
Role of Terpenophenolics in Modulating Inflammation and Apoptosis in Cardiovascular Diseases: A Review

Abstract
:1. Introduction
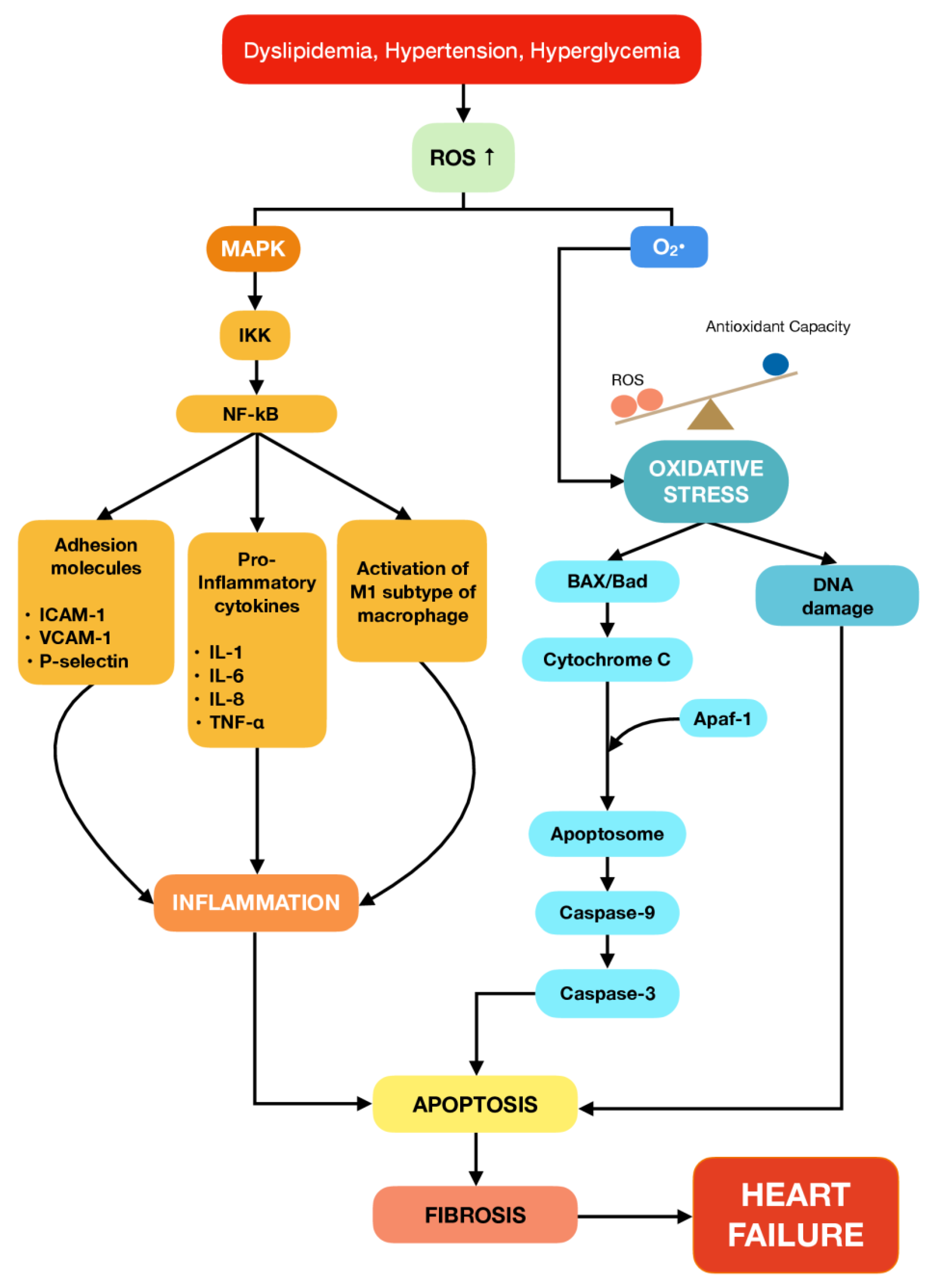
2. The Role of Inflammation and Apoptosis in Cardiovascular Diseases
3. Terpenes and Their Subclasses
4. Role of Terpenophenolic Compounds on Inflammation and Apoptosis in Cardiovascular Diseases
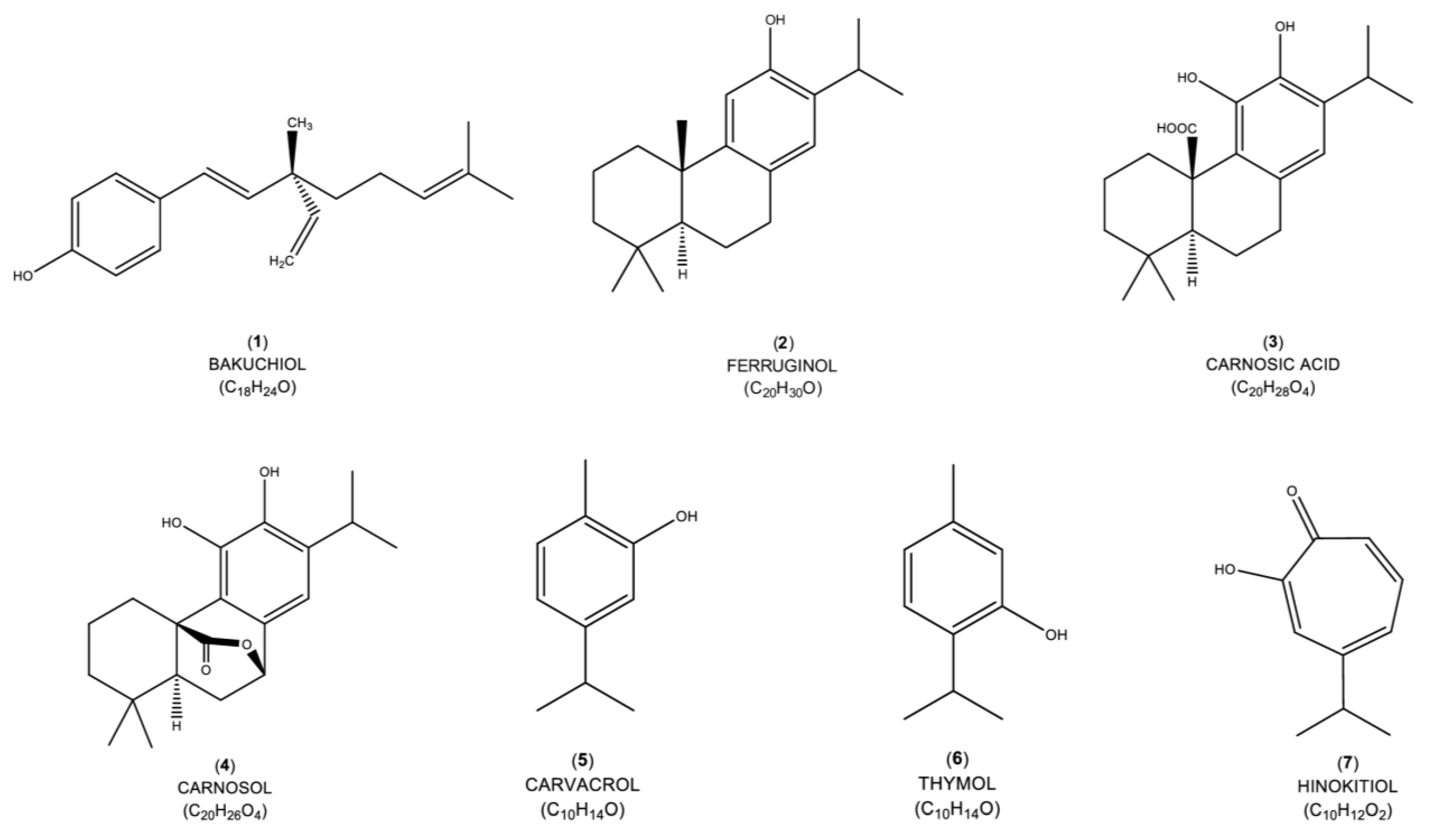
4.1. Bakuchiol
4.2. Ferruginol
4.3. Carnosic Acid
4.4. Carnosol
4.5. Carvacrol
4.6. Thymol
4.7. Hinokitiol
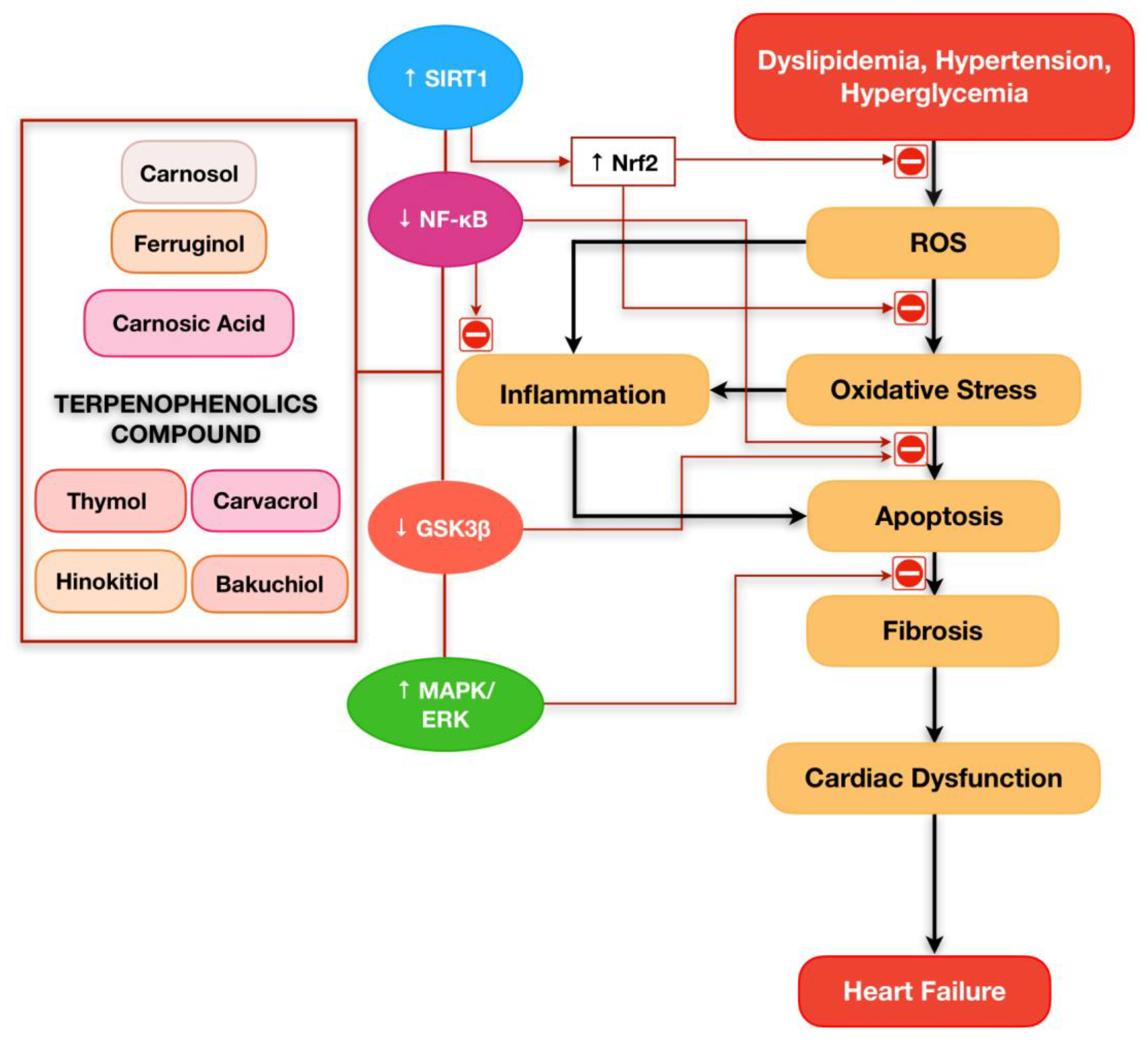
5. Prospective of Terpenophenolics Compound Playing a Critical Role in Cardiovascular Diseases
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | Serine/threonine kinase |
| ANP | Atrial natriuretic peptide |
| aP2 | Adipocyte protein 2 |
| ATG-5 | Autophagy-related genes-5 |
| Bad | Bcl-2 associated death |
| Bax | Bcl2-associated X protein |
| Bcl-2 | B-cell leukemia/lymphoma 2 |
| Bcl-xL | B-cell lymphoma-extra large |
| BNP | Brain natriuretic peptide |
| CAT | Catalase |
| CGTF | Connective tissue growth factor |
| CK | Creatine kinase |
| CK-MB | Creatine kinase-MB |
| Col 1 | Collagen 1 |
| COX-2 | Cyclooxygenase-2 |
| cTnI | Cardiac troponin I |
| cTnT | Cardiac troponin T |
| DOX | Doxorubicin |
| E-selectin | Endothelial-selectin |
| eNOS | Endothelial nitric oxide synthase |
| ERK | Extracellular signal-regulated kinase |
| GSH | Glutathione |
| GSH-Px | Glutathione peroxidase |
| GSK3β | Glycogen synthase kinase 3β |
| GST | Glutathione S-transferases |
| H/R | Hypoxia/Reoxygenation |
| HMVEC | Human microvascular endothelial cell |
| I/R | Ischemia/Reperfusion |
| IL-6 | Interleukin-6 |
| LC3BII | Light chain 3B II |
| LDH | Lactate dehydrogenase |
| LDL | Low-density lipoprotein |
| LPL | Lipoprotein lipase |
| MAP | Mean atrial pressure |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MDA | Malondialdehyde |
| MMP | Matrix metalloproteinase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NF-κB | Nuclear factor kappa B |
| NOX | NAPDH oxidase |
| NRCM | Neonatal rat cardiomyocytes |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| p-p38 | Phosphorylated p38 |
| PPAR-γ | Peroxisome proliferator-activated receptor gamma |
| ROS | Reactive oxygen species |
| Ser9 | Serine 9 |
| sICAM-1 | Soluble intercellular adhesion molecule-1 |
| SIRT1 | Silent information regulator 1 |
| Smad3 | Suppressor of mothers against decapentaplegic 3 |
| SOD | Superoxide dismutase |
| sVCAM-1 | Soluble vascular cell adhesion molecule-1 |
| TAC | Total antioxidant capacity |
| TNF-α | Tumor necrosis factor alpha |
| TUNEL | Terminal deoxynucleotidyl transferase biotin-dUTP nick end labeling |
| α-SMA | Alpha smooth muscle actin |
| β-MHC | Beta myosin heavy chain |
References
- WHO. Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 15 July 2022).
- CDC. Heart Disease Facts. Available online: https://www.cdc.gov/heartdisease/facts.htm (accessed on 1 August 2022).
- Chen, X.; Li, X.; Xu, X.; Li, L.; Liang, N.; Zhang, L.; Lv, J.; Wu, Y.-C.; Yin, H. Ferroptosis and cardiovascular disease: Role of free radical-induced lipid peroxidation. Free Radic. Res. 2021, 55, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Hajar, R. Risk Factors for Coronary Artery Disease: Historical Perspectives. Heart Views 2017, 18, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Majid, H.A.; Hairi, F.; Thangiah, N.; Bulgiba, A.; Su, T.T. Prevalence and determinants of cardiovascular disease risk factors among the residents of urban community housing projects in Malaysia. BMC Public Health 2014, 14 (Suppl. 3), S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, A.P.; Chauhan, J.; Yadav, D.K.; Gangwar, R.; Maurya, V.K. Chapter 11—Nutraceuticals and their impact on human health. In Preparation of Phytopharmaceuticals for the Management of Disorders; Egbuna, C., Mishra, A.P., Goyal, M.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 229–254. [Google Scholar]
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2665–2691. [Google Scholar]
- Buckle, J. Basic Plant Taxonomy, Basic Essential Oil Chemistry, Extraction, Biosynthesis, and Analysis. In Clinical Aromatherapy: Essential Oils in Healthcare, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 37–72. [Google Scholar]
- da Silva, L.B.; Camargo, S.B.; Moraes, R.D.; Medeiros, C.F.; Jesus, A.D.; Evangelista, A.; Villarreal, C.F.; Quintans, L.J.; Silva, D.F. Antihypertensive effect of carvacrol is improved after incorporation in beta-cyclodextrin as a drug delivery system. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.; Mai, Y.; Qiu, X.; Yuan, W.; Li, Y.; Luo, C.; Liu, Y.; Zhang, G.; Zhao, G.; Luo, J.D. Carvacrol Attenuates Diabetic Cardiomyopathy by Modulating the PI3K/AKT/GLUT4 Pathway in Diabetic Mice. Front. Pharm. 2019, 10, 998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wei, J.; Ma, K.T.; Li, C.L.; Mai, Y.P.; Qiu, X.X.; Wei, H.; Hou, N.; Luo, J.D. Carvacrol protects against diabetes-induced hypercontractility in the aorta through activation of the PI3K/Akt pathway. Biomed. Pharm. 2020, 125, 109825. [Google Scholar] [CrossRef]
- Xu, S.; Ilyas, I.; Little, P.J.; Li, H.; Kamato, D.; Zheng, X.; Luo, S.; Li, Z.; Liu, P.; Han, J.; et al. Endothelial Dysfunction in Atherosclerotic Cardiovascular Diseases and Beyond: From Mechanism to Pharmacotherapies. Pharmacol. Rev. 2021, 73, 924. [Google Scholar] [CrossRef]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The vascular endothelium and human diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Little, P.J.; Askew, C.D.; Xu, S.; Kamato, D. Endothelial Dysfunction and Cardiovascular Disease: History and Analysis of the Clinical Utility of the Relationship. Biomedicines 2021, 9, 699. [Google Scholar] [CrossRef]
- Li, Q.; Atochin, D.; Kashiwagi, S.; Earle, J.; Wang, A.; Mandeville, E.; Hayakawa, K.; d’Uscio, L.V.; Lo, E.H.; Katusic, Z.; et al. Deficient eNOS Phosphorylation Is a Mechanism for Diabetic Vascular Dysfunction Contributing to Increased Stroke Size. Stroke 2013, 44, 3183–3188. [Google Scholar] [CrossRef] [Green Version]
- Kostov, K. The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation. Life 2021, 11, 986. [Google Scholar] [CrossRef]
- Soehnlein, O.; Libby, P. Targeting inflammation in atherosclerosis—From experimental insights to the clinic. Nat. Rev. Drug Discov. 2021, 20, 589–610. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Ahmad, W.; Budin, S.B.; Zainalabidin, S. Implication of dietary phenolic acids on inflammation in cardiovascular disease. Rev. Cardiovasc. Med. 2020, 21, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Deng, K.; Zang, Y.; Zhang, Z.; Zhao, B.; Fan, J.; Huang, L. Exploring the regulatory roles of circular RNAs in the pathogenesis of atherosclerosis. Vascul. Pharmacol. 2021, 141, 106898. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.A.; Aminuddin, A.; Anuar, N.N.M.; Mansor, N.I.; Ahmad, M.F.; Saleh, M.S.M.; Mokhtar, M.H.; Ugusman, A. Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells. Life 2022, 12, 1462. [Google Scholar] [CrossRef] [PubMed]
- Mohd Nor, N.A.; Budin, S.B.; Zainalabidin, S.; Jalil, J.; Sapian, S.; Jubaidi, F.F.; Mohamad Anuar, N.N. The Role of Polyphenol in Modulating Associated Genes in Diabetes-Induced Vascular Disorders. Int. J. Mol. Sci. 2022, 23, 6396. [Google Scholar] [CrossRef]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Abdul Hamid, Z.; Mohamad Anuar, N.N.; Jalil, J.; Mohd Nor, N.A.; Budin, S.B. The Role of PKC-MAPK Signalling Pathways in the Development of Hyperglycemia-Induced Cardiovascular Complications. Int. J. Mol. Sci. 2022, 23, 8582. [Google Scholar] [CrossRef]
- Zeng, Y.; Xiong, Y.; Yang, T.; Wang, Y.; Zeng, J.; Zhou, S.; Luo, Y.; Li, L. Icariin and its metabolites as potential protective phytochemicals against cardiovascular disease: From effects to molecular mechanisms. Biomed. Pharmacother. 2022, 147, 112642. [Google Scholar] [CrossRef]
- Sapian, S.; Taib, I.S.; Latip, J.; Katas, H.; Chin, K.Y.; Mohd Nor, N.A.; Jubaidi, F.F.; Budin, S.B. Therapeutic Approach of Flavonoid in Ameliorating Diabetic Cardiomyopathy by Targeting Mitochondrial-Induced Oxidative Stress. Int. J. Mol. Sci. 2021, 22, 1616. [Google Scholar] [CrossRef]
- Si, L.Y.; Ali, S.A.M.; Latip, J.; Fauzi, N.M.; Budin, S.B.; Zainalabidin, S. Roselle is cardioprotective in diet-induced obesity rat model with myocardial infarction. Life Sci. 2017, 191, 157–165. [Google Scholar] [CrossRef]
- Kim, N.-H.; Kang, P.M. Apoptosis in Cardiovascular Diseases: Mechanism and Clinical Implications. Korean Circ. J. 2010, 40, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, W.; Wang, Z.; Xu, T.; Li, D. Anti-apoptotic effects and mechanisms of salvianolic acid A on cardiomyocytes in ischemia-reperfusion injury. Histol. Histopathol. 2019, 34, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Yalamarti, B.; Kang, P.M. Role of caspase-independent apoptosis in cardiovascular diseases. Front. Biosci. 2008, 13, 2495–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinderer, S.; Schenke-Layland, K. Cardiac fibrosis—A short review of causes and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 77–82. [Google Scholar] [CrossRef]
- Gunawardena, G. Isoprene Rule. Available online: https://chem.libretexts.org/@go/page/39398 (accessed on 3 June 2022).
- Perveen, S.; Al-Taweel, A. Terpenes and Terpenoids; BoD–Books on Demand: Norderstedt, Germany, 2018. [Google Scholar]
- Yadav, N.; Yadav, R.; Goyal, A. Chemistry of Terpenoids. Int. J. Pharm. Sci 2014, 27, 272–278. [Google Scholar]
- Santos, M.R.; Moreira, F.V.; Fraga, B.P.; Souza, D.P.D.; Bonjardim, L.R.; Quintans-Junior, L.J. Cardiovascular effects of monoterpenes: A review. Rev. Bras. Farmacogn. 2011, 21, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Ashour, M.; Wink, M.; Gershenzon, J. Biochemistry of Terpenoids: Monoterpenes, Sesquiterpenes and Diterpenes. In Annual Plant Reviews Volume 40: Biochemistry of Plant Secondary Metabolism; Wiley Online Library: Hoboken, NJ, USA, 2010; pp. 258–303. [Google Scholar]
- Harborne, J.B. Phytochemical Methods, 1st ed.; Springer: Dordrecht, The Netherlands, 1988; p. 278. [Google Scholar]
- Morrow, G.W.; Morrow, G.W. 258The Shikimate Pathway: Biosynthesis of Phenolic Products from Shikimic Acid. In Bioorganic Synthesis: An Introduction; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Paduch, R.; Kandefer-Szerszeń, M.; Trytek, M.; Fiedurek, J. Terpenes: Substances useful in human healthcare. Arch. Immunol. Exp. 2007, 55, 315–327. [Google Scholar] [CrossRef]
- Breitmaier, E. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones; Wiley Online Library: Hoboken, NJ, USA, 2006; pp. 1–214. [Google Scholar]
- Wang, G.; Tang, W.; Bidigare, R. Terpenoids as Therapeutic Drugs and Pharmaceutical Agents. In Natural Products: Drug Discovery and Therapeutic Medicine; Humana Press: Totowa, NJ, USA, 2005; Volume 3, pp. 197–227. [Google Scholar]
- Tiwari, G.; Tiwari, R.; Sriwastawa, B.; Bhati, L.; Pandey, S.; Pandey, P.; Bannerjee, S.K. Drug delivery systems: An updated review. Int. J. Pharm. Investig. 2012, 2, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Duchêne, D.; Wouessidjewe, D.; Ponchel, G. Cyclodextrins and carrier systems. J. Control. Release 1999, 62, 263–268. [Google Scholar] [CrossRef]
- Ma, W.; Guo, W.; Shang, F.; Li, Y.; Li, W.; Liu, J.; Ma, C.; Teng, J. Bakuchiol Alleviates Hyperglycemia-Induced Diabetic Cardiomyopathy by Reducing Myocardial Oxidative Stress via Activating the SIRT1/Nrf2 Signaling Pathway. Oxid. Med. Cell Longev. 2020, 2020, 3732718. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, L.; Xiao, L.; Kong, L.; Shi, H.; Tian, X.; Zhao, L. Bakuchiol protects against pathological cardiac hypertrophy by blocking NF-κB signaling pathway. Biosci. Rep. 2018, 38, bsr20181043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, X.; Wang, C.; Li, H.; Wang, L.; Chen, Y.; Feng, J.; Ali Alharbi, S.; Deng, Y. Ameliorative effect of ferruginol on isoprenaline hydrochloride-induced myocardial infarction in rats. Environ. Toxicol. 2021, 36, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cao, J.; Wang, X.; Zhang, Y.; Sun, Q.; Jiang, Y.; Yao, J.; Li, C.; Wang, Y.; Wang, W. Ferruginol Restores SIRT1-PGC-1α-Mediated Mitochondrial Biogenesis and Fatty Acid Oxidation for the Treatment of DOX-Induced Cardiotoxicity. Front. Pharm. 2021, 12, 773834. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Lim, W.; Sung, M.K. Carnosic Acid Modulates Increased Hepatic Lipogenesis and Adipocytes Differentiation in Ovariectomized Mice Fed Normal or High-Fat Diets. Nutrients 2018, 10, 1984. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.J.; Xu, H.J.; Chen, J.J.; Yang, X.; Xiong, J.; Wang, J.; Cheng, F. Carnosic acid protects against pressure overload-induced cardiac remodelling by inhibiting the AKT/GSK3 beta/NOX4 signalling pathway. Exp. Ther. Med. 2020, 20, 3709–3719. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Yang, J.J.; Zhang, H.S. Carvedilol (CAR) combined with carnosic acid (CAA) attenuates doxorubicin-induced cardiotoxicity by suppressing excessive oxidative stress, inflammation, apoptosis and autophagy. Biomed. Pharmacother. 2019, 109, 71–83. [Google Scholar] [CrossRef]
- Baradaran Rahimi, V.; Momeni-Moghaddam, M.A.; Chini, M.G.; Saviano, A.; Maione, F.; Bifulco, G.; Rahmanian-Devin, P.; Jebalbarezy, A.; Askari, V.R. Carnosol Attenuates LPS-Induced Inflammation of Cardiomyoblasts by Inhibiting NF-κB: A Mechanistic in Vitro and in Silico Study. Evid.-Based Complement. Altern. Med. 2022, 2022, 7969422. [Google Scholar] [CrossRef]
- Ou, S.; Lv, J.; Peng, L.; Zhao, J.; Chi, L. Carnosol promotes endothelial differentiation under H2O2-induced oxidative stress. Arch. Biol. Sci. 2017, 69, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ba, L.; Huang, W.; Liu, Y.; Pan, H.; Mingyao, E.; Shi, P.; Wang, Y.; Li, S.; Qi, H.; et al. Role of carvacrol in cardioprotection against myocardial ischemia/reperfusion injury in rats through activation of MAPK/ERK and Akt/eNOS signaling pathways. Eur. J. Pharm. 2017, 796, 90–100. [Google Scholar] [CrossRef]
- Sadeghzadeh, S.; Hejazian, S.H.; Jamhiri, M.; Hafizibarjin, Z.; Sadeghzadeh, S.; Safari, F. The effect of carvacrol on transcription levels of Bcl-2 family proteins in hypertrophied heart of rats. Physiol. Pharmacol. 2018, 22, 54–62. [Google Scholar]
- Bayatmakoo, R.; Rashtchizadeh, N.; Yaghmaei, P.; Farhoudi, M.; Karimi, P. Thymol decreases apoptosis and carotid inflammation induced by hypercholesterolemia through a discount in oxidative stress. Crescent J. Med. Biol. Sci. 2017, 4, 186–193. [Google Scholar]
- El-Marasy, S.A.; El Awdan, S.A.; Hassan, A.; Abdallah, H.M.I. Cardioprotective effect of thymol against adrenaline-induced myocardial injury in rats. Heliyon 2020, 6, e04431. [Google Scholar] [CrossRef] [PubMed]
- Shih, M.F.; Pan, K.H.; Liu, C.C.; Shen, C.R.; Cherng, J.Y. Treatment of beta-thujaplicin counteracts di(2-ethylhexyl)phthalate (DEHP)-exposed vascular smooth muscle activation, inflammation and atherosclerosis progression. Regul. Toxicol. Pharmacol. 2018, 92, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Liang, S.; Cai, Q.; Liu, J.; Jin, L.; Yang, Z.; Chen, X. Hinokitiol Protects Cardiomyocyte from Oxidative Damage by Inhibiting GSK3β-Mediated Autophagy. Oxid. Med. Cell Longev. 2022, 2022, 2700000. [Google Scholar] [CrossRef]
- Khushboo, P.S.; Jadhav, V.M.; Kadam, V.J.; Sathe, N.S. Psoralea corylifolia Linn.-”Kushtanashini”. Pharm. Rev. 2010, 4, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.H.; Ha, T.Y.; Ahn, J.; Kim, S. Estrogenic activities of Psoralea corylifolia L. seed extracts and main constituents. Phytomedicine 2011, 18, 425–430. [Google Scholar] [CrossRef]
- Ulubelen, A.; Topçu, G. Chemical and biological investigations of Salvia species growing in Turkey. In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 1997; Volume 20, pp. 659–718. [Google Scholar]
- Ulubelen, A. Cardioactive and antibacterial terpenoids from some Salvia species. Phytochemistry 2003, 64, 395–399. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.d.P.; Herrero, M. Rosemary (Rosmarinus officinalis) as a functional ingredient: Recent scientific evidence. Curr. Opin. Food Sci. 2017, 14, 13–19. [Google Scholar] [CrossRef]
- Liu, P.; Dong, J. Protective effects of carnosic acid against mitochondria-mediated injury in H9c2 cardiomyocytes induced by hypoxia/reoxygenation. Exp. Med. 2017, 14, 5629–5634. [Google Scholar] [CrossRef]
- Ghasemzadeh Rahbardar, M.; Hosseinzadeh, H. Therapeutic effects of rosemary (Rosmarinus officinalis L.) and its active constituents on nervous system disorders. Iran. J. Basic Med. Sci. 2020, 23, 1100–1112. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Q.; Hou, N.; Li, J.; Liu, M.; Peng, S.; Zhang, Y.; Luo, Y.; Zhao, B.; Wang, S.; et al. Carnosol as a Nrf2 activator improves endothelial barrier function through antioxidative mechanisms. Int. J. Mol. Sci. 2019, 20, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedan, A.A.; Dezfoulian, O.; Alirezaei, M. Satureja khuzistanica Jamzad essential oil prevents doxorubicin-induced apoptosis via extrinsic and intrinsic mitochondrial pathways. Res. Pharm. Sci. 2020, 15, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, F.; Khazaei, M.; Boskabady, M.H. Cardiovascular Effects of Nigella sativa L. and its Constituents. Indian J. Pharm. Sci. 2018, 80, 971–983. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Gutiérrez-Praena, D.; Pichardo, S.; Moreno, F.J.; Bermúdez, J.M.; Aucejo, S.; Cameán, A.M. Cytotoxicity and morphological effects induced by carvacrol and thymol on the human cell line Caco-2. Food Chem. Toxicol. 2014, 64, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Agus, H.H. Chapter 4—Terpene toxicity and oxidative stress. In Toxicology; Patel, V.B., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 33–42. [Google Scholar]
- El-Sayed, S.M.; Mansour, A.M.; Abdul-Hameed, M.S. Thymol and Carvacrol Prevent Doxorubicin-Induced Cardiotoxicity by Abrogation of Oxidative Stress, Inflammation, and Apoptosis in Rats. J. Biochem. Mol. Toxicol. 2016, 30, 37–44. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Galaly, S.R.; Raslan, M.; Mostafa, M. Thyme oil and thymol abrogate doxorubicin-induced nephrotoxicity and cardiotoxicity in Wistar rats via repression of oxidative stress and enhancement of antioxidant defense mechanisms. Biocell 2020, 44, 41–53. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Javed, H.; Al Taee, H.; Azimullah, S.; Ojha, S.K. Pharmacological Properties and Molecular Mechanisms of Thymol: Prospects for Its Therapeutic Potential and Pharmaceutical Development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.M.; Chao, T.Y.; Chang, W.C.; Chang, M.J.; Lee, M.F. Thymol reduces oxidative stress, aortic intimal thickening, and inflammation-related gene expression in hyperlipidemic rabbits. J. Food Drug Anal. 2016, 24, 556–563. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, J.; Takashima, Y.; Ishiguri, F.; Sanpe, H.; Ohshima, J.; Iizuka, K.; Yokota, S. Differences in β-thujaplicin content of wood between plantation- and naturally grown Thujopsis dolabrata var. hondae (hinokiasunaro) trees in Shimokita Peninsula, Aomori, Japan. J. Wood Sci. 2019, 65, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Jayakumar, T.; Hsu, W.-H.; Yen, T.-L.; Luo, J.-Y.; Kuo, Y.-C.; Fong, T.-H.; Sheu, J.-R. Hinokitiol, a Natural Tropolone Derivative, Offers Neuroprotection from Thromboembolic Stroke In Vivo. Evid. Based Complement. Altern. Med. 2013, 2013, 840487. [Google Scholar] [CrossRef] [Green Version]
- University of California, D. Comparison of the Cosmetic Effects of Bakuchiol and Retinol. Available online: https://ClinicalTrials.gov/show/NCT03112863 (accessed on 5 February 2023).
- Instituto Venezolano de Investigaciones, C. Study to Verify the Effectiveness and Safety of Isothymol or Carvacrol Compound against SARS-CoV-2 in COVID-19 Patients. Available online: https://ClinicalTrials.gov/show/NCT05445089 (accessed on 5 February 2023).
- Centro Universitario de Ciencias de la Salud, M. Thymol on Netrin-1 on Obese Patients. Available online: https://ClinicalTrials.gov/show/NCT05427721 (accessed on 5 February 2023).




{kind=link}
{kind=link}
{kind=link}
| Terpene | Number of Isoprene Units | Number of Carbon Atoms |
|---|---|---|
| Hemiterpenes, C5H8 | 1 | 5 |
| Monoterpenes, C10H16 | 2 | 10 |
| Sesquiterpenes, C15H24 | 3 | 15 |
| Diterpenes, C20H32 | 4 | 20 |
| Sesterterpenes, C25H40 | 5 | 25 |
| Triterpenes, C30H48 | 6 | 30 |
| Tetraterpenes, C40H64 | 8 | 40 |
| Polyterpenes | Many | <40 |
| Molecular Structure | Name |
|---|---|
| Chain, no ring | acyclic |
| One ring | cyclic |
| Two rings | bicyclic |
| Three rings | tricyclic |
| Four rings | tetracyclic |
| Terpenophenolics | Terpenophenolics Subclasses | Study Design | Dose | Findings | Conclusion | References |
|---|---|---|---|---|---|---|
| Bakuchiol | Meroterpenoids | In vivo & in vitro; C57BL6 male mice (20–25 g) & H9c2 cells | 60 mg/kg/day; 2, 5, 10 μM |
| Protective effect of bakuchiol in limiting the synthesis fibrosis, preventing oxidative damage and cell death in diabetic myocardium may be via the SIRT1 and Nrf2 signalling pathway. | [42] |
| In vivo & in vitro; C57BL/6J mice & NRCM cells | 10 mg/kg/day; 1, 5, 10 μM |
| Bakuchiol exerts antihypertrophy effects by modulating synthesis of fibrosis and inflammatory responses may be via the NF-κB pathway. | [43] | ||
| Ferruginol | Diterpenoids | In vivo; Wistar rats | 50 mg/kg/day |
| Cardioprotective effect of ferruginol against myocardial infarction via modulation of inflammatory response and upregulation of antioxidant enzymes. | [44] |
| In vivo & in vitro; C57BL/6 mice (20 ± 2) & H9c2 cells | 20 mg/kg/day; 0.1 μM |
| The cardioprotective action of ferruginol is proven by preserving the mitochondrial from the production of ROS, limiting damage to heart function and attenuating the apoptotic process, possibly via the SIRT1 pathway that mediates mitochondrial biogenesis and fatty acid oxidation. | [45] | ||
| Carnosic Acid | Diterpenoids | In vivo; C57BL/6 mice | 0.02% |
| Carnosic acid exhibit antiobesity effect by improving hormone homeostasis and reduced genes expression of liver lipogenesis possibly via the PPAR-γ pathway. | [46] |
| In vivo; C57BL/6 mice | 50 mg/kg |
| Cardioprotective of carnosic acid against myocardial remodelling by modulation oxidative stress and apoptosis via the AKT/GSK3β/NOX 4 signalling pathway. | [47] | ||
| In vivo & in vitro; C57BL/6 mice & H9c2 cells | 5 mg/kg; 10 μM |
| Carnosic acid protects the heart against toxicity by suppression of oxidative damage, inflammation, apoptosis, and autophagy. | [48] | ||
| Carnosol | Diterpenoids | In vitro; H9c2 cells | 5, 10, 20 μM |
| The protective effect of carnosol against inflammation in the cardiomyoblasts may be via the NF-κB signalling pathway. | [49] |
| In vitro; MAPC cells | 0.2 μM |
| Carnosol promotes vascular health by regulating redox status and downregulating inflammation and apoptosis. | [50] | ||
| Carvacrol | Monoterpenoids | In vivo & in vitro; Wistar rats & NRCM cells | 25, 50, 100 mg/kg; 0.6 mM |
| Carvacrol was found to possess cardioprotective properties, which may be related to its antioxidant and antiapoptotic properties in myocardial I/R injury through activation of MAPK/ERK and Akt-eNOS signalling pathways. | [51] |
| In vivo; Wistar rats | 5, 10, 25, 50 mg/kg |
| Protective effect of carvacrol against myocardial hypertrophy by improving blood pressure and inhibiting apoptosis via regulation of the Bcl-2 family protein. | [52] | ||
| Thymol | Monoterpenoids | In vivo; Wistar rats | 24 mg/kg |
| Thymol preserves carotid tissue by reducing apoptosis and inflammation, which may be a result of its direct antioxidant properties. | [53] |
| In vivo; Albino Wistar rats | 15, 30, 60 mg/kg |
| Thymol has been revealed to have a cardioprotective effect against myocardial infarction via modulating oxidative stress, inflammation, and apoptosis. | [54] | ||
| Hinokitiol | Monoterpenoids | In vitro; SEVC4-10 cells and A7r5 cells | 4, 12 μM |
| Hinokitiol exerts a protective effect against atherosclerosis through modulating cell adhesion molecules and members of the matrix metalloproteinase (MMP) family. | [55] |
| In vitro; AC16 cells | 20 μM |
| Hinokitiol protects cardiomyocytes from oxidative damage by regulating apoptosis and autophagy, probably through the GSK3β signalling pathway. | [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Ghani, M.A.; Ugusman, A.; Latip, J.; Zainalabidin, S. Role of Terpenophenolics in Modulating Inflammation and Apoptosis in Cardiovascular Diseases: A Review. Int. J. Mol. Sci. 2023, 24, 5339. https://doi.org/10.3390/ijms24065339
Abdul Ghani MA, Ugusman A, Latip J, Zainalabidin S. Role of Terpenophenolics in Modulating Inflammation and Apoptosis in Cardiovascular Diseases: A Review. International Journal of Molecular Sciences. 2023; 24(6):5339. https://doi.org/10.3390/ijms24065339
Chicago/Turabian StyleAbdul Ghani, Muhamad Adib, Azizah Ugusman, Jalifah Latip, and Satirah Zainalabidin. 2023. "Role of Terpenophenolics in Modulating Inflammation and Apoptosis in Cardiovascular Diseases: A Review" International Journal of Molecular Sciences 24, no. 6: 5339. https://doi.org/10.3390/ijms24065339