

A Data-Mining Approach to Identify NF-kB-Responsive microRNAs in Tissues Involved in Inflammatory Processes: Potential Relevance in Age-Related Diseases
 , ,
, ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Putative NF-kB Responsive miRNAs
2.2. Genomic Features of Putative NF-kB Responsive miRNAs
2.3. Characterization of the Interplay Linking NF-kB, miRNAs, and Their Host Genes
- NFYC (Nuclear transcription factor Y subunit gamma) is a component of the sequence-specific heterotrimeric TF (NF-Y) which specifically recognizes a 5′- CCAAT-3′ box motif found in the promoters of its target genes. NF-Y can function as both an activator and a repressor, depending on its interacting cofactors [39];
- ZRANB2 (Zinc finger Ran-binding domain-containing protein 2) is a splicing factor required for alternative splicing of TRA2B/SFRS10 transcripts. May interfere with constitutive 5′-splice site selection [40];
- IARS2 (Isoleucine-tRNA ligase, mitochondrial) is a nuclear gene encoding mitochondrial isoleucyl-tRNA synthetase on which depends the translation of mitochondrial-encoded proteins [41];
- SMC4 (Structural maintenance of chromosomes protein 4) is the central component of the condensin complex, a complex required for the conversion of interphase chromatin into mitotic-like condense chromosomes [42];
- MCM7 (DNA replication licensing factor MCM7) acts as a component of the MCM2-7 complex (MCM complex) which is the replicative helicase essential for “once per cell cycle” DNA replication initiation and elongation in eukaryotic cells. It is the core component of CDC45-MCM-GINS (CMG) helicase, the molecular machine that unwinds template DNA during replication, and around which the replisome is built [43,44,45,46,47,48];
- NR6A1 (Nuclear receptor subfamily 6 group A member 1) is an orphan nuclear receptor that binds to a response element containing the sequence 5′-TCAAGGTCA-3′. By similarity, it may be involved in the regulation of gene expression in germ cell development during gametogenesis. It is involved in regulating embryonic stem cell differentiation, reproduction, and neuronal differentiation [49];
- TENM4 (Teneurin-4) is involved in neural development, regulating the establishment of proper connectivity within the nervous system. It plays a role in the establishment of the anterior–posterior axis during gastrulation. Moreover, it regulates the differentiation and cellular process formation of oligodendrocytes and myelination of small-diameter axons in the central nervous system (CNS) [50];
- WWP2 (NEDD4-like E3 ubiquitin-protein ligase WWP2) plays an important role in protein ubiquitination and inhibits activation-induced T cell death by catalyzing EGR2 ubiquitination [58]. In human embryonic stem cells, WWP2 promotes the degradation of TF OCT4, which not only plays an essential role in maintaining the pluripotent and self-renewing state of embryonic stem cells but also acts as a cell fate determinant through a gene dosage effect [55];
- HOXB3 (Homeobox protein Hox-B3) is a sequence-specific TF that is part of a developmental regulatory system that provides cells with specific positional identities on the anterior–posterior axis. Therefore, it may regulate gene expression, morphogenesis, and differentiation [59];
- SREBF1 (Sterol regulatory element-binding protein 1) is a precursor of the TF form (Processed sterol regulatory element-binding protein 1), which is embedded in the endoplasmic reticulum membrane [60]. Its processed form is a key TF that regulates the expression of genes involved in cholesterol biosynthesis and lipid homeostasis [60,61,62];
- PANK2 (Pantothenate kinase 2) is the mitochondrial isoform that catalyzes the phosphorylation of pantothenate to generate 4′-phosphopantothenate in the first and rate-determining step of coenzyme A (CoA) synthesis [63,64,65,66]. It is required for angiogenic activity of the umbilical vein of endothelial cells (HUVEC) [67].
2.4. Pathways Targeted by the 68 Putative NF-kB Responsive miRNAs
2.5. The 68 Putative NF-kB Responsive miRNAs and Previously Identified Inflammamirs
2.6. mRNAs Targeted by the 68 Putative NF-kB Responsive miRNAs Belonging to Pathways Involved in Aging Process and/or Age-Related Diseases
3. Discussion
- “Intronic” promoters can explain cases of poor correlation between host gene and miRNA expression, functioning as a real alternative promoter [23]. As shown in Figure S1, the expression levels of NF-kB-miRNAs modulated by both “host gene” and “intronic” promoters (i.e., miR-16, miR-103, miR-186, and miR-33b) or by both “host gene” and “intergenic promoters” (i.e., miR-194) are not correlated with the expression levels of their host gene, whereas most of the miRNAs that share the host gene promoters are characterized by directly (e.g., miR-15b) or inversely (e.g., miR-30c, miR-616, and miR-93) correlated transcription levels.
- “Intronic” promoters are expressed in a tissue-specific manner, but “host gene” promoters are considered primarily for housekeeping gene regulation [23]. Housekeeping genes are required for the maintenance of essential functions of any cell type, so they are expected to be constitutively expressed in all cells and at any development stage [100]. Among the NF-kB-miRNA host genes, COPZ1, NFYC, and ZRANB2 have been cataloged as housekeeping genes (Table 2).
- “Intronic” promoters are mainly triggered by tissue-specific master regulator TFs, instead of TFs of “host gene” promoters, which broadly overlap with those of protein coding genes and can be considered mainly for housekeeping (“intergenic” promoters are regulated by a combination of intronic-specific and host-gene specific TFs). This suggests a different evolutionary mechanism [23]. In this study, the expression levels of the three housekeeping host genes (COPZ1, ZRANB2, and NFYC) and their miRNAs (miR-148b-3p, miR-186-5p, and, lastly, miR-30c-5p and miR-30e-5p, respectively) are mainly inversely correlated or not showing clear correlation trends (Figure S1).
- “Intronic” miRNA promoters are less evolutionarily conserved than either “intergenic” or “host gene” promoters [23].
- Conversely, evolutionarily conserved miRNAs are more likely to be regulated by an “intronic” promoter [23].
- Moreover, those intragenic miRNAs that share the promoters of the host gene interact with their own host genes (miR-16-2::MSC4; miR-106b::MCM7, miR-181b-2::NR6A1, miR-708::TENM4, miR-148b::COPZ1, and miR-10a::HOXB3), but also with the other functionally related host genes, creating a complex regulatory mechanism (Figure 2). NFKB1, REL, miR-16-5p, miR-103a-3p, and NR6A1 are the most important hub nodes in the network, whereas miR-10a-5p connects the hub nodes identified by NFKB1, NR6A1, and HOXB3, and miR-30e-5p connects REL, NR6A1, and ZRAMB2 hubs. Interestingly, in the network, it is possible to identify a clear TF-miRNA feed-forward loop involving DDIT3, miR-16-5p, and NFYC. In a TF-miRNA feed-forward loop, TF and miRNA co-regulate the target genes: in a “coherent” feed-forward loop, the TF and miRNA have the same effects on their common targets, whereas, in an “incoherent” feed-forward loop, the TF and miRNA carry out opposing effects, which precisely fine-tune gene expressions to minimize noise and maintain stability [68,101]. TF-miRNA feed-forward loops have a specific function in noise buffering effects, which can minimize the response of stochastic signaling noise to maintain steady-state target levels [102,103]. Disruption of feed-forward loops could lead to serious dysregulations at the origin of diseases and cancers, e.g., interference in the NF-kB/miR-19/CYLD loop can induce T cell leukemogenesis [103,104]. Therefore, investigating the regulatory motifs among DDIT3, 16-5p, and NFYC could provide valuable insights to dissect the molecular mechanisms underlying biological processes and diseases triggered by NF-kB constitutive activation.
Limitation of the Study
4. Methods
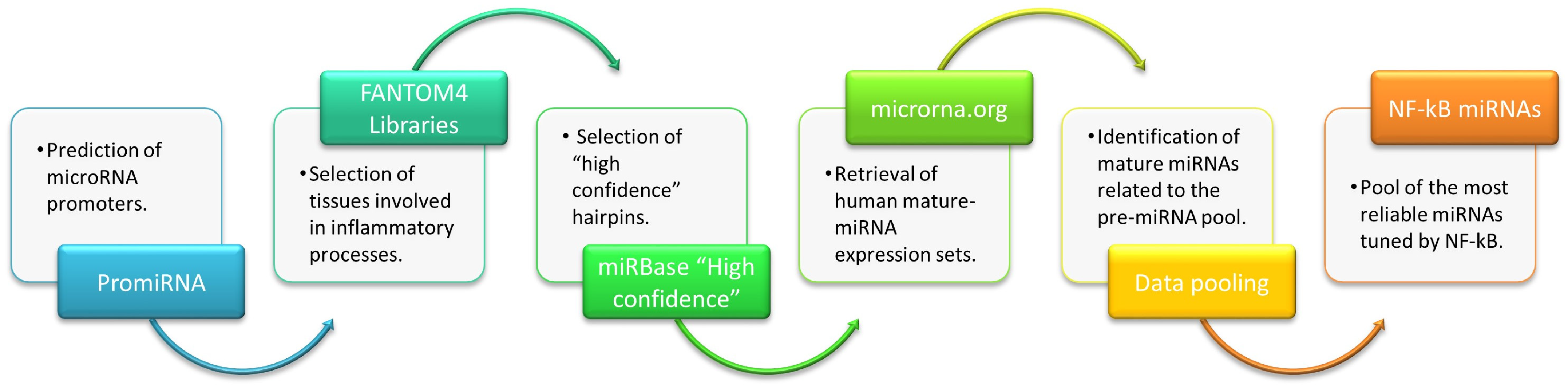
4.1. Data Mining Process
4.1.1. Database Selection
- PROmiRNA provides an interesting approach for miRNA promoter annotation based on a semi-supervised statistical model trained on deepCAGE data and sequence features [23]. It was used to identify all human miRNAs potentially modulated by NF-kB, i.e., “NF-kappaB”, “NFKB1”, “REL”, and “RELA”.
- FANTOM4 libraries, generated by the FANTOM4 project [26], collects a wide range of genome-scale data from several tissues. The analysis of FANTOM4 libraries retrieved those miRNAs showing “expression at the promoter level” in different human tissues. The following libraries from healthy tissues were selected: “blood”, “bone marrow”, “immune system cells”, “liver”, “monocytic-cells”, “T cells”, and “T cells 2”.
- miRBase database is the public repository for all published miRNA sequences and associated annotations [27,32,123,124,125,126]. The “High confidence human hairpins” dataset [27] was downloaded to identify all human miRBase entries with high confidence levels assessed using the deep sequencing data sets collated in miRbase (The original dataset is provided in Supplementary File S1, and it has been downloaded from this link: https://www.mirbase.org/blog/2014/07/high-confidence-mirna-set-available-for-mirbase-21/, accessed on 10 January 2023).
- Finally, microRNA.org [28], a comprehensive resource of miRNA target predictions and expression profiles, was searched to extract the “Human Expression dataset”, meaning the mature miRNA expression profiles in various tissues as presented by Landgraf et al. [127]. Expressed miRNAs from the following healthy tissues were selected (library names for each sample type are indicated in brackets): liver (hsa_Liver), pluripotent hematopoietic stem cell (hsa_HSC-CD34), B cells from peripheral blood (hsa_B-cell-CD19, hsa_B-cell-CD19-2, hsa_B-cell-CD19-pool), T-lymphocytes (hsa_T-cell-CD4, hsa_T-cell-CD4-2, hsa_T-cell-CD4-effector, hsa_T-cell-CD4-memory, hsa_T-cell-CD4-naïve, hsa_T-cell-CD8, hsa_T-cell-CD8-2→hsa_T-cell-CD8-naïve), NK cells (hsa_NK-CD56), monocytes (hsa_Monocytes-CD14), granulocytes (hsa_Granulocytes-CD15), and Dendritic cells (hsa_DC-unstim). Libraries from cell lines, tumor samples, genetic disorders, and so on, have been discharged. (The original dataset is provided in Supplementary File S2, and it has been downloaded from this link: http://www.microrna.org/microrna/getDownloads.do, accessed on 10 January 2023).
4.1.2. Data Extraction and Integration
4.1.3. Data Cleaning and Transformation
4.1.4. Assessment of the Results
4.2. Bioinformatic Evaluations
4.2.1. Evaluation of miRNA-Host Gene-Transcription Factor Interactions
4.2.2. Ingenuity Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K. SIRT1 longevity factor suppresses NF-kappaB-driven immune responses: Regulation of aging via NF-kappaB acetylation? Bioessays 2008, 30, 939–942. [Google Scholar] [CrossRef]
- Salminen, A.; Huuskonen, J.; Ojala, J.; Kauppinen, A.; Kaarniranta, K.; Suuronen, T. Activation of innate immunity system during aging: NF-kB signaling is the molecular culprit of inflamm-aging. Ageing Res. Rev. 2008, 7, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, Y.; Wang, W.; Bai, Y.; Jia, H.; Yuan, Z.; Yang, Z. Role and mechanisms of the NF-kB signaling pathway in various developmental processes. Biomed. Pharm. 2022, 153, 113513. [Google Scholar] [CrossRef] [PubMed]
- Dorrington, M.G.; Fraser, I.D.C. NF-kappaB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Prattichizzo, F.; Giuliani, A.; Matacchione, G.; Rippo, M.R.; Sabbatinelli, J.; Bonafe, M. miR-21 and miR-146a: The microRNAs of inflammaging and age-related diseases. Ageing Res. Rev. 2021, 70, 101374. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Adler, A.S.; Sinha, S.; Kawahara, T.L.; Zhang, J.Y.; Segal, E.; Chang, H.Y. Motif module map reveals enforcement of aging by continual NF-kappaB activity. Genes Dev. 2007, 21, 3244–3257. [Google Scholar] [CrossRef] [Green Version]
- Tilstra, J.S.; Clauson, C.L.; Niedernhofer, L.J.; Robbins, P.D. NF-kappaB in Aging and Disease. Aging Dis. 2011, 2, 449–465. [Google Scholar]
- Bernal-Lopez, M.R.; Garrido-Sanchez, L.; Gomez-Carrillo, V.; Gallego-Perales, J.L.; Llorente-Cortes, V.; Calleja, F.; Gomez-Huelgas, R.; Badimon, L.; Tinahones, F.J. Antioxidized LDL antibodies are associated with different metabolic pathways in patients with atherosclerotic plaque and type 2 diabetes. Diabetes Care 2013, 36, 1006–1011. [Google Scholar] [CrossRef] [Green Version]
- Bektas, A.; Zhang, Y.; Lehmann, E.; Wood, W.H., 3rd; Becker, K.G.; Madara, K.; Ferrucci, L.; Sen, R. Age-associated changes in basal NF-kappaB function in human CD4+ T lymphocytes via dysregulation of PI3 kinase. Aging 2014, 6, 957–974. [Google Scholar] [CrossRef]
- Rothschild, D.E.; McDaniel, D.K.; Ringel-Scaia, V.M.; Allen, I.C. Modulating inflammation through the negative regulation of NF-kappaB signaling. J. Leukoc. Biol. 2018, 103, 1131–1150. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Rana, R.; Chhabra, A.; Jaiswal, A.; Rani, V. miRNA-transcription factor interactions: A combinatorial regulation of gene expression. Mol. Genet. Genom. 2013, 288, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Iwama, H. Coordinated networks of microRNAs and transcription factors with evolutionary perspectives. Adv. Exp. Med. Biol. 2013, 774, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Elizarova, A.; Ozturk, M.; Guler, R.; Medvedeva, Y.A. MIREyA: A computational approach to detect miRNA-directed gene activation. F1000Research 2021, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, G.; Teng, Z.Q.; Wingo, T.S. Editorial: Non-Coding RNAs and Human Diseases. Front. Genet. 2020, 11, 523. [Google Scholar] [CrossRef]
- Peng, Y. Non-coding RNAs in human cancer. Semin. Cancer Biol. 2021, 75, 1–2. [Google Scholar] [CrossRef]
- Akhtar, M.M.; Micolucci, L.; Islam, M.S.; Olivieri, F.; Procopio, A.D. Bioinformatic tools for microRNA dissection. Nucleic Acids Res. 2016, 44, 24–44. [Google Scholar] [CrossRef] [Green Version]
- Lukiw, W.J. NF-kappaB-regulated, proinflammatory miRNAs in Alzheimer’s disease. Alzheimers Res. Ther. 2012, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.L.; Huang, Y.; Wang, G.; Chen, S.D. The potential role of microRNA-146 in Alzheimer’s disease: Biomarker or therapeutic target? Med. Hypotheses 2012, 78, 398–401. [Google Scholar] [CrossRef]
- Yang, M.; Ye, L.; Wang, B.; Gao, J.; Liu, R.; Hong, J.; Wang, W.; Gu, W.; Ning, G. Decreased miR-146 expression in peripheral blood mononuclear cells is correlated with ongoing islet autoimmunity in type 1 diabetes patients 1miR-146. J. Diabetes 2015, 7, 158–165. [Google Scholar] [CrossRef]
- Markopoulos, G.S.; Roupakia, E.; Tokamani, M.; Alabasi, G.; Sandaltzopoulos, R.; Marcu, K.B.; Kolettas, E. Roles of NF-kappaB Signaling in the Regulation of miRNAs Impacting on Inflammation in Cancer. Biomedicines 2018, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Wang, C.C.; Guan, N.N. Computational Models in Non-Coding RNA and Human Disease. Int. J. Mol. Sci. 2020, 21, 1557. [Google Scholar] [CrossRef] [Green Version]
- Marsico, A.; Huska, M.R.; Lasserre, J.; Hu, H.; Vucicevic, D.; Musahl, A.; Orom, U.; Vingron, M. PROmiRNA: A new miRNA promoter recognition method uncovers the complex regulation of intronic miRNAs. Genome Biol. 2013, 14, R84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Ding, J.; Yang, J.; Guo, X.; Zheng, Y. MicroRNA Roles in the Nuclear Factor Kappa B Signaling Pathway in Cancer. Front. Immunol. 2018, 9, 546. [Google Scholar] [CrossRef] [Green Version]
- Amini-Farsani, Z.; Yadollahi-Farsani, M.; Arab, S.; Forouzanfar, F.; Yadollahi, M.; Asgharzade, S. Prediction and analysis of microRNAs involved in COVID-19 inflammatory processes associated with the NF-kB and JAK/STAT signaling pathways. Int. Immunopharmacol. 2021, 100, 108071. [Google Scholar] [CrossRef] [PubMed]
- Kawaji, H.; Severin, J.; Lizio, M.; Waterhouse, A.; Katayama, S.; Irvine, K.M.; Hume, D.A.; Forrest, A.R.; Suzuki, H.; Carninci, P.; et al. The FANTOM web resource: From mammalian transcriptional landscape to its dynamic regulation. Genome Biol. 2009, 10, R40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [Green Version]
- Thomas-Chollier, M.; Hufton, A.; Heinig, M.; O’Keeffe, S.; Masri, N.E.; Roider, H.G.; Manke, T.; Vingron, M. Transcription factor binding predictions using TRAP for the analysis of ChIP-seq data and regulatory SNPs. Nat. Protoc. 2011, 6, 1860–1869. [Google Scholar] [CrossRef]
- Portales-Casamar, E.; Thongjuea, S.; Kwon, A.T.; Arenillas, D.; Zhao, X.; Valen, E.; Yusuf, D.; Lenhard, B.; Wasserman, W.W.; Sandelin, A. JASPAR 2010: The greatly expanded open-access database of transcription factor binding profiles. Nucleic Acids Res. 2010, 38, D105–D110. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Hui, J.H.; Marco, A.; Ronshaugen, M. MicroRNA evolution by arm switching. EMBO Rep. 2011, 12, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Schanen, B.C.; Li, X. Transcriptional regulation of mammalian miRNA genes. Genomics 2011, 97, 1–6. [Google Scholar] [CrossRef]
- Davis, B.N.; Hata, A. Regulation of MicroRNA Biogenesis: A miRiad of mechanisms. Cell Commun. Signal. 2009, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozsolak, F.; Poling, L.L.; Wang, Z.; Liu, H.; Liu, X.S.; Roeder, R.G.; Zhang, X.; Song, J.S.; Fisher, D.E. Chromatin structure analyses identify miRNA promoters. Genes Dev. 2008, 22, 3172–3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteys, A.M.; Spengler, R.M.; Wan, J.; Tecedor, L.; Lennox, K.A.; Xing, Y.; Davidson, B.L. Structure and activity of putative intronic miRNA promoters. RNA 2010, 16, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Carninci, P.; Sandelin, A.; Lenhard, B.; Katayama, S.; Shimokawa, K.; Ponjavic, J.; Semple, C.A.; Taylor, M.S.; Engstrom, P.G.; Frith, M.C.; et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat. Genet. 2006, 38, 626–635. [Google Scholar] [CrossRef]
- Nakshatri, H.; Bhat-Nakshatri, P.; Currie, R.A. Subunit association and DNA binding activity of the heterotrimeric transcription factor NF-Y is regulated by cellular redox. J. Biol. Chem. 1996, 271, 28784–28791. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.J.; van der Weyden, L.; Mayeda, A.; Stamm, S.; Morris, B.J.; Rasko, J.E. ZNF265—A novel spliceosomal protein able to induce alternative splicing. J. Cell Biol. 2001, 154, 25–32. [Google Scholar] [CrossRef]
- Schwartzentruber, J.; Buhas, D.; Majewski, J.; Sasarman, F.; Papillon-Cavanagh, S.; Thiffault, I.; Sheldon, K.M.; Massicotte, C.; Patry, L.; Simon, M.; et al. Mutation in the nuclear-encoded mitochondrial isoleucyl-tRNA synthetase IARS2 in patients with cataracts, growth hormone deficiency with short stature, partial sensorineural deafness, and peripheral neuropathy or with Leigh syndrome. Hum. Mutat. 2014, 35, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Cuvier, O.; Hirano, T. Chromosome condensation by a human condensin complex in Xenopus egg extracts. J. Biol. Chem. 2001, 276, 5417–5420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzechorzek, N.J.; Hardwick, S.W.; Jatikusumo, V.A.; Chirgadze, D.Y.; Pellegrini, L. CryoEM structures of human CMG-ATPgammaS-DNA and CMG-AND-1 complexes. Nucleic Acids Res. 2020, 48, 6980–6995. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Baris, Y.; Taylor, M.R.G.; Yeeles, J.T.P. Structure of a human replisome shows the organisation and interactions of a DNA replication machine. EMBO J. 2021, 40, e108819. [Google Scholar] [CrossRef] [PubMed]
- Jenkyn-Bedford, M.; Jones, M.L.; Baris, Y.; Labib, K.P.M.; Cannone, G.; Yeeles, J.T.P.; Deegan, T.D. A conserved mechanism for regulating replisome disassembly in eukaryotes. Nature 2021, 600, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Baris, Y.; Taylor, M.R.G.; Aria, V.; Yeeles, J.T.P. Fast and efficient DNA replication with purified human proteins. Nature 2022, 606, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Ishimi, Y.; Irie, D. G364R mutation of MCM4 detected in human skin cancer cells affects DNA helicase activity of MCM4/6/7 complex. J. Biochem. 2015, 157, 561–569. [Google Scholar] [CrossRef]
- Ishimi, Y. A DNA helicase activity is associated with an MCM4, -6, and -7 protein complex. J. Biol. Chem. 1997, 272, 24508–24513. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Dai, X.; Liu, Z.; Yin, P.; Wang, N.; Zhang, P. NR6A1 couples with cAMP response element binding protein and regulates vascular smooth muscle cell migration. Int. J. Biochem. Cell Biol. 2015, 69, 225–232. [Google Scholar] [CrossRef]
- Hor, H.; Francescatto, L.; Bartesaghi, L.; Ortega-Cubero, S.; Kousi, M.; Lorenzo-Betancor, O.; Jimenez-Jimenez, F.J.; Gironell, A.; Clarimon, J.; Drechsel, O.; et al. Missense mutations in TENM4, a regulator of axon guidance and central myelination, cause essential tremor. Hum. Mol. Genet. 2015, 24, 5677–5686. [Google Scholar] [CrossRef] [Green Version]
- Beck, R.; Rawet, M.; Wieland, F.T.; Cassel, D. The COPI system: Molecular mechanisms and function. FEBS Lett. 2009, 583, 2701–2709. [Google Scholar] [CrossRef] [Green Version]
- Razi, M.; Chan, E.Y.; Tooze, S.A. Early endosomes and endosomal coatomer are required for autophagy. J. Cell Biol. 2009, 185, 305–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, W.A.; Tsai, T.M.; Breidenbach, W. Per Primam thumb replantation for all patients with traumatic amputations. Clin. Orthop. Relat. Res. 1991, 266, 90–95. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, Y.; Ma, Y.; Ni, S.; Wikerholmen, T.; Xi, K.; Zhao, F.; Zhao, Z.; Wang, J.; Huang, B.; et al. Loss of COPZ1 induces NCOA4 mediated autophagy and ferroptosis in glioblastoma cell lines. Oncogene 2021, 40, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wang, H.G. CHOP is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J. Biol. Chem. 2004, 279, 45495–45502. [Google Scholar] [CrossRef] [Green Version]
- Ohoka, N.; Yoshii, S.; Hattori, T.; Onozaki, K.; Hayashi, H. TRB3, a novel ER stress-inducible gene, is induced via ATF4-CHOP pathway and is involved in cell death. EMBO J. 2005, 24, 1243–1255. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.J.; Pinto, J.P.; Picarote, G.; Costa, V.M.; Carvalho, F.; Rangel, M.; de Sousa, M.; de Almeida, S.F. ER stress-inducible factor CHOP affects the expression of hepcidin by modulating C/EBPalpha activity. PLoS ONE 2009, 4, e6618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Gao, B.; Zhang, J.; McEwen, T.; Ye, S.Q.; Zhang, D.; Fang, D. The HECT-type E3 ubiquitin ligase AIP2 inhibits activation-induced T-cell death by catalyzing EGR2 ubiquitination. Mol. Cell Biol. 2009, 29, 5348–5356. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Arya, R.; Brahmachari, V. Identification of a polycomb responsive region in human HoxA cluster and its long-range interaction with polycomb enriched genomic regions. Gene 2022, 845, 146832. [Google Scholar] [CrossRef]
- Xu, D.; Wang, Z.; Xia, Y.; Shao, F.; Xia, W.; Wei, Y.; Li, X.; Qian, X.; Lee, J.H.; Du, L.; et al. The gluconeogenic enzyme PCK1 phosphorylates INSIG1/2 for lipogenesis. Nature 2020, 580, 530–535. [Google Scholar] [CrossRef]
- Yokoyama, C.; Wang, X.; Briggs, M.R.; Admon, A.; Wu, J.; Hua, X.; Goldstein, J.L.; Brown, M.S. SREBP-1, a basic-helix-loop-helix-leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell 1993, 75, 187–197. [Google Scholar] [CrossRef]
- Amemiya-Kudo, M.; Shimano, H.; Hasty, A.H.; Yahagi, N.; Yoshikawa, T.; Matsuzaka, T.; Okazaki, H.; Tamura, Y.; Iizuka, Y.; Ohashi, K.; et al. Transcriptional activities of nuclear SREBP-1a, -1c, and -2 to different target promoters of lipogenic and cholesterogenic genes. J. Lipid Res. 2002, 43, 1220–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotzbauer, P.T.; Truax, A.C.; Trojanowski, J.Q.; Lee, V.M. Altered neuronal mitochondrial coenzyme A synthesis in neurodegeneration with brain iron accumulation caused by abnormal processing, stability, and catalytic activity of mutant pantothenate kinase 2. J. Neurosci. 2005, 25, 689–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, R.; Zhang, Y.M.; Lykidis, A.; Rock, C.O.; Jackowski, S. Localization and regulation of mouse pantothenate kinase 2. FEBS Lett. 2007, 581, 4639–4644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, R.; Rock, C.O.; Jackowski, S.; Zhang, Y.M. Activation of human mitochondrial pantothenate kinase 2 by palmitoylcarnitine. Proc. Natl. Acad. Sci. USA 2007, 104, 1494–1499. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Rock, C.O.; Jackowski, S. Biochemical properties of human pantothenate kinase 2 isoforms and mutations linked to pantothenate kinase-associated neurodegeneration. J. Biol. Chem. 2006, 281, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Pagani, F.; Trivedi, A.; Khatri, D.; Zizioli, D.; Garrafa, E.; Mitola, S.; Finazzi, D. Silencing of pantothenate kinase 2 reduces endothelial cell angiogenesis. Mol. Med. Rep. 2018, 18, 4739–4746. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. miRNet 2.0: Network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Pavlopoulos, G.A.; Kontou, P.I.; Pavlopoulou, A.; Bouyioukos, C.; Markou, E.; Bagos, P.G. Bipartite graphs in systems biology and medicine: A survey of methods and applications. Gigascience 2018, 7, 1–31. [Google Scholar] [CrossRef]
- Fan, Y.; Xia, J. miRNet-Functional Analysis and Visual Exploration of miRNA-Target Interactions in a Network Context. Methods Mol. Biol. 2018, 1819, 215–233. [Google Scholar] [CrossRef]
- Zhu, X.; Gerstein, M.; Snyder, M. Getting connected: Analysis and principles of biological networks. Genes Dev. 2007, 21, 1010–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Shin, V.Y.; Jin, H.; Ng, E.K.; Cheng, A.S.; Chong, W.W.; Wong, C.Y.; Leung, W.K.; Sung, J.J.; Chu, K.M. NF-kappaB targets miR-16 and miR-21 in gastric cancer: Involvement of prostaglandin E receptors. Carcinogenesis 2011, 32, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Mu, N.; Gu, J.; Huang, T.; Zhang, C.; Shu, Z.; Li, M.; Hao, Q.; Li, W.; Zhang, W.; Zhao, J.; et al. A novel NF-kappaB/YY1/microRNA-10a regulatory circuit in fibroblast-like synoviocytes regulates inflammation in rheumatoid arthritis. Sci. Rep. 2016, 6, 20059. [Google Scholar] [CrossRef] [Green Version]
- Toury, L.; Frankel, D.; Airault, C.; Magdinier, F.; Roll, P.; Kaspi, E. miR-140-5p and miR-140-3p: Key Actors in Aging-Related Diseases? Int. J. Mol. Sci. 2022, 23, 11439. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, J.; Chen, F.; Sun, Y. MiR-148b suppressed non-small cell lung cancer progression via inhibiting ALCAM through the NF-kappaB signaling pathway. Thorac. Cancer 2020, 11, 415–425. [Google Scholar] [CrossRef]
- Zhu, B.; Ye, J.; Ashraf, U.; Li, Y.; Chen, H.; Song, Y.; Cao, S. Transcriptional regulation of miR-15b by c-Rel and CREB in Japanese encephalitis virus infection. Sci. Rep. 2016, 6, 22581. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Deng, R.; Wang, Y.; Zhang, H.; Dou, J.; Li, L.; Du, Y.; Chen, R.; Cheng, J.; Yu, J. Twist1/Dnmt3a and miR186 establish a regulatory circuit that controls inflammation-associated prostate cancer progression. Oncogenesis 2017, 6, e315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prattichizzo, F.; Micolucci, L.; Cricca, M.; De Carolis, S.; Mensa, E.; Ceriello, A.; Procopio, A.D.; Bonafe, M.; Olivieri, F. Exosome-based immunomodulation during aging: A nano-perspective on inflamm-aging. Mech. Ageing Dev. 2017, 168, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jo, H.; Kwon, Y.; Jeong, M.S.; Jung, H.S.; Kim, Y.; Jeoung, D. MiR-154-5p-MCP1 Axis Regulates Allergic Inflammation by Mediating Cellular Interactions. Front. Immunol. 2021, 12, 663726. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Y.; Yang, Z.; Sun, Q.; Yang, X.; Hu, Y.; Xie, H.; Gao, H.J.; Guo, L.M.; Yi, J.Y.; Liu, M.; et al. miR-377-3p drives malignancy characteristics via upregulating GSK-3beta expression and activating NF-kappaB pathway in hCRC cells. J. Cell Biochem. 2018, 119, 2124–2134. [Google Scholar] [CrossRef]
- Sole, C.; Domingo, S.; Penzo, E.; Moline, T.; Porres, L.; Aparicio, G.; Ferrer, B.; Cortes-Hernandez, J. Downregulation of miR-885-5p Promotes NF-kappaB Pathway Activation and Immune Recruitment in Cutaneous Lupus Erythematosus. J. Investig. Dermatol. 2023, 143, 209–219.e213. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Vamadevan, A.S.; Abreu, M.T. Toll-like receptors (TLRs) and Nod-like receptors (NLRs) in inflammatory disorders. Semin. Immunol. 2009, 21, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. Emerging role of NF-kappaB signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal. 2012, 24, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, M.S.; Ghosh, S. NF-kappaB, the first quarter-century: Remarkable progress and outstanding questions. Genes Dev. 2012, 26, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Garcia, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-kappaB in Ageing and Age-Related Diseases: Lessons from Genetically Modified Mouse Models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef]
- Bass, V.L.; Wong, V.C.; Bullock, M.E.; Gaudet, S.; Miller-Jensen, K. TNF stimulation primarily modulates transcriptional burst size of NF-kappaB-regulated genes. Mol. Syst. Biol. 2021, 17, e10127. [Google Scholar] [CrossRef]
- Kizilirmak, C.; Bianchi, M.E.; Zambrano, S. Insights on the NF-kappaB System Using Live Cell Imaging: Recent Developments and Future Perspectives. Front. Immunol. 2022, 13, 886127. [Google Scholar] [CrossRef]
- Liu, X.; Shao, Y.; Zhou, J.; Qian, G.; Ma, Z. Nuclear Factor kappaB Signaling and Its Related Non-coding RNAs in Cancer Therapy. Mol. Ther. Nucleic Acids 2020, 19, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Rodier, F.; Coppe, J.P.; Patil, C.K.; Hoeijmakers, W.A.; Munoz, D.P.; Raza, S.R.; Freund, A.; Campeau, E.; Davalos, A.R.; Campisi, J. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat. Cell Biol. 2009, 11, 973–979. [Google Scholar] [CrossRef]
- Laberge, R.M.; Zhou, L.; Sarantos, M.R.; Rodier, F.; Freund, A.; de Keizer, P.L.; Liu, S.; Demaria, M.; Cong, Y.S.; Kapahi, P.; et al. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging Cell 2012, 11, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freund, A.; Orjalo, A.V.; Desprez, P.Y.; Campisi, J. Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2017, 8, 1960. [Google Scholar] [CrossRef] [Green Version]
- Songkiatisak, P.; Rahman, S.M.T.; Aqdas, M.; Sung, M.H. NF-kappaB, a culprit of both inflamm-ageing and declining immunity? Immun. Ageing 2022, 19, 20. [Google Scholar] [CrossRef]
- Josephson, A.M.; Leclerc, K.; Remark, L.H.; Lopez, E.M.; Leucht, P. Systemic NF-kappaB-mediated inflammation promotes an aging phenotype in skeletal stem/progenitor cells. Aging 2021, 13, 13421–13429. [Google Scholar] [CrossRef]
- Haga, M.; Okada, M. Systems approaches to investigate the role of NF-kappaB signaling in aging. Biochem. J. 2022, 479, 161–183. [Google Scholar] [CrossRef]
- Hounkpe, B.W.; Chenou, F.; de Lima, F.; De Paula, E.V. HRT Atlas v1.0 database: Redefining human and mouse housekeeping genes and candidate reference transcripts by mining massive RNA-seq datasets. Nucleic Acids Res. 2021, 49, D947–D955. [Google Scholar] [CrossRef]
- Bracken, C.P.; Scott, H.S.; Goodall, G.J. A network-biology perspective of microRNA function and dysfunction in cancer. Nat. Rev. Genet. 2016, 17, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef]
- Xie, G.Y.; Xia, M.; Miao, Y.R.; Luo, M.; Zhang, Q.; Guo, A.Y. FFLtool: A web server for transcription factor and miRNA feed forward loop analysis in human. Bioinformatics 2020, 36, 2605–2607. [Google Scholar] [CrossRef]
- Ye, H.; Liu, X.; Lv, M.; Wu, Y.; Kuang, S.; Gong, J.; Yuan, P.; Zhong, Z.; Li, Q.; Jia, H.; et al. MicroRNA and transcription factor co-regulatory network analysis reveals miR-19 inhibits CYLD in T-cell acute lymphoblastic leukemia. Nucleic Acids Res. 2012, 40, 5201–5214. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, M.L.; Shaban, M.S.; Albert, B.V.; Gokcen, A.; Kracht, M. The Crosstalk of Endoplasmic Reticulum (ER) Stress Pathways with NF-kappaB: Complex Mechanisms Relevant for Cancer, Inflammation and Infection. Biomedicines 2018, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Sandhu, U.G.; Quintana-Quezada, R.A.; Hoang, N.M.; Fujiwara, K.; Abe, J.I. Flow signaling and atherosclerosis. Cell Mol. Life Sci. 2017, 74, 1835–1858. [Google Scholar] [CrossRef] [Green Version]
- Dolfini, D.; Gatta, R.; Mantovani, R. NF-Y and the transcriptional activation of CCAAT promoters. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Vaiman, D.; Calicchio, R.; Miralles, F. Landscape of transcriptional deregulations in the preeclamptic placenta. PLoS ONE 2013, 8, e65498. [Google Scholar] [CrossRef]
- Mato-Basalo, R.; Morente-Lopez, M.; Arntz, O.J.; van de Loo, F.A.J.; Fafian-Labora, J.; Arufe, M.C. Therapeutic Potential for Regulation of the Nuclear Factor Kappa-B Transcription Factor p65 to Prevent Cellular Senescence and Activation of Pro-Inflammatory in Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 3367. [Google Scholar] [CrossRef]
- Slota, J.A.; Booth, S.A. MicroRNAs in Neuroinflammation: Implications in Disease Pathogenesis, Biomarker Discovery and Therapeutic Applications. Noncoding RNA 2019, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Grants, J.M.; Wegrzyn, J.; Hui, T.; O’Neill, K.; Shadbolt, M.; Knapp, D.; Parker, J.; Deng, Y.; Gopal, A.; Docking, T.R.; et al. Altered microRNA expression links IL6 and TNF-induced inflammaging with myeloid malignancy in humans and mice. Blood 2020, 135, 2235–2251. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Lu, L.F.; Rudensky, A.Y.; Baltimore, D. An NF-kappaB-microRNA regulatory network tunes macrophage inflammatory responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalife, J.; Ghose, J.; Martella, M.; Viola, D.; Rocci, A.; Troadec, E.; Terrazas, C.; Satoskar, A.R.; Gunes, E.G.; Dona, A.; et al. MiR-16 regulates crosstalk in NF-kappaB tolerogenic inflammatory signaling between myeloma cells and bone marrow macrophages. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Mensa, E.; Guescini, M.; Giuliani, A.; Bacalini, M.G.; Ramini, D.; Corleone, G.; Ferracin, M.; Fulgenzi, G.; Graciotti, L.; Prattichizzo, F.; et al. Small extracellular vesicles deliver miR-21 and miR-217 as pro-senescence effectors to endothelial cells. J. Extracell. Vesicles 2020, 9, 1725285. [Google Scholar] [CrossRef]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-kappaB and AP-1 connection: Mechanism of NF-kappaB-dependent regulation of AP-1 activity. Mol. Cell Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [Green Version]
- Del Mar Diaz-Gonzalez, S.; Rodriguez-Aguilar, E.D.; Meneses-Acosta, A.; Valadez-Graham, V.; Deas, J.; Gomez-Ceron, C.; Tavira-Montalvan, C.A.; Arizmendi-Heras, A.; Ramirez-Bello, J.; Zurita-Ortega, M.E.; et al. Transregulation of microRNA miR-21 promoter by AP-1 transcription factor in cervical cancer cells. Cancer Cell Int. 2019, 19, 214. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, F.; Capri, M.; Bonafe, M.; Morsiani, C.; Jung, H.J.; Spazzafumo, L.; Vina, J.; Suh, Y. Circulating miRNAs and miRNA shuttles as biomarkers: Perspective trajectories of healthy and unhealthy aging. Mech. Ageing Dev. 2017, 165, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Zhavoronkov, A.; Li, R.; Ma, C.; Mamoshina, P. Deep biomarkers of aging and longevity: From research to applications. Aging 2019, 11, 10771–10780. [Google Scholar] [CrossRef]
- Hamdan, Y.; Mazini, L.; Malka, G. Exosomes and Micro-RNAs in Aging Process. Biomedicines 2021, 9, 968. [Google Scholar] [CrossRef]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum. Genet. 2020, 139, 291–308. [Google Scholar] [CrossRef]
- Rovcanin Dragovic, I.; Popovic, N.; Zdralevic, M.; Radulovic, L.; Vukovic, T.; Marzano, F.; Tullo, A.; Radunovic, M. Inflammation-Related microRNAs-146a and -155 Are Upregulated in Mild Cognitive Impairment Subjects Among Older Age Population in Montenegro. J. Alzheimers Dis. 2022, 90, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Sahu, H.; Shrma, S.; Gondhalakar, S. A Brief Overview on Data Mining Survey. Int. J. Comput. Technol. Electron. Eng. 2011, 1, 114–121. [Google Scholar]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. miRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36, D154–D158. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [Green Version]
- Hinske, L.C.; Franca, G.S.; Torres, H.A.; Ohara, D.T.; Lopes-Ramos, C.M.; Heyn, J.; Reis, L.F.; Ohno-Machado, L.; Kreth, S.; Galante, P.A. miRIAD-integrating microRNA inter- and intragenic data. Database 2014, 2014. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Cho, J.W.; Lee, S.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Siklenka, K.; Arora, S.K.; Ribeiro, P.; Kimmins, S.; Xia, J. miRNet—Dissecting miRNA-target interactions and functional associations through network-based visual analysis. Nucleic Acids Res. 2016, 44, W135–W141. [Google Scholar] [CrossRef] [PubMed]
- Aging Atlas, C. Aging Atlas: A multi-omics database for aging biology. Nucleic Acids Res. 2021, 49, D825–D830. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Russo, F.; Di Bella, S.; Vannini, F.; Berti, G.; Scoyni, F.; Cook, H.V.; Santos, A.; Nigita, G.; Bonnici, V.; Lagana, A.; et al. miRandola 2017: A curated knowledge base of non-invasive biomarkers. Nucleic Acids Res. 2018, 46, D354–D359. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative NF-kB Responsive miRNAs * | Mature miRNA Expression ** in Tissues Linked to Inflammatory Processes | Prediction of Promoter Location According to PROmiRNA | miRNA Age | Chromosomes |
|---|---|---|---|---|
| hsa-miR-101 | Immune system | intergenic | v | 1 |
| hsa-miR-103 | Liver and Immune system | host gene-intronic | v | 20 |
| hsa-miR-106b | Immune system | host gene | m | 7 |
| hsa-miR-10a | Immune system | host gene | v | 17 |
| hsa-miR-132 | Immune system | intergenic | m | 17 |
| hsa-miR-136 | Immune system | intergenic | m | 14 |
| hsa-miR-140-3p | Liver and Immune system | host gene | v | 16 |
| hsa-miR-140-5p | Liver and Immune system | host gene | v | 16 |
| hsa-miR-144 | Liver and Immune system | intergenic | v | 17 |
| hsa-miR-146a | Immune system | intronic | m | 5 |
| hsa-miR-148b | Immune system | host gene | m | 12 |
| hsa-miR-150 | Immune system | intergenic | m | 19 |
| hsa-miR-154 | Liver | intergenic | m | 14 |
| hsa-miR-155 | Immune system | intronic | v | 21 |
| hsa-miR-15a | Liver and Immune system | host gene-intronic | v | 13 |
| hsa-miR-15b | Liver and Immune system | host gene | v | 3 |
| hsa-miR-16 | Liver and Immune system | host gene-intronic | v | 13|3 |
| hsa-miR-17 | Liver and Immune system | intronic | v | 13 |
| hsa-miR-181b | Immune system | host gene | v | 9 |
| hsa-miR-183 | Immune system | intergenic | m | 7 |
| hsa-miR-186 | Liver and Immune system | host gene-intronic | m | 1 |
| hsa-miR-188-3p | Immune system | intronic | m | X |
| hsa-miR-188-5p | Immune system | intronic | m | X |
| hsa-miR-18a | Immune system | intronic | v | 13 |
| hsa-miR-192 | Liver and Immune system | intergenic | m | 11 |
| hsa-miR-194 | Liver and Immune system | host gene-intergenic | m|v | 11 |
| hsa-miR-195 | Liver | intronic | m | 17 |
| hsa-miR-19a | Immune system | intronic | v | 13 |
| hsa-miR-19b | Immune system | intronic | m|v | 13 |
| hsa-miR-20a | Immune system | intronic | v | 13 |
| hsa-miR-221 | Immune system | intergenic | v | X |
| hsa-miR-222 | Immune system | intergenic | v | X |
| hsa-miR-23a | Liver and Immune system | intergenic | m | 19 |
| hsa-miR-25 | Liver and Immune system | host gene | m | 7 |
| hsa-miR-27a | Liver and Immune system | intergenic | m | 19 |
| hsa-miR-296-5p | Immune system | intergenic | m | 20 |
| hsa-miR-30c | Liver and Immune system | host gene | v | 1 |
| hsa-miR-30e | Liver and Immune system | host gene | v | 1 |
| hsa-miR-31 | Immune system | intronic | v | 9 |
| hsa-miR-32 | Immune system | intronic | v | 9 |
| hsa-miR-339-3p | Immune system | intronic | m | 7 |
| hsa-miR-339-5p | Immune system | intronic | m | 7 |
| hsa-miR-33a | Immune system | intronic | m | 22 |
| hsa-miR-33b | Immune system | host gene-intronic | m | 17 |
| hsa-miR-362-5p | Immune system | intronic | m | X |
| hsa-miR-374a | Immune system | host gene | m | X |
| hsa-miR-374b | Immune system | intergenic | m | X |
| hsa-miR-377 | Liver | intergenic | m | 14 |
| hsa-miR-424 | Liver and Immune system | intergenic | m | X |
| hsa-miR-450a | Immune system | intergenic | m | X |
| hsa-miR-491-5p | Immune system | intronic | m | 9 |
| hsa-miR-500 | Liver | intronic | m | X |
| hsa-miR-503 | Immune system | intergenic | m | X |
| hsa-miR-532-3p | Immune system | intronic | m | X |
| hsa-miR-542-5p | Immune system | intergenic | m | X |
| hsa-miR-545 | Immune system | host gene | m | X |
| hsa-miR-548d-5p | Immune system | intronic | p | 8 |
| hsa-miR-570 | Immune system | intronic | p | 3 |
| hsa-miR-616 | Immune system | host gene | p | 12 |
| hsa-miR-625 | Immune system | intronic | p | 14 |
| hsa-miR-629 | Immune system | intronic | p | 15 |
| hsa-miR-708 | Immune system | host gene | m | 11 |
| hsa-miR-874 | Liver and Immune system | intronic | m | 5 |
| hsa-miR-885-5p | Liver | intronic | p | 3 |
| hsa-miR-92a | Liver and Immune system | intronic | m | 13 |
| hsa-miR-93 | Immune system | host gene | m | 7 |
| hsa-miR-98 | Liver and Immune system | intronic | m | X |
| hsa-miR-99a | Liver and Immune system | intronic | v | 21 |
| miRNA Hairpin | Host Gene Information | Intragenic miRNA Information | ||||||
|---|---|---|---|---|---|---|---|---|
| Ch | Host Gene | Entrez Gene ID | Host Gene Biological Process | Intron n° | Distance from Upstream Exon | Direction | Mature miRNA ID in miRBase 22.1 Release (In Parentheses Previous IDs) § | |
| hsa-mir-30c-1 H | 1 | NFYC | 4802 | Transcription regulation ‡ | 4 | 4038 | sense | miR-30c-5p (miR-30c); miR-30c-1-3p (miR-30c-1*) |
| hsa-mir-30e H | 1 | NFYC | 4802 | Transcription regulation ‡ | 4 | 1109 | sense | miR-30e-5p (miR-30e); miR-30e-3p (miR-30e*) |
| hsa-mir-186 H, I | 1 | ZRANB2 | 9406 | mRNA processing ‡ | 8 | 1560 | sense | miR-186-5p (miR-186); miR-186-3p (miR-186*) |
| hsa-mir-194-1 H, Ig | 1, 11 | IARS2 | 55699 | Protein biosynthesis | 12 | 6996 | antisense | miR-194-5p (miR-194); miR-194-3p (miR-194*) |
| hsa-mir-15b H | 3 | SMC4 | 10051 | DNA condensation | 3 | 84 | sense | miR-15b-5p (miR-15b); miR-15b-3p (miR-15b*) |
| hsa-mir-16-2 H, I | 3, 13 | SMC4 | 10051 | DNA condensation | 3 | 241 | sense | miR-16-5p (miR-16); miR-16-2-3p (miR-16-2*) |
| hsa-mir-106b H | 7 | MCM7 | 4176 | DNA replication | 13 | 99 | sense | miR-106b-5p (miR-106b); miR-106b-3p (miR-106b*) |
| hsa-mir-25 H | 7 | MCM7 | 4176 | DNA replication | 13 | 530 | sense | miR-25-5p (miR-25*); miR-25-3p (miR-25) |
| hsa-mir-93 H | 7 | MCM7 | 4176 | DNA replication | 13 | 326 | sense | miR-93-5p (miR-93); miR-93-3p (miR-93*) |
| hsa-mir-181b-2 H | 9 | NR6A1 | 2649 | Transcription regulation | 2 | 139,140 | antisense | miR-181b-5p (miR-181b); miR-181b-2-3p |
| hsa-mir-708 H | 11 | TENM4 | 26011 | Differentiation | 1 | 38,400 | sense | miR-708-5p (miR-708); miR-708-3p (miR-708*) |
| hsa-mir-148b H | 12 | COPZ1 | 22818 | Protein transport ‡ | 1 | 12,035 | sense | miR-148b-5p (miR-148b*); miR-148b-3p (miR-148b) |
| hsa-mir-616 H | 12 | DDIT3 | 1649 | Transcription regulation | 1 | 1159 | sense | miR-616-5p (miR-616, miR-616*); miR-616-3p (miR-616) |
| hsa-mir-140 H | 16 | WWP2 | 11060 | Ubl conjugation pathway | 6 | 1191 | sense | miR-140-5p (miR-140); miR-140-3p |
| hsa-mir-10a H | 17 | HOXB3 | 3213 | Transcription regulation | 1 | 2202 | sense | miR-10a-5p (miR-10a); miR-10a-3p (miR-10a*) |
| hsa-mir-33b H, I | 17 | SREBF1 | 6720 | Transcription regulation | 12 | 314 | sense | miR-33b-5p (miR-33b); miR-33b-3p (miR-33b*) |
| hsa-mir-103a-2 H, I | 20 | PANK2 | 80025 | Coenzyme A biosynthesis | 5 | 444 | sense | miR-103a-2-5p (miR-103-2*; miR-103a-2*); miR-103a-3p (miR-103, miR-103a) |
| Differentially Expressed Gene | Cell/Tissue | Treatment/Condition | log2 Fold Change | p-Value | q-Value | DOI |
|---|---|---|---|---|---|---|
| DDIT3 | Human diploid fibroblasts IMR90 | Replicative senescence | −1.09474 | 1.55 × 10−23 | 2.63 × 10−22 | 10.1093/nar/gkz555 |
| DDIT3 | Lung | COVID-19 vs. Control | 3.04617 | 1.11 × 10−17 | 2.55 × 10−16 | 10.1038/s41556-021-00796-6 |
| HOXB3 | Human induced pluripotent stem (iPS) cell-derived neural progenitor cells | Alzheimer’s disease | 1.30000 | 5.10 × 10−4 | 3.40 × 10−2 | 10.1016/j.celrep.2019.01.023 |
| MCM7 | Human arterial endothelial cell | Ionizing radiation | −1.07367 | 2.66 × 10−33 | 1.94 × 10−31 | 10.1093/nar/gkz555 |
| MCM7 | Human diploid fibroblasts WI38 | Ionizing radiation | −1.60665 | 1.53 × 10−10 | 5.99 × 10−9 | 10.1093/nar/gkz555 |
| MCM7 | WRN-/- mesenchymal stem cell | Vitamin C | 1.90980 | 5.08 × 10−51 | 10.1007/s13238-016-0278-1 | |
| SMC4 | Human arterial endothelial cell | Ionizing radiation | −1.14304 | 5.14 × 10−38 | 4.56 × 10−36 | 10.1093/nar/gkz555 |
| SMC4 | Human diploid fibroblasts WI38 | Ionizing radiation | −2.31474 | 1.24 × 10−14 | 8.41 × 10−13 | 10.1093/nar/gkz555 |
| SMC4 | Lung | COVID19 vs. Control | −2.85324 | 4.44 × 10−10 | 3.79 × 10−9 | 10.1038/s41556-021-00796-6 |
| SMC4 | Human diploid fibroblasts WI38 | Replicative senescence | −1.44078 | 8.14 × 10−7 | 1.75 × 10−5 | 10.1093/nar/gkz555 |
| SMC4 | WRN-/- mesenchymal stem cell | Vitamin C | 2.18610 | 4.24 × 10−55 | 10.1007/s13238-016-0278-1 | |
| TENM4 | Human diploid fibroblasts WI38 | Replicative senescence | −4.84098 | 1.97 × 10−52 | 10.1093/nar/gkz555 | |
| TENM4 | Human diploid fibroblasts IMR90 | Ionizing radiation | −1.63819 | 3.26 × 10−23 | 5.65 × 10−22 | 10.1093/nar/gkz555 |
| TENM4 | Human diploid fibroblasts WI38 | Ionizing radiation | 1.82630 | 6.17 × 10−12 | 3.09 × 10−10 | 10.1093/nar/gkz555 |
| TENM4 | Human diploid fibroblasts IMR90 | Replicative senescence | −1.03845 | 1.66 × 10−10 | 1.24 × 10−9 | 10.1093/nar/gkz555 |
| TENM4 | Human induced pluripotent stem (iPS) cell-derived neurons | Alzheimer’s disease | 1.70000 | 5.50 × 10−4 | 1.70 × 10−2 | 10.1016/j.celrep.2019.01.023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micolucci, L.; Matacchione, G.; Albertini, M.C.; Marra, M.; Ramini, D.; Giuliani, A.; Sabbatinelli, J.; Procopio, A.D.; Olivieri, F.; Marsico, A.; et al. A Data-Mining Approach to Identify NF-kB-Responsive microRNAs in Tissues Involved in Inflammatory Processes: Potential Relevance in Age-Related Diseases. Int. J. Mol. Sci. 2023, 24, 5123. https://doi.org/10.3390/ijms24065123
Micolucci L, Matacchione G, Albertini MC, Marra M, Ramini D, Giuliani A, Sabbatinelli J, Procopio AD, Olivieri F, Marsico A, et al. A Data-Mining Approach to Identify NF-kB-Responsive microRNAs in Tissues Involved in Inflammatory Processes: Potential Relevance in Age-Related Diseases. International Journal of Molecular Sciences. 2023; 24(6):5123. https://doi.org/10.3390/ijms24065123
Chicago/Turabian StyleMicolucci, Luigina, Giulia Matacchione, Maria Cristina Albertini, Massimo Marra, Deborah Ramini, Angelica Giuliani, Jacopo Sabbatinelli, Antonio Domenico Procopio, Fabiola Olivieri, Annalisa Marsico, and et al. 2023. "A Data-Mining Approach to Identify NF-kB-Responsive microRNAs in Tissues Involved in Inflammatory Processes: Potential Relevance in Age-Related Diseases" International Journal of Molecular Sciences 24, no. 6: 5123. https://doi.org/10.3390/ijms24065123