
In Vitro and In Vivo Effects of the Combination of Polypurine Reverse Hoogsteen Hairpins against HER-2 and Trastuzumab in Breast Cancer Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
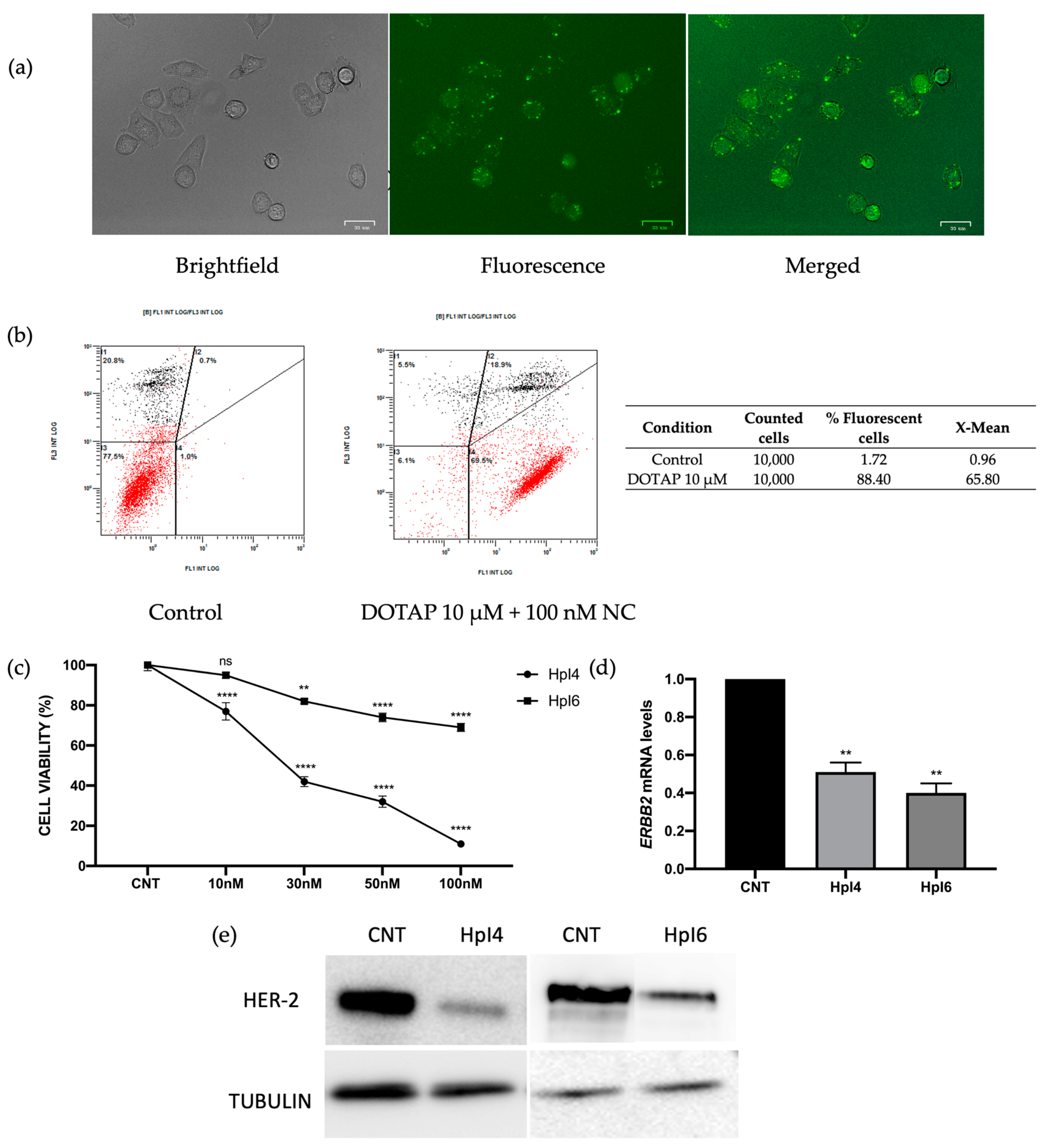
2.1. Effects of PPRHs on SKBR-3 Cells
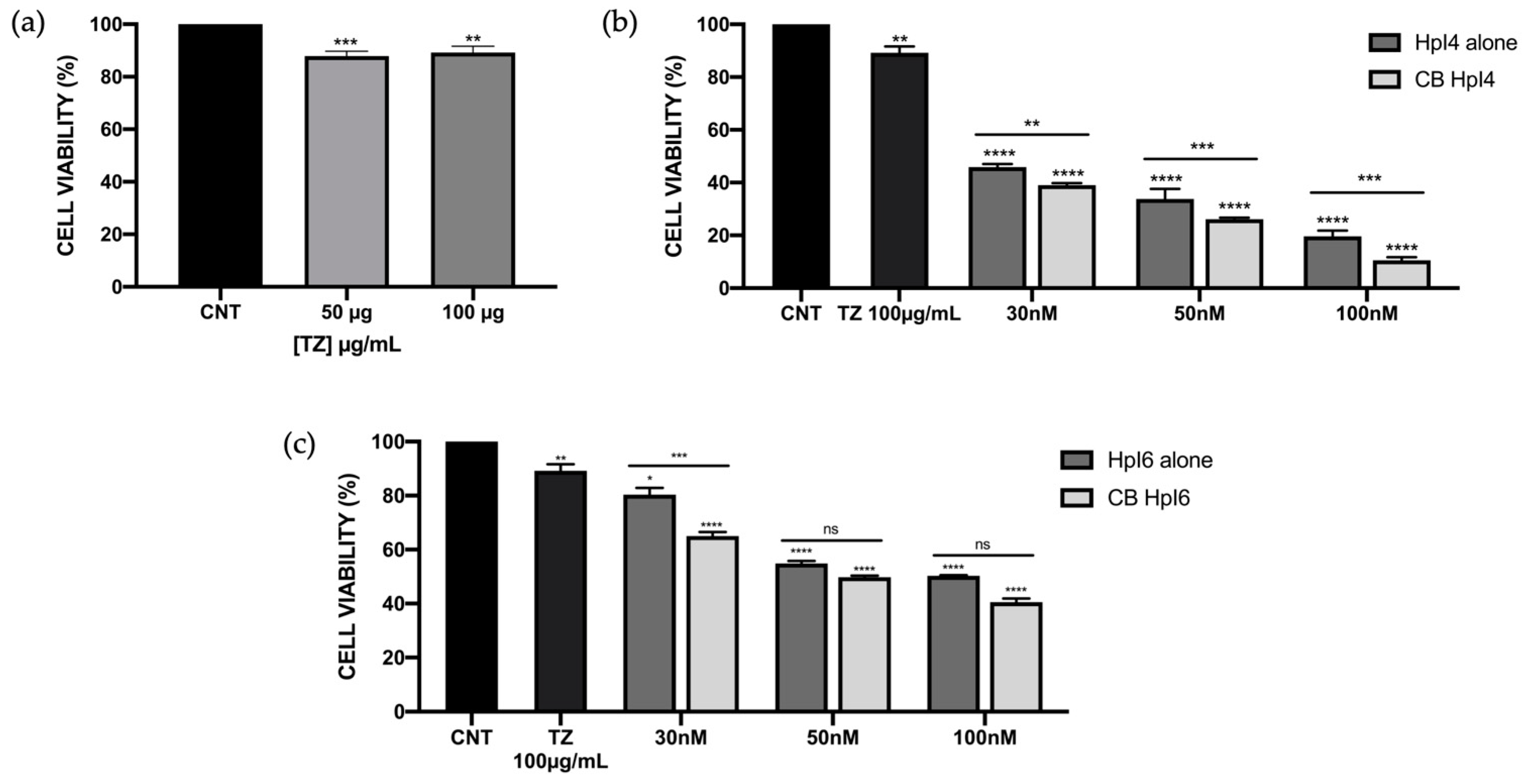
2.2. Effects of the Combination of PPRHs and Trastuzumab on SKBR-3 Cell Viability
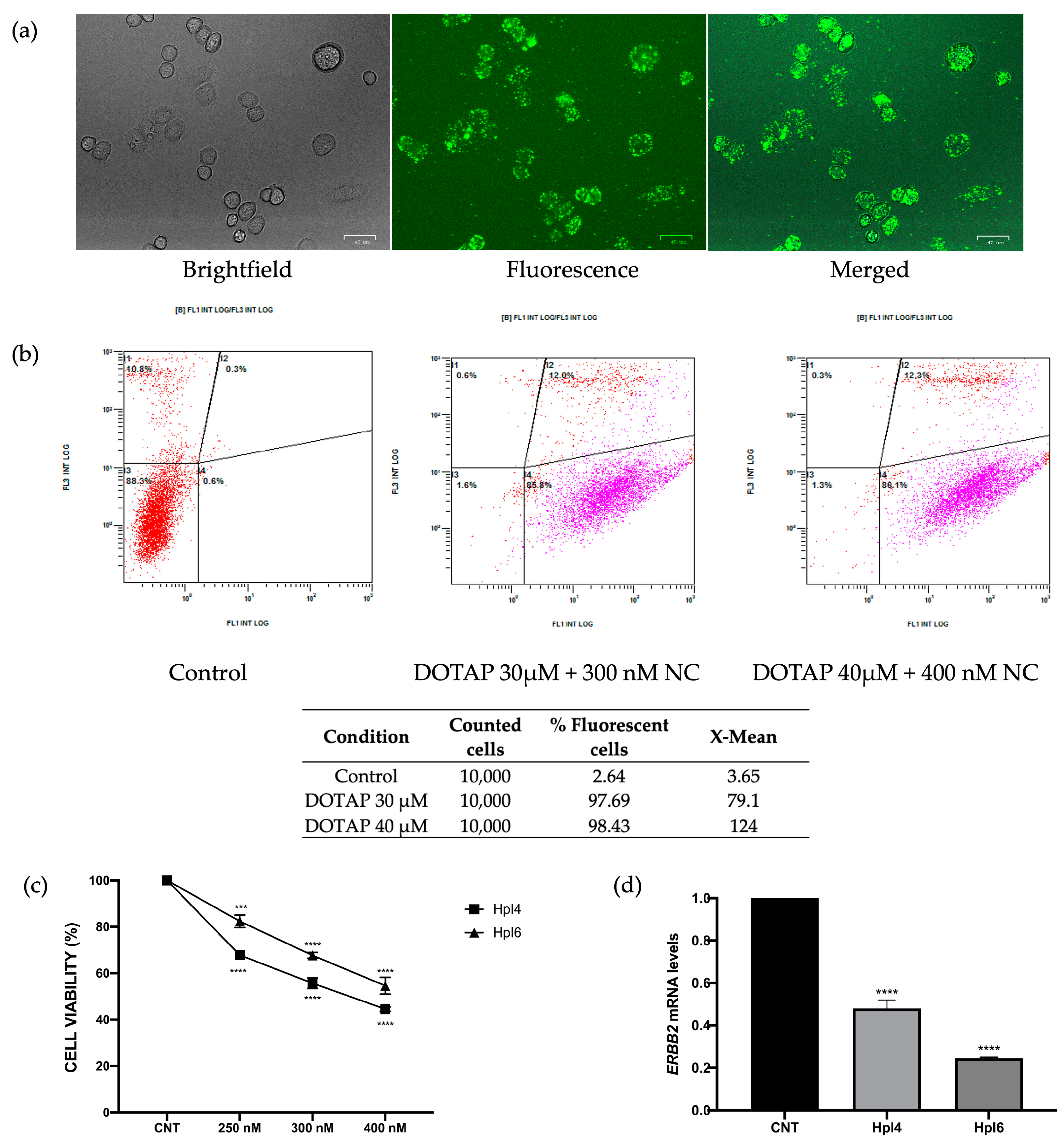
2.3. In Vitro Effects of PPRHs in MDA-MB-453 Cells
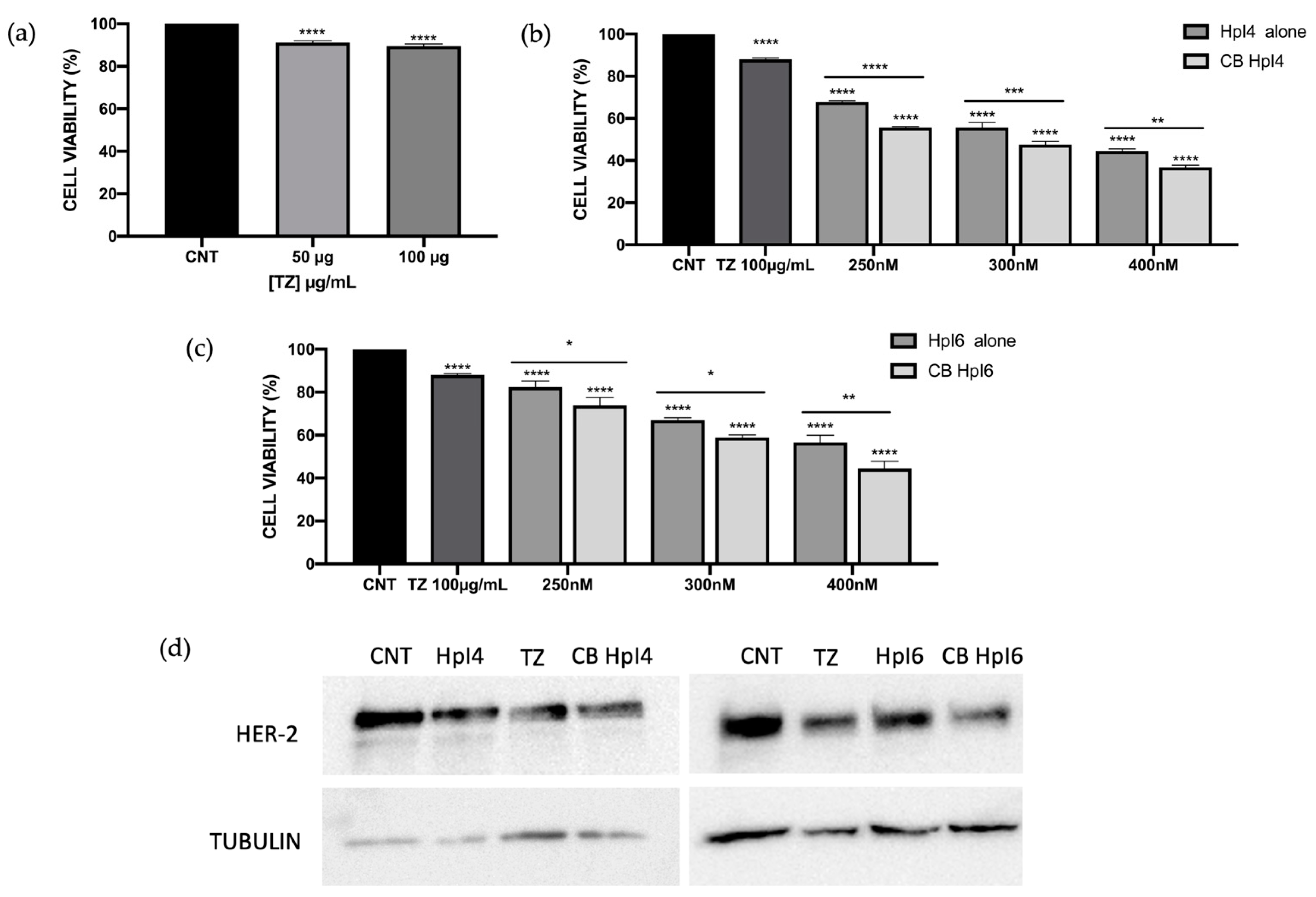
2.4. Effects of the Combination of PPRHs and Trastuzumab on MDA-MB-453 Cell Viability
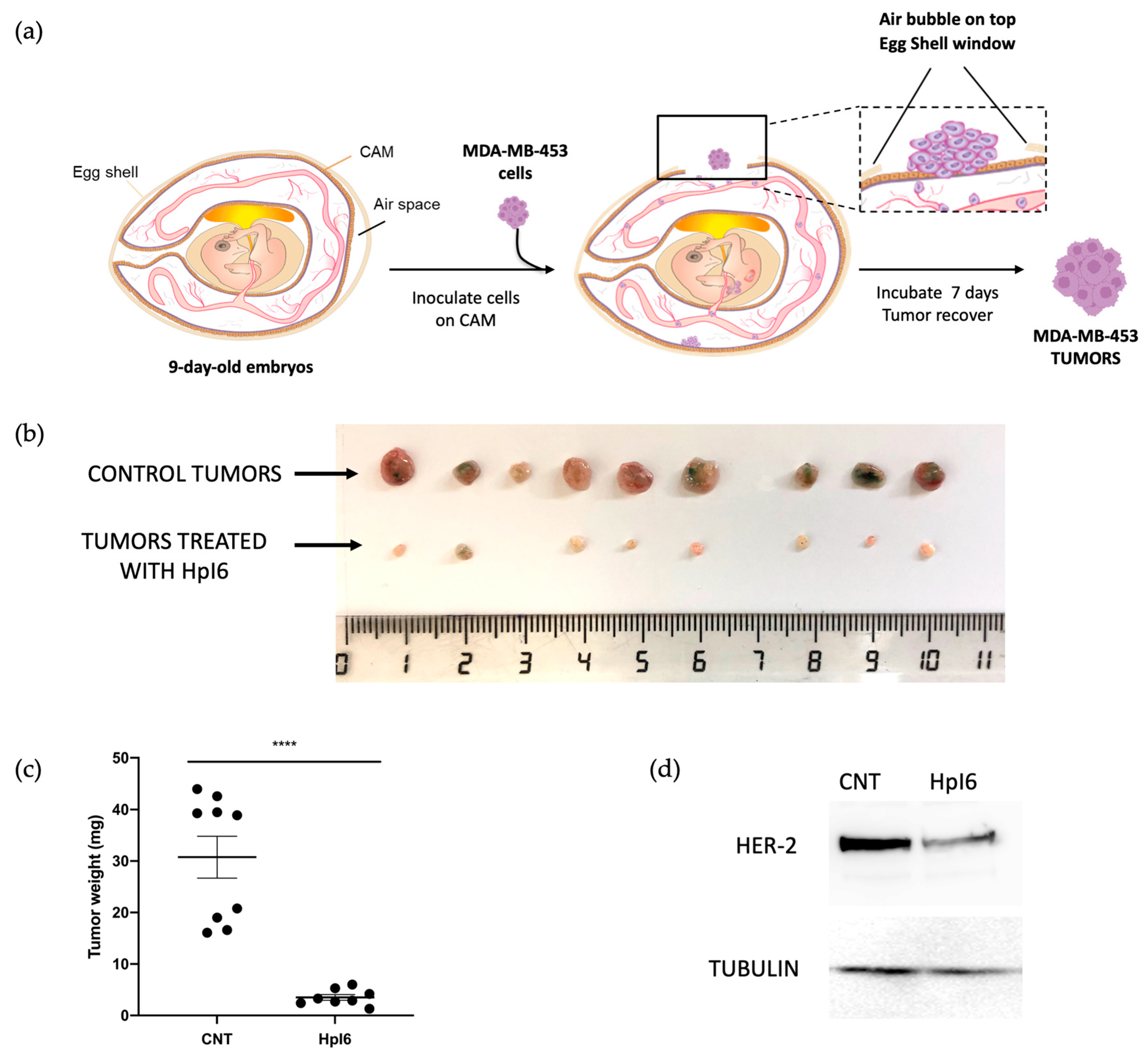
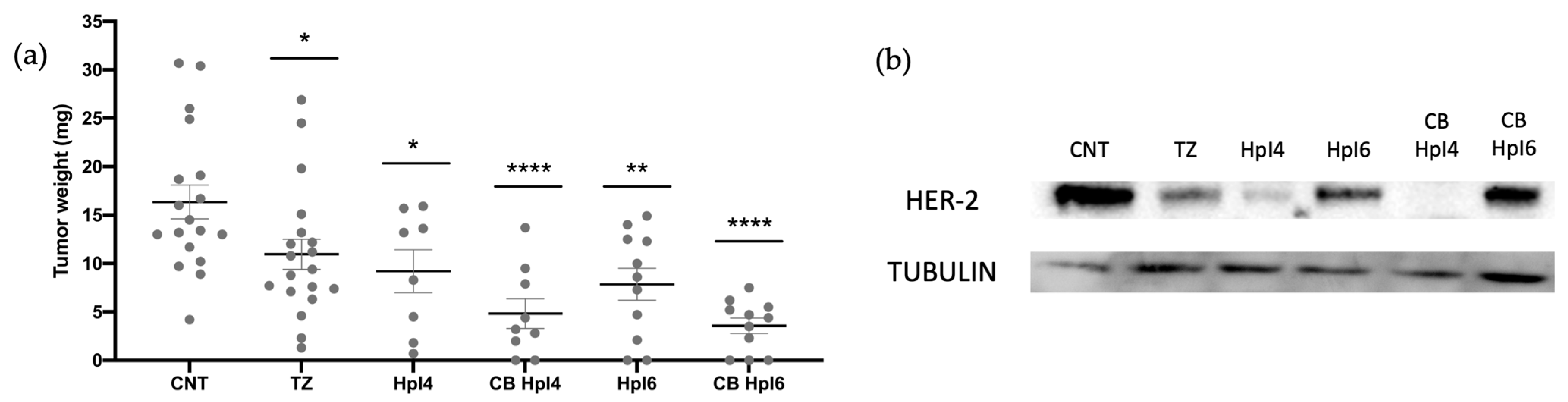
2.5. In Vivo Assays in the CAM Model
3. Discussion
4. Materials and Methods
4.1. Oligonucleotides
4.2. Cell Culture
4.3. Transfection of PPRHs
4.4. Fluorescent Microscopy and Flow Cytometry
4.5. MTT Assay and Synergism Analyses
4.6. RNA Extraction and Analysis
4.7. Western Blot Analyses
4.8. CAM Assays
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciudad, C.J.; Rodríguez, L.; Villalobos, X.; Félix, A.J.; Noé, V. Polypurine Reverse Hoogsteen Hairpins as a Gene Silencing Tool for Cancer. Curr. Med. Chem. 2017, 24, 2809–2826. [Google Scholar] [CrossRef] [PubMed]
- De Almagro, M.C.; Mencia, N.; Noé, V.; Ciudad, C.J. Coding polypurine hairpins cause target-induced cell death in breast cancer cells. Hum. Gene Ther. 2011, 22, 451–463. [Google Scholar] [CrossRef] [PubMed]
- De Almagro, M.C.; Coma, S.; Noé, V.; Ciudad, C.J. Polypurine hairpins directed against the template strand of DNA knock down the expression of mammalian genes. J. Biol. Chem. 2009, 284, 11579–11589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos, X.; Rodríguez, L.; Prévot, J.; Oleaga, C.; Ciudad, C.J.; Noé, V. Stability and immunogenicity properties of the gene-silencing polypurine reverse hoogsteen hairpins. Mol. Pharm. 2014, 11, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; Villalobos, X.; Dakhel, S.; Padilla, L.; Hervas, R.; Luis Hernández, J.; Ciudad, C.J.; Noe, V. Polypurine reverse Hoogsteen hairpins as a gene therapy tool against survivin in human prostate cancer PC3 cells in vitro and in vivo n.d. Biochem. Pharmacol. 2013, 86, 1541–1554. [Google Scholar] [CrossRef] [PubMed]
- Bener, G.; Félix, A.J.; Sánchez de Diego, C.; Pascual Fabregat, I.; Ciudad, C.J.; Noé, V. Silencing of CD47 and SIRPα by Polypurine reverse Hoogsteen hairpins to promote MCF-7 breast cancer cells death by PMA-differentiated THP-1 cells. BMC Immunol. 2016, 17, 32. [Google Scholar] [CrossRef] [Green Version]
- Enríquez, M.M.M.; Félix, A.J.; Ciudad, C.J.; Noé, V. Cancer immunotherapy using PolyPurine Reverse Hoogsteen hairpins targeting the PD-1/PD-L1 pathway in human tumor cells. PLoS ONE 2018, 13, e0206818. [Google Scholar] [CrossRef]
- Aubets, E.; Noé, V.; Ciudad, C.J. Targeting replication stress response using polypurine reverse hoogsteen hairpins directed against WEE1 and CHK1 genes in human cancer cells. Biochem. Pharmacol. 2020, 175, 113911. [Google Scholar] [CrossRef]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef]
- Kreutzfeldt, J.; Rozeboom, B.; Dey, N.; De, P. The trastuzumab era: Current and upcoming targeted HER2+ breast cancer therapies. Am. J. Cancer Res. 2020, 10, 1045–1067. [Google Scholar]
- Hasmann, M. Targeting HER2 by monoclonal antibodies for cancer therapy. In Introduction to Biological and Small Molecule Drug Research and Development: Theory and Case Studies; Elsevier: Amsterdam, The Netherlands, 2013; pp. 283–305. [Google Scholar] [CrossRef]
- Rodríguez, L.; Villalobos, X.; Solé, A.; Lliberós, C.; Ciudad, C.J.; Noé, V. Improved design of PPRHs for gene silencing. Mol. Pharm. 2015, 12, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Ayala de la Peña, F.; Andrés, R.; Garcia-Sáenz, J.A.; Manso, L.; Margelí, M.; Dalmau, E.; Pernas, S. SEOM clinical guidelines in early stage breast cancer (2018). Clin. Transl. Oncol. 2019, 21, 18–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandó, P.; Rivero, S.G.; Rizzo, M.M.; Pinkasz, M.; Levy, E.M. Targeting ADCC: A different approach to HER2 breast cancer in the immunotherapy era. Breast 2021, 60, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Nogueira, P.; Mancino, M.; Fuster, G.; Lopez-Plana, A.; Jauregui, P.; Almendro, V.; Bragado, P. Tumor-Associated Fibroblasts Promote HER2-Targeted Therapy Resistance through FGFR2 Activation n.d. Clin. Cancer Res 2020, 26, 1432–1448. [Google Scholar] [CrossRef] [Green Version]
- Villalobos, X.; Rodríguez, L.; Solé, A.; Lliberós, C.; Mencia, N.; Ciudad, C.J.; Noe, V. Effect of Polypurine Reverse Hoogsteen Hairpins on Relevant Cancer Target Genes in Different Human Cell Lines. Nucleic Acid Ther. 2015, 25, 198–208. [Google Scholar] [CrossRef]
- Rait, A.S.; Pirollo, K.F.; Rait, V.; Krygier, J.E.; Xiang, L.; Chang, E.H. Inhibitory effects of the combination of HER-2 antisense oligonucleotide and chemotherapeutic agents used for the treatment of human breast cancer. Cancer Gene Ther. 2001, 8, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Xu, Y.; Song, S.; Wu, Z.; Duan, H. Antisense oligodeoxynucleotide targeting HER2 mRNA sensitized docetaxel in breast cancer treatment. Pharm. Biol. 2011, 49, 1167–1172. [Google Scholar] [CrossRef]
- Colomer, R.; Lupu, R.; Bacus, S.S.; Gelmann, E.P. erbB-2 antisense oligonucleotides inhibit the proliferation of breast carcinoma cells with erbB-2 oncogene amplification. Br. J. Cancer 1994, 70, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, A.; Charo, J.; Parapuram, S.K.; Hunt, R.C.; Hunt, D.M.; Seliger, B.; Kiessling, R. Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int. J. Cancer 2004, 108, 71–77. [Google Scholar] [CrossRef]
- Gu, S.; Hu, Z.; Ngamcherdtrakul, W.; Castro, D.J.; Morry, J.; Reda, M.M.; Gray, J.W.; Yantasee, W. Therapeutic siRNA for drug-resistant HER2-positive breast cancer. Oncotarget 2016, 7, 14727–14741. [Google Scholar] [CrossRef] [Green Version]
- PDF, Temporary Ex Vivo Inhibition of the Expression of the Human Oncogene HER2 (NEU) by a Triple Helix-Forming Oligonucleotide n.d. Available online: https://www.researchgate.net/publication/14635491_Temporary_ex_vivo_inhibition_of_the_expression_of_the_human_oncogene_HER2_NEU_by_a_triple_helix-forming_oligonucleotide (accessed on 28 January 2023).
- Kaushik Tiwari, M.; Colon-Rios, D.A.; Tumu, H.C.R.; Liu, Y.; Quijano, E.; Krysztofiak, A.; Chan, C.; Song, E. Direct targeting of amplified gene loci for proapoptotic anticancer therapy. Nat. Biotechnol. 2021, 40, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.D.; Lee, H.J.; Yadav, D.K. Clinical updates on tyrosine kinase inhibitors in HER2-positive breast cancer. Front. Pharmacol. 2022, 13, 5276. [Google Scholar] [CrossRef] [PubMed]
- Jagosky, M.; Tan, A.R. Combination of Pertuzumab and Trastuzumab in the Treatment of HER2-Positive Early Breast Cancer: A Review of the Emerging Clinical Data. Breast Cancer Targets Ther. 2021, 13, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.H.; Kacew, A.J.; Dreyer, M.; Serritella, A.V.; Knoebel, R.W.; Strohbehn, G.W.; Ratain, M.J. Alternative trastuzumab dosing strategies in HER2-positive early breast cancer are associated with patient out-of-pocket savings. NPJ Breast Cancer 2022, 8, 32. [Google Scholar] [CrossRef]
- DeBord, L.C.; Pathak, R.R.; Villaneuva, M.; Liu, H.-C.; Harrington, D.A.; Yu, W.; Sikora, A.G. The chick chorioallantoic membrane (CAM) as a versatile patient-derived xenograft (PDX) platform for precision medicine and preclinical research. Am. J. Cancer Res. 2018, 8, 1642. [Google Scholar]
- Lokman, N.A.; Elder, A.S.F.; Ricciardelli, C.; Oehler, M.K. Chick chorioallantoic membrane (CAM) assay as an in vivo model to study the effect of newly identified molecules on ovarian cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef] [Green Version]
- Pawlikowska, P.; Tayoun, T.; Oulhen, M.; Faugeroux, V.; Rouffiac, V.; Aberlenc, A.; Farace, F. Exploitation of the chick embryo chorioallantoic membrane (CAM) as a platform for anti-metastatic drug testing. Sci. Rep. 2020, 10, 16876. [Google Scholar] [CrossRef]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef]
- Rousset, X.; Maillet, D.; Grolleau, E.; Barthelemy, D.; Calattini, S.; Brevet, M.; Payen, L. Embryonated Chicken Tumor Xenografts Derived from Circulating Tumor Cells as a Relevant Model to Study Metastatic Dissemination: A Proof of Concept. Cancers 2022, 14, 4085. [Google Scholar] [CrossRef]
- Triplex-Forming Oligonucleotide Target Sequence Search—The University of Texas MD Anderson Cancer Center n.d. Available online: http://utw10685.utweb.utexas.edu/tfo/ (accessed on 8 December 2022).
- Chou, T.C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Kain, K.H.; Miller, J.W.I.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 2014, 243, 216–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubeldia-Plazaola, A.; Recalde-Percaz, L.; Moragas, N.; Alcaraz, M.; Chen, X.; Mancino, M.; Fuster, G. Glucocorticoids promote transition of ductal carcinoma in situ to invasive ductal carcinoma by inducing myoepithelial cell apoptosis. Breast Cancer Res. 2018, 20, 65. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose HpI4 (nM) | Dose HpI6 (nM) | Dose TZ (µg/mL) | CI Media | Description |
|---|---|---|---|---|
| 30 | - | 100 | 0.76 | Moderate synergism |
| 50 | - | 100 | 0.72 | Moderate synergism |
| 100 | - | 100 | 0.51 | Synergism |
| - | 30 | 100 | 0.66 | Synergism |
| - | 50 | 100 | 0.60 | Synergism |
| - | 100 | 100 | 0.93 | Nearly additive |
| Dose HpI4 (nM) | Dose HpI6 (nM) | Dose TZ (µg/mL) | CI Media | Description |
|---|---|---|---|---|
| 250 | - | 100 | 0.80 | Moderate synergism |
| 300 | - | 100 | 0.81 | Moderate synergism |
| 400 | - | 100 | 0.86 | Slight synergism |
| - | 250 | 100 | 0.90 | Slight synergism |
| - | 300 | 100 | 0.81 | Moderate synergism |
| - | 400 | 100 | 0.85 | Moderate synergism |
| Condition | Mean Weight (mg) | SEM | N | % Reduction vs. Control |
|---|---|---|---|---|
| Control | 16.35 | 1.75 | 18 | - |
| TZ 100 µg/mL | 10.96 | 1.56 | 19 | 32.97 |
| HpI4 300 nM | 9,21 | 2.21 | 8 | 43.65 |
| CB HpI4 | 4.83 | 1.54 | 9 | 70.44 |
| HpI6 300 nM | 7.86 | 1.65 | 11 | 51.96 |
| CB HpI6 | 3.57 | 0.80 | 11 | 78.15 |
| PPRH Name | Sequence from 5′ to 3′ | Target Location in ERBB2 |
|---|---|---|
| HpHer2-I4-T | GGGAGAGGGAGTGGGAACAGAGTGGG (T)5 GGGAGAGGGAGTGGGAACAGAGTGGG | Intron 4 |
| HpHer2-I6-C | AGATGAGAGGTGAGAAGGAAGGAGAGAG (T)5 AGATGAGAGGTGAGAAGGAAGGAGAGAG | Intron 6 |
| Negative control | AAGAAGAGAAGAAGAAGAA (T)5 AAGAAGAAGAAGAGAAGAA | None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Aguilar, E.; Fernández-Nogueira, P.; Fuster, G.; Carbó, N.; Ciudad, C.J.; Noé, V. In Vitro and In Vivo Effects of the Combination of Polypurine Reverse Hoogsteen Hairpins against HER-2 and Trastuzumab in Breast Cancer Cells. Int. J. Mol. Sci. 2023, 24, 7073. https://doi.org/10.3390/ijms24087073
López-Aguilar E, Fernández-Nogueira P, Fuster G, Carbó N, Ciudad CJ, Noé V. In Vitro and In Vivo Effects of the Combination of Polypurine Reverse Hoogsteen Hairpins against HER-2 and Trastuzumab in Breast Cancer Cells. International Journal of Molecular Sciences. 2023; 24(8):7073. https://doi.org/10.3390/ijms24087073
Chicago/Turabian StyleLópez-Aguilar, Ester, Patricia Fernández-Nogueira, Gemma Fuster, Neus Carbó, Carlos J. Ciudad, and Véronique Noé. 2023. "In Vitro and In Vivo Effects of the Combination of Polypurine Reverse Hoogsteen Hairpins against HER-2 and Trastuzumab in Breast Cancer Cells" International Journal of Molecular Sciences 24, no. 8: 7073. https://doi.org/10.3390/ijms24087073