
Oral Administration of Lipopolysaccharide Enhances Insulin Signaling-Related Factors in the KK/Ay Mouse Model of Type 2 Diabetes Mellitus

 ,
,
Abstract
:1. Introduction
2. Results
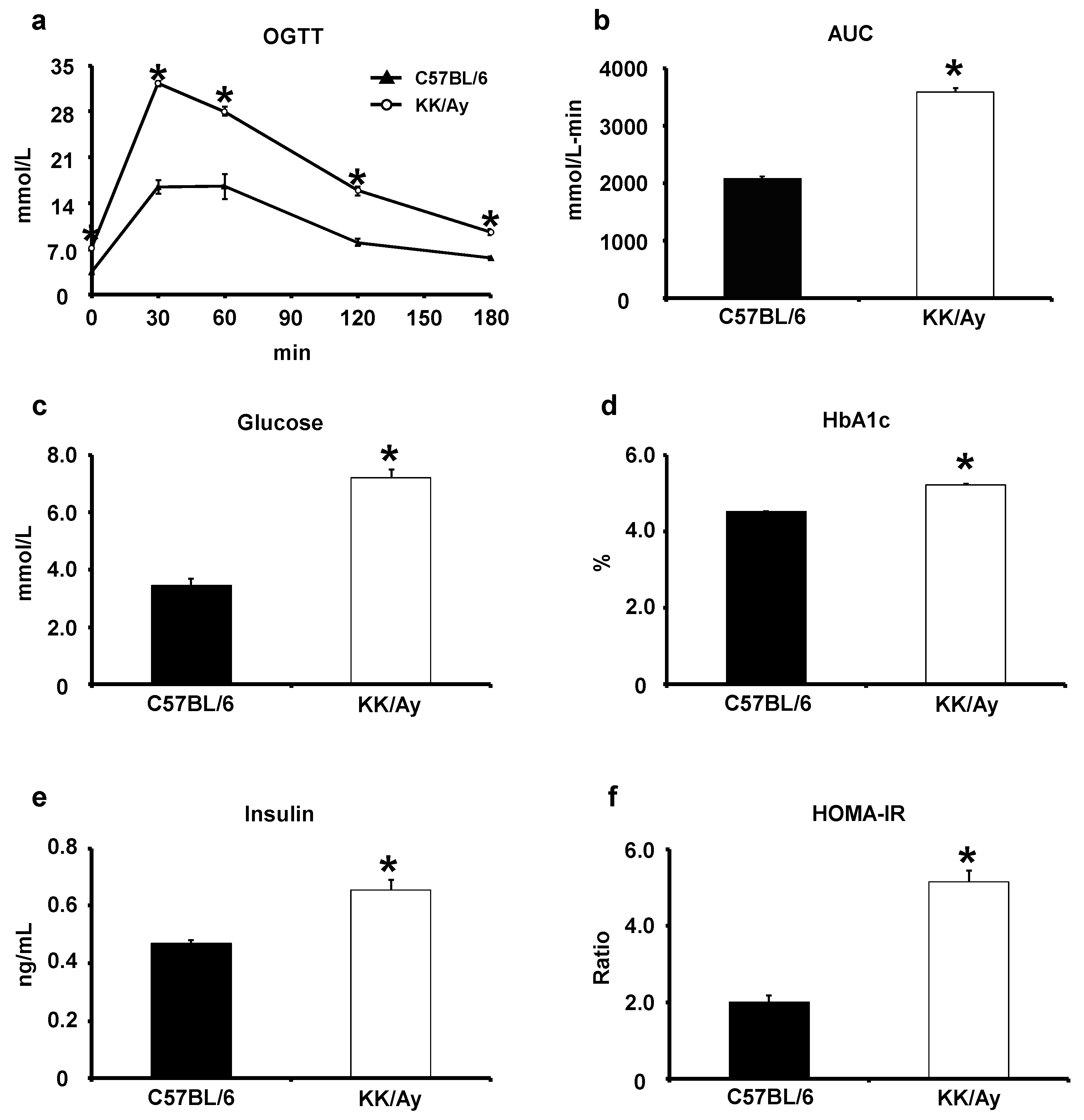
2.1. Verification of the Development of Type 2 Diabetes Mellitus in KK/Ay Mice
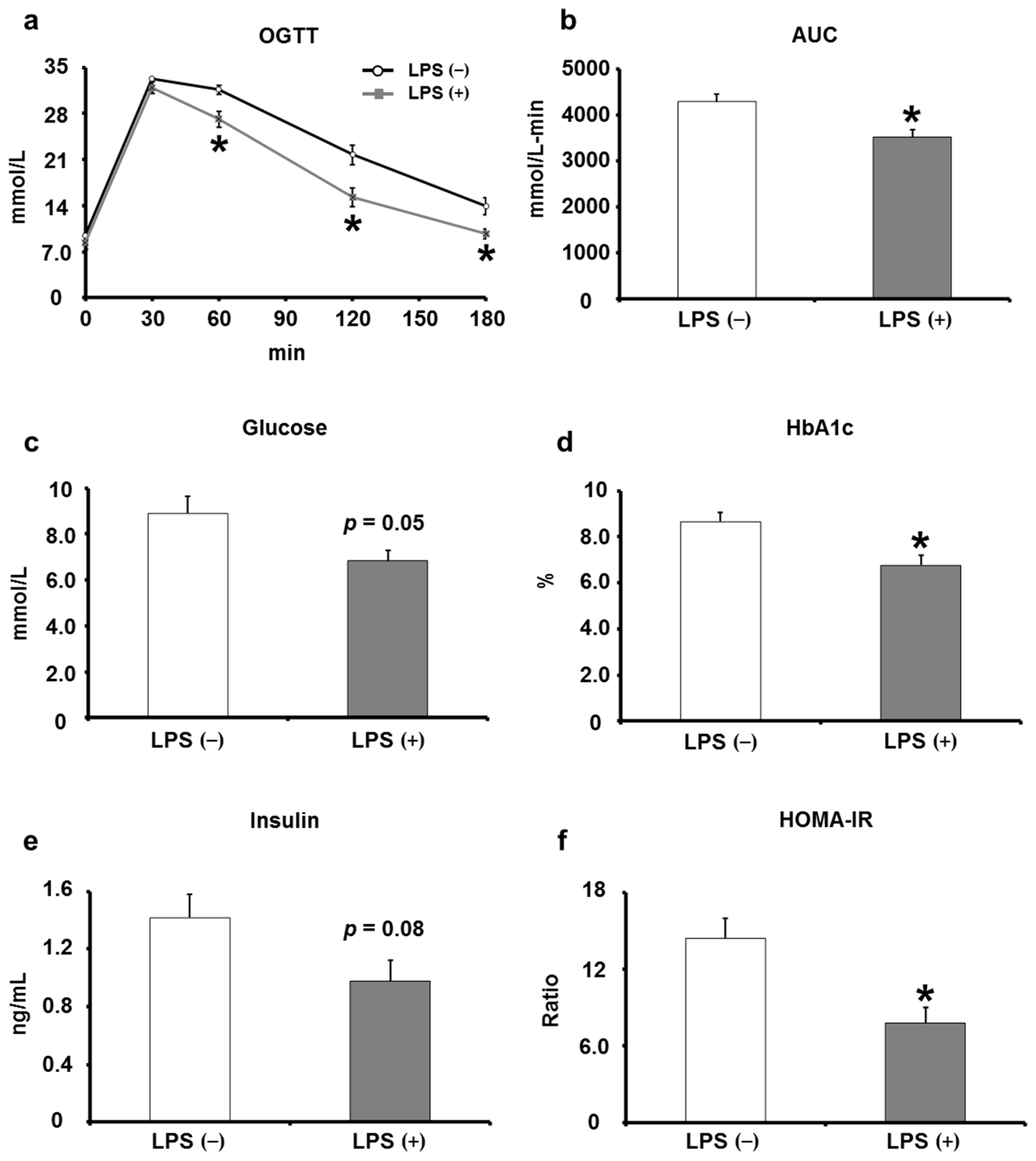
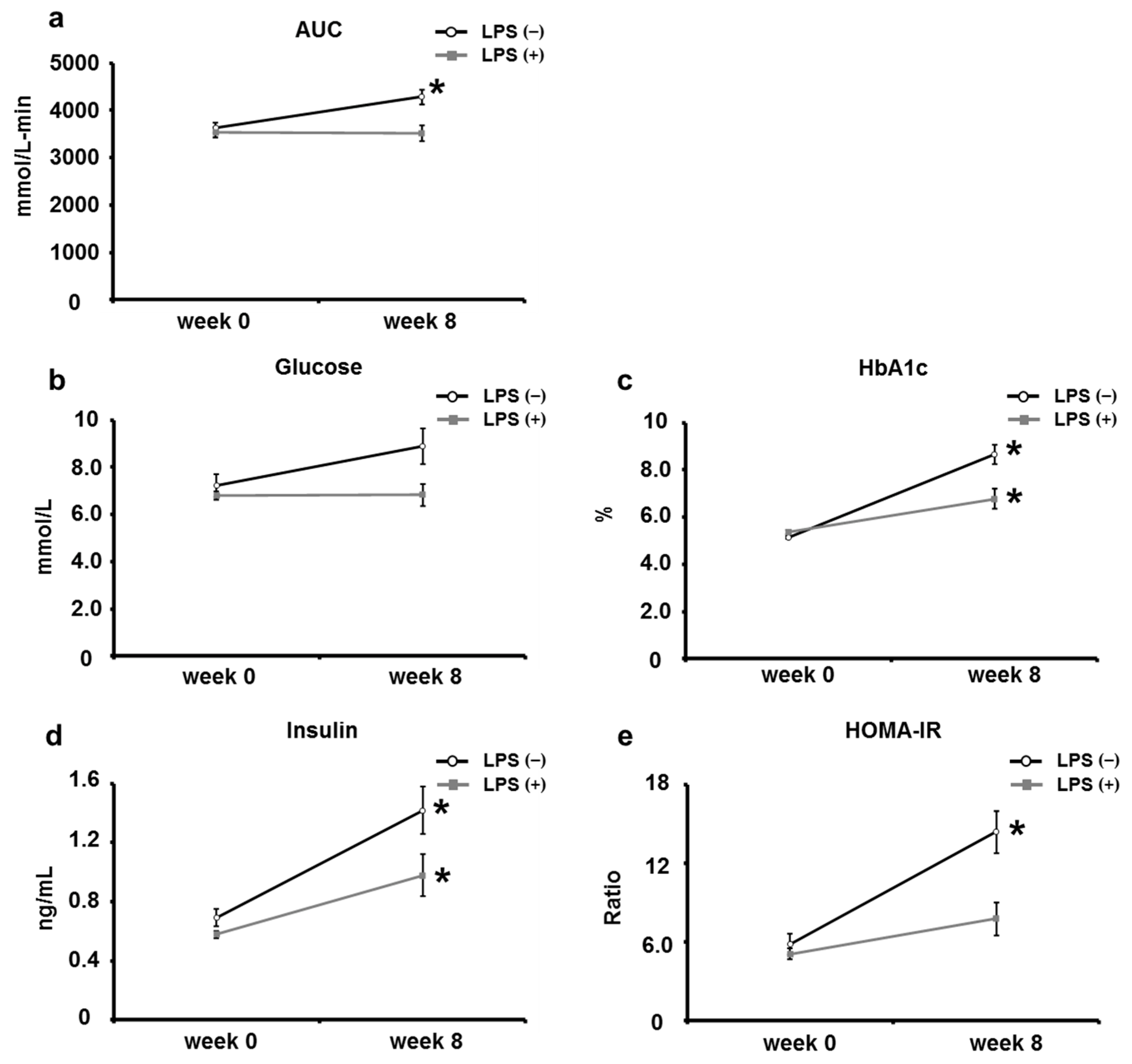
2.2. Oral LPS Administration Suppresses Type 2 Diabetes Mellitus in KK/Ay Mice
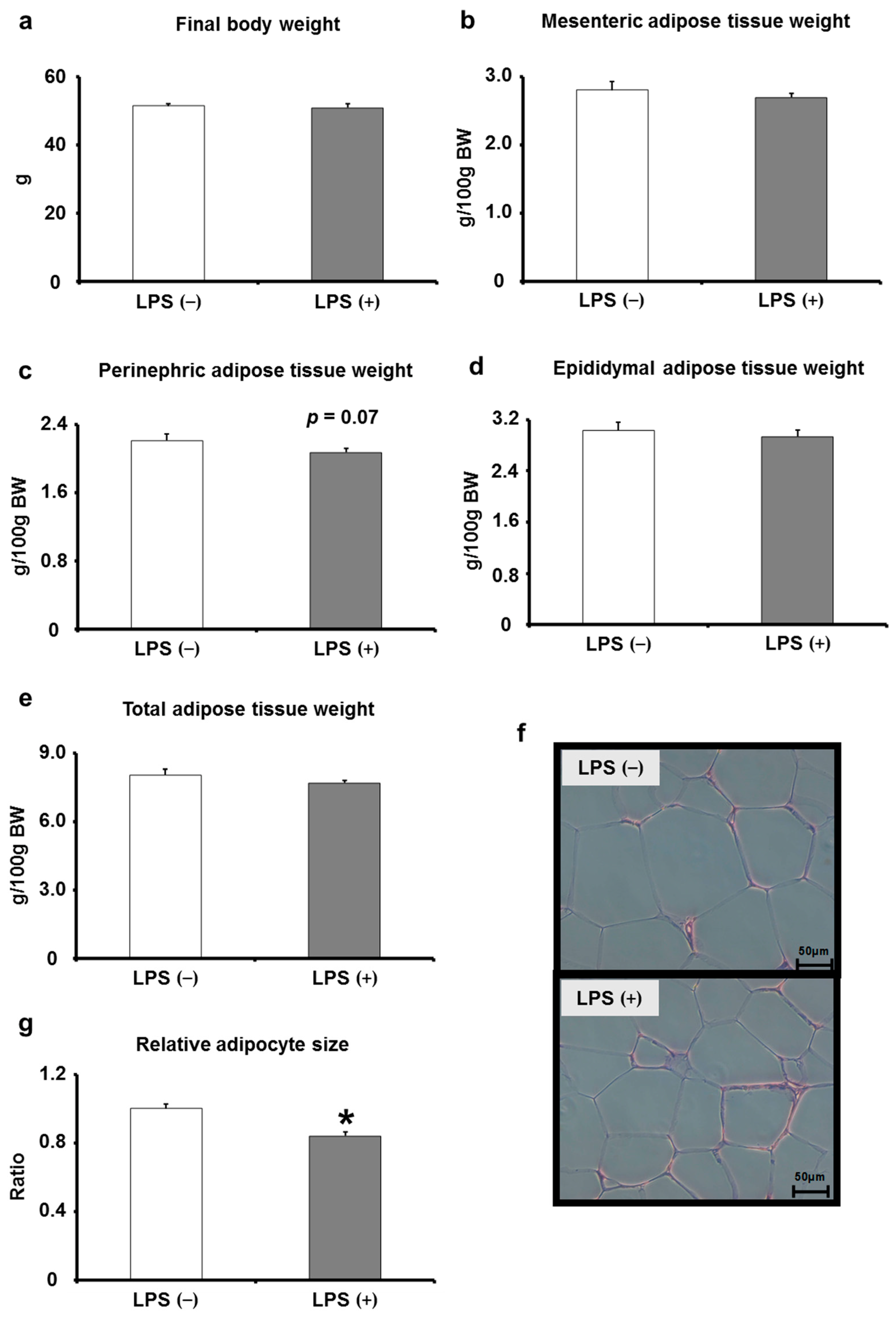
2.3. Effects of Oral LPS Administration on Adipose Tissues
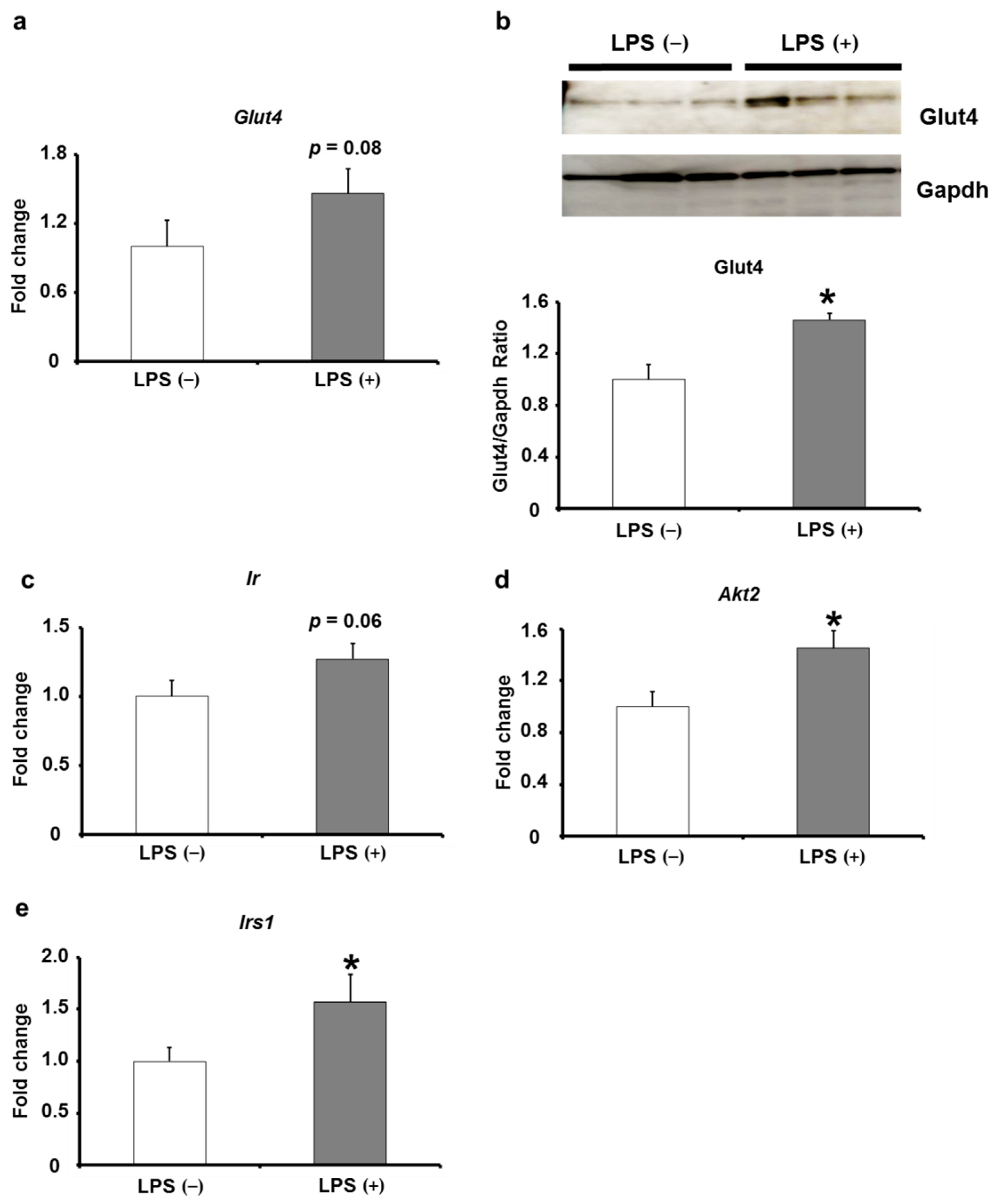
2.4. Effects of Oral LPS Administration on the Expression Levels of Insulin Signaling-Related Factors in Adipose Tissues of KK/Ay Mice
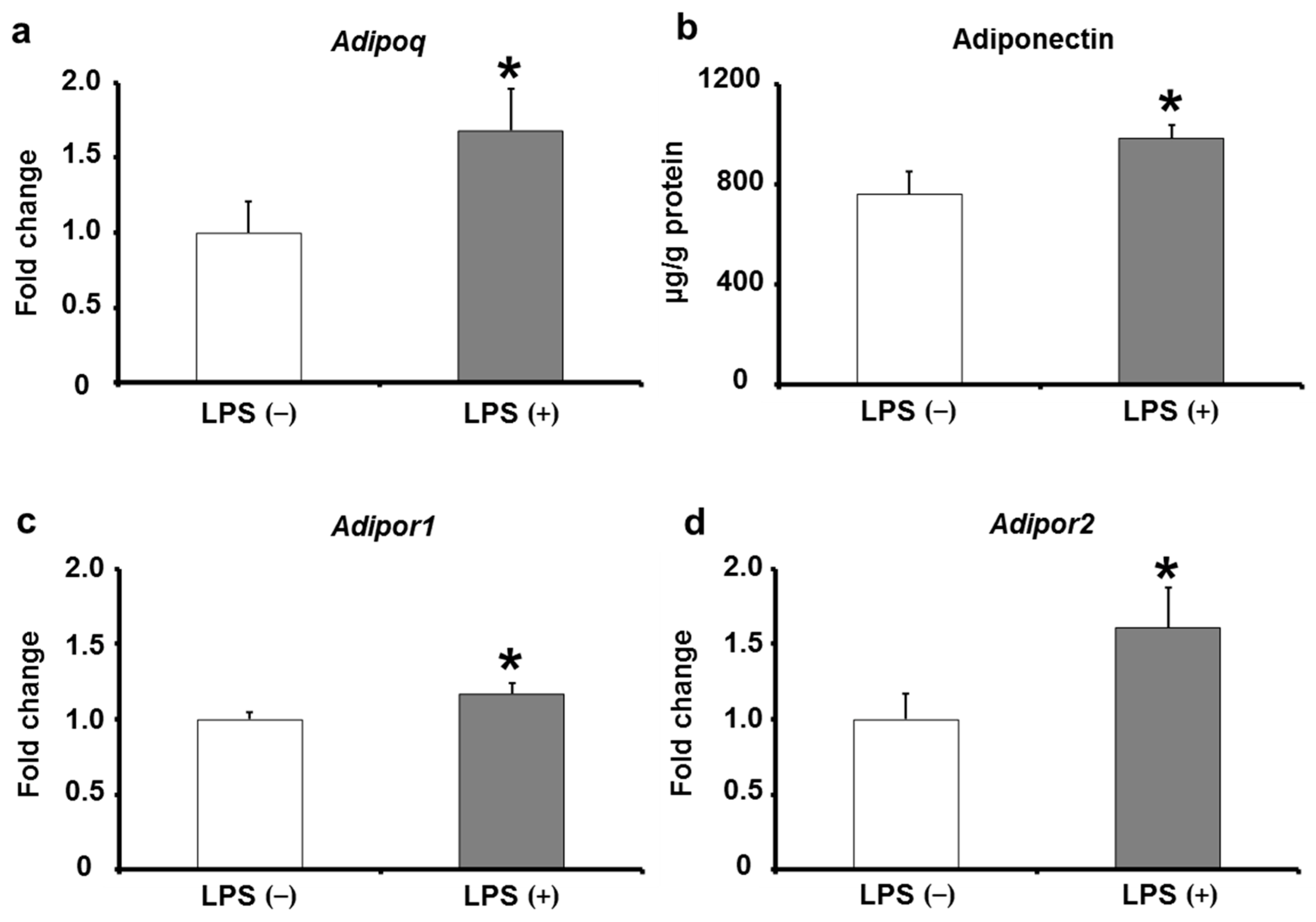
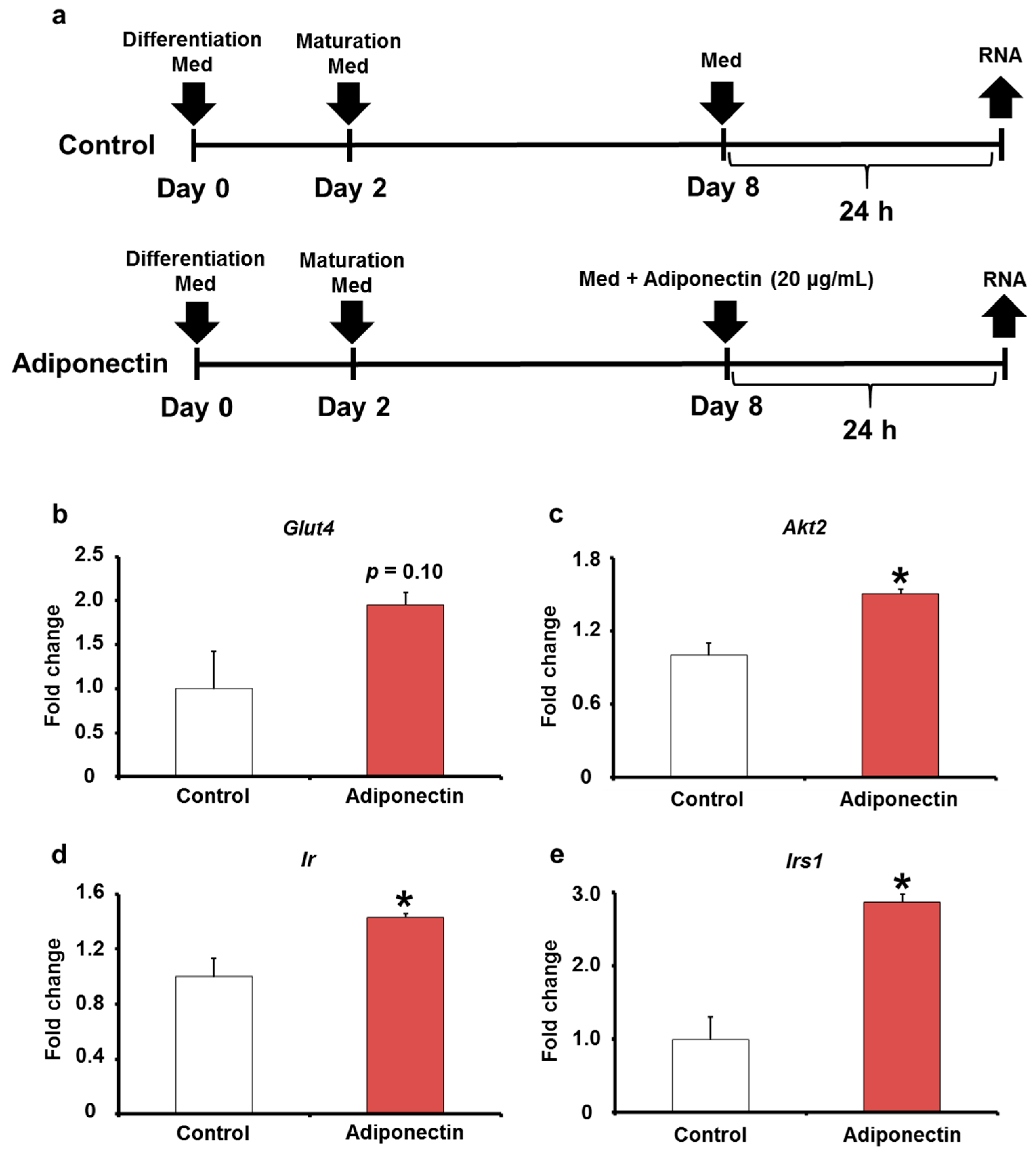
2.5. Adiponectin Expression in Adipose Tissues of KK/Ay Mice by Oral LPS Administration
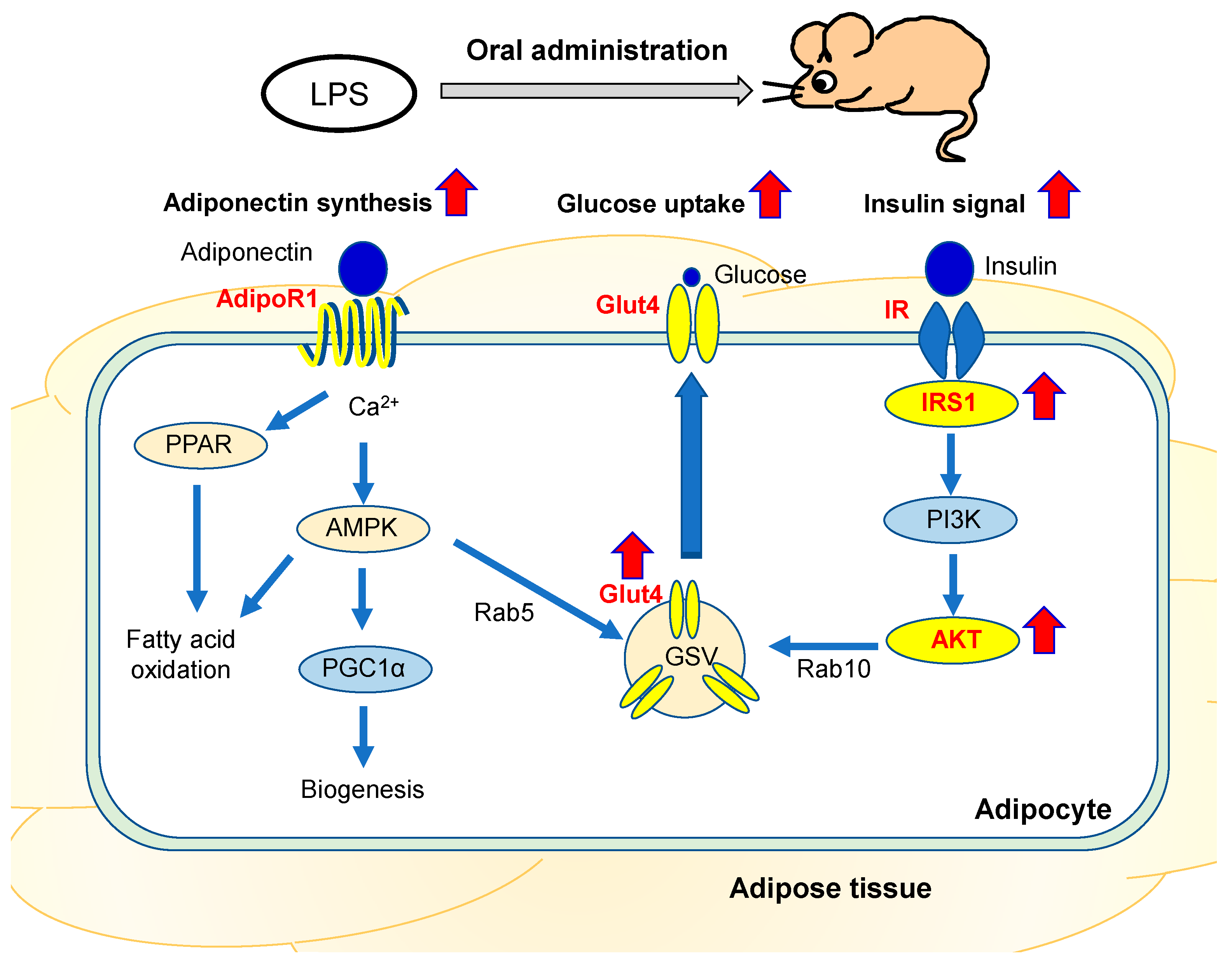
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. OGTTs
4.3. Biochemical Analyses of Serum or Plasma, Whole Blood, and Epididymal ADIPOSE Tissues
4.4. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
4.5. Western Blot Analysis
4.6. Cell Culture
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leuwer, M.; Welters, I.; Marx, G.; Rushton, A.; Bao, H.; Hunter, L.; Trayhurn, P. Endotoxaemia leads to major increases in inflammatory adipokine gene expression in white adipose tissue of mice. Pflug. Arch. 2009, 457, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.N.; McGillicuddy, F.C.; Anderson, P.D.; Hinkle, C.C.; Shah, R.; Pruscino, L.; Tabita-Martinez, J.; Sellers, K.F.; Rickels, M.R.; Reilly, M.P. Experimental endotoxemia induces adipose inflammation and insulin resistance in humans. Diabetes 2010, 59, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anhê, F.F.; Barra, N.G.; Cavallari, J.F.; Henriksbo, B.D.; Schertzer, J.D. Metabolic endotoxemia is dictated by the type of lipopolysaccharide. Cell Rep. 2021, 36, 109691. [Google Scholar] [CrossRef] [PubMed]
- Luche, E.; Cousin, B.; Garidou, L.; Serino, M.; Waget, A.; Barreau, C.; André, M.; Valet, P.; Courtney, M.; Casteilla, L.; et al. Metabolic endotoxemia directly increases the proliferation of adipocyte precursors at the onset of metabolic diseases through a CD14-dependent mechanism. Mol. Metab. 2013, 2, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, A.J.; Rietschel, E.T.; Zähringer, U.; Heine, H. Lipopolysaccharide: Structure, Bioactivity, Receptors, and Signal Transduction. Trends Glycosci. Glycotechnol. 2002, 14, 53–68. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef]
- Festi, D.; Schiumerini, R.; Eusebi, L.H.; Marasco, G.; Taddia, M.; Colecchia, A. Gut microbiota and metabolic syndrome. World J. Gastroenterol. 2014, 20, 16079–16094. [Google Scholar] [CrossRef]
- Soma, G.-I.; Inagawa, H.; Nishizawa, T.; Yoshioka, N.; Taniguchi, Y.; Kohchi, C. Preventative and therapeutic potential of lipopolysaccharide derived from edible Gram-negative bacteria to various diseases. Current. Drug Ther. 2008, 3, 26–32. [Google Scholar] [CrossRef]
- Inagawa, H.; Kobayashi, Y.; Kohchi, C.; Zhang, R.; Shibasaki, Y.; Soma, G. Primed Activation of Macrophages by Oral Administration of Lipopolysaccharide Derived from Pantoea agglomerans. In Vivo 2016, 30, 205–211. [Google Scholar]
- Mizobuchi, H.; Yamamoto, K.; Yamashita, M.; Nakata, Y.; Inagawa, H.; Kohchi, C.; Soma, G.I. Prevention of Diabetes-Associated Cognitive Dysfunction Through Oral Administration of Lipopolysaccharide Derived From Pantoea agglomerans. Front. Immunol. 2021, 12, 650176. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Inagawa, H.; Kohchi, C.; Kazumura, K.; Tsuchiya, H.; Miwa, T.; Okazaki, K.; Soma, G.I. Oral administration of Pantoea agglomerans-derived lipopolysaccharide prevents metabolic dysfunction and Alzheimer's disease-related memory loss in senescence-accelerated prone 8 (SAMP8) mice fed a high-fat diet. PLoS ONE 2018, 13, e0198493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.; Inagawa, H.; Kohchi, C.; Kazumura, K.; Tsuchiya, H.; Miwa, T.; Okazaki, K.; Soma, G.I. Oral administration of Pantoea agglomerans-derived lipopolysaccharide prevents development of atherosclerosis in high-fat diet-fed apoE-deficient mice via ameliorating hyperlipidemia, pro-inflammatory mediators and oxidative responses. PLoS ONE 2018, 13, e0195008. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.Q.; Yu, Y.T.; Jin, M.D.; Duan, H.L.; Huang, G.; Zhang, M.L. Early 1,25-Dihydroxyvitamin D(3) Supplementation Effectively Lowers the Incidence of Type 2 Diabetes Mellitus via Ameliorating Inflammation In KK-A(y) Mice. J. Nutr. Sci. Vitaminol. 2021, 67, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Wang, Y.N.; Ma, L.Y.; Liu, Z.S.; Ye, F.; Yang, J.H. Uncarboxylated osteocalcin ameliorates hepatic glucose and lipid metabolism in KKAy mice via activating insulin signaling pathway. Acta Pharmacol. Sin. 2020, 41, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Marcano, R.; Rojo, M. Pathological and Therapeutic Approach to Endotoxin-Secreting Bacteria Involved in Periodontal Disease. Toxins 2021, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manco, M. Endotoxin as a missed link among all the metabolic abnormalities in the metabolic syndrome. Atherosclerosis 2009, 206, 36. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [Green Version]
- Kawano, M.; Miyoshi, M.; Ogawa, A.; Sakai, F.; Kadooka, Y. Lactobacillus gasseri SBT2055 inhibits adipose tissue inflammation and intestinal permeability in mice fed a high-fat diet. J. Nutr. Sci. 2016, 5, e23. [Google Scholar] [CrossRef] [Green Version]
- Harris, H.W.; Grunfeld, C.; Feingold, K.R.; Read, T.E.; Kane, J.P.; Jones, A.L.; Eichbaum, E.B.; Bland, G.F.; Rapp, J.H. Chylomicrons alter the fate of endotoxin, decreasing tumor necrosis factor release and preventing death. J. Clin. Invest. 1993, 91, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, S.; Witta, J.; Zhong, J.; de Villiers, W.; Eckhardt, E. Chylomicrons promote intestinal absorption of lipopolysaccharides. J. Lipid Res. 2009, 50, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumwenda, Z.L.; Wong, C.B.; Johnson, J.A.; Gosnell, J.E.; Welch, W.J.; Harris, H.W. Chylomicron-bound endotoxin selectively inhibits NF-kappaB activation in rat hepatocytes. Shock 2002, 18, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K. Inbred strains resulting from Japanese mice. Bull Exp. Anim. 1957, 6, 107–112. [Google Scholar]
- Nakamura, M.; Yamada, K. Studies on a diabetic (KK) strain of the mouse. Diabetologia 1967, 3, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Ishida, T.; Jobu, K.; Morisawa, S.; Kawada, K.; Yoshioka, S.; Miyamura, M. Juzentaihoto Suppresses Muscle Atrophy in KKAy Mice. Biol. Pharm. Bull. 2022, 45, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, K.; Nakano, G.; Ehara, N.; Sonoda, K.; Ito, J.; Uchida, H.; Kobayashi, J. Dietary nitrite supplementation improves insulin resistance in type 2 diabetic KKA(y) mice. Nitric. Oxide 2015, 44, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Yamamotoya, T.; Nakatsu, Y.; Ueda, K.; Matsunaga, Y.; Inoue, M.K.; Sakoda, H.; Fujishiro, M.; Ono, H.; Kikuchi, T.; et al. Xanthine Oxidase Inhibitor Febuxostat Exerts an Anti-Inflammatory Action and Protects against Diabetic Nephropathy Development in KK-Ay Obese Diabetic Mice. Int. J. Mol. Sci. 2019, 20, 4680. [Google Scholar] [CrossRef] [Green Version]
- Nakata, K.; Taniguchi, Y.; Yoshioka, N.; Yoshida, A.; Inagawa, H.; Nakamoto, T.; Yoshimura, H.; Miyake, S.; Kohchi, C.; Kuroki, M.; et al. A mixture of Salacia oblonga extract and IP-PA1 reduces fasting plasma glucose (FPG) and low-density lipoprotein (LDL) cholesterol levels. Nutr. Res. Pract. 2011, 5, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Smith, U.; Kahn, B.B. Adipose tissue regulates insulin sensitivity: Role of adipogenesis, de novo lipogenesis and novel lipids. J. Intern. Med. 2016, 280, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Ohlsson, C.; Hammarstedt, A.; Vandenput, L.; Saarinen, N.; Ryberg, H.; Windahl, S.H.; Farman, H.H.; Jansson, J.O.; Movérare-Skrtic, S.; Smith, U.; et al. Increased adipose tissue aromatase activity improves insulin sensitivity and reduces adipose tissue inflammation in male mice. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E450–E462. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, J.; Wang, Z.; Zhang, G.; Yin, J.; Wang, X.; Wang, S.; Yu, Z. Resveratrol reduces the inflammatory response in adipose tissue and improves adipose insulin signaling in high-fat diet-fed mice. PeerJ 2018, 6, e5173. [Google Scholar] [CrossRef] [PubMed]
- Burhans, M.S.; Hagman, D.K.; Kuzma, J.N.; Schmidt, K.A.; Kratz, M. Contribution of Adipose Tissue Inflammation to the Development of Type 2 Diabetes Mellitus. Compr. Physiol. 2018, 9, 1–58. [Google Scholar] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gong, L.; Liu, S.; Zhang, Y.; Zhang, C.; Tian, M.; Lu, H.; Bu, P.; Yang, J.; Ouyang, C.; et al. Adipose HuR protects against diet-induced obesity and insulin resistance. Nat. Commun. 2019, 10, 2375. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Lee, H.; Han, J.; Lee, H.; Kattia, R.O.; Nelson, Z.V.; Choi, S.; Kim, S.-Y.; Park, H.-Y.; Jeong, H.G.; et al. Viburnum stellato-tomentosum Extract Suppresses Obesity and Hyperglycemia through Regulation of Lipid Metabolism in High-Fat Diet-Fed Mice. Molecules 2021, 26, 1052. [Google Scholar] [CrossRef]
- Hauffe, R.; Stein, V.; Chudoba, C.; Flore, T.; Rath, M.; Ritter, K.; Schell, M.; Wardelmann, K.; Deubel, S.; Kopp, J.F.; et al. GPx3 dysregulation impacts adipose tissue insulin receptor expression and sensitivity. JCI Insight 2020, 5, e136283. [Google Scholar] [CrossRef]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef]
- Softic, S.; Boucher, J.; Solheim, M.H.; Fujisaka, S.; Haering, M.F.; Homan, E.P.; Winnay, J.; Perez-Atayde, A.R.; Kahn, C.R. Lipodystrophy Due to Adipose Tissue-Specific Insulin Receptor Knockout Results in Progressive NAFLD. Diabetes 2016, 65, 2187–2200. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [CrossRef]
- Carvalho, E.; Kotani, K.; Peroni, O.D.; Kahn, B.B. Adipose-specific overexpression of GLUT4 reverses insulin resistance and diabetes in mice lacking GLUT4 selectively in muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E551–E561. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Aprahamian, T.R. Elevated adiponectin expression promotes adipose tissue vascularity under conditions of diet-induced obesity. Metabolism 2013, 62, 1730–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Luo, N.; Klein, R.L.; Garvey, W.T. Adiponectin promotes adipocyte differentiation, insulin sensitivity, and lipid accumulation. J. Lipid Res. 2005, 46, 1369–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chen, Q.; Pu, H.; Wei, Q.; Duan, M.; Zhang, C.; Jiang, T.; Shou, X.; Zhang, J.; Yang, Y. Adiponectin improves NF-κB-mediated inflammation and abates atherosclerosis progression in apolipoprotein E-deficient mice. Lipids Health Dis. 2016, 15, 33. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Perakakis, N.; Gong, H.; Chamberland, J.P.; Brinkoetter, M.T.; Hamnvik, O.P.R.; Mantzoros, C.S. Adiponectin administration prevents weight gain and glycemic profile changes in diet-induced obese immune deficient Rag1-/- mice lacking mature lymphocytes. Metabolism 2016, 65, 1720–1730. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Yamashita, M.; Inagawa, H.; Kohchi, C.; Soma, G.I. Anti-inflammatory and Insulin Signaling Phenotype Induced by Repeated Lipopolysaccharide Stimulation in 3T3-L1 Adipocytes. Anticancer. Res. 2022, 42, 3983–3991. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank ID | Target Gene | Primer | Primer Sequence (5′–3′) |
|---|---|---|---|
| NM_009605 | Adipoq | F | TGTTCCTCTTAATCCTGCCCA |
| R | CCAACCTGCACAAGTTCCCTT | ||
| NM_001306069 | Adipor1 | F | AACGGGCCATCCATTTTTG |
| R | TTAGCCGGGCTACATCAAGG | ||
| NM_197985 | Adipor2 | F | CCACACAACACAAGAATCCG |
| R | CCCTTCTTCTTGGGAGAATGG | ||
| NM_001110208 | Akt2 | F | ACGTGGTGAATACATCAAGACC |
| R | GCTACAGAGAAATTGTTCAGGGG | ||
| NM_001289726 | Gapdh | F | CGACTTCAACAGCAACTCCCACTCTTCC |
| R | TGGGTGGTCCAGGGTTTCTTACTCCTT | ||
| NM_001359114 | Glut4 | F | GTAACTTCATTGTCGGCATGG |
| R | AGCTGAGATCTGGTCAAACG | ||
| NM_008361 | Il1b | F | GAAAGACGGCACACCCACCCT |
| R | GCTCTGCTTGTGAGGTGCTGATGTA | ||
| NM_031168 | Il6 | F | CCAGAGATACAAAGAAATGATGG |
| R | ACTCCAGAAGACCAGAGGAAAT | ||
| NM_001303244 | Il12b | F | ACAGCACCAGCTTCTTCATCAG |
| R | TCTTCAAAGGCTTCATCTGCAA | ||
| NM_010568 | Ir | F | TTTGTCATGGATGGAGGCTA |
| R | CCTCATCTTGGGGTTGAACT | ||
| NM_010570 | Irs1 | F | CCATGAGCGATGAGTTTCGC |
| R | GCAGTGATGCTCTCAGTTCG | ||
| NM_011333 | Mcp1 | F | AACTGCATCTGCCCTAAGGT |
| R | ACTGTCACACTGGTCACTCC | ||
| NM_013693 | TNFα | F | CTGTGAAGGGAATGGGTGTT |
| R | GGTCACTGTCCCAGCATCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, K.; Yamashita, M.; Oda, M.; Tjendana Tjhin, V.; Inagawa, H.; Soma, G.-I. Oral Administration of Lipopolysaccharide Enhances Insulin Signaling-Related Factors in the KK/Ay Mouse Model of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2023, 24, 4619. https://doi.org/10.3390/ijms24054619
Yamamoto K, Yamashita M, Oda M, Tjendana Tjhin V, Inagawa H, Soma G-I. Oral Administration of Lipopolysaccharide Enhances Insulin Signaling-Related Factors in the KK/Ay Mouse Model of Type 2 Diabetes Mellitus. International Journal of Molecular Sciences. 2023; 24(5):4619. https://doi.org/10.3390/ijms24054619
Chicago/Turabian StyleYamamoto, Kazushi, Masashi Yamashita, Masataka Oda, Vindy Tjendana Tjhin, Hiroyuki Inagawa, and Gen-Ichiro Soma. 2023. "Oral Administration of Lipopolysaccharide Enhances Insulin Signaling-Related Factors in the KK/Ay Mouse Model of Type 2 Diabetes Mellitus" International Journal of Molecular Sciences 24, no. 5: 4619. https://doi.org/10.3390/ijms24054619