
Platelets, Protean Cells with All-Around Functions and Multifaceted Pharmacological Applications
 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
1.1. Platelet Granules and Receptors
1.1.1. α-Granules
1.1.2. Dense Granules
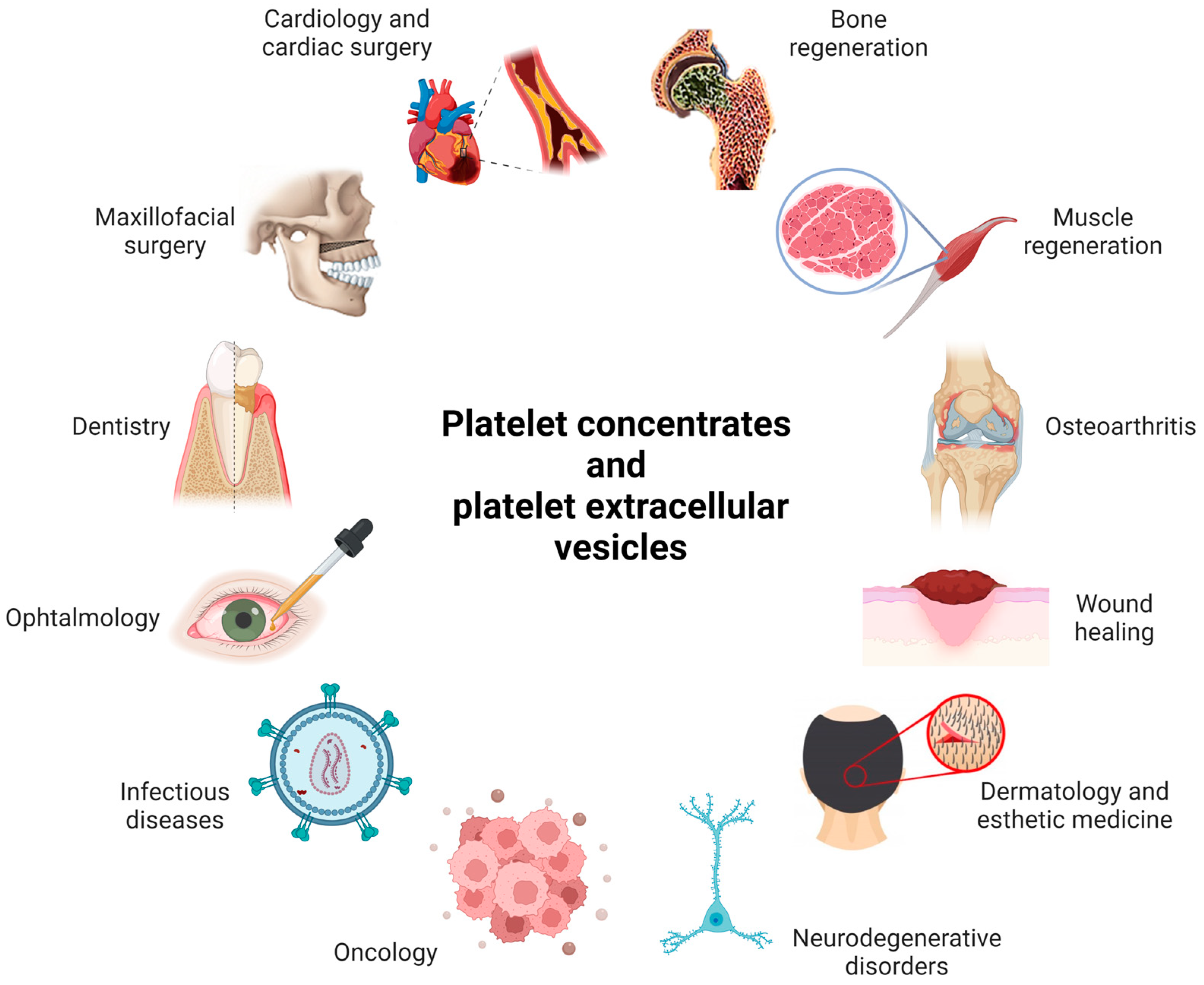
2. Platelet Derivatives
2.1. Examples of Medical Applications of PCs
2.2. Limitations in the Use of PCs
3. Platelet Role in Disease
3.1. Platelets and Atherosclerosis
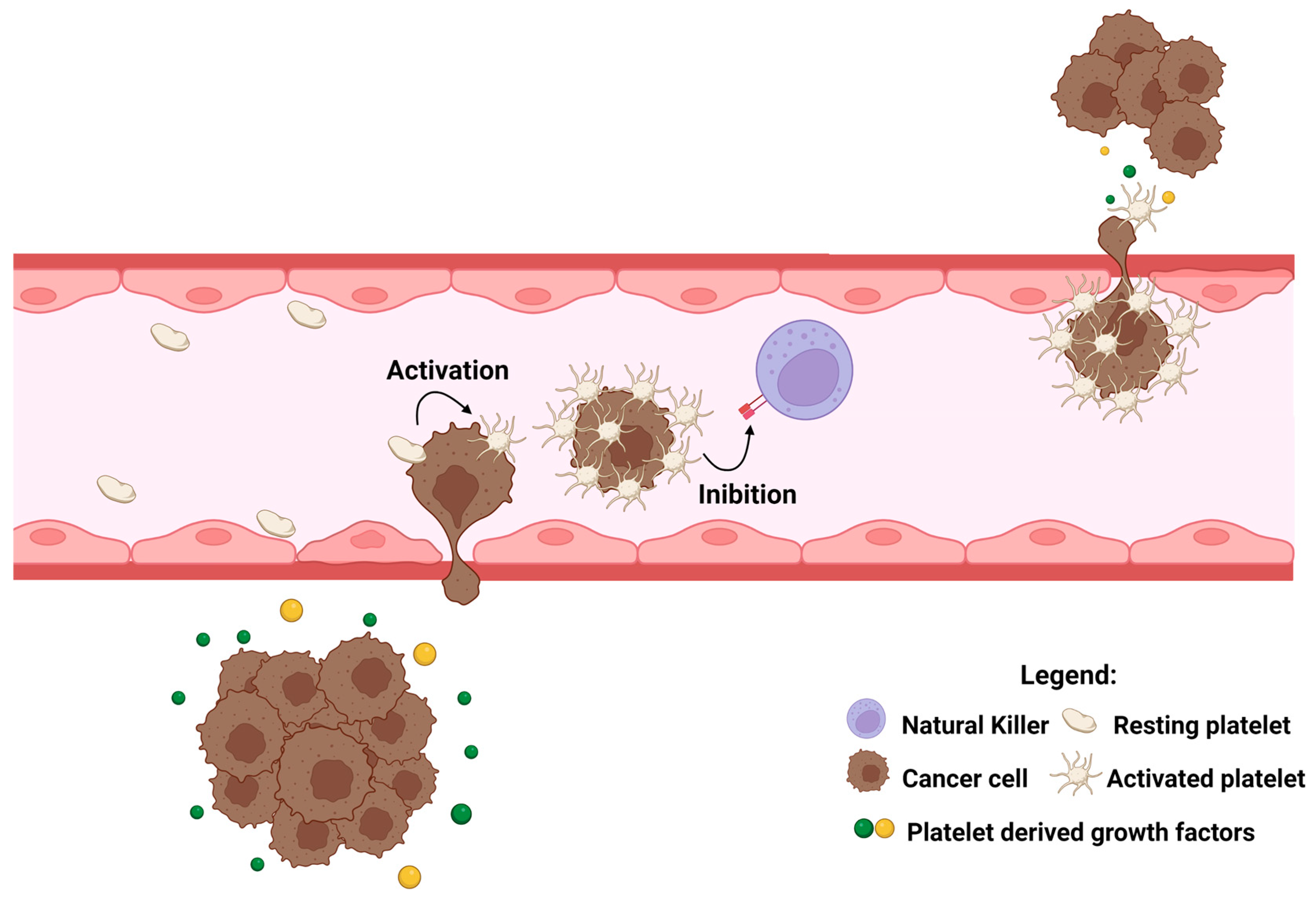
3.2. Platelets and Cancer
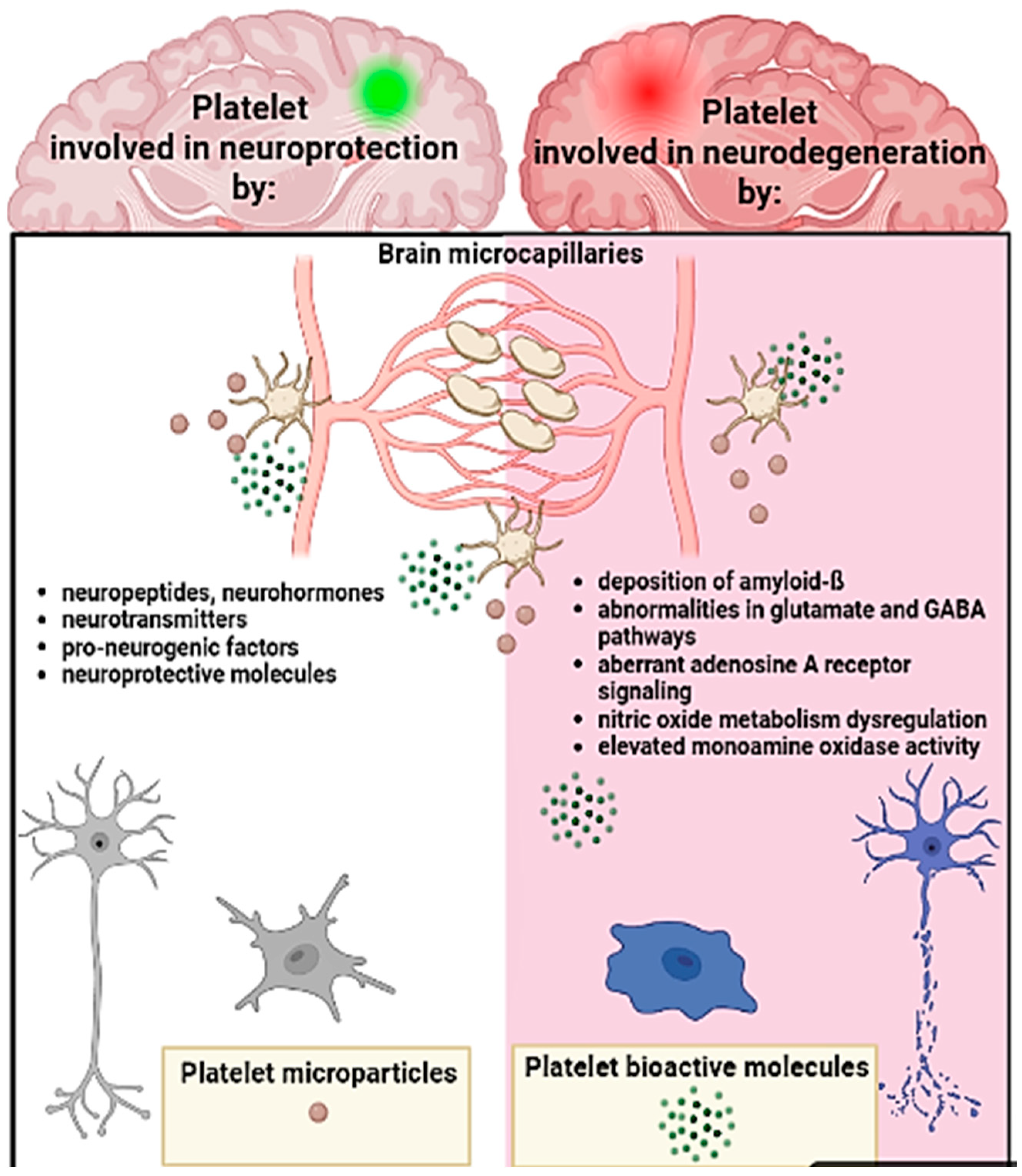
3.3. Platelets, the Brain, and Neurodegenerative Conditions
4. Platelet Extracellular Vesicles
4.1. General Overview of Extracellular Vesicles and Their Clinical Applications
4.2. From “Platelet Dust” to pEV Full Recognition
4.3. pEV Isolation, Characterization, and Production Methods
4.4. Therapeutic Applications of pEVs
5. Therapeutic Hitchhiking: Platelets as Drug Delivery Vehicles
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rolla, R.; Puricelli, C.; Bertoni, A.; Boggio, E.; Gigliotti, C.L.; Chiocchetti, A.; Cappellano, G.; Dianzani, U. Platelets: “multiple choice” effectors in the immune response and their implication in COVID-19 thromboinflammatory process. Int. J. Lab. Hematol. 2021, 43, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Diacovo, T.G.; Roth, S.J.; Buccola, J.M.; Bainton, D.F.; Springer, T.A. Neutrophil Rolling, Arrest, and Transmigration Across Activated, Surface-Adherent Platelets Via Sequential Action of P-Selectin and the &-Integrin CDllb/CDl8. Blood 1996, 88, 146–157. [Google Scholar] [PubMed]
- Shiraki, R.; Inoue, N.; Kawasaki, S.; Takei, A.; Kadotani, M.; Ohnishi, Y.; Ejiri, J.; Kobayashi, S.; Hirata, K.I.; Kawashima, S.; et al. Expression of Toll-like receptors on human platelets. Thromb. Res. 2004, 113, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Clancy, L.; Corkrey, H.A.; Freedman, J.E. Circulating platelets as mediators of immunity, inflammation, and thrombosis. Circ. Res. 2018, 122, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Boilard, E.; Paré, G.; Rousseau, M.; Cloutier, N.; Dubuc, I.; Lévesque, T.; Borgeat, P.; Flamand, L. Influenza virus H1N1 activates platelets through FcγRIIA signaling and thrombin generation. Blood 2014, 123, 2854–2863. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Tourdot, B.E.; Fernandez-Perez, P.; Vesci, J.; Ren, J.; Smyrniotis, C.J.; Luci, D.K.; Jadhav, A.; Simeonov, A.; Maloney, D.J.; et al. Platelet 12-LOX is essential for FcγRIIa-mediated platelet activation. Blood 2014, 124, 2271–2279. [Google Scholar] [CrossRef] [Green Version]
- Holinstat, M. Normal platelet function. Cancer Metastasis Rev. 2017, 36, 198. [Google Scholar] [CrossRef]
- McDonald, B.; Urrutia, R.; Yipp, B.G.; Jenne, C.N.; Kubes, P. Intravascular neutrophil extracellular traps capture bacteria from the bloodstream during sepsis. Cell Host Microbe 2012, 12, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Ouseph, M.M.; Huang, Y.; Banerjee, M.; Joshi, S.; MacDonald, L.; Zhong, Y.; Liu, H.; Li, X.; Xiang, B.; Zhang, G.; et al. Autophagy is induced upon platelet activation and is essential for hemostasis and thrombosis. Blood 2015, 126, 1233. [Google Scholar] [CrossRef] [Green Version]
- Yeung, J.; Li, W.; Holinstat, M. Platelet Signaling and Disease: Targeted Therapy for Thrombosis and Other Related Diseases. Pharmacol. Rev. 2018, 70, 526–548. [Google Scholar] [CrossRef] [Green Version]
- Rendu, F.; Brohard-Bohn, B. The platelet release reaction: Granules’ constituents, secretion and functions. Platelets 2001, 12, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Maynard, D.M.; Heijnen, H.F.G.; Horne, M.K.; White, J.G.; Gahl, W.A. Proteomic analysis of platelet α-granules using mass spectrometry. J. Thromb. Haemost. 2007, 5, 1945–1955. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.; Masse, J.; Cramer, E. Alpha-granule membrane mirrors the platelet plasma membrane and contains the glycoproteins Ib, IX, and V. Blood 1996, 87, 1385–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Murasaki, K.; Kodama, K.; Takayama, H. Intracellular localization of glycoprotein VI in human platelets and its surface expression upon activation. Br. J. Haematol. 2003, 121, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Coppinger, J.A.; Cagney, G.; Toomey, S.; Kislinger, T.; Belton, O.; McRedmond, J.P.; Cahill, D.J.; Emili, A.; Fitzgerald, D.J.; Maguire, P.B. Characterization of the proteins released from activated platelets leads to localization of novel platelet proteins in human atherosclerotic lesions. Blood 2004, 103, 2096–2104. [Google Scholar] [CrossRef] [Green Version]
- Falati, S.; Liu, Q.; Gross, P.; Merrill-Skoloff, G.; Chou, J.; Vandendries, E.; Celi, A.; Croce, K.; Furie, B.C.; Furie, B. Accumulation of Tissue Factor into Developing Thrombi In Vivo Is Dependent upon Microparticle P-Selectin Glycoprotein Ligand 1 and Platelet P-Selectin. J. Exp. Med. 2003, 197, 1598. [Google Scholar] [CrossRef]
- Kasper, B.; Brandt, E.; Bulfone-Paus, S.; Petersen, F. Platelet factor 4 (PF-4)–induced neutrophil adhesion is controlled by src-kinases, whereas PF-4–mediated exocytosis requires the additional activation of p38 MAP kinase and phosphatidylinositol 3-kinase. Blood 2004, 103, 1602–1610. [Google Scholar] [CrossRef]
- Kasper, B.; Brandt, E.; Ernst, M.; Petersen, F. Neutrophil adhesion to endothelial cells induced by platelet factor 4 requires sequential activation of Ras, Syk, and JNK MAP kinases. Blood 2006, 107, 1768–1775. [Google Scholar] [CrossRef]
- Nomura, S.; Ishii, K.; Kanazawa, S.; Inami, N.; Kamitsuji, Y.; Uoshima, N.; Ishida, H.; Yoshihara, T.; Kitayama, H.; Hayashi, K. Role of platelet-derived chemokines (RANTES and ENA-78) after stem cell transplantation. Transpl. Immunol. 2006, 15, 247–253. [Google Scholar] [CrossRef]
- Smith, D.F.; Galkina, E.; Ley, K.; Huo, Y. GRO family chemokines are specialized for monocyte arrest from flow. Am. J. Physiol. Circ. Physiol. 2005, 289, H1976–H1984. [Google Scholar] [CrossRef]
- Cha, J.K.; Jeong, M.H.; Bae, H.R.; Han, J.Y.; Jeong, S.J.; Jin, H.J.; Lim, Y.J.; Kim, S.H.; Kim, J.W. Activated platelets induce secretion of interleukin-1beta, monocyte chemotactic protein-1, and macrophage inflammatory protein-1alpha and surface expression of intercellular adhesion molecule-1 on cultured endothelial cells. J. Korean Med. Sci. 2000, 15, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleissner, C.A.; Von Hundelshausen, P.; Ley, K. Platelet chemokines in vascular disease. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.P.; Ataga, K.I.; Orringer, E.P.; Phillips, D.R.; Parise, L.V. Biologically Active CD40 Ligand Is Elevated in Sickle Cell Anemia. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Klesney-Tait, J.; Turnbull, I.R.; Colonna, M. The TREM receptor family and signal integration. Nat. Immunol. 2006, 7, 1266–1273. [Google Scholar] [CrossRef]
- Haselmayer, P.; Grosse-Hovest, L.; von Landenberg, P.; Schild, H.; Radsak, M.P. TREM-1 ligand expression on platelets enhances neutrophil activation. Blood 2007, 110, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Au, A.E.; Josefsson, E.C. Regulation of platelet membrane protein shedding in health and disease. Platelets 2017, 28, 342–353. [Google Scholar] [CrossRef]
- Ruiz, F.A.; Lea, C.R.; Oldfield, E.; Docampo, R. Human Platelet Dense Granules Contain Polyphosphate and Are Similar to Acidocalcisomes of Bacteria and Unicellular Eukaryotes. J. Biol. Chem. 2004, 279, 44250–44257. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.; Mutch, N.J.; Schenk, W.A.; Smith, S.A.; Esterl, L.; Spronk, H.M.; Schmidbauer, S.; Gahl, W.A.; Morrissey, J.H.; Renné, T. Platelet polyphosphates are proinflammatory and procoagulant mediators in vivo. Cell 2009, 139, 1156. [Google Scholar] [CrossRef] [Green Version]
- Youssefian, T.; Massé, J.M.; Rendu, F.; Guichard, J.; Cramer, E.M. Platelet and Megakaryocyte Dense Granules Contain Glycoproteins Ib and IIb-IIIa. Blood 1997, 89, 4047–4057. [Google Scholar] [CrossRef] [Green Version]
- Jenne, C.N.; Urrutia, R.; Kubes, P. Platelets: Bridging hemostasis, inflammation, and immunity. Int. J. Lab. Hematol. 2013, 35, 254–261. [Google Scholar] [CrossRef]
- Mohammadi, M.; Zinkle, A. A threshold model for receptor tyrosine kinase signaling specificity and cell fate determination. F1000Research 2018, 7, 872. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Tang, X.Y.; Qu, Z.Y.; Sun, Z.W.; Ji, C.F.; Li, Y.J.; Guo, S.D. Targeting the PDGF/PDGFR signaling pathway for cancer therapy: A review. Int. J. Biol. Macromol. 2022, 202, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Khan, M.I.; Wang, J.; Ali, R.; Ali, S.W.; Kazmi, A.; Lolai, A.; Huang, Y.L.; Hussain, A.; Bilal, M.; et al. Role of receptor tyrosine kinases mediated signal transduction pathways in tumor growth and angiogenesis—New insight and futuristic vision. Int. J. Biol. Macromol. 2021, 180, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Piccin, A.; Di Pierro, A.M.; Canzian, L.; Primerano, M.; Corvetta, D.; Negri, G.; Mazzoleni, G.; Gastl, G.; Steurer, M.; Gentilini, I.; et al. Platelet gel: A new therapeutic tool with great potential. Blood Transfus. 2017, 15, 333–340. [Google Scholar] [CrossRef]
- Everts, P.A.M.; Knape, J.T.A.; Weibrich, G.; Schönberger, J.P.A.M.; Hoffmann, J.; Overdevest, E.P.; Box, H.A.M.; Van Zundert, A. Platelet-Rich Plasma and Platelet Gel: A Review. J. Extra. Corpor. Technol. 2006, 38, 187. [Google Scholar]
- Piccin, A.; Di Pierro, A.M.; Calabrese, L.; Fontanella, F.; Daves, M. Platelet gel: The “holy water” of regenerative medicine. La Riv. Ital. della Med. di Lab.-Ital. J. Lab. Med. 2018 144 2018, 14, 201–202. [Google Scholar] [CrossRef]
- De Pascale, M.R.; Sommese, L.; Casamassimi, A.; Napoli, C. Platelet Derivatives in Regenerative Medicine: An Update. Transfus. Med. Rev. 2015, 29, 52–61. [Google Scholar] [CrossRef]
- Cicha, I.; Garlichs, C.D.; Daniel, W.G.; Goppelt-Struebe, M. Activated human platelets release connective tissue growth factor. Thromb. Haemost. 2004, 91, 755–760. [Google Scholar] [CrossRef]
- Dhurat, R.; Sukesh, M. Principles and Methods of Preparation of Platelet-Rich Plasma: A Review and Author’s Perspective. J. Cutan. Aesthet. Surg. 2014, 7, 197. [Google Scholar] [CrossRef]
- Zamani, M.; Yaghoubi, Y.; Movassaghpour, A.; Shakouri, K.; Mehdizadeh, A.; Pishgahi, A.; Yousefi, M. Novel therapeutic approaches in utilizing platelet lysate in regenerative medicine: Are we ready for clinical use? J. Cell. Physiol. 2019, 234, 17172–17186. [Google Scholar] [CrossRef]
- Chacón-Fernández, P.; Säuberli, K.; Colzani, M.; Moreau, T.; Ghevaert, C.; Barde, Y.A. Brain-derived Neurotrophic Factor in Megakaryocytes. J. Biol. Chem. 2016, 291, 9881. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 387. [Google Scholar] [CrossRef]
- Chicharro-Alcántara, D.; Rubio-Zaragoza, M.; Damiá-Giménez, E.; Carrillo-Poveda, J.M.; Cuervo-Serrato, B.; Peláez-Gorrea, P.; Sopena-Juncosa, J.J. Platelet Rich Plasma: New Insights for Cutaneous Wound Healing Management. J. Funct. Biomater. 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusti, I.; D’Ascenzo, S.; Mancò, A.; Di Stefano, G.; Di Francesco, M.; Rughetti, A.; Dal Mas, A.; Properzi, G.; Calvisi, V.; Dolo, V. Platelet Concentration in Platelet-Rich Plasma Affects Tenocyte Behavior In Vitro. BioMed Res. Int. 2014, 2014, 630870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieudonné, S.C.; Foo, P.; Van Zoelen, E.J.J.; Burger, E.H. Inhibiting and stimulating effects of TGF-beta 1 on osteoclastic bone resorption in fetal mouse bone organ cultures. J. Bone Miner. Res. 1991, 6, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Weibrich, G.; Hansen, T.; Kleis, W.; Buch, R.; Hitzler, W.E. Effect of platelet concentration in platelet-rich plasma on peri-implant bone regeneration. Bone 2004, 34, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Zumarán, C.C.; Parra, M.V.; Olate, S.A.; Fernández, E.G.; Muñoz, F.T.; Haidar, Z.S. The 3 R’s for Platelet-Rich Fibrin: A “Super” Tri-Dimensional Biomaterial for Contemporary Naturally-Guided Oro-Maxillo-Facial Soft and Hard Tissue Repair, Reconstruction and Regeneration. Materials 2018, 11, 1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohan Ehrenfest, D.M.; Rasmusson, L.; Albrektsson, T. Classification of platelet concentrates: From pure platelet-rich plasma (P-PRP) to leucocyte- and platelet-rich fibrin (L-PRF). Trends Biotechnol. 2009, 27, 158–167. [Google Scholar] [CrossRef]
- Del Fante, C.; Perotti, C.; Bonferoni, M.C.; Rossi, S.; Sandri, G.; Ferrari, F.; Scudeller, L.; Caramella, C.M. Platelet Lysate Mucohadesive Formulation to Treat Oral Mucositis in Graft Versus Host Disease Patients: A New Therapeutic Approach. AAPS PharmSciTech 2011, 12, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Passaretti, F.; Tia, M.; D’esposito, V.; De Pascale, M.; Del Corso, M.; Sepulveres, R.; Liguoro, D.; Valentino, R.; Beguinot, F.; Formisano, P.; et al. Growth-promoting action and growth factor release by different platelet derivatives. Platelets 2014, 25, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Choukroun, J.I.; Braccini, F.; Diss, A.; Giordano, G.; Doglioli, P.; Dohan, D.M. Influence of platelet rich fibrin (PRF) on proliferation of human preadipocytes and tympanic keratinocytes: A new opportunity in facial lipostructure (Coleman’s technique) and tympanoplasty? Rev. Laryngol. Otol. Rhinol. 2007, 128, 27–32. [Google Scholar] [PubMed]
- Diss, A.; Dohan, D.M.; Mouhyi, J.; Mahler, P. Osteotome sinus floor elevation using Choukroun’s platelet-rich fibrin as grafting material: A 1-year prospective pilot study with microthreaded implants. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2008, 105, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Del Corso, M.; Vervelle, A.; Simonpieri, A.; Jimbo, R.; Inchingolo, F.; Sammartino, G.; M. Dohan Ehrenfest, D. Current Knowledge and Perspectives for the Use of Platelet-Rich Plasma (PRP) and Platelet-Rich Fibrin (PRF) in Oral and Maxillofacial Surgery Part 1: Periodontal and Dentoalveolar Surgery. Curr. Pharm. Biotechnol. 2012, 13, 1207–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonpieri, A.; Del Corso, M.; Vervelle, A.; Jimbo, R.; Inchingolo, F.; Sammartino, G.; M. Dohan Ehrenfest, D. Current knowledge and perspectives for the use of platelet-rich plasma (PRP) and platelet-rich fibrin (PRF) in oral and maxillofacial surgery part 2: Bone graft, implant and reconstructive surgery. Curr. Pharm. Biotechnol. 2012, 13, 1231–1256. [Google Scholar] [CrossRef]
- Braccini, F.; Dohan, D.M. The relevance of Choukroun’s platelet rich fibrin (PRF) during facial aesthetic lipostructure (Coleman’s technique): Preliminary results. Rev. Laryngol. Otol. Rhinol. 2007, 128, 255–260. [Google Scholar]
- Aprili, G.; Gandini, G.; Guaschino, R.; Mazzucco, L.; Salvaneschi, L.; Vaglio, S. SIMTI recommendations on blood components for non-transfusional use. Blood Transfus. 2013, 11, 622. [Google Scholar] [CrossRef]
- Rodgers, G.M. Immune-mediated coagulopathy associated with topical bovine thrombin: Review of the pediatric literature. HJournal Pediatr. Hematol. 2011, 33, 86–88. [Google Scholar] [CrossRef]
- Clark, J.; Crean, S.; Reynolds, M.W. Topical bovine thrombin and adverse events: A review of the literature. Curr. Med. Res. Opin. 2008, 24, 2071–2087. [Google Scholar] [CrossRef]
- Harrison, S.; Vavken, P.; Kevy, S.; Jacobson, M.; Zurakowski, D.; Murray, M.M. Platelet activation by collagen provides sustained release of anabolic cytokines. Am. J. Sport. Med. 2011, 39, 729–734. [Google Scholar] [CrossRef]
- Geerling, G.; MacLennan, S.; Hartwig, D. Autologous serum eye drops for ocular surface disorders. Br. J. Ophthalmol. 2004, 88, 1474. [Google Scholar] [CrossRef] [Green Version]
- Quinto, G.G.; Campos, M.; Behrens, A. Autologous serum for ocular surface diseases. Arq. Bras. Oftalmol. 2008, 71, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geremicca, W.; Fonte, C.; Vecchio, S. Blood components for topical use in tissue regeneration: Evaluation of corneal lesions treated with platelet lysate and considerations on repair mechanisms. Blood Transfus. 2010, 8, 112. [Google Scholar] [CrossRef]
- Harmon, K.; Hanson, R.; Bowen, J.; Greenberg, S.; Magaziner, E.; Vandenbosch, J.; Harshfield, D.; Shiple, B.; Audley, D. Guidelines for the Use of Platelet Rich Plasma—Draft. Int. Cell. Med. Soc. 2011. Available online: https://www.scribd.com/document/159334949/206-ICMS-Guidelines-for-the-Use-of-Platelet-Rich-Plasma-Draftob-oasbonasdandbowndoww (accessed on 24 December 2022).
- Ferrari, M.; Zia, S.; Valbonesi, M.; Henriquet, F.; Venere, G.; Spagnolo, S.; Grasso, M.A.; Panzani, I. A new technique for hemodilution, preparation of autologous platelet-rich plasma and intraoperative blood salvage in cardiac surgery. Int. J. Artif. Organs 1987, 10, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.C.; Jeevanandam, V.; Smith, C.R.; Williams, M.R.; Kaynar, A.M.; Frank, R.A.; Mosca, R.; Reiss, R.F.; Rose, E.A. Autologous fibrin glue from intraoperatively collected platelet-rich plasma. Ann. Thorac. Surg. 1992, 53, 530–531. [Google Scholar] [CrossRef] [PubMed]
- Tawes, R.L.; Sydorak, G.R.; DuVall, T.B. Autologous fibrin glue: The last step in operative hemostasis. Am. J. Surg. 1994, 168, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Piccin, A.; Di Pierro, A.M.; Tagnin, M.; Russo, C.; Fustos, R.; Corvetta, D.; Primerano, M.; Magri, E.; Conci, V.; Gentilini, I.; et al. Healing of a soft tissue wound of the neck and jaw osteoradionecrosis using platelet gel. Regen. Med. 2016, 11, 459–463. [Google Scholar] [CrossRef] [PubMed]
- San Sebastian, K.M.; Lobato, I.; Hernández, I.; Burgos-Alonso, N.; Gomez-Fernandez, M.C.; López, J.L.; Rodríguez, B.; March, A.G.; Grandes, G.; Andia, I. Efficacy and safety of autologous platelet rich plasma for the treatment of vascular ulcers in primary care: Phase III study. BMC Fam. Pract. 2014, 15, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Merchán, W.H.; Gómez, L.A.; Chasoy, M.E.; Alfonso-Rodríguez, C.A.; Muñoz, A.L. Platelet-rich plasma, a powerful tool in dermatology. J. Tissue Eng. Regen. Med. 2019, 13, 892–901. [Google Scholar] [CrossRef]
- McCarrel, T.M.; Mall, N.A.; Lee, A.S.; Cole, B.J.; Butty, D.C.; Fortier, L.A. Considerations for the use of platelet-rich plasma in orthopedics. Sport. Med. 2014, 44, 1025–1036. [Google Scholar] [CrossRef]
- L. Alio, J.; Arnalich-Montiel, F.; E. Rodriguez, A. The role of “eye platelet rich plasma” (E-PRP) for wound healing in ophthalmology. Curr. Pharm. Biotechnol. 2012, 13, 1257–1265. [Google Scholar] [CrossRef]
- Ye, F.; Li, H.; Qiao, G.; Chen, F.; Tao, H.; Ji, A.; Hu, Y. Platelet-rich plasma gel in combination with Schwann cells for repair of sciatic nerve injury. Neural Regen. Res. 2012, 7, 2292. [Google Scholar] [CrossRef]
- Chou, M.L.; Wu, J.W.; Gouel, F.; Jonneaux, A.; Tillerman, K.; Renn, T.Y.; Laloux, C.; Chang, H.M.; Lin, L.T.; Devedjian, J.C.; et al. Tailor-made purified human platelet lysate concentrated in neurotrophins for treatment of Parkinson’s disease. Biomaterials 2017, 142, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Krupski, W.; Reilly, L.; Perez, S.; Moss, K.; Crombleholme, P.; Rapp, J. A prospective randomized trial of autologous platelet-derived wound healing factors for treatment of chronic nonhealing wounds: A preliminary report. J. Vasc. Surg. 1991, 14, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.J.; Fylling, C.P.; Parnell, L.K.S. Use of Platelet Rich Plasma Gel on Wound Healing: A Systematic Review and Meta-Analysis. Eplasty 2011, 11, e38. [Google Scholar]
- Alsousou, J.; Thompson, M.; Hulley, P.; Noble, A.; Willett, K. The biology of platelet-rich plasma and its application in trauma and orthopaedic surgery: A review of the literature. J. Bone Jt. Surgery. 2009, 91, 987–996. [Google Scholar] [CrossRef]
- Forni, F.; Marzagalli, M.; Tesei, P.; Grassi, A. Platelet gel: Applications in dental regenerative surgery. Blood Transfus. 2013, 11, 102–107. [Google Scholar] [CrossRef]
- Kim, M.; Won, J.Y.; Choi, S.Y.; Kim, M.; Ra, H.; Jee, D.; Kwon, J.W.; Kang, K.D.; Roh, Y.J.; Park, Y.G.; et al. Therapeutic efficacy of autologous platelet concentrate injection on macular holes with high myopia, large macular holes, or recurrent macular holes: A multicenter randomized controlled trial. J. Clin. Med. 2021, 10, 2727. [Google Scholar] [CrossRef]
- Fox, R.I.; Chan, R.; Michelson, J.B.; Belmont, J.B.; Michelson, P.E. Beneficial effect of artificial tears made with autologous serum in patients with keratoconjunctivitis sicca. Arthritis Rheum. 1984, 27, 459–461. [Google Scholar] [CrossRef]
- De Vos, R.J.; Windt, J.; Weir, A. Strong evidence against platelet-rich plasma injections for chronic lateral epicondylar tendinopathy: A systematic review. Br. J. Sport. Med. 2014, 48, 952–956. [Google Scholar] [CrossRef] [Green Version]
- De Vos, R.J.; Weir, A.; Van Schie, H.T.M.; Bierma-Zeinstra, S.M.A.; Verhaar, J.A.N.; Weinans, H.; Tol, J.L. Platelet-rich plasma injection for chronic Achilles tendinopathy: A randomized controlled trial. JAMA 2010, 303, 144–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuffler, D.P. Platelet-rich plasma and the elimination of neuropathic pain. Mol. Neurobiol. 2013, 48, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Malahias, M.A.; Johnson, E.O.; Babis, G.C.; Nikolaou, V.S. Single injection of platelet-rich plasma as a novel treatment of carpal tunnel syndrome. Neural Regen. Res. 2015, 10, 1856–1859. [Google Scholar] [CrossRef] [PubMed]
- Centeno, C.; Markle, J.; Dodson, E.; Stemper, I.; Hyzy, M.; Williams, C.; Freeman, M. The use of lumbar epidural injection of platelet lysate for treatment of radicular pain. J. Exp. Orthop. 2017, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Crovetti, G.; Martinelli, G.; Issi, M.; Barone, M.; Guizzardi, M.; Campanati, B.; Moroni, M.; Carabelli, A. Platelet gel for healing cutaneous chronic wounds. Transfus. Apher. Sci. 2004, 30, 145–151. [Google Scholar] [CrossRef]
- Antich-Rosselló, M.; Forteza-Genestra, M.A.; Monjo, M.; Ramis, J.M. Platelet-Derived Extracellular Vesicles for Regenerative Medicine. Int. J. Mol. Sci. 2021, 22, 8580. [Google Scholar] [CrossRef]
- Phipps, R.P. Atherosclerosis: The emerging role of inflammation and the CD40-CD40 ligand system. Proc. Natl. Acad. Sci. USA 2000, 97, 6930–6932. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Ley, K.F. Role of Platelets in the Development of Atherosclerosis. Trends Cardiovasc. Med. 2004, 14, 18–22. [Google Scholar] [CrossRef]
- Fitzgerald, D.J.; Roy, L.; Catella, F.; FitzGerald, G.A. Platelet activation in unstable coronary disease. N. Engl. J. Med. 1986, 315, 983–989. [Google Scholar] [CrossRef]
- Steinhubl, S.R.; Badimon, J.J.; Bhatt, D.L.; Herbert, J.M.; Lüscher, T. Clinical evidence for anti-inflammatory effects of antiplatelet therapy in patients with atherothrombotic disease. Vasc. Med. 2007, 12, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Cohen Arazi, H.; Badimon, J.J. Anti-inflammatory effects of anti-platelet treatment in atherosclerosis. Curr. Pharm. Des. 2012, 18, 4311–4325. [Google Scholar] [CrossRef]
- Iyengar, S.; Rabbani, L.R.E. Beyond platelet inhibition: Potential pleiotropic effects of ADP-receptor antagonists. J. Thromb. Thrombolysis 2009, 27, 300–306. [Google Scholar] [CrossRef]
- Klinkhardt, U.; Bauersachs, R.; Adams, J.; Graff, J.; Lindhoff-Last, E.; Harder, S. Clopidogrel but not aspirin reduces P-selectin expression and formation of platelet-leukocyte aggregates in patients with atherosclerotic vascular disease. Clin. Pharmacol. Ther. 2003, 73, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Hu, H.; Hjemdahl, P. Aspirin treatment does not attenuate platelet or leukocyte activation as monitored by whole blood flow cytometry. Thromb. Res. 2003, 111, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Diehl, P.; Olivier, C.; Haischeid, C.; Helbing, T.; Bode, C.; Moser, M. Clopidogrel affects leukocyte dependent platelet aggregation by P2Y 12 expressing leukocytes. Basic Res. Cardiol. 2010, 105, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.R.; Marso, S.P.; Dai, D.; Anstrom, K.J.; Shunk, K.A.; Curtus, J.P.; Brennan, J.M.; Sedrakyan, A.; Messenger, J.C.; Douglas, P.S. Comparative Effectiveness of Drug-Eluting Versus Bare-Metal Stents in Elderly Patients Undergoing Revascularization of Chronic Total Coronary Occlusions: Results From the National Cardiovascular Data Registry, 2005–2008. JACC Cardiovasc. Interv. 2012, 5, 1054–1061. [Google Scholar] [CrossRef] [Green Version]
- Crimi, G.; Gritti, V.; Galiffa, V.A.; Scotti, V.; Leonardi, S.; Ferrario, M.; Ferlini, M.; De Ferrari, G.M.; Oltrona Visconti, L.; Klersy, C. Drug eluting stents are superior to bare metal stents to reduce clinical outcome and stent-related complications in CKD patients, a systematic review, meta-analysis and network meta-analysis. J. Interv. Cardiol. 2018, 31, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccolo, R.; Bonaa, K.H.; Efthimiou, O.; Varenne, O.; Baldo, A.; Urban, P.; Kaiser, C.; Remkes, W.; Räber, L.; de Belder, A.; et al. Drug-eluting or bare-metal stents for percutaneous coronary intervention: A systematic review and individual patient data meta-analysis of randomised clinical trials. Lancet 2019, 393, 2503–2510. [Google Scholar] [CrossRef]
- Pickard, A.S.; Becker, R.C.; Schumock, G.T.; Frye, C.B. Clopidogrel-Associated Bleeding and Related Complications in Patients Undergoing Coronary Artery Bypass Grafting. Pharmacotherapy 2008, 28, 376–392. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, W.; Zou, D.; Kou, F.; Hou, Y.; Yasin, A.; Zhang, K. Comparison of conjugating chondroitin sulfate A and B on amine-rich surface: For deeper understanding on directing cardiovascular cells fate. Compos. Part B Eng. 2022, 228, 109430. [Google Scholar] [CrossRef]
- Han, Z.; Guo, H.; Zhou, Y.; Wang, L.; Zhang, K.; Li, J.A. Composite Coating Prepared with Ferulic Acid to Improve the Corrosion Resistance and Blood Compatibility of Magnesium Alloy. Metals 2022, 12, 545. [Google Scholar] [CrossRef]
- Li, J.; Zhang, K.; Hou, Y. From selective cardiovascular cells adhesion to regulating spatiotemporal orderliness of function: Understanding based on biomaterials surface modification with functional molecules. Curr. Top. Med. Med. Res. 2020, 5, 155–161. [Google Scholar]
- Haemmerle, M.; Stone, R.L.; Menter, D.G.; Afshar-Kharghan, V.; Sood, A.K. The Platelet Lifeline to Cancer: Challenges and Opportunities. Cancer Cell 2018, 33, 965–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios-Acedo, A.L.; Mège, D.; Crescence, L.; Dignat-George, F.; Dubois, C.; Panicot-Dubois, L. Platelets, Thrombo-Inflammation, and Cancer: Collaborating With the Enemy. Front. Immunol. 2019, 10, 1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.R.; Yousef, G.M.; Ni, H. Cancer and platelet crosstalk: Opportunities and challenges for aspirin and other antiplatelet agents. Blood 2018, 131, 1777–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, M. Role of platelets and platelet receptors in cancer metastasis. J. Hematol. Oncol. 2018, 11, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Cappellano, G.; Raineri, D.; Rolla, R.; Giordano, M.; Puricelli, C.; Vilardo, B.; Manfredi, M.; Cantaluppi, V.; Sainaghi, P.P.; Castello, L.; et al. Circulating Platelet-Derived Extracellular Vesicles Are a Hallmark of SARS-CoV-2 Infection. Cells 2021, 10, 85. [Google Scholar] [CrossRef]
- Olsson, A.K.; Cedervall, J. The pro-inflammatory role of platelets in cancer. Platelets 2018, 29, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Patrignani, P.; Patrono, C. Aspirin, platelet inhibition and cancer prevention. Platelets 2018, 29, 779–785. [Google Scholar] [CrossRef]
- Mezouar, S.; Darbousset, R.; Dignat-George, F.; Panicot-Dubois, L.; Dubois, C. Inhibition of platelet activation prevents the P-selectin and integrin-dependent accumulation of cancer cell microparticles and reduces tumor growth and metastasis in vivo. Int. J. Cancer 2015, 136, 462–475. [Google Scholar] [CrossRef] [Green Version]
- Bibbins-Domingo, K.; Grossman, D.C.; Curry, S.J.; Davidson, K.W.; Epling, J.W.; García, F.A.R.; Gillman, M.; Harper, D.M.; Kemper, A.R.; Krist, A.H.; et al. Aspirin Use for the Primary Prevention of Cardiovascular Disease and Colorectal Cancer: U.S. Preventive Services Task Force Recommendation Statement. Ann. Intern. Med. 2016, 164, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Kolandaivelu, K.; Bhatt, D. Novel antiplatelet therapies. In Platelets; Michelson, A.D., Ed.; Academic Press: Amsterdam, The Netherlands, 2013; pp. 1185–1213. ISBN 9780123878373/0123878373. [Google Scholar]
- Jain, S.; Russell, S.; Ware, J. Platelet glycoprotein VI facilitates experimental lung metastasis in syngenic mouse models. J. Thromb. Haemost. 2009, 7, 1713–1717. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Ma, L.; Carrim, N.; Yougbare, I.; Li, J.; Chen, P.; Zhu, G.; Ni, H. Platelet GPIba Is Important for Thrombopoietin Production and Thrombopoietin-Induced Platelet Generation. Blood 2015, 126, 12. [Google Scholar] [CrossRef]
- Leiter, O.; Walker, T.L. Platelets: The missing link between the blood and brain? Prog. Neurobiol. 2019, 183, 101695. [Google Scholar] [CrossRef] [PubMed]
- Gaertner, F.; Ahmad, Z.; Rosenberger, G.; Fan, S.; Nicolai, L.; Busch, B.; Yavuz, G.; Luckner, M.; Ishikawa-Ankerhold, H.; Hennel, R.; et al. Migrating Platelets Are Mechano-scavengers that Collect and Bundle Bacteria. Cell 2017, 171, 1368–1382.e23. [Google Scholar] [CrossRef] [PubMed]
- Hayon, Y.; Dashevsky, O.; Shai, E.; Varon, D.; Leker, R.R. Platelet lysates stimulate angiogenesis, neurogenesis and neuroprotection after stroke. Thromb. Haemost. 2013, 110, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotnikov, I.; Veremeyko, T.; Starossom, S.C.; Barteneva, N.; Weiner, H.L.; Ponomarev, E.D. Platelets recognize brain-specific glycolipid structures, respond to neurovascular damage and promote neuroinflammation. PLoS ONE 2013, 8, e58979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukhinova, M.; Kuznetsova, I.; Kopeikina, E.; Veniaminova, E.; Yung, A.W.Y.; Veremeyko, T.; Levchuk, K.; Barteneva, N.S.; Wing-Ho, K.K.; Yung, W.H.; et al. Platelets mediate protective neuroinflammation and promote neuronal plasticity at the site of neuronal injury. Brain Behav. Immun. 2018, 74, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Wasielewska, J.M.; Grönnert, L.; Rund, N.; Donix, L.; Rust, R.; Sykes, A.M.; Hoppe, A.; Roers, A.; Kempermann, G.; Walker, T.L. Mast cells increase adult neural precursor proliferation and differentiation but this potential is not realized in vivo under physiological conditions. Sci. Rep. 2017, 7, 17759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjayani, S.; Wirohadidjojo, Y.W.; Adam, A.M.; Suwandi, D.; Seweng, A.; Amiruddin, M.D. Sensory improvement of leprosy peripheral neuropathy in patients treated with perineural injection of platelet-rich plasma. Int. J. Dermatol. 2014, 53, 109–113. [Google Scholar] [CrossRef]
- Chen, N.F.; Sung, C.S.; Wen, Z.H.; Chen, C.H.; Feng, C.W.; Hung, H.C.; Yang, S.N.; Tsui, K.H.; Chen, W.F. Therapeutic effect of platelet-rich plasma in rat spinal cord injuries. Front. Neurosci. 2018, 12, 252. [Google Scholar] [CrossRef]
- Borhani-Haghighi, M.; Mohamadi, Y. The therapeutic effect of platelet-rich plasma on the experimental autoimmune encephalomyelitis mice. J. Neuroimmunol. 2019, 333, 476958. [Google Scholar] [CrossRef] [PubMed]
- Hayon, Y.; Dashevsky, O.; Shai, E.; Varon, D.; Leker, R.R. Platelet microparticles promote neural stem cell proliferation, survival and differentiation. J. Mol. Neurosci. 2012, 47, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Gouel, F.; Do Van, B.; Chou, M.L.; Jonneaux, A.; Moreau, C.; Bordet, R.; Burnouf, T.; Devedjian, J.C.; Devos, D. The protective effect of human platelet lysate in models of neurodegenerative disease: Involvement of the Akt and MEK pathways. J. Tissue Eng. Regen. Med. 2017, 11, 3236–3240. [Google Scholar] [CrossRef] [PubMed]
- Abubaker, A.A.; Vara, D.; Visconte, C.; Eggleston, I.; Torti, M.; Canobbio, I.; Pula, G. Amyloid Peptide β 1-42 Induces Integrin α IIb β 3 Activation, Platelet Adhesion, and Thrombus Formation in a NADPH Oxidase-Dependent Manner. Oxid. Med. Cell. Longev. 2019, 2019, 1050476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiter, O.; Walker, T.L. Platelets in Neurodegenerative Conditions—Friend or Foe? Front. Immunol. 2020, 11, 760. [Google Scholar] [CrossRef]
- Inyushin, M.Y.; Sanabria, P.; Rojas, L.; Kucheryavykh, Y.; Kucheryavykh, L. A β Peptide Originated from Platelets Promises New Strategy in Anti-Alzheimer’s Drug Development. BioMed Res. Int. 2017, 2017, 3948360. [Google Scholar] [CrossRef] [Green Version]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, É.; Pap, E.; Kittel, Á.; et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [Green Version]
- van der Pol, E.; Böing, A.N.; Gool, E.L.; Nieuwland, R. Recent developments in the nomenclature, presence, isolation, detection and clinical impact of extracellular vesicles. J. Thromb. Haemost. 2016, 14, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating Extracellular Vesicles in Human Disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef]
- Au Yeung, C.L.; Co, N.N.; Tsuruga, T.; Yeung, T.L.; Kwan, S.Y.; Leung, C.S.; Li, Y.; Lu, E.S.; Kwan, K.; Wong, K.K.; et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun. 2016, 7, 11150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, S.A.; Sugimoto, H.; O’Connell, J.T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vitkin, E.; Perelman, L.T.; Melo, C.A.; et al. Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Signal Transduct. Target. Ther. 2020, 5, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat. Cell Biol. 2015, 17, 816–826. [Google Scholar] [CrossRef]
- Ji, Q.; Zhou, L.; Sui, H.; Yang, L.; Wu, X.; Song, Q.; Jia, R.; Li, R.; Sun, J.; Wang, Z.; et al. Primary tumors release ITGBL1-rich extracellular vesicles to promote distal metastatic tumor growth through fibroblast-niche formation. Nat. Commun. 2020, 11, 1211–1228. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.; Garcia, V.; Rodriguez, M.; Compte, M.; Cisneros, E.; Veguillas, P.; Garcia, J.M.; Dominguez, G.; Campos-Martin, Y.; Cuevas, J.; et al. Analysis of exosome release and its prognostic value in human colorectal cancer. Genes. Chromosom. Cancer 2012, 51, 409–418. [Google Scholar] [CrossRef]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar] [CrossRef]
- Kim, M.S.; Haney, M.J.; Zhao, Y.; Mahajan, V.; Deygen, I.; Klyachko, N.L.; Inskoe, E.; Piroyan, A.; Sokolsky, M.; Okolie, O.; et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine 2016, 12, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.J.; Sun, X.Y.; Huang, K.M.; Zhang, L.; Yang, Z.S.; Zou, D.D.; Wang, B.; Warnock, G.L.; Dai, L.J.; Luo, J. Therapeutic potential of CAR-T cell-derived exosomes: A cell-free modality for targeted cancer therapy. Oncotarget 2015, 6, 44179–44190. [Google Scholar] [CrossRef] [Green Version]
- Emanueli, C.; Shearn, A.I.U.; Laftah, A.; Fiorentino, F.; Reeves, B.C.; Beltrami, C.; Mumford, A.; Clayton, A.; Gurney, M.; Shantikumar, S.; et al. Coronary Artery-Bypass-Graft Surgery Increases the Plasma Concentration of Exosomes Carrying a Cargo of Cardiac MicroRNAs: An Example of Exosome Trafficking Out of the Human Heart with Potential for Cardiac Biomarker Discovery. PLoS ONE 2016, 11, e0154274. [Google Scholar] [CrossRef] [Green Version]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, F.; Wang, H.; Przybilla, D.; Franklin, B.S.; Dolf, A.; Pfeifer, P.; Schmitz, T.; Flender, A.; Endl, E.; Nickenig, G.; et al. Vascular endothelial microparticles-incorporated microRNAs are altered in patients with diabetes mellitus. Cardiovasc. Diabetol. 2016, 15, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, Y.X.; Liu, H.; Zhang, L.S.; Lv, W.; Hu, X.Y. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6, 37043–37053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetzl, E.J.; Abner, E.L.; Jicha, G.A.; Kapogiannis, D.; Schwartz, J.B. Declining levels of functionally specialized synaptic proteins in plasma neuronal exosomes with progression of Alzheimer’s disease. FASEB J. 2018, 32, 888–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, R.A.; Tripodis, Y.; Baugh, C.M.; Fritts, N.G.; Martin, B.M.; Chaisson, C.; Cantu, R.C.; Joyce, J.A.; Shah, S.; Ikezu, T.; et al. Preliminary Study of Plasma Exosomal Tau as a Potential Biomarker for Chronic Traumatic Encephalopathy. J. Alzheimer’s Dis. 2016, 51, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Gu, H.; Guo, Q.; Liu, X.; Li, F.; Liu, H.; Sun, L.; Ma, H.; Zhao, K. Circulating Small Extracellular Vesicle-Derived miR-342-5p Ameliorates Beta-Amyloid Formation via Targeting Beta-site APP Cleaving Enzyme 1 in Alzheimer’s Disease. Cells 2022, 11, 3830. [Google Scholar] [CrossRef]
- Garcia-Contreras, M.; Thakor, A. Extracellular vesicles in Alzheimer’s disease: From pathology to therapeutic approaches. Neural Regen. Res. 2023, 18, 22. [Google Scholar] [CrossRef]
- Apodaca, L.A.; Baddour, A.A.D.; Garcia, C.; Alikhani, L.; Giedzinski, E.; Ru, N.; Agrawal, A.; Acharya, M.M.; Baulch, J.E. Human neural stem cell-derived extracellular vesicles mitigate hallmarks of Alzheimer’s disease. Alzheimer’s Res. Ther. 2021, 13, 57. [Google Scholar] [CrossRef]
- Brisson, A.R.; Tan, S.; Linares, R.; Gounou, C.; Arraud, N. Extracellular vesicles from activated platelets: A semiquantitative cryo-electron microscopy and immuno-gold labeling study. Platelets 2017, 28, 263–271. [Google Scholar] [CrossRef]
- Puhm, F.; Boilard, E.; Machlus, K.R. Platelet extracellular vesicles: Beyond the blood. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 87–96. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Pachler, K.; Lener, T.; Streif, D.; Dunai, Z.A.; Desgeorges, A.; Feichtner, M.; Öller, M.; Schallmoser, K.; Rohde, E.; Gimona, M. A Good Manufacturing Practice-grade standard protocol for exclusively human mesenchymal stromal cell-derived extracellular vesicles. Cytotherapy 2017, 19, 458–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Wu, Y.W.; Blyth, C.; Lichtfuss, G.; Goubran, H.; Burnouf, T. Prospective Therapeutic Applications of Platelet Extracellular Vesicles. Trends Biotechnol. 2021, 39, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, V.; Burnouf, P.A.; Burnouf, T.; Agrahari, V. Nanoformulation properties, characterization, and behavior in complex biological matrices: Challenges and opportunities for brain-targeted drug delivery applications and enhanced translational potential. Adv. Drug Deliv. Rev. 2019, 148, 146–180. [Google Scholar] [CrossRef]
- Kola, S.M.; Kumar, P.; Choonara, Y.E.; du Toit, L.C.; Pillay, V. Hypothesis: Can drug-loaded platelets be used as delivery vehicles for blood-brain barrier penetration? Med. Hypotheses 2019, 125, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Burnouf, T.; Goubran, H.A.; Chou, M.L.; Devos, D.; Radosevic, M. Platelet microparticles: Detection and assessment of their paradoxical functional roles in disease and regenerative medicine. Blood Rev. 2014, 28, 155–166. [Google Scholar] [CrossRef]
- Boudreau, L.H.; Duchez, A.C.; Cloutier, N.; Soulet, D.; Martin, N.; Bollinger, J.; Paré, A.; Rousseau, M.; Naika, G.S.; Lévesque, T.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood 2014, 124, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Boilard, E.; Duchez, A.C.; Brisson, A. The diversity of platelet microparticles. Curr. Opin. Hematol. 2015, 22, 437–444. [Google Scholar] [CrossRef]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef]
- Antich-Rosselló, M.; Forteza-Genestra, M.A.; Calvo, J.; Gayà, A.; Monjo, M.; Ramis, J.M. Platelet-derived extracellular vesicles promote osteoinduction of mesenchymal stromal cells. Bone Jt. Res. 2020, 9, 667–674. [Google Scholar] [CrossRef]
- Otahal, A.; Kuten-Pella, O.; Kramer, K.; Neubauer, M.; Lacza, Z.; Nehrer, S.; De Luna, A. Functional repertoire of EV-associated miRNA profiles after lipoprotein depletion via ultracentrifugation and size exclusion chromatography from autologous blood products. Sci. Rep. 2021, 11, 5823. [Google Scholar] [CrossRef]
- Aatonen, M.T.; Öhman, T.; Nyman, T.A.; Laitinen, S.; Grönholm, M.; Siljander, P.R.M. Isolation and characterization of platelet-derived extracellular vesicles. J. Extracell. Vesicles 2014, 3, 24692. [Google Scholar] [CrossRef] [PubMed]
- Milioli, M.; Ibáñez-Vea, M.; Sidoli, S.; Palmisano, G.; Careri, M.; Larsen, M.R. Quantitative proteomics analysis of platelet-derived microparticles reveals distinct protein signatures when stimulated by different physiological agonists. J. Proteom. 2015, 121, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Shai, E.; Rosa, I.; Parguiña, A.F.; Motahedeh, S.; Varon, D.; García, Á. Comparative analysis of platelet-derived microparticles reveals differences in their amount and proteome depending on the platelet stimulus. J. Proteom. 2012, 76, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Dean, W.L.; Lee, M.J.; Cummins, T.D.; Schultz, D.J.; Powell, D.W.; Dean, B. Proteomic and functional characterisation of platelet microparticle size classes. Thromb. Haemost. 2009, 102, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Richter, M.; de Rossi, C.; Lehr, C.M.; Fuhrmann, K.; Fuhrmann, G. Extracellular vesicles protect glucuronidase model enzymes during freeze-drying. Sci. Rep. 2018, 8, 12377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.W.; Huang, C.C.; Changou, C.A.; Lu, L.S.; Goubran, H.; Burnouf, T. Clinical-grade cryopreserved doxorubicin-loaded platelets: Role of cancer cells and platelet extracellular vesicles activation loop. J. Biomed. Sci. 2020, 27, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Lopez, E.; Srivastava, A.K.; Burchfield, J.; Wang, Y.W.; Cardenas, J.C.; Togarrati, P.P.; Miyazawa, B.; Gonzalez, E.; Holcomb, J.B.; Pati, S.; et al. Platelet-derived- Extracellular Vesicles Promote Hemostasis and Prevent the Development of Hemorrhagic Shock. Sci. Rep. 2019, 9, 17676. [Google Scholar] [CrossRef] [Green Version]
- Qin, B.; Zhang, Q.; Hu, X.M.; Mi, T.Y.; Yu, H.Y.; Liu, S.S.; Zhang, B.; Tang, M.; Huang, J.F.; Xiong, K. How does temperature play a role in the storage of extracellular vesicles? J. Cell. Physiol. 2020, 235, 7663–7680. [Google Scholar] [CrossRef]
- Miyazawa, B.; Trivedi, A.; Togarrati, P.P.; Potter, D.; Baimukanova, G.; Vivona, L.; Lin, M.; Lopez, E.; Callcut, R.; Srivastava, A.K.; et al. Regulation of endothelial cell permeability by platelet-derived extracellular vesicles. J. Trauma Acute Care Surg. 2019, 86, 942. [Google Scholar] [CrossRef] [PubMed]
- Sinauridze, E.I.; Kireev, D.A.; Popenko, N.Y.; Pichugin, A.V.; Panteleev, M.A.; Krymskaya, O.V.; Ataullakhanov, F.I. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb. Haemost. 2007, 97, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, H.; Song, J.; Choi, E.S.; You, G.; Mok, H. Activated Platelet-Derived Vesicles for Efficient Hemostatic Activity. Macromol. Biosci. 2020, 20, e1900338. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.C.; Tao, S.C.; Yin, W.J.; Qi, X.; Yuan, T.; Zhang, C.Q. Exosomes derived from platelet-rich plasma promote the re-epithelization of chronic cutaneous wounds via activation of YAP in a diabetic rat model. Theranostics 2017, 7, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, F.; Carton, F.; Gino, S.; Migliario, M.; Renò, F. Platelet rich plasma-derived microvesicles increased in vitro wound healing. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9658–9664. [Google Scholar] [CrossRef]
- Iyer, S.R.; Scheiber, A.L.; Yarowsky, P.; Henn, R.F.; Otsuru, S.; Lovering, R.M. Exosomes Isolated From Platelet-Rich Plasma and Mesenchymal Stem Cells Promote Recovery of Function After Muscle Injury. Am. J. Sport. Med. 2020, 48, 2277–2286. [Google Scholar] [CrossRef]
- Liang, C.; Huang, J.; Luo, P.; Wang, Z.; He, J.; Wu, S.; Peng, C.; Cao, X. Platelet-Derived Microparticles Mediate the Intra-Articular Homing of Mesenchymal Stem Cells in Early-Stage Cartilage Lesions. Stem Cells Dev. 2020, 29, 414–424. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Ma, C.; Wang, G.; Zhang, Y.; Sun, S. Exosomes derived from platelet-rich plasma present a novel potential in alleviating knee osteoarthritis by promoting proliferation and inhibiting apoptosis of chondrocyte via Wnt/β-catenin signaling pathway. J. Orthop. Surg. Res. 2019, 14, 470–485. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.C.; Yuan, T.; Rui, B.Y.; Zhu, Z.Z.; Guo, S.C.; Zhang, C.Q. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics 2017, 7, 733–750. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Zambuzzi, W.F. Platelet microparticles load a repertory of miRNAs programmed to drive osteogenic phenotype. J. Biomed. Mater. Res. A 2021, 109, 1502–1511. [Google Scholar] [CrossRef]
- Hayon, Y.; Dashevsky, O.; Shai, E.; Brill, A.; Varon, D.; R. Leker, R. Platelet microparticles induce angiogenesis and neurogenesis after cerebral ischemia. Curr. Neurovasc. Res. 2012, 9, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Brill, A.; Dashevsky, O.; Rivo, J.; Gozal, Y.; Varon, D. Platelet-derived microparticles induce angiogenesis and stimulate post-ischemic revascularization. Cardiovasc. Res. 2005, 67, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Mause, S.F.; Ritzel, E.; Liehn, E.A.; Hristov, M.; Bidzhekov, K.; Müller-Newen, G.; Soehnlein, O.; Weber, C. Platelet microparticles enhance the vasoregenerative potential of angiogenic early outgrowth cells after vascular injury. Circulation 2010, 122, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaldivia, M.T.K.; McFadyen, J.D.; Lim, B.; Wang, X.; Peter, K. Platelet-Derived Microvesicles in Cardiovascular Diseases. Front. Cardiovasc. Med. 2017, 4, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Goubran, H.; Sabry, W.; Kotb, R.; Seghatchian, J.; Burnouf, T. Platelet microparticles and cancer: An intimate cross-talk. Transfus. Apher. Sci. 2015, 53, 168–172. [Google Scholar] [CrossRef]
- Franco, A.T.; Ware, J. Pathophysiology 2: The Role of Platelets in Cancer Biology. Cancer Treat. Res. 2019, 179, 37–54. [Google Scholar] [CrossRef]
- Michael, J.V.; Wurtzel, J.G.T.; Mao, G.F.; Rao, A.K.; Kolpakov, M.A.; Sabri, A.; Hoffman, N.E.; Rajan, S.; Tomar, D.; Madesh, M.; et al. Platelet microparticles infiltrating solid tumors transfer miRNAs that suppress tumor growth. Blood 2017, 130, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ai, Y.; Wang, L.; Bu, P.; Sharkey, C.C.; Wu, Q.; Wun, B.; Roy, S.; Shen, X.; King, M.R. Targeted drug delivery to circulating tumor cells via platelet membrane-functionalized particles. Biomaterials 2016, 76, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Sun, W.; Qian, C.; Wang, C.; Bomba, H.N.; Gu, Z. Anticancer platelet-mimicking nanovehicles. Adv. Mater. 2015, 27, 7050. [Google Scholar] [CrossRef]
- Soleymani, S.; Yari, F.; Bolhassani, A.; Bakhshandeh, H. Platelet microparticles: An effective delivery system for anti-viral drugs. J. Drug Deliv. Sci. Technol. 2019, 51, 290–296. [Google Scholar] [CrossRef]
- Jung, C.; Sörensson, P.; Saleh, N.; Arheden, H.; Rydén, L.; Pernow, J. Circulating endothelial and platelet derived microparticles reflect the size of myocardium at risk in patients with ST-elevation myocardial infarction. Atherosclerosis 2012, 221, 226–231. [Google Scholar] [CrossRef]
- Mause, S.F.; Von Hundelshausen, P.; Zernecke, A.; Koenen, R.R.; Weber, C. Platelet microparticles: A transcellular delivery system for RANTES promoting monocyte recruitment on endothelium. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1512–1518. [Google Scholar] [CrossRef] [Green Version]
- Böing, A.N.; Hau, C.M.; Sturk, A.; Nieuwland, R. Platelet microparticles contain active caspase 3. Platelets 2008, 19, 96–103. [Google Scholar] [CrossRef]
- Leroyer, A.S.; Isobe, H.; Lesèche, G.; Castier, Y.; Wassef, M.; Mallat, Z.; Binder, B.R.; Tedgui, A.; Boulanger, C.M. Cellular origins and thrombogenic activity of microparticles isolated from human atherosclerotic plaques. J. Am. Coll. Cardiol. 2007, 49, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Pawlowski, C.L.; Li, W.; Sun, M.; Ravichandran, K.; Hickman, D.S.; Kos, C.; Kaur, G.; Sen Gupta, A. Platelet microparticle-inspired clot-responsive nanomedicine for targeted fibrinolysis. Biomaterials 2017, 128, 108. [Google Scholar] [CrossRef]
- Shan, L.Y.; Li, J.Z.; Zu, L.Y.; Niu, C.G.; Ferro, A.; Zhang, Y.D.; Zheng, L.M.; Ji, Y. Platelet-derived microparticles are implicated in remote ischemia conditioning in a rat model of cerebral infarction. CNS Neurosci. Ther. 2013, 19, 917–925. [Google Scholar] [CrossRef]
- Ma, F.; Liu, H.; Shen, Y.; Zhang, Y.; Pan, S. Platelet-derived microvesicles are involved in cardio-protective effects of remote preconditioning. Int. J. Clin. Exp. Pathol. 2015, 8, 10839. [Google Scholar]
- Dai, Z.; Zhao, T.; Song, N.; Pan, K.; Yang, Y.; Zhu, X.; Chen, P.; Zhang, J.; Xia, C. Platelets and platelet extracellular vesicles in drug delivery therapy: A review of the current status and future prospects. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Sarkar, S.; Alam, M.A.; Shaw, J.; Dasgupta, A.K. Drug delivery using platelet cancer cell interaction. Pharm. Res. 2013, 30, 2785–2794. [Google Scholar] [CrossRef]
- Xu, P.; Zuo, H.; Chen, B.; Wang, R.; Ahmed, A.; Hu, Y.; Ouyang, J. Doxorubicin-loaded platelets as a smart drug delivery system: An improved therapy for lymphoma. Sci. Rep. 2017, 7, 42632. [Google Scholar] [CrossRef] [Green Version]
- Yarmush, M.L.; Golberg, A.; Serša, G.; Kotnik, T.; Miklavčič, D. Electroporation-Based Technologies for Medicine: Principles, Applications, and Challenges. Annu. Rev. Biomed. Eng. 2014, 16, 295–320. [Google Scholar] [CrossRef] [Green Version]
- Rao, L.; Bu, L.-L.; Ma, L.; Wang, W.; Liu, H.; Wan, D.; Liu, J.-F.; Li, A.; Guo, S.-S.; Zhang, L.; et al. Platelet-Facilitated Photothermal Therapy of Head and Neck Squamous Cell Carcinoma. Angew. Chem. 2018, 130, 998–1003. [Google Scholar] [CrossRef]
- Modery-Pawlowski, C.L.; Tian, L.L.; Pan, V.; McCrae, K.R.; Mitragotri, S.; Sen Gupta, A. Approaches to synthetic platelet analogs. Biomaterials 2013, 34, 526–541. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Lynn Modery-Pawlowski, C.; Menegatti, S.; Kumar, S.; Vogus, D.R.; Tian, L.L.; Chen, M.; Squires, T.M.; Gupta, A.S.; Mitragotri, S. Platelet-like Nanoparticles: Mimicking Shape, Flexibility, and Surface Biology of Platelets To Target Vascular Injuries. ACS Nano 2014, 8, 11253. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Factor | Source | Function | Refs. |
|---|---|---|---|
| BDNF | Platelets, neurons, heart, lung, liver, skeletal muscle | MSC survival and proliferation, angiogenesis, neuron activity, survival, and function | [40,41,42] |
| b-FGF | Platelets, macrophages, T lymphocytes, mast cells, endothelial cells, fibroblasts, chondrocytes, osteoblasts, mesenchymal cells | Mitogenic effect on fibroblasts, endothelial cells, MSCs, chondroblasts, and osteoblasts, angiogenesis, bone regeneration, corneal tissue repair, hair growth, wound healing, soft dental tissue regeneration, embryogenesis | [34,35,37,39,40,43] |
| BMP | Platelets, bone, CNS and many other tissues | Immune system regulation, cell maturation and differentiation, angiogenesis, cartilage and bone formation, fracture repair, tooth development, tissue fibrosis, cancer cell inhibition, CNS function | [37,40] |
| CTGF | Platelets after endocytosis from bone marrow extracellular environment | Found in platelets at a concentration that is 20-fold higher than any other PGF, involved in platelet adhesion, angiogenesis, cartilage regeneration, fibrosis, platelet adhesion, white blood cell migration, angiogenesis, regulation of collagen synthesis | [34,35,37,38,39] |
| EGF | Platelets, macrophages, monocytes | Endothelial chemotaxis, epithelial and mesenchymal mitogenesis, regulation of collagenase secretion, keratinocyte and fibroblast migration and proliferation, angiogenesis, re-epithelialization, corneal tissue repair | [34,35,37,39,40,43] |
| IGF-1 | Platelets, epithelial cells, endothelial cells, fibroblasts, smooth muscle cells, osteoblasts | Differentiation of osteoblasts in bone and of myeloblastic tissue in muscle, proliferation, migration, angiogenesis, neuroprotection and re-myelination, bone regeneration, regulation of collagen production, synergistic effect with PDGF | [39,40,43] |
| PDGF | Platelets, endothelial cells, macrophages, monocytes, smooth muscle cells, osteoblasts, keratinocytes | First growth factor to be released in a wound, mitogenesis, chemotaxis of macrophages and neutrophils, regulation of collagen synthesis and collagenase activity, matrix formation and remodeling, angiogenesis, neurogenesis, bone regeneration, re-epithelialization, wound healing, corneal tissue repair, synergistic effect with TGF-β | [34,35,37,39,40,43] |
| TGF (α-β) | Platelets, activated Th1 cells, NK cells, monocytes, macrophages, endothelial cells, neutrophils, keratinocytes, fibroblasts, muscle cells, bone and cartilage ECM | MSC proliferation, mitogenesis of fibroblasts, osteoblasts, and endothelial cells, chemotaxis of endothelial cells, angiogenesis, regulation of collagen synthesis and collagenase secretion, extracellular matrix formation and connective tissue regeneration, regulation of mitogenesis mediated by other growth factors, bone formation and regeneration, re-epithelialization, wound healing, cancer metastasis, inhibition of macrophage and lymphocyte proliferation, synergistic effect with PDGF, possible anti-proliferative effect at high concentrations | [34,35,37,39,40,43] |
| VEGF | Platelets, endothelial cells, fibroblasts, keratinocytes | Differentiation and mitogenesis of endothelial cells, chemotaxis, angiogenesis, increased vascular permeability, vascularization, neuroprotection, bone regeneration | [34,35,37,39,40,43] |
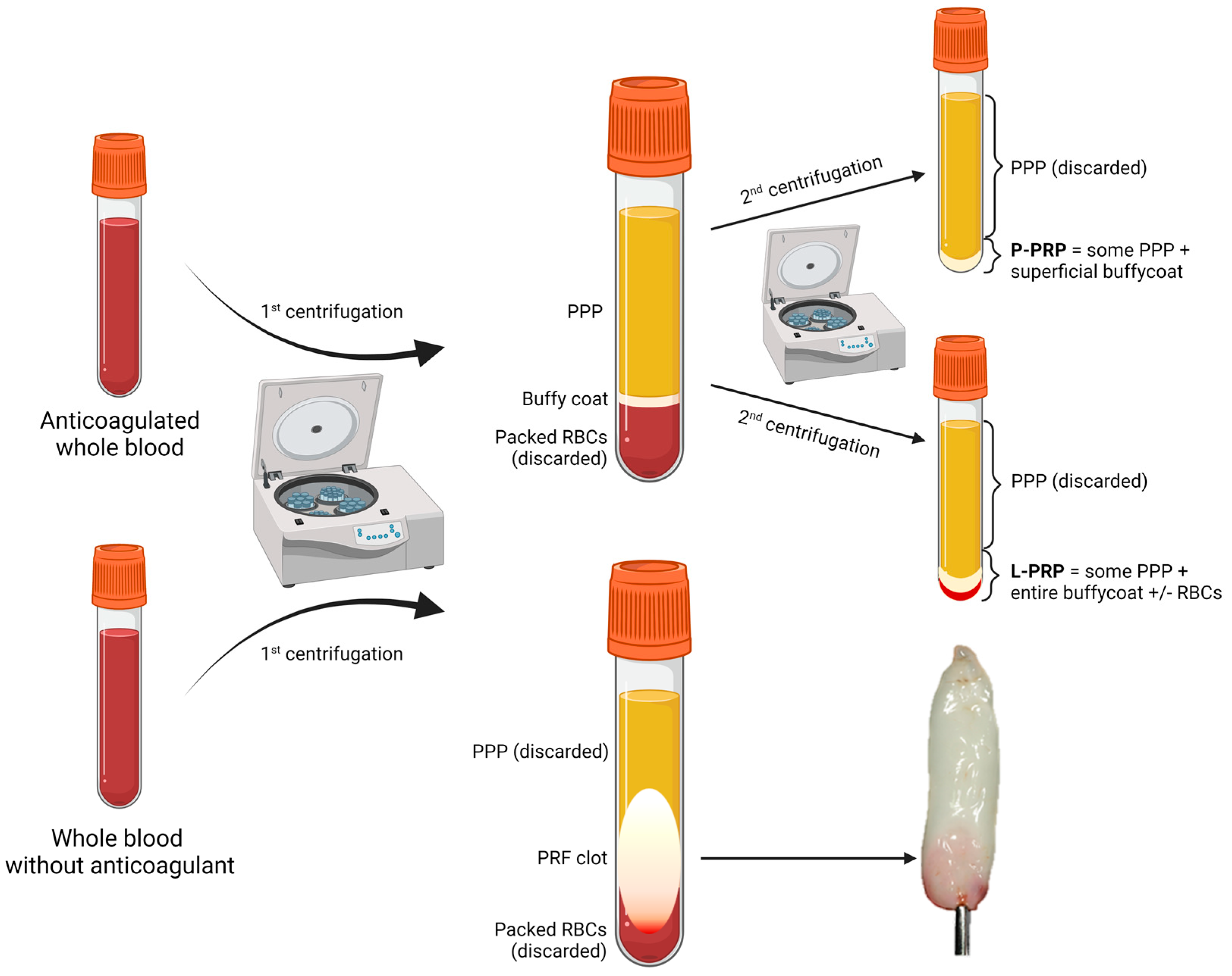
| Product | Source | Preparation Steps | Characteristics | Refs. |
|---|---|---|---|---|
| P-PRP L-PRP | ACD-anticoagulated whole blood or apheresis | 1. Short soft-spin centrifugation to separate PPP, BC, and packed RBCs; 2. Collection of PPP and BC; 3. Long hard-spin centrifugation and discarding of the supernatant (PPP) to obtain P-PRP or L-PRP. |
| [35,48,49] |
| PRF | ACD-anticoagulated whole blood or apheresis | Immediate centrifugation to obtain a flexible fibrin clot. |
| [37,48,50,51,52,53,54,55] |
| PG | ACD-anticoagulated whole blood or apheresis | Same preparation as P-PRP or L-PRP but additional activation step to form a semi-solid (gelated) product by the action of calcium chloride or calcium gluconate with or without the addition of human thrombin, batroxobin, or synthetic TRAP (a thrombin receptor agonist), often added directly at the application site. |
| [35,37,56,57,58,59] |
| PL | Whole blood or apheresis platelets | Platelet degranulation after freeze-thaw cycles, sonication, treatment with solvents and detergents, possibly followed by activation by thrombin or batroxobin just before application. |
| [40] |
| E-S | Whole blood | Clotting at room temperature for 2–72 h, centrifugation to obtain serum and dilution with saline solution. |
| [37,60,61] |
| E-PRP | Whole blood or apheresis platelets | Platelet degranulation after freeze-thaw cycles, dilution of the supernatant with saline solution. | Higher content of PGFs in a smaller volume, more suitable for intraocular instillation. | [37,56,62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puricelli, C.; Boggio, E.; Gigliotti, C.L.; Stoppa, I.; Sutti, S.; Giordano, M.; Dianzani, U.; Rolla, R. Platelets, Protean Cells with All-Around Functions and Multifaceted Pharmacological Applications. Int. J. Mol. Sci. 2023, 24, 4565. https://doi.org/10.3390/ijms24054565
Puricelli C, Boggio E, Gigliotti CL, Stoppa I, Sutti S, Giordano M, Dianzani U, Rolla R. Platelets, Protean Cells with All-Around Functions and Multifaceted Pharmacological Applications. International Journal of Molecular Sciences. 2023; 24(5):4565. https://doi.org/10.3390/ijms24054565
Chicago/Turabian StylePuricelli, Chiara, Elena Boggio, Casimiro Luca Gigliotti, Ian Stoppa, Salvatore Sutti, Mara Giordano, Umberto Dianzani, and Roberta Rolla. 2023. "Platelets, Protean Cells with All-Around Functions and Multifaceted Pharmacological Applications" International Journal of Molecular Sciences 24, no. 5: 4565. https://doi.org/10.3390/ijms24054565