 2020, 40, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, I.E.; Ayoub, R.; Sawaya, R.; Saba, S.C. Iatrogenic sciatic nerve injury during liposuction and fat tissue grafting: A preventable surgical complication with devastating patient outcomes. Patient Saf. Surg. 2020, 14, 40. [Google Scholar] [CrossRef]
- Elsaadany, B.; Zakaria, M.; Mousa, M.R. Transplantation of Bone Marrow-Derived Mesenchymal Stem Cells Preserve the Salivary Glands Structure after Head and Neck Radiation in Rats. Open Access Maced. J. Med. Sci. 2019, 7, 1588–1592. [Google Scholar] [CrossRef]
- AbuBakr, N.; Haggag, T.; Sabry, D.; Salem, Z.A. Functional and histological evaluation of bone marrow stem cell-derived exosomes therapy on the submandibular salivary gland of diabetic Albino rats through TGFβ/Smad3 signaling pathway. Heliyon 2020, 6, e03789. [Google Scholar] [CrossRef]
- Liang, L.; Wang, J.; Zhang, Y.; Shen, Z.; Zheng, J.; Li, J.; Su, Z.; Cai, J.; Jiang, W.; Sun, M. Transdifferentiation of bone marrow-derived mesenchymal stem cells into salivary gland-like cells using a novel culture method. Biotechnol. Lett. 2015, 37, 1505–1513. [Google Scholar] [CrossRef]
- Aghedo, B.O.; Gupta, V. Filgrastim. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Ono, H.; Obana, A.; Usami, Y.; Sakai, M.; Nohara, K.; Egusa, H.; Sakai, T. Regenerating Salivary Glands in the Microenvironment of Induced Pluripotent Stem Cells. BioMed Res. Int. 2015, 2015, 293570. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Janebodin, K.; Horst, O.V.; Ieronimakis, N.; Balasundaram, G.; Reesukumal, K.; Pratumvinit, B.; Reyes, M. Isolation and characterization of neural crest-derived stem cells from dental pulp of neonatal mice. PLoS ONE 2011, 6, e27526. [Google Scholar] [CrossRef]
- Yamazaki, H.; Tsuneto, M.; Yoshino, M.; Yamamura, K.-I.; Hayashi, S.-I. Potential of dental mesenchymal cells in developing teeth. Stem Cells 2007, 25, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Jiang, X.; Ito, Y.; Bringas, P.J.; Han, J.; Rowitch, D.H.; Soriano, P.; McMahon, A.P.; Sucov, H.M. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development 2000, 127, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- American Association of Oral and Maxillofacial Surgeons. Available online: https://www.aaoms.org/images/uploads/pdfs/management_third_molar_supporting_information.pdf (accessed on 10 August 2021).
- Pilbauerová, N.; Suchánek, J. Cryopreservation of Dental Stem Cells. Acta Med. Hradec Kral. 2018, 61, 1–7. [Google Scholar] [CrossRef]
- Liu, L.; Wei, X.; Ling, J.; Wu, L.; Xiao, Y. Expression pattern of Oct-4, Sox2, and c-Myc in the primary culture of human dental pulp derived cells. J. Endod. 2011, 37, 466–472. [Google Scholar] [CrossRef]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef]
- Wells, K.L.; Gaete, M.; Matalova, E.; Deutsch, D.; Rice, D.; Tucker, A.S. Dynamic relationship of the epithelium and mesenchyme during salivary gland initiation: The role of Fgf10. Biol. Open 2013, 2, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Rosa, V.; Dubey, N.; Islam, I.; Min, K.-S.; Nör, J.E. Pluripotency of Stem Cells from Human Exfoliated Deciduous Teeth for Tissue Engineering. Stem Cells Int. 2016, 2016, 5957806. [Google Scholar] [CrossRef]
- Cho, Y.-A.; Noh, K.; Jue, S.-S.; Lee, S.-Y.; Kim, E.-C. Melatonin promotes hepatic differentiation of human dental pulp stem cells: Clinical implications for the prevention of liver fibrosis. J. Pineal Res. 2015, 58, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Mao, J.; Liu, Y. Pulp stem cells derived from human permanent and deciduous teeth: Biological characteristics and therapeutic applications. Stem Cells Transl. Med. 2020, 9, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, L.; Jin, Y.; Shi, S. Fas ligand regulates the immunomodulatory properties of dental pulp stem cells. J. Dent. Res. 2012, 91, 948–954. [Google Scholar] [CrossRef]
- Földes, A.; Kádár, K.; Kerémi, B.; Zsembery, Á.; Gyires, K.; Zádori, Z.S.; Varga, G. Mesenchymal Stem Cells of Dental Origin-Their Potential for Antiinflammatory and Regenerative Actions in Brain and Gut Damage. Curr. Neuropharmacol. 2016, 14, 914–934. [Google Scholar] [CrossRef]
- Lo Monaco, M.; Gervois, P.; Beaumont, J.; Clegg, P.; Bronckaers, A.; Vandeweerd, J.-M.; Lambrichts, I. Therapeutic Potential of Dental Pulp Stem Cells and Leukocyte- and Platelet-Rich Fibrin for Osteoarthritis. Cells 2020, 9, 980. [Google Scholar] [CrossRef]
- Ishikawa, J.; Takahashi, N.; Matsumoto, T.; Yoshioka, Y.; Yamamoto, N.; Nishikawa, M.; Hibi, H.; Ishigro, N.; Ueda, M.; Furukawa, K.; et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental rheumatoid arthritis. Bone 2016, 83, 210–219. [Google Scholar] [CrossRef]
- Racz, G.Z.; Kadar, K.; Foldes, A.; Kallo, K.; Perczel-Kovach, K.; Keremi, B.; Nagy, A.; Varga, G. Immunomodulatory and potential therapeutic role of mesenchymal stem cells in periodontitis. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2014, 65, 327–339. [Google Scholar]
- Al-Zer, H.; Apel, C.; Heiland, M.; Friedrich, R.E.; Jung, O.; Kroeger, N.; Eichhorn, W.; Smeets, R. Enrichment and Schwann Cell Differentiation of Neural Crest-derived Dental Pulp Stem Cells. In Vivo 2015, 29, 319–326. [Google Scholar]
- Patel, V.N.; Hoffman, M.P. Salivary gland development: A template for regeneration. Semin. Cell Dev. Biol. 2014, 25–26, 52–60. [Google Scholar] [CrossRef]
- Morayma Reyes, K.J. Neural Crest-Derived Dental Pulp Stem Cells Function as Ectomesenchyme to Support Salivary Gland Tissue Formation. Dentistry 2013, 13, 2161-1122. [Google Scholar] [CrossRef]
- Gancheva, M.R.; Kremer, K.L.; Gronthos, S.; Koblar, S.A. Using Dental Pulp Stem Cells for Stroke Therapy. Front. Neurol. 2019, 10, 422. [Google Scholar] [CrossRef]
- Yamamura, Y.; Yamada, H.; Sakurai, T.; Ide, F.; Inoue, H.; Muramatsu, T.; Mishima, K.; Hamada, Y.; Saito, I. Treatment of salivary gland hypofunction by transplantation with dental pulp cells. Arch. Oral Biol. 2013, 58, 935–942. [Google Scholar] [CrossRef]
- Narmada, I.B.; Laksono, V.; Nugraha, A.P.; Ernawati, D.S.; Winias, S.; Prahasanti, C.; Dinaryanti, A.; Susilowati, H.; Hendrianto, E.; Ihsan, I.S.; et al. Regeneration of salivary gland defects of diabetic wistar rats post human dental pulp stem cells intraglandular transplantation on acinar cell vacuolization and interleukin-10 serum level. Pesqui. Bras. Odontopediatria Clín. Integr. 2019, 19, e5002. [Google Scholar] [CrossRef]
- Suciadi, S.P.; Nugraha, A.P.; Ernawati, D.S.; Ayuningtyas, N.F.; Narmada, I.B.; Prahasanti, C.; Dinaryanti, A.; Ihsan, I.S.; Hendrinto, E.; Susilowati, H.; et al. The efficacy of human dental pulp stem cells in regenerating submandibular gland defects in diabetic wistar rats (Rattus novergicus). Res. J. Pharm. Technol. 2019, 12, 1573. [Google Scholar] [CrossRef]
- Law, S.; Chaudhuri, S. Mesenchymal stem cell and regenerative medicine: Regeneration versus immunomodulatory challenges. Am. J. Stem Cells 2013, 2, 22–38. [Google Scholar]
- Chen, S.; Cui, G.; Peng, C.; Lavin, M.F.; Sun, X.; Zhang, E.; Yang, Y.; Guan, Y.; Du, Z.; Shao, H. Transplantation of adipose-derived mesenchymal stem cells attenuates pulmonary fibrosis of silicosis via anti-inflammatory and anti-apoptosis effects in rats. Stem Cell Res. Ther. 2018, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Stonesifer, C.; Corey, S.; Ghanekar, S.; Diamandis, Z.; Acosta, S.A.; Borlongan, C.V. Stem cell therapy for abrogating stroke-induced neuroinflammation and relevant secondary cell death mechanisms. Prog. Neurobiol. 2017, 158, 94–131. [Google Scholar] [CrossRef]
- Al-Ghadban, S.; Bunnell, B.A. Adipose Tissue-Derived Stem Cells: Immunomodulatory Effects and Therapeutic Potential. Physiology 2020, 35, 125–133. [Google Scholar] [CrossRef]
- Abdelmawgoud, H.; Saleh, A. Anti-inflammatory and antioxidant effects of mesenchymal and hematopoietic stem cells in a rheumatoid arthritis rat model. Adv. Clin. Exp. Med. 2018, 27, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Luo, H.; Chen, F.; Shi, Y.; Sun, M. Human umbilical cord mesenchymal stem cells polarize RAW264.7 macrophages to an anti-inflammatory subpopulation. Int. J. Clin. Exp. Pathol. 2018, 11, 1446–1452. [Google Scholar] [PubMed]
- Prockop, D.J.; Oh, J.Y. Mesenchymal stem/stromal cells (MSCs): Role as guardians of inflammation. Mol. Ther. 2012, 20, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S.; Pontecorvi, P.; Anastasiadou, E.; Napoli, C.; Marchese, C. Immunomodulatory Effect of Adipose-Derived Stem Cells: The Cutting Edge of Clinical Application. Front. Cell Dev. Biol. 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Verstockt, B.; Ferrante, M.; Vermeire, S.; Van Assche, G. New treatment options for inflammatory bowel diseases. J. Gastroenterol. 2018, 53, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Riordan, N.H.; Hincapié, M.L.; Morales, I.; Fernández, G.; Allen, N.; Leu, C.; Madrigal, M.; Paz Rodríguez, J.; Novarro, N. Allogeneic Human Umbilical Cord Mesenchymal Stem Cells for the Treatment of Autism Spectrum Disorder in Children: Safety Profile and Effect on Cytokine Levels. Stem Cells Transl. Med. 2019, 8, 1008–1016. [Google Scholar] [CrossRef]
- Nagpal, A.; Kremer, K.L.; Hamilton-Bruce, M.A.; Kaidonis, X.; Milton, A.G.; Levi, C.; Shi, S.; Carey, L.; Hillier, S.; Rose, M.; et al. TOOTH (The Open study of dental pulp stem cell Therapy in Humans): Study protocol for evaluating safety and feasibility of autologous human adult dental pulp stem cell therapy in patients with chronic disability after stroke. Int. J. Stroke Off. J. Int. Stroke Soc. 2016, 11, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Cubuk, S.; Oduncuoglu, B.F.; Alaaddinoglu, E.E. The effect of dental pulp stem cells and L-PRF when placed into the extraction sockets of impacted mandibular third molars on the periodontal status of adjacent second molars: A split-mouth, randomized, controlled clinical trial. Oral Maxillofac. Surg. 2023, 27, 59–68. [Google Scholar] [CrossRef]
- Koga, S.; Horiguchi, Y. Efficacy of a cultured conditioned medium of exfoliated deciduous dental pulp stem cells in erectile dysfunction patients. J. Cell. Mol. Med. 2022, 26, 195–201. [Google Scholar] [CrossRef]
- Ferrarotti, F.; Romano, F.; Gamba, M.N.; Quirico, A.; Giraudi, M.; Audagna, M.; Aimetti, M. Human intrabony defect regeneration with micrografts containing dental pulp stem cells: A randomized controlled clinical trial. J. Clin. Periodontol. 2018, 45, 841–850. [Google Scholar] [CrossRef]
- Barbier, L.; Ramos, E.; Mendiola, J.; Rodriguez, O.; Santamaria, G.; Santamaria, J.; Arteagoitia, I. Autologous dental pulp mesenchymal stem cells for inferior third molar post-extraction socket healing: A split-mouth randomised clinical trial. Med. Oral Patol. Oral Cir. Bucal 2018, 23, e469–e477. [Google Scholar] [CrossRef]
- Brizuela, C.; Meza, G.; Urrejola, D.; Quezada, M.A.; Concha, G.; Ramírez, V.; Angelopoulos, I.; Cadiz, M.I.; Tapia-Limonchi, R.; Khoury, M. Cell-Based Regenerative Endodontics for Treatment of Periapical Lesions: A Randomized, Controlled Phase I/II Clinical Trial. J. Dent. Res. 2020, 99, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Matsumura-Kawashima, M.; Moriyama, M.; Kawado, T.; Nakamura, S. Dental pulp-derived stem cell-conditioned media attenuates secondary Sjögren’s syndrome via suppression of inflammatory cytokines in the submandibular glands. Regen. Ther. 2021, 16, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Bao, L.; Gu, Z.; Zhou, Q.; Liang, Y.; Zheng, Y.; Xu, Y.; Zhang, X.; Feng, X. Comparison of immunomodulatory properties of exosomes derived from bone marrow mesenchymal stem cells and dental pulp stem cells. Immunol. Res. 2019, 67, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Matsumura-Kawashima, M.; Ogata, K.; Moriyama, M.; Murakami, Y.; Kawado, T.; Nakamura, S. Secreted factors from dental pulp stem cells improve Sjögren’s syndrome via regulatory T cell-mediated immunosuppression. Stem Cell Res. Ther. 2021, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.H.; Ding, C.; Zhang, Q.; Zhang, Y.; Ge, X.Y.; Li, S.L.; Yu, G.Y. Stem cells from exfoliated deciduous teeth alleviate hyposalivation caused by Sjögren syndrome. Oral Dis. 2019, 25, 1530–1544. [Google Scholar] [CrossRef]
- Kunimatsu, R.; Nakajima, K.; Awada, T.; Tsuka, Y.; Abe, T.; Ando, K.; Hiraki, T.; Kimura, A.; Tanimoto, K. Comparative characterization of stem cells from human exfoliated deciduous teeth, dental pulp, and bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2018, 501, 193–198. [Google Scholar] [CrossRef]
- Nakajima, K.; Kunimatsu, R.; Ando, K.; Hiraki, T.; Rikitake, K.; Tsuka, Y.; Abe, T.; Tanimoto, K. Success rates in isolating mesenchymal stem cells from permanent and deciduous teeth. Sci. Rep. 2019, 9, 16764. [Google Scholar] [CrossRef]
- Al-Serwi, R.H.; El-Kersh, A.O.F.O.; El-Akabawy, G. Human dental pulp stem cells attenuate streptozotocin-induced parotid gland injury in rats. Stem Cell Res. Ther. 2021, 12, 577. [Google Scholar] [CrossRef]
- Takeuchi, H.; Takahashi, H.; Tanaka, A. Effects of Human Dental Pulp Stem Cell-Derived Conditioned Medium on Atrophied Submandibular Gland after the Release from Ligation of the Main Excretory Duct in Mice. J. Hard Tissue Biol. 2020, 29, 183–192. [Google Scholar] [CrossRef]
- le Roux, J.; Kleynhans, J.; Rubow, S. The use of HEPES-buffer in the production of gallium-68 radiopharmaceuticals–time to reconsider strict pharmacopoeial limits? EJNMMI Radiopharm. Chem. 2021, 6, 15. [Google Scholar] [CrossRef]
- de Silva, R.; Dasanayake, W.M.D.K.; Wickramasinhe, G.D.; Karunatilake, C.; Weerasinghe, N.; Gunasekera, P.; Malavige, G.N. Sensitization to bovine serum albumin as a possible cause of allergic reactions to vaccines. Vaccine 2017, 35, 1494–1500. [Google Scholar] [CrossRef]
- Baquir, B.; Hancock, R.E.W. Exosomes, your body’s answer to immune health. Ann. Transl. Med. 2017, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuang, S.; Zhang, Y.; Yang, M.; Qin, W.; Shi, X.; Lin, Z. Chitosan hydrogel incorporated with dental pulp stem cell-derived exosomes alleviates periodontitis in mice via a macrophage-dependent mechanism. Bioact. Mater. 2020, 5, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Sakai, K.; Koma, Y.; Watanabe, J.; Liu, K.; Maruyama, H.; Sakaguchi, K.; Hibi, H. Dental pulp stem cell-derived small extracellular vesicle in irradiation-induced senescence. Biochem. Biophys. Res. Commun. 2021, 575, 28–35. [Google Scholar] [CrossRef]
- Akashi, Y.; Nemoto, A.; Nakajima, K.; Kokubun, K.; Murakami, S.; Inoue, T.; Matsuzaka, K. The effect of fibroblast growth factor 7 on human dental pulp stem cells for differentiation to AQP5-positive and αSMA-positive cells in vitro and in vivo. Clin. Exp. Dent. Res. 2021, 7, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Morsczeck, C.; Reichert, T.E. Dental stem cells in tooth regeneration and repair in the future. Expert Opin. Biol. Ther. 2018, 18, 187–196. [Google Scholar] [CrossRef]
- Croci, S.; Bonacini, M.; Dolci, G.; Massari, M.; Facciolongo, N.; Pignatti, E.; Pisciotta, A.; Carnevale, G.; Negro, A.; Cassone, G.; et al. Human Dental Pulp Stem Cells Modulate Cytokine Production in vitro by Peripheral Blood Mononuclear Cells from Coronavirus Disease 2019 Patients. Front. Cell Dev. Biol. 2020, 8, 609204. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Z.; Wang, Z. Dental Pulp Stem Cells Ameliorate Elastase-Induced Pulmonary Emphysema by Regulating Inflammation and Oxidative Stress. J. Inflamm. Res. 2023, 16, 1497–1508. [Google Scholar] [CrossRef]
- Tang, X.; Li, W.; Wen, X.; Zhang, Z.; Chen, W.; Yao, G.; Chen, H.; Wang, D.; Shi, S.; Sun, L. Transplantation of dental tissue-derived mesenchymal stem cells ameliorates nephritis in lupus mice. Ann. Transl. Med. 2019, 7, 132. [Google Scholar] [CrossRef]
- Inada, R.; Mendoza, H.Y.; Tanaka, T.; Horie, T.; Satomi, T. Preclinical study for the treatment of diabetes mellitus using β-like cells derived from human dental pulp stem cells. Regen. Med. 2022, 17, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, N.; Liu, X.; Su, J.; Cong, X.; Wu, L.; Zhang, Y.; Yu, G. Autonomic reinnervation and functional regeneration in autologous transplanted submandibular glands in patients with severe keratoconjunctivitis sicca. Int. J. Oral Sci. 2018, 10, 14. [Google Scholar] [CrossRef]
- Scheller, E.L.; Krebsbach, P.H.; Kohn, D.H. Tissue engineering: State of the art in oral rehabilitation. J. Oral Rehabil. 2009, 36, 368–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-S.; Feng, Z.-H.; Wu, G.-F.; Bai, S.-Z.; Dong, Y.; Chen, F.-M.; Zhao, Y.-M. The use of platelet-rich fibrin combined with periodontal ligament and jaw bone mesenchymal stem cell sheets for periodontal tissue engineering. Sci. Rep. 2016, 6, 28126. [Google Scholar] [CrossRef] [PubMed]
- Duin, S.; Schütz, K.; Ahlfeld, T.; Lehmann, S.; Lode, A.; Ludwig, B.; Gelinsky, M. 3D Bioprinting of Functional Islets of Langerhans in an Alginate/Methylcellulose Hydrogel Blend. Adv. Healthc. Mater. 2019, 8, e1801631. [Google Scholar] [CrossRef]
- Ahlfeld, T.; Köhler, T.; Czichy, C.; Lode, A.; Gelinsky, M. A Methylcellulose Hydrogel as Support for 3D Plotting of Complex Shaped Calcium Phosphate Scaffolds. Gels 2018, 4, 68. [Google Scholar] [CrossRef]
- Kopp, A.; Smeets, R.; Gosau, M.; Friedrich, R.E.; Fuest, S.; Behbahani, M.; Barbeck, M.; Rutkowski, R.; Burg, S.; Kluwe, L.; et al. Production and Characterization of Porous Fibroin Scaffolds for Regenerative Medical Application. In Vivo 2019, 33, 757–762. [Google Scholar] [CrossRef]
- Wu, D.T.; Munguia-Lopez, J.G.; Cho, Y.W.; Ma, X.; Song, V.; Zhu, Z.; Tran, S.D. Polymeric Scaffolds for Dental, Oral, and Craniofacial Regenerative Medicine. Molecules 2021, 26, 7043. [Google Scholar] [CrossRef] [PubMed]



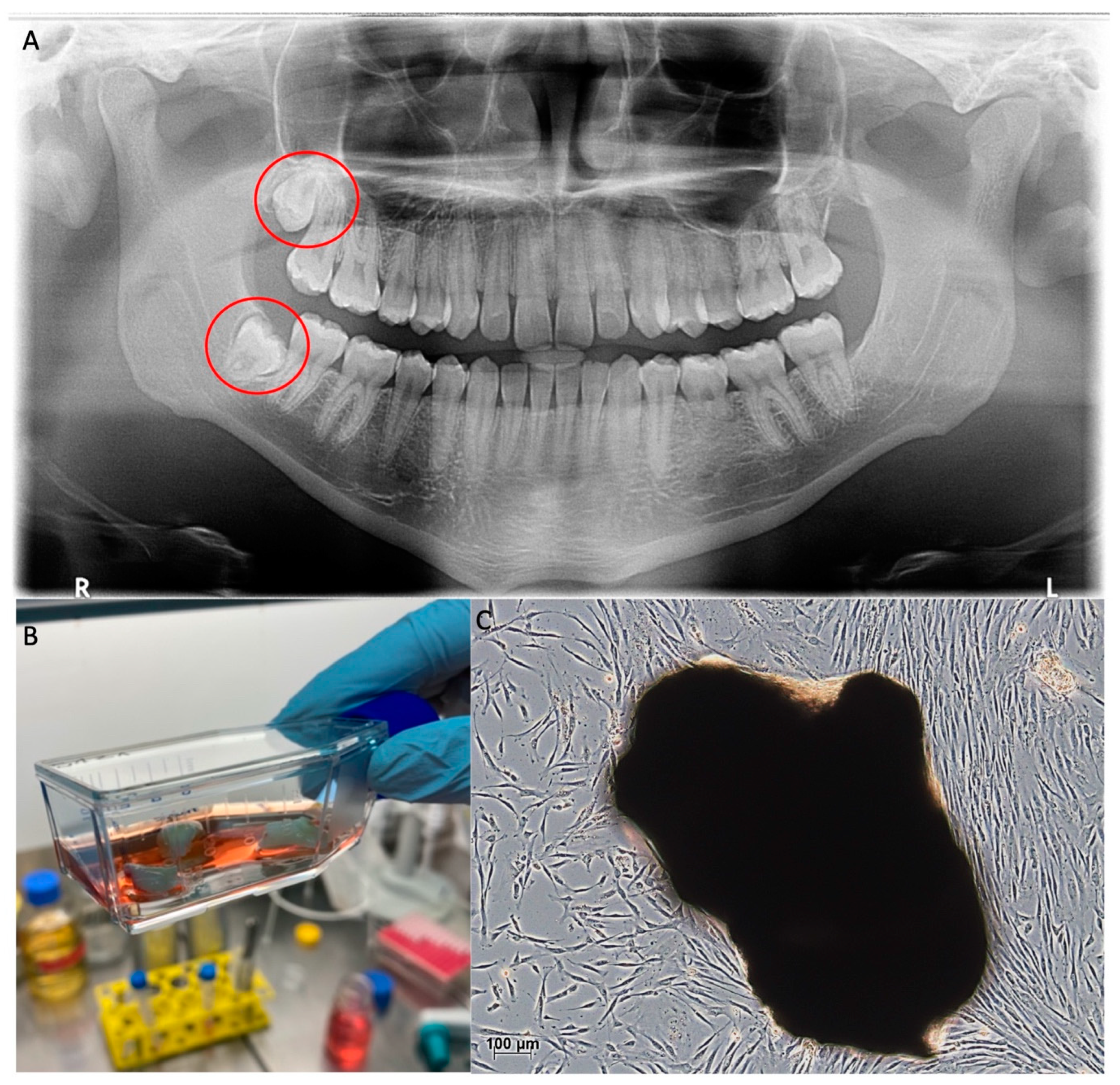{kind=link}
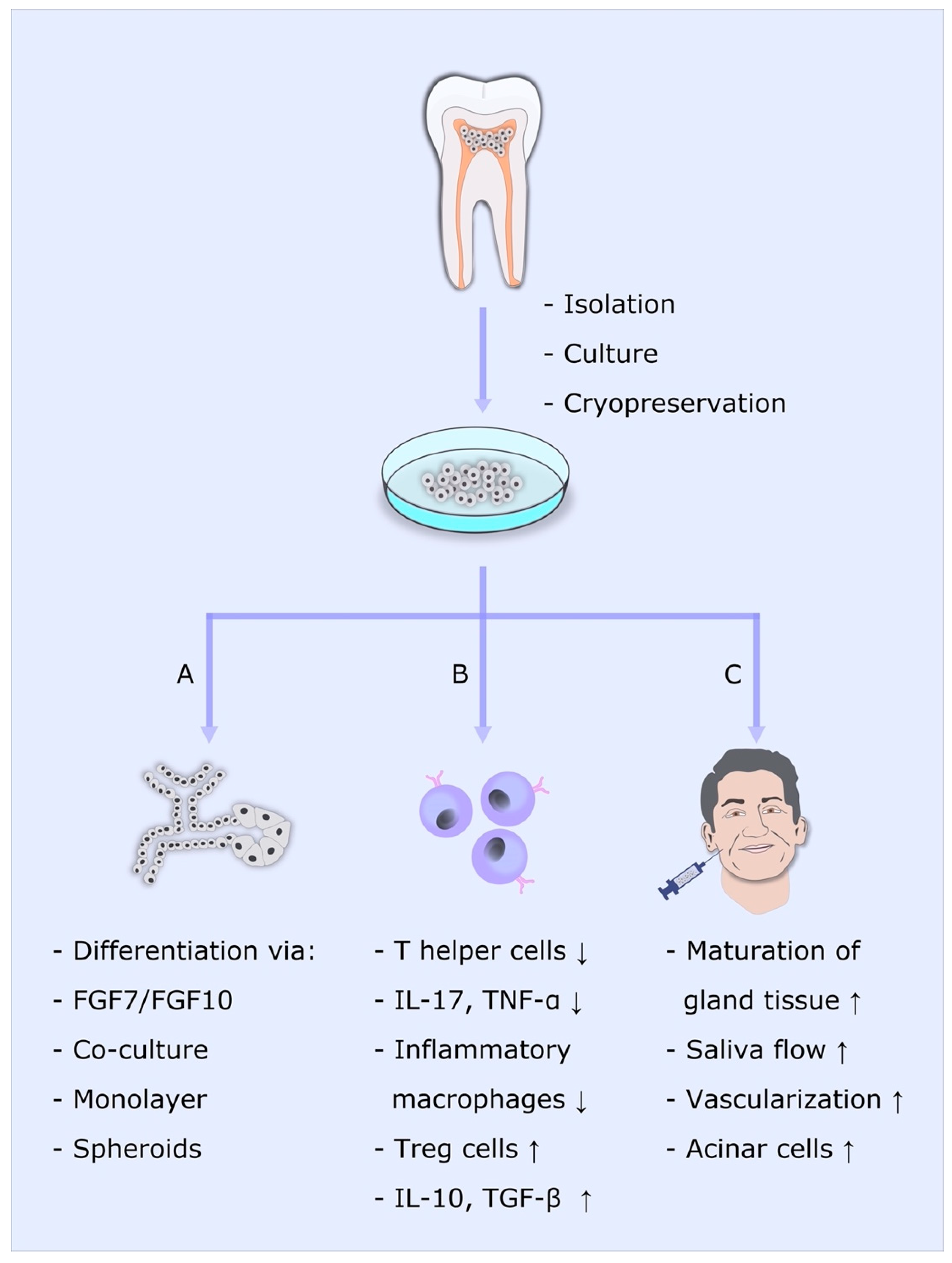{kind=link}
| Mechanism | Author, Year |
|---|---|
| Effect on primary cells | |
| Increased development of acinar structures and expression of LAMP-1 and CD44 after coculture of human salivary gland cells with DPSCs | [55] Reyes et al., 2013 |
| Increased saliva flow after DPSC injection into radiated salivary glands of mice | [57] Yamamura et al., 2013 |
| Decreased acinar cell vacuolization and increased IL-10 serum levels after DPSC injection into diabetic rats’ salivary glands | [58] Narmada et al., 2019 |
| Increase of vascularization, TGF-β serum level and acinar cell number after DPSC injection into diabetic rats’ salivary glands | [59] Suciadi et al., 2019 |
| Immunomodulatory effects | |
| Decreased apoptotic cell number in salivary glands of diabetic rats after injection of DPSCs in tail veins; also reduced expression of ATG5 and Beclin-1 as well as suppression of Th1 and Tfh cells in spleen while increased number of Treg cells | [79] Du et al., 2019 |
| Inhibition of CD4+T cells’ differentiation into T helper 17 cells and reduction of IL-17 and TNF-α, promotion of Treg cells and increased release of IL-10 and TGF-β | [77] Ji et al., 2019 |
| Downregulation of caspase-3 and upregulation of VEGF, decreased blood glucose, improved gland weight and salivary flow in diabetic rats after injection of DPSCs into the tail vein | [82] Al-Serwi et al., 2021 |
| Treatment of mice in salivary gland duct ligation model with DPSC-conditioned medium leads to increased expression of CK5, AQP5 | [83] Takeuchi et al., 2020 |
| DPSC exosomes caused macrophages to transform from proinflammatory phenotype to anti-inflammatory phenotype | [87] Shen et al., 2020 |
| Differentiation of DPSC to acinar cells | |
| Differentiation of DPSC via coculture with acinar cells in monolayer and expression of specific acinar morphology and markers such as CK8, amylase | [15] Yan et al., 2020 |
| 3D culture of DPSCs differentiated into acinar-like cells using FGF 10 | [14] Adine et al., 2018 |
| Differentiation of DPSC into acinar-like cells expressing AQP5 and αSMA after induction via FGF 7 | [79] Akashi et al., 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muallah, D.; Matschke, J.; Kappler, M.; Kroschwald, L.M.; Lauer, G.; Eckert, A.W. Dental Pulp Stem Cells for Salivary Gland Regeneration—Where Are We Today? Int. J. Mol. Sci. 2023, 24, 8664. https://doi.org/10.3390/ijms24108664
Muallah D, Matschke J, Kappler M, Kroschwald LM, Lauer G, Eckert AW. Dental Pulp Stem Cells for Salivary Gland Regeneration—Where Are We Today? International Journal of Molecular Sciences. 2023; 24(10):8664. https://doi.org/10.3390/ijms24108664
Chicago/Turabian StyleMuallah, David, Jan Matschke, Matthias Kappler, Lysann Michaela Kroschwald, Günter Lauer, and Alexander W. Eckert. 2023. "Dental Pulp Stem Cells for Salivary Gland Regeneration—Where Are We Today?" International Journal of Molecular Sciences 24, no. 10: 8664. https://doi.org/10.3390/ijms24108664