
Eupatilin Ameliorates Hepatic Fibrosis and Hepatic Stellate Cell Activation by Suppressing β-catenin/PAI-1 Pathway
Abstract
:1. Introduction
2. Results
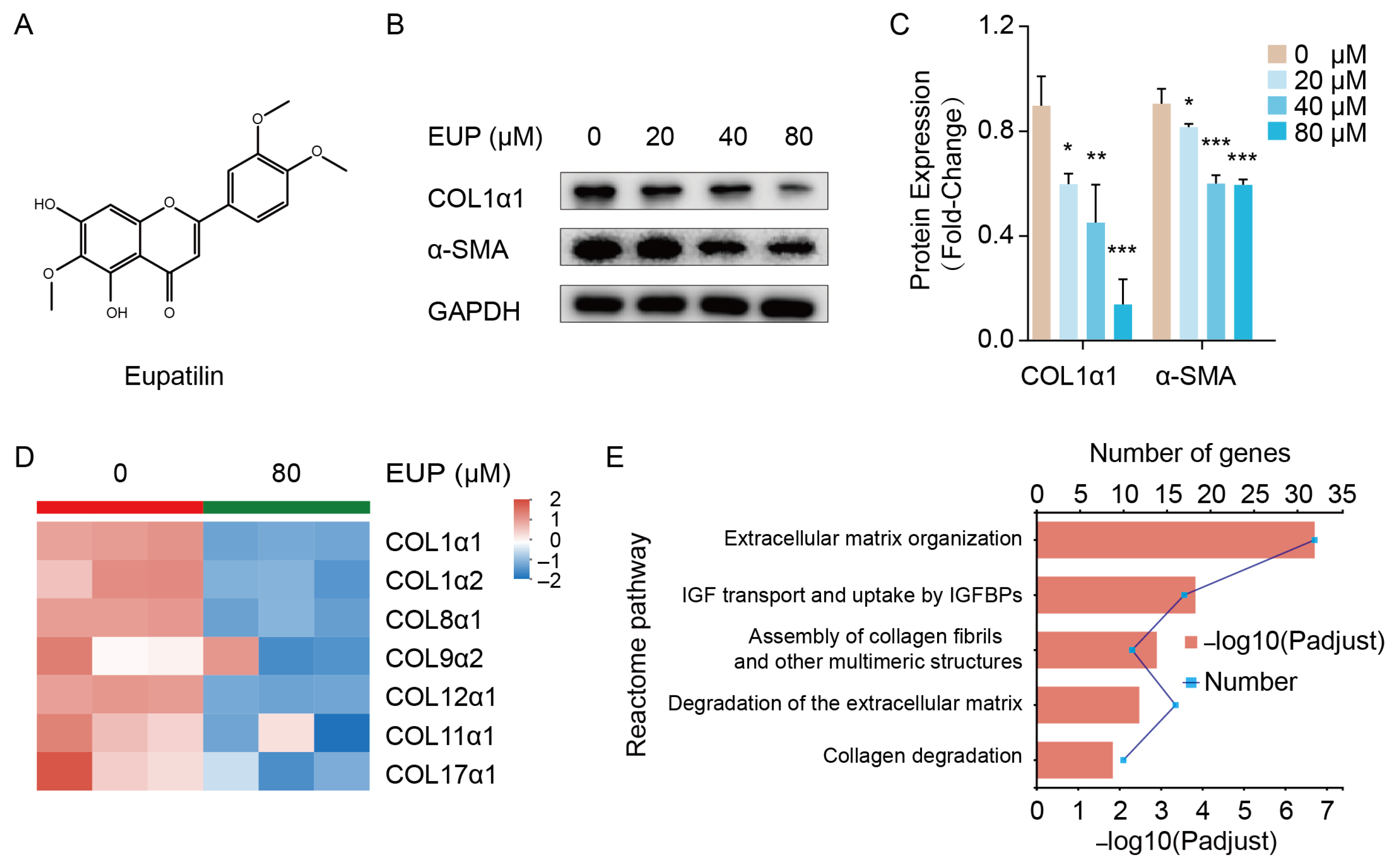
2.1. Eupatilin Represses the Activation of HSCs
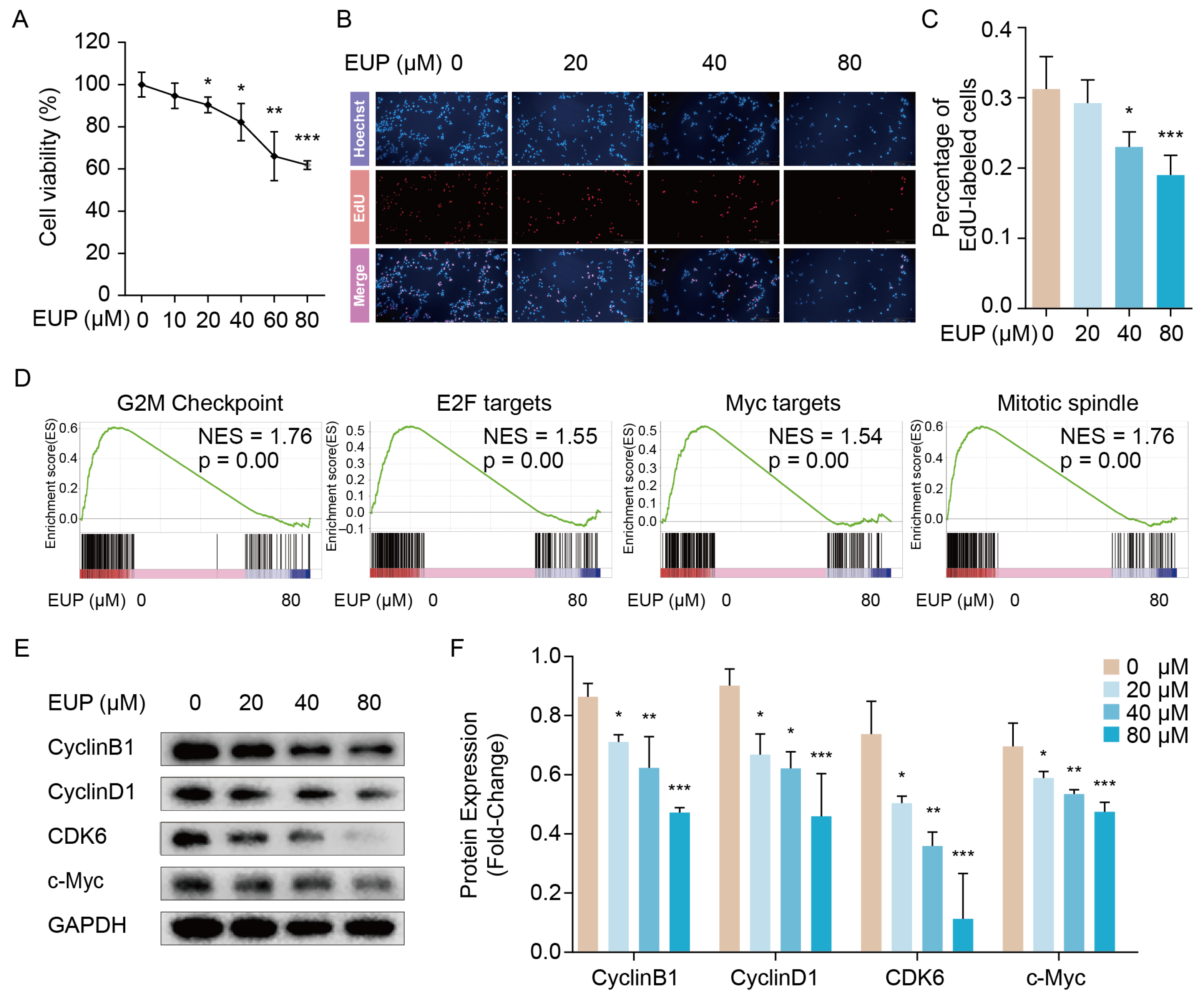
2.2. Eupatilin Inhibits the Proliferation of HSCs
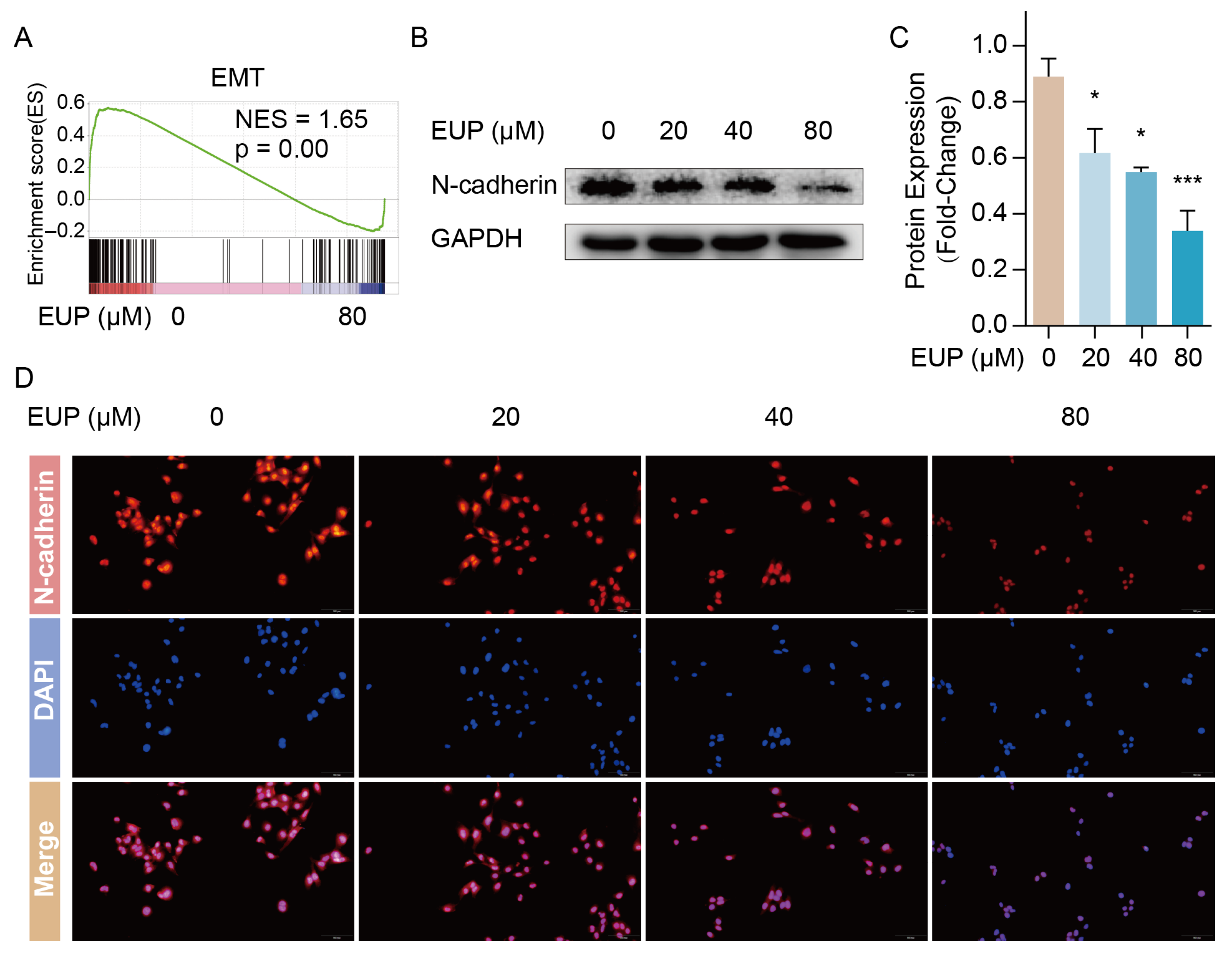
2.3. Eupatilin Inhibits the EMT of HSCs
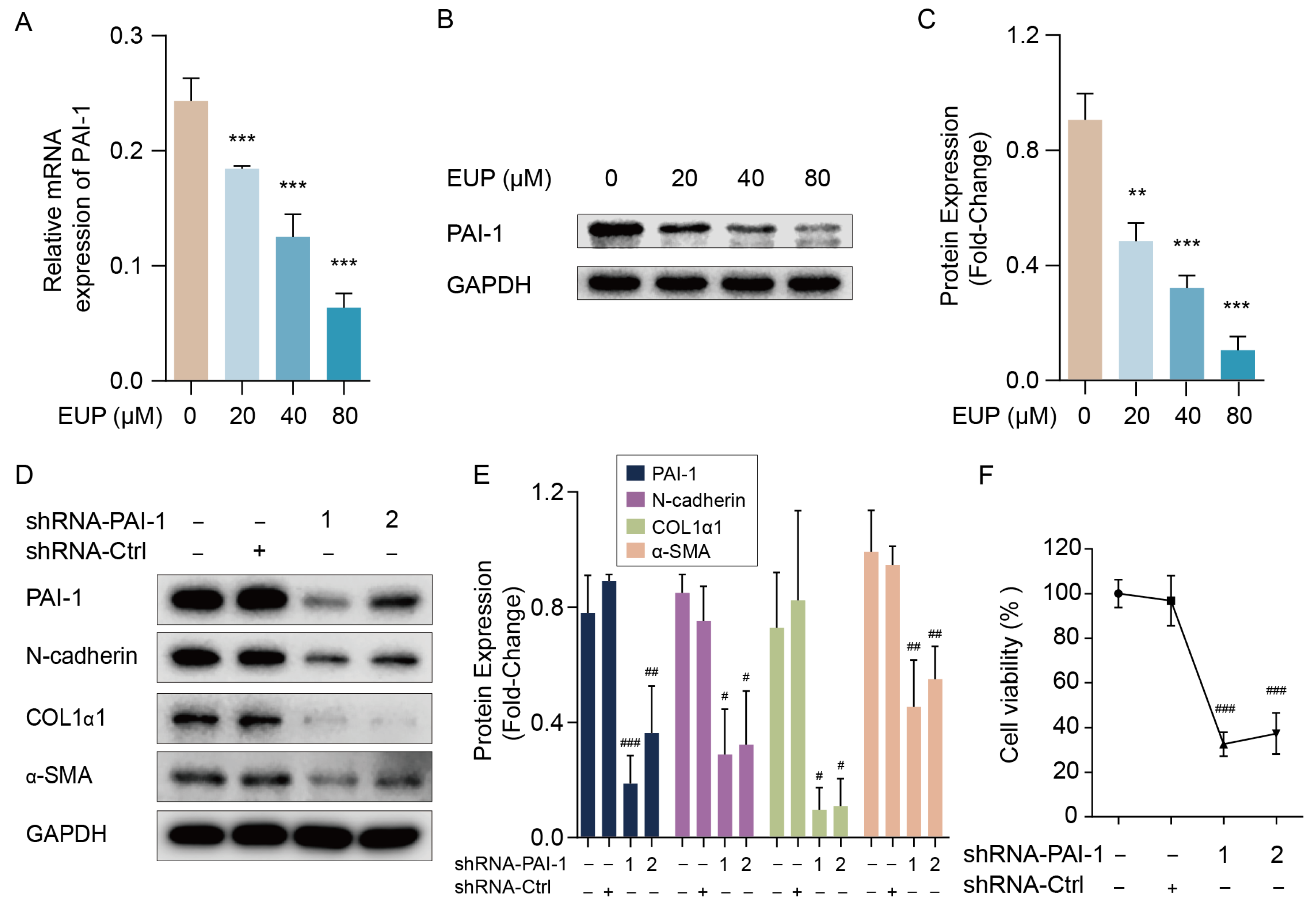
2.4. PAI-1 Regulates the EMT and Activation of HSCs
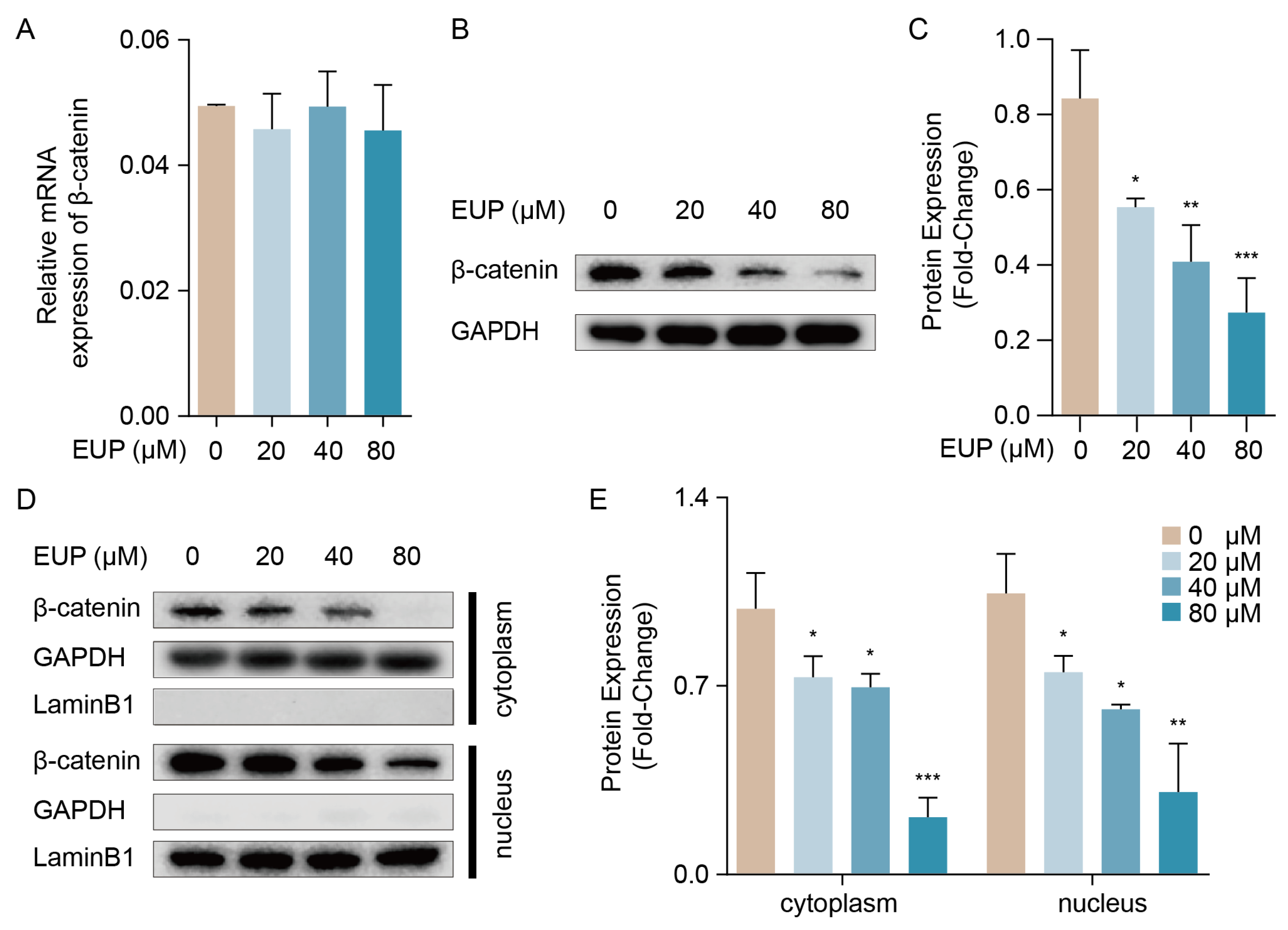
2.5. The β-Catenin Pathway Is Necessary for Eupatilin-Induced EMT Repression in HSCs
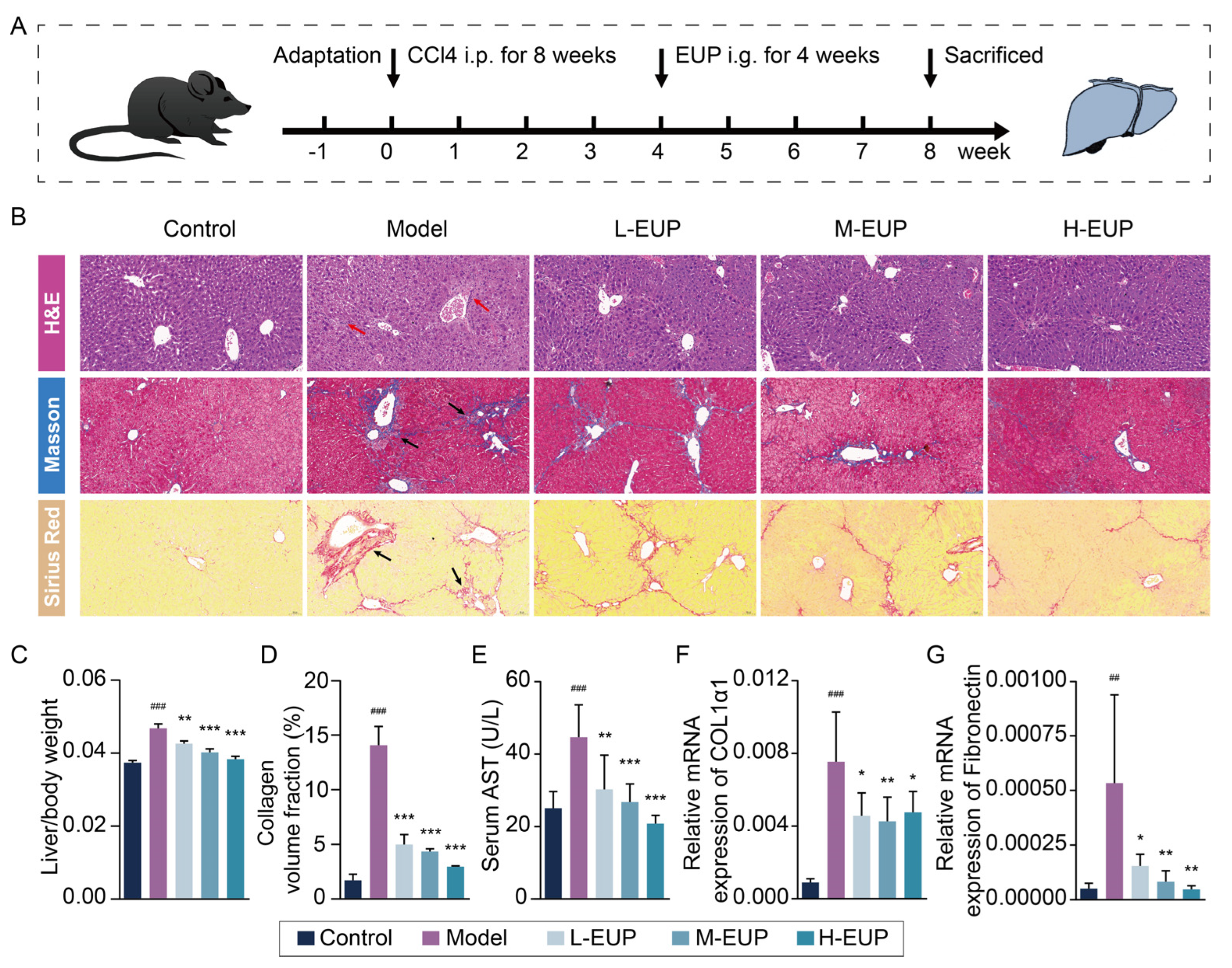
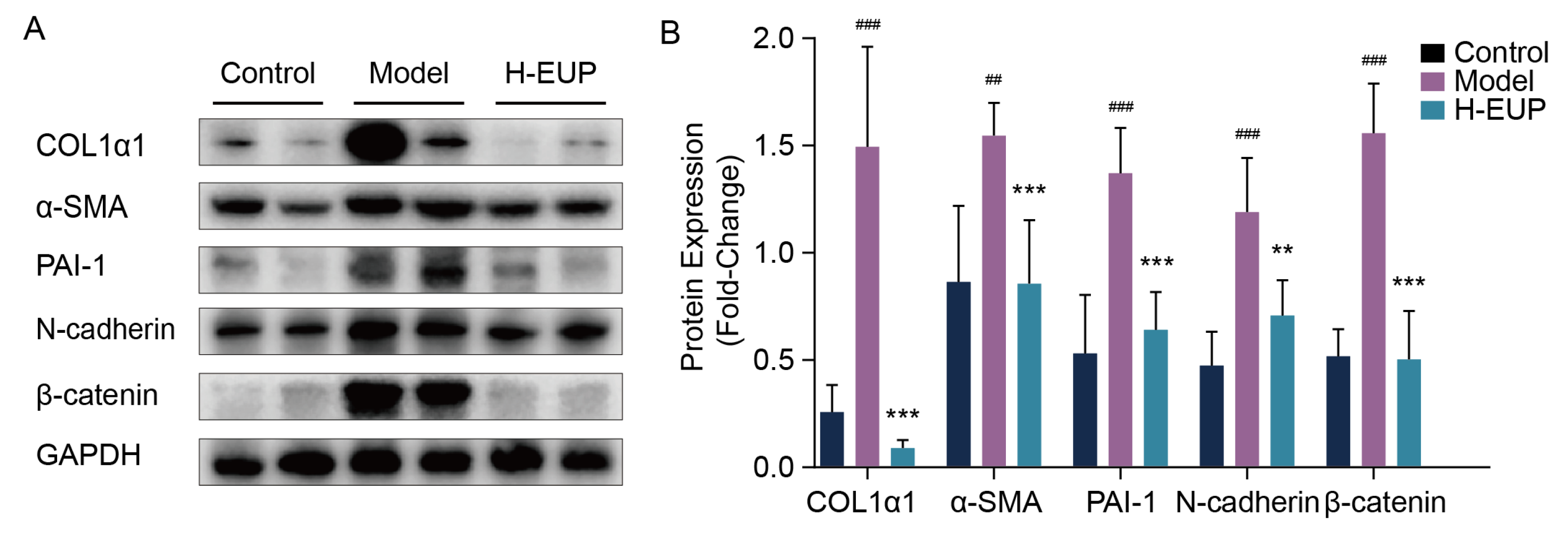
2.6. Eupatilin Alleviates Hepatic Fibrosis in CCl4-Treated Mouse Model
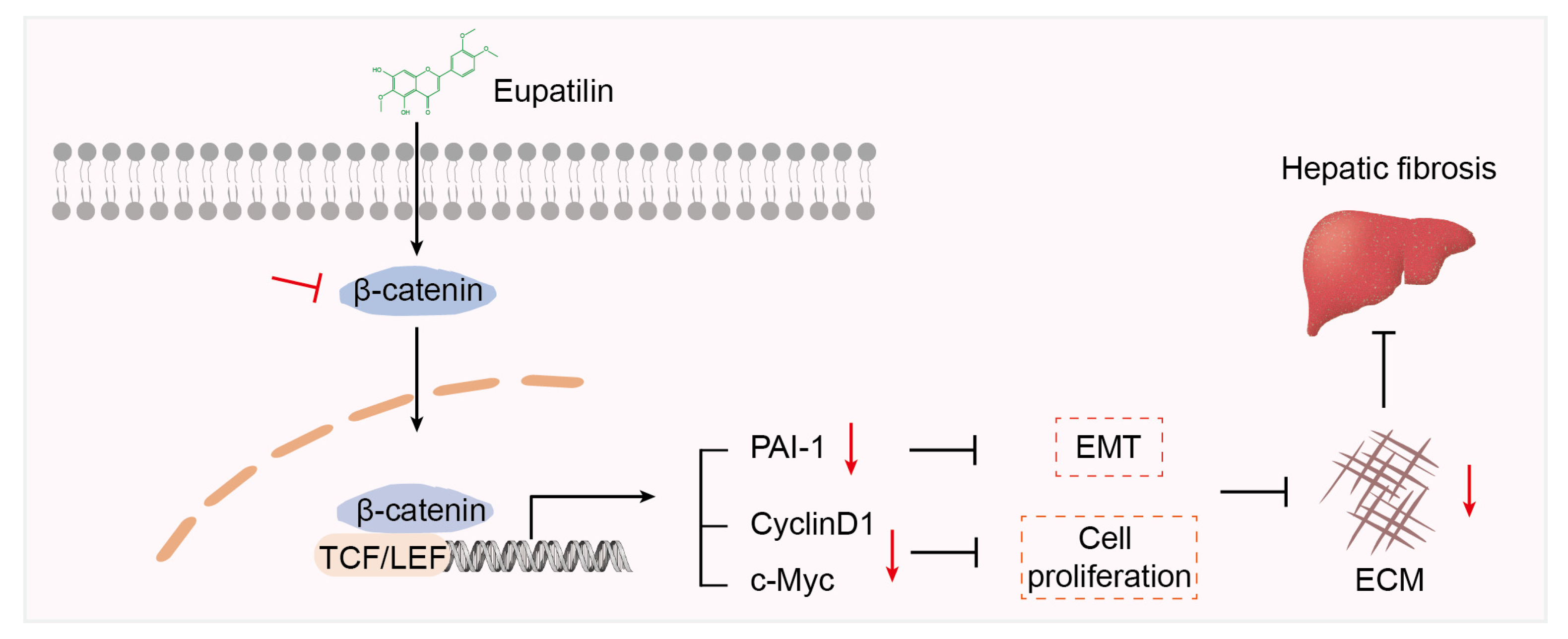
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Cell Viability
4.4. EdU Assay
4.5. RNA Isolation and Real-Time PCR
4.6. Western Blotting
4.7. RNA Sequencing Analysis
4.8. shRNA Interference
4.9. Animal Procedures and Treatments
4.10. Liver Histopathology
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.S.; Runyon, B.A. Treatment of Patients with Cirrhosis. N. Engl. J. Med. 2016, 375, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, L.; Liu, Z.; Ci, L.; Shuai, C.; Lv, X.; Li, J. Research progress on the anti-hepatic fibrosis action and mechanism of natural products. Int. Immunopharmacol. 2019, 75, 105765. [Google Scholar] [CrossRef]
- Li, N.; Zhang, X.; Zhou, J.; Li, W.; Shu, X.; Wu, Y.; Long, M. Multiscale biomechanics and mechanotransduction from liver fibrosis to cancer. Adv. Drug Deliv. Rev. 2022, 188, 114448. [Google Scholar] [CrossRef]
- Bobowski-Gerard, M.; Zummo, F.P.; Staels, B.; Lefebvre, P.; Eeckhoute, J. Retinoids Issued from Hepatic Stellate Cell Lipid Droplet Loss as Potential Signaling Molecules Orchestrating a Multicellular Liver Injury Response. Cells 2018, 7, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, R.M.; El-Meguid, M.A.; Salum, G.M.; El Awady, M.K. Key Players of Hepatic Fibrosis. J. Interf. Cytokine Res. 2020, 40, 472–489. [Google Scholar] [CrossRef]
- Ma, L.; Wei, J.; Zeng, Y.; Liu, J.; Xiao, E.; Kang, Y.; Kang, Y. Mesenchymal stem cell-originated exosomal circDIDO1 suppresses hepatic stellate cell activation by miR-141-3p/PTEN/AKT pathway in human liver fibrosis. Drug Deliv. 2022, 29, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Li, Z.; Wang, G.; Ling, L.; Wu, Y.; Zhang, C. Carvedilol attenuates liver fibrosis by suppressing autophagy and promoting apoptosis in hepatic stellate cells. Biomed. Pharmacother. 2018, 108, 1617–1627. [Google Scholar] [CrossRef]
- Zhang, C.-Y.; Yuan, W.-G.; He, P.; Lei, J.-H.; Wang, C.-X. Liver fibrosis and hepatic stellate cells: Etiology, pathological hallmarks and therapeutic targets. World J. Gastroenterol. 2016, 22, 10512–10522. [Google Scholar] [CrossRef]
- Hu, P.-F.; Chen, H.; Zhong, W.; Lin, Y.; Zhang, X.; Chen, Y.-X.; Xie, W.-F. Adenovirus-mediated transfer of siRNA against PAI-1 mRNA ameliorates hepatic fibrosis in rats. J. Hepatol. 2009, 51, 102–113. [Google Scholar] [CrossRef]
- Matsuo, S.; Lepez-Guisa, J.M.; Cai, X.; Okamura, D.M.; Alpers, C.E.; Bumgarner, R.E.; Peters, M.A.; Zhang, G.; Eddy, A.A. Multifunctionality of PAI-1 in fibrogenesis: Evidence from obstructive nephropathy in PAI-1–overexpressing mice. Kidney Int. 2005, 67, 2221–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, R.; Kaji, K.; Namisaki, T.; Moriya, K.; Kawaratani, H.; Kitade, M.; Takaya, H.; Aihara, Y.; Douhara, A.; Asada, K.; et al. Novel oral plasminogen activator inhibitor-1 inhibitor TM5275 attenuates hepatic fibrosis under metabolic syndrome via suppression of activated hepatic stellate cells in rats. Mol. Med. Rep. 2020, 22, 2948–2956. [Google Scholar] [CrossRef]
- Lee, S.-J.; Kim, K.-H.; Park, K.-K. Mechanisms of fibrogenesis in liver cirrhosis: The molecular aspects of epithelial-mesenchymal transition. World J. Hepatol. 2014, 6, 207–216. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, W.; Tang, L.; Chen, W.; Guan, X. Epithelial-mesenchymal transition induced PAI-1 is associated with prognosis of triple-negative breast cancer patients. Gene 2018, 670, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, Y.; Kawami, M.; Ojima, T.; Futatsugi, S.; Yumoto, R.; Takano, M. Role of plasminogen activator inhibitor-1 in methotrexate-induced epithelial-mesenchymal transition in alveolar epithelial A549 cells. Biochem. Biophys. Res. Commun. 2020, 525, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Sun, T.; Lu, F.; Shen, Y.; Zhang, Y.; Zhang, B.; Yu, G.; Li, H.; Hao, J. Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice. Int. J. Mol. Sci. 2022, 23, 1582. [Google Scholar] [CrossRef]
- Lu, X.; Deng, T.; Dong, H.; Han, J.; Yu, Y.; Xiang, D.; Nie, G.; Hu, B. Novel Application of Eupatilin for Effectively Attenuating Cisplatin-Induced Auditory Hair Cell Death via Mitochondrial Apoptosis Pathway. Oxidative Med. Cell. Longev. 2022, 2022, 1090034. [Google Scholar] [CrossRef]
- Liu, H.; Hao, J.; Wu, C.; Liu, G.; Wang, X.; Yu, J.; Liu, Y.; Zhao, H. Eupatilin Alleviates Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammation and Oxidative Stress. Experiment 2019, 25, 8289–8296. [Google Scholar] [CrossRef]
- Song, E.-H.; Chung, K.-S.; Kang, Y.-M.; Lee, J.-H.; Lee, M.; An, H.-J. Eupatilin suppresses the allergic inflammatory response in vitro and in vivo. Phytomedicine 2018, 42, 1–8. [Google Scholar] [CrossRef]
- Lee, C.-J.; Hong, S.-H.; Yoon, M.-J.; Lee, K.-A.; Choi, D.H.; Kwon, H.; Ko, J.-J.; Koo, H.S.; Kang, Y.-J. Eupatilin treatment inhibits transforming growth factor beta-induced endometrial fibrosis in vitro. Clin. Exp. Reprod. Med. 2020, 47, 108–113. [Google Scholar] [CrossRef]
- Park, S.J.; Choi, H.; Kim, J.H.; Kim, C.-S. Antifibrotic effects of eupatilin on TGF-β1-treated human vocal fold fibroblasts. PLoS ONE 2021, 16, e0249041. [Google Scholar] [CrossRef]
- Kim, H.-S.; Yoon, Y.-M.; Meang, M.K.; Park, Y.E.; Lee, J.Y.; Lee, T.H.; Lee, J.E.; Kim, I.-H.; Youn, B.-S. Reversion of in vivo fibrogenesis by novel chromone scaffolds. Ebiomedicine 2019, 39, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Hui, A.Y.; Friedman, S.L. Molecular basis of hepatic fibrosis. Expert Rev. Mol. Med. 2003, 5, 1–23. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flevaris, P.; Vaughan, D. The Role of Plasminogen Activator Inhibitor Type-1 in Fibrosis. Semin. Thromb. Hemost. 2017, 43, 169–177. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Tan, R.; Dai, C.; Li, Y.; Wang, D.; Hao, S.; Kahn, M.; Liu, Y. Plasminogen Activator Inhibitor-1 Is a Transcriptional Target of the Canonical Pathway of Wnt/β-Catenin Signaling. J. Biol. Chem. 2010, 285, 24665–24675. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [Green Version]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yao, L.; Liu, Y.; Pan, Z.; Peng, S.; Wan, G.; Cheng, J.; Wang, J.; Cao, W. Astragaloside IV regulates NF-κB-mediated cellular senescence and apoptosis of hepatic stellate cells to suppress PDGF-BB-induced activation. Exp. Ther. Med. 2019, 18, 3741–3750. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.L.; Pinzani, M. Hepatic fibrosis 2022: Unmet needs and a blueprint for the future. Hepatology 2022, 75, 473–488. [Google Scholar] [CrossRef]
- Wei, H.-T.; Xu, Y.; Tan, X.-Y.; Jing, H.-Y.; Ma, Y.-R. ShenKang Injection Attenuates Renal Fibrosis by Inhibiting EMT and Regulating the Wnt/β-Catenin Pathway. Evidence-Based Complement. Altern. Med. 2022, 2022, 9705948. [Google Scholar] [CrossRef]
- Guo, Y.; Xiao, L.; Sun, L.; Liu, F. Wnt/β-Catenin Signaling: A Promising New Target for Fibrosis Diseases. Physiol. Res. 2012, 61, 337–346. [Google Scholar] [CrossRef]
- Ge, W.-S.; Wang, Y.-J.; Wu, J.-X.; Fan, J.-G.; Chen, Y.-W.; Zhu, L. β-catenin is overexpressed in hepatic fibrosis and blockage of Wnt/β-catenin signaling inhibits hepatic stellate cell activation. Mol. Med. Rep. 2014, 9, 2145–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef] [PubMed]
- Anthony, C.C.; Robbins, D.J.; Ahmed, Y.; Lee, E. Nuclear Regulation of Wnt/β-Catenin Signaling: It’s a Complex Situation. Genes 2020, 11, 886. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Z. Inhibition of PAI-1: A New Anti-thrombotic Approach. Curr. Drug Target-Cardiovascular Hematol. Disord. 2002, 2, 27–42. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Hautekeete, M.L.; Geerts, A. The hepatic stellate (Ito) cell: Its role in human liver disease. Virchows Arch. 1997, 430, 195–207. [Google Scholar] [CrossRef]
- Sato, M.; Suzuki, S.; Senoo, H. Hepatic Stellate Cells: Unique Characteristics in Cell Biology and Phenotype. Cell Struct. Funct. 2003, 28, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, Y.; Heuckeroth, R.O. PAI-1 deficiency reduces liver fibrosis after bile duct ligation in mice through activation of tPA. FEBS Lett. 2007, 581, 3098–3104. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-P.; Li, W.-B.; Wang, W.-L.; Liu, J.; Song, S.-X.; Bai, L.-L.; Hu, Y.-Y.; Yuan, Y.-D.; Zhang, M. siRNA against plasminogen activator inhibitor-1 ameliorates bleomycin-induced lung fibrosis in rats. Acta Pharmacol. Sin. 2012, 33, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.J.; Fogo, A.B. PAI-1 and kidney fibrosis. Front. Biosci. (Landmark Ed.) 2009, 14, 2028–2041. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.K.; Donahue, D.L.; Sandoval-Cooper, M.J.; Castellino, F.J.; Ploplis, V.A. Plasminogen Activator Inhibitor-1 Protects Mice Against Cardiac Fibrosis by Inhibiting Urokinase-type Plasminogen Activator-mediated Plasminogen Activation. Sci. Rep. 2017, 7, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward primer (5′→3′) | Reverse primer (5′→3′) | |
|---|---|---|---|
| Mouse | GAPDH | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
| COL 1α1 | TAAGGGTCCCCAATGGTGAGA | GGGTCCCTCGACTCCTACAT | |
| Fibronectin | AAAAGTTTGTGGGAGTCGTTCT | GGCCCTGTTCTTCCATCCAG | |
| Human | GAPDH | GGCATGGACTGTGGTCATGAG | TGCACCACCAACTGCTTAGC |
| PAI-1 | GCACCACAGACGCGATCTT | ACCTCTGAAAAGTCCACTTGC | |
| β-catenin | CATCTACACAGTTTGATGCTGCT | GCAGTTTTGTCAGTTCAGGGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Liu, Y.; Pan, Z.; Huang, X.; Wang, J.; Cao, W.; Chen, Z. Eupatilin Ameliorates Hepatic Fibrosis and Hepatic Stellate Cell Activation by Suppressing β-catenin/PAI-1 Pathway. Int. J. Mol. Sci. 2023, 24, 5933. https://doi.org/10.3390/ijms24065933
Hu J, Liu Y, Pan Z, Huang X, Wang J, Cao W, Chen Z. Eupatilin Ameliorates Hepatic Fibrosis and Hepatic Stellate Cell Activation by Suppressing β-catenin/PAI-1 Pathway. International Journal of Molecular Sciences. 2023; 24(6):5933. https://doi.org/10.3390/ijms24065933
Chicago/Turabian StyleHu, Jinyuan, Yuanyuan Liu, Zheng Pan, Xuekuan Huang, Jianwei Wang, Wenfu Cao, and Zhiwei Chen. 2023. "Eupatilin Ameliorates Hepatic Fibrosis and Hepatic Stellate Cell Activation by Suppressing β-catenin/PAI-1 Pathway" International Journal of Molecular Sciences 24, no. 6: 5933. https://doi.org/10.3390/ijms24065933