
Therapeutic Hypothermia Attenuates Cortical Interneuron Loss after Cerebral Ischemia in Near-Term Fetal Sheep
 , , and
, , and
Abstract
:1. Introduction
2. Results
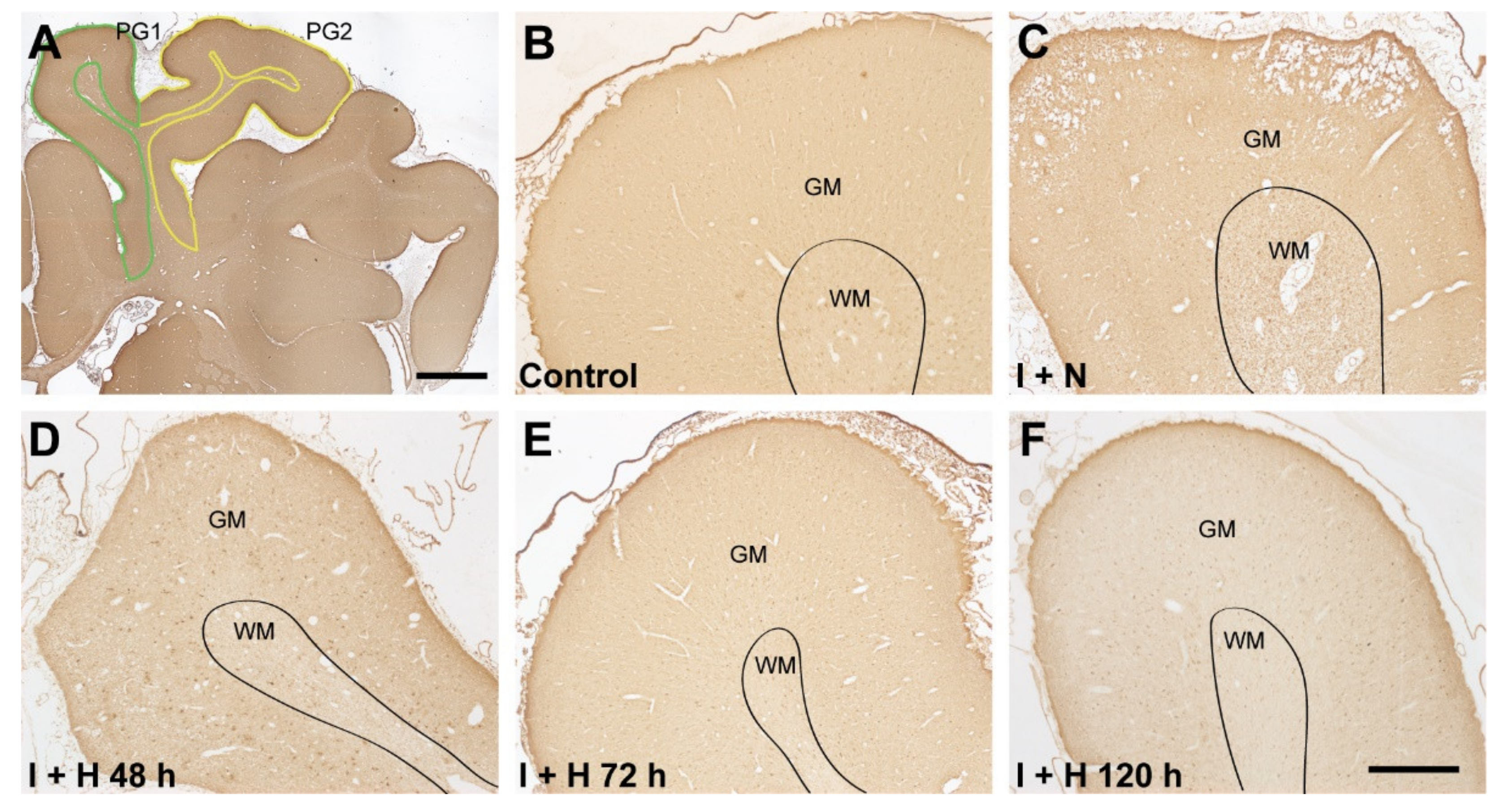
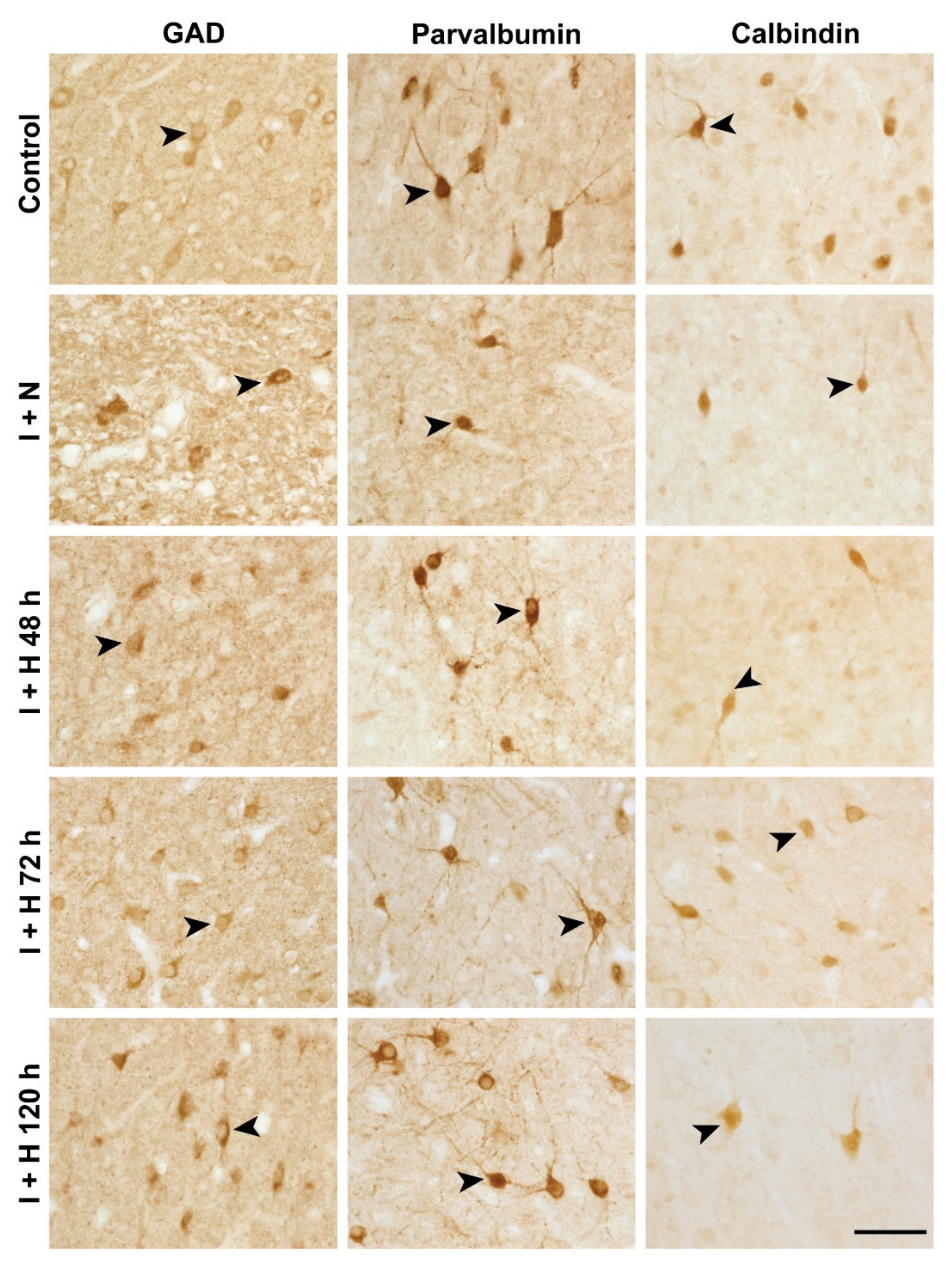
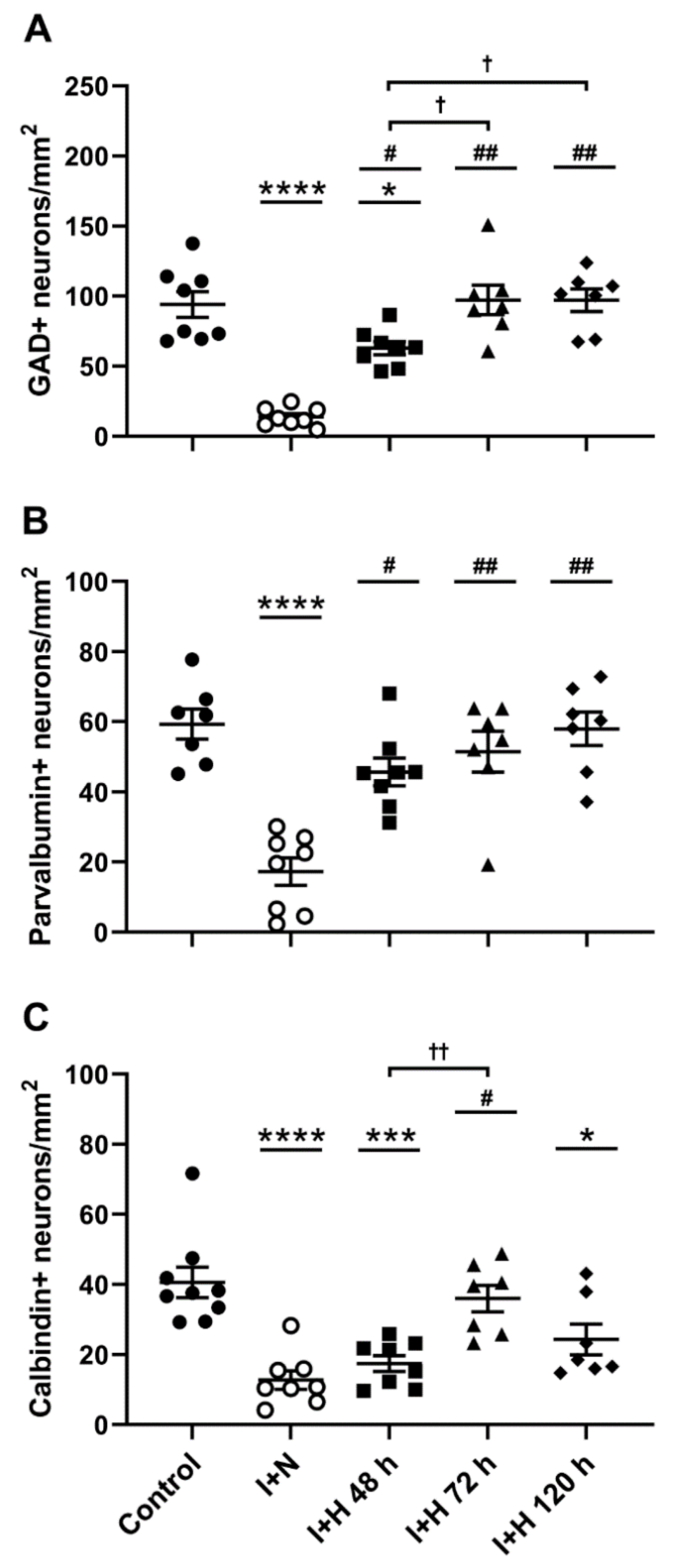
2.1. Survival of GABAergic Interneurons in the Parasagittal Cortex after Cerebral Ischemia and Therapeutic Hypothermia
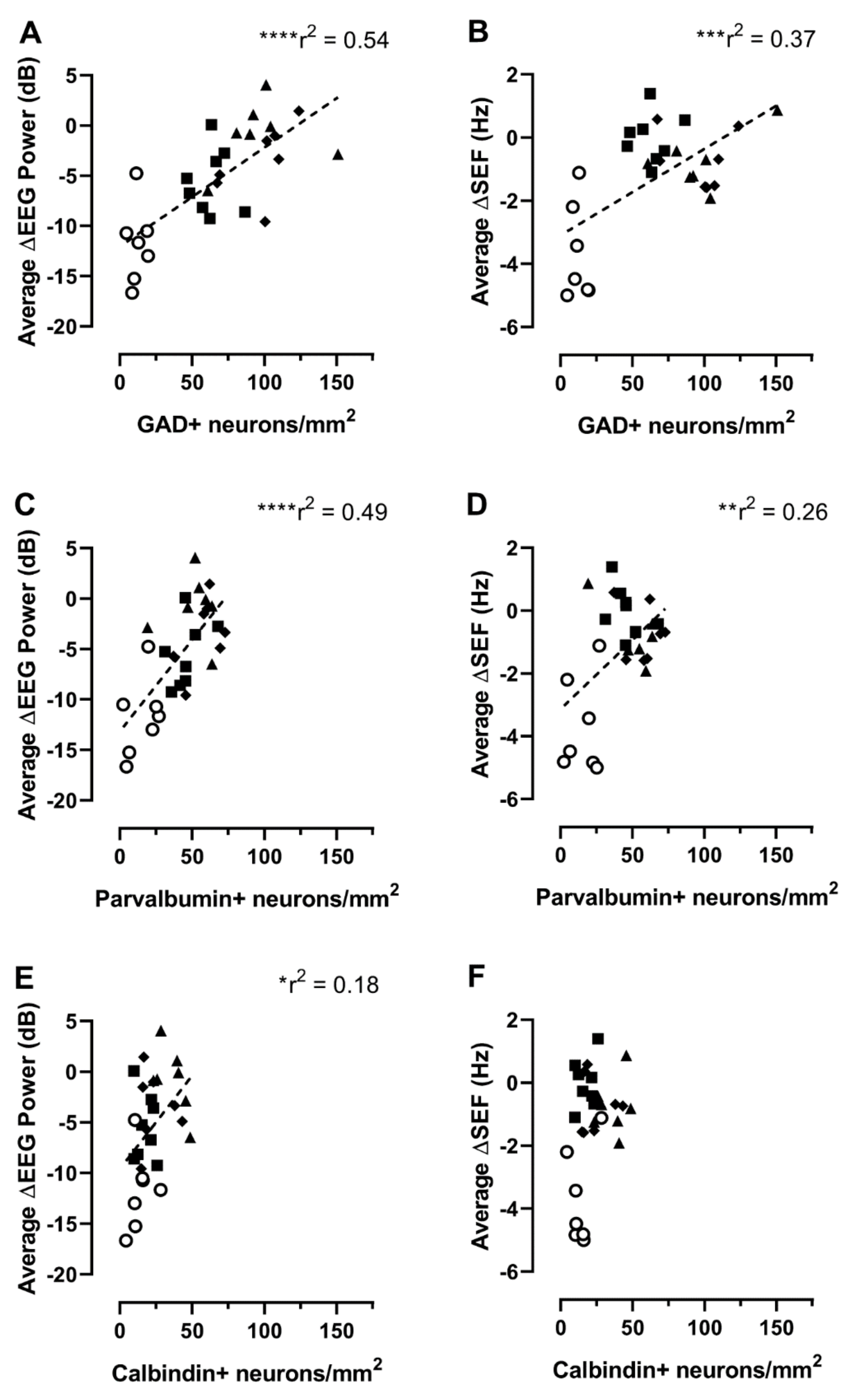
2.2. Relationship between Interneuron Survival and EEG or Spectral Edge Frequency
3. Discussion
4. Materials and Methods
4.1. Animals and Surgery
4.2. Postoperative Care
4.3. Experimental Protocols
4.4. Electrophysiological Data Recording and Analysis
4.5. Tissue Preparation
4.6. Immunohistochemical Staining
4.7. Image Analysis
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wassink, G.; Davidson, J.O.; Lear, C.A.; Juul, S.E.; Northington, F.; Bennet, L.; Gunn, A.J. A working model for hypothermic neuroprotection. J. Physiol. 2018, 596, 5641–5654. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.; Statnikov, Y.; Jawad, S.; Uthaya, S.N.; Modi, N. Neonatal brain injuries in England: Population-based incidence derived from routinely recorded clinical data held in the National Neonatal Research Database. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F301–F306. [Google Scholar] [CrossRef] [PubMed]
- Glass, H.C.; Shellhaas, R.A.; Wusthoff, C.J.; Chang, T.; Abend, N.S.; Chu, C.J.; Cilio, M.R.; Glidden, D.V.; Bonifacio, S.L.; Massey, S.; et al. Contemporary Profile of Seizures in Neonates: A Prospective Cohort Study. J. Pediatr. 2016, 174, 98–103.e1. [Google Scholar] [CrossRef] [PubMed]
- Osmond, E.; Billetop, A.; Jary, S.; Likeman, M.; Thoresen, M.; Luyt, K. Neonatal seizures: Magnetic resonance imaging adds value in the diagnosis and prediction of neurodisability. Acta Paediatr. 2014, 103, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Barkovich, A.J.; Truwit, C.L. Brain damage from perinatal asphyxia: Correlation of MR findings with gestational age. Am. J. Neuroradiol. 1990, 11, 1087–1096. [Google Scholar]
- Bano, S.; Chaudhary, V.; Garga, U.C. Neonatal hypoxic-ischemic encephalopathy: A radiological review. J. Pediatr. Neurosci. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- Rocha-Ferreira, E.; Hristova, M. Plasticity in the Neonatal Brain following Hypoxic-Ischaemic Injury. Neural Plast. 2016, 2016, 4901014. [Google Scholar] [CrossRef]
- Fatemi, A.; Wilson, M.A.; Johnston, M.V. Hypoxic-Ischemic Encephalopathy in the Term Infant. Clin. Perinatol. 2009, 36, 835–857. [Google Scholar] [CrossRef]
- Luhmann, H.J.; Kirischuk, S.; Sinning, A.; Kilb, W. Early GABAergic circuitry in the cerebral cortex. Curr. Opin. Neurobiol. 2014, 26, 72–78. [Google Scholar] [CrossRef]
- Tau, G.Z.; Peterson, B.S. Normal Development of Brain Circuits. Neuropsychopharmacology 2010, 35, 147–168. [Google Scholar] [CrossRef]
- Le Magueresse, C.; Monyer, H. GABAergic Interneurons Shape the Functional Maturation of the Cortex. Neuron 2013, 77, 388–405. [Google Scholar] [CrossRef]
- Lim, L.; Mi, D.; Llorca, A.; Marín, O. Development and functional diversification of cortical interneurons. Neuron 2018, 100, 294–313. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Scheinost, D.; Lacadie, C.; Benjamin, J.; Myers, E.H.; Qiu, M.; Schneider, K.C.; Rothman, D.L.; Constable, R.T.; Ment, L.R. GABA, Resting-State Connectivity and the Developing Brain. Neonatology 2014, 106, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Schuler, V.; Lüscher, C.; Blanchet, C.; Klix, N.; Sansig, G.; Klebs, K.; Schmutz, M.; Heid, J.; Gentry, C.; Urban, L.; et al. Epilepsy, Hyperalgesia, Impaired Memory, and Loss of Pre- and Postsynaptic GABAB Responses in Mice Lacking GABAB(1). Neuron 2001, 31, 47–58. [Google Scholar] [CrossRef]
- Panda, S.; Dohare, P.; Jain, S.; Parikh, N.; Singla, P.; Mehdizadeh, R.; Klebe, D.W.; Kleinman, G.M.; Cheng, B.; Ballabh, P. Estrogen treatment reverses prematurity-induced disruption in cortical interneuron population. J. Neurosci. 2018, 38, 7378–7391. [Google Scholar] [CrossRef] [PubMed]
- Stolp, H.B.; Fleiss, B.; Arai, Y.; Supramaniam, V.; Vontell, R.; Birtles, S.; Yates, A.G.; Baburamani, A.A.; Thornton, C.; Rutherford, M.; et al. Interneuron development is disrupted in preterm brains with diffuse white matter injury: Observations in mouse and human. Front. Physiol. 2019, 10, 955. [Google Scholar] [CrossRef]
- Robinson, S.; Li, Q.; DeChant, A.; Cohen, M.L. Neonatal loss of γ–aminobutyric acid pathway expression after human perinatal brain injury. J. Neurosurg. Pediatr. 2006, 104 (Suppl. S6), 396–408. [Google Scholar] [CrossRef]
- Lacaille, H.; Vacher, C.-M.; Bakalar, D.; O’Reilly, J.J.; Salzbank, J.; Penn, A.A. Impaired interneuron development in a novel model of neonatal brain injury. eNeuro 2019, 6, ENEURO.0300-18.2019. [Google Scholar] [CrossRef]
- Ardalan, M.; Svedin, P.; Baburamani, A.A.; Supramaniam, V.G.; Ek, J.; Hagberg, H.; Mallard, C. Dysmaturation of somatostatin interneurons following umbilical cord occlusion in preterm fetal sheep. Front. Physiol. 2019, 10, 563. [Google Scholar] [CrossRef]
- Nisimov, H.; Orenbuch, A.; Pleasure, S.J.; Golan, H.M. Impaired Organization of GABAergic Neurons Following Prenatal Hypoxia. Neuroscience 2018, 384, 300–313. [Google Scholar] [CrossRef]
- Komitova, M.; Xenos, D.; Salmaso, N.; Tran, K.M.; Brand, T.; Schwartz, M.L.; Ment, L.; Vaccarino, F.M. Hypoxia-induced developmental delays of inhibitory interneurons are reversed by environmental enrichment in the postnatal mouse forebrain. J. Neurosci. 2013, 33, 13375–13387. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Valdez, R.; Emerson, P.; Goffigan-Holmes, J.; Kirkwood, A.; Martin, L.J.; Northington, F.J. Delayed injury of hippocampal interneurons after neonatal hypoxia-ischemia and therapeutic hypothermia in a murine model. Hippocampus 2018, 28, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Davidson, J.O.; Fowke, T.M.; Galinsky, R.; Wassink, G.; Karunasinghe, R.N.; Prasad, J.D.; Ranasinghe, S.; Green, C.R.; Bennet, L.; et al. Connexin Hemichannel Mimetic Peptide Attenuates Cortical Interneuron Loss and Perineuronal Net Disruption Following Cerebral Ischemia in Near-Term Fetal Sheep. Int. J. Mol. Sci. 2020, 21, 6475. [Google Scholar] [CrossRef] [PubMed]
- Gunn, A.J.; Thoresen, M. Hypothermic neuroprotection. NeuroRx 2006, 3, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, G.; Laptook, A.; Shankaran, S. Therapeutic hypothermia: How can we optimize this therapy to further improve outcomes? Clin. Perinatol. 2018, 45, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.; Wassink, G.; Bennet, L.; Gunn, A.J.; Davidson, J.O. Can we further optimize therapeutic hypothermia for hypoxic-ischemic encephalopathy? Neural Regen. Res. 2019, 14, 1678–1683. [Google Scholar] [PubMed]
- Lemyre, B.; Chau, V. Hypothermia for newborns with hypoxic-ischemic encephalopathy. Paediatr. Child Health 2018, 23, 285–291. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischaemic encephalopathy. Cochrane Database Syst. Rev. 2013, 2013, CD003311. [Google Scholar] [CrossRef]
- Shankaran, S.; Laptook, A.R.; Pappas, A.; McDonald, S.A.; Das, A.; Tyson, J.E.; Poindexter, B.B.; Schibler, K.; Bell, E.F.; Heyne, R.J.; et al. Effect of depth and duration of cooling on death or disability at age 18 months among neonates with hypoxic-ischemic encephalopathy: A randomized clinical trial. JAMA 2017, 318, 57–67. [Google Scholar] [CrossRef]
- Davidson, J.O.; Wassink, G.; Yuill, C.A.; Zhang, F.G.; Bennet, L.; Gunn, A.J. How long is too long for cerebral cooling after ischemia in fetal sheep? J. Cereb. Blood Flow Metab. 2015, 35, 751–758. [Google Scholar] [CrossRef]
- Davidson, J.O.; Draghi, V.; Whitham, S.; Dhillon, S.K.; Wassink, G.; Bennet, L.; Gunn, A.J. How long is sufficient for optimal neuroprotection with cerebral cooling after ischemia in fetal sheep? J. Cereb. Blood Flow Metab. 2018, 38, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, X.; Tang, K. Interneuron development and dysfunction. FEBS J. 2022, 289, 2318–2336. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Anderson, S.A. Development of Cortical Interneurons. Neuropsychopharmacology 2015, 40, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Levitt, P.; Eagleson, K.L.; Powell, E.M. Regulation of neocortical interneuron development and the implications for neurodevelopmental disorders. Trends Neurosci. 2004, 27, 400–406. [Google Scholar] [CrossRef]
- Fowke, T.M.; Galinsky, R.; Davidson, J.O.; Wassink, G.; Karunasinghe, R.N.; Prasad, J.D.; Bennet, L.; Gunn, A.J.; Dean, J.M. Loss of interneurons and disruption of perineuronal nets in the cerebral cortex following hypoxia-ischaemia in near-term fetal sheep. Sci. Rep. 2018, 8, 17686. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.; Meng, X.; Liu, J.; Mikhailova, A.; Kao, J.P.Y.; McQuillen, P.; O Kanold, P. Neonatal Hypoxia–Ischemia Causes Functional Circuit Changes in Subplate Neurons. Cereb. Cortex 2019, 29, 765–776. [Google Scholar] [CrossRef]
- Failor, S.; Nguyen, V.; Darcy, D.P.; Cang, J.; Wendland, M.F.; Stryker, M.; McQuillen, P.S. Neonatal Cerebral Hypoxia-Ischemia Impairs Plasticity in Rat Visual Cortex. J. Neurosci. 2010, 30, 81–92. [Google Scholar] [CrossRef]
- Verney, C.; Rees, S.; Biran, V.; Thompson, M.; Inder, T.; Gressens, P. Neuronal Damage in the Preterm Baboon: Impact of the Mode of Ventilatory Support. J. Neuropathol. Exp. Neurol. 2010, 69, 473–482. [Google Scholar] [CrossRef]
- Tibrewal, M.; Cheng, B.; Dohare, P.; Hu, F.; Mehdizadeh, R.; Wang, P.; Zheng, D.; Ungvari, Z.; Ballabh, P. Disruption of interneuron neurogenesis in premature newborns and reversal with estrogen treatment. J. Neurosci. 2018, 38, 1100–1113. [Google Scholar] [CrossRef]
- Molloy, E.J.; El-Dib, M.; Juul, S.E.; Benders, M.; Gonzalez, F.; Bearer, C.; Wu, Y.W.; Robertson, N.J.; Hurley, T.; Branagan, A.; et al. Neuroprotective therapies in the NICU in term infants: Present and future. Pediatr. Res. 2022, 1–9. [Google Scholar] [CrossRef]
- Tremblay, R.; Lee, S.; Rudy, B. GABAergic interneurons in the neocortex: From cellular properties to circuits. Neuron 2016, 91, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.O.; Heuij, L.G.V.D.; Dhillon, S.K.; Miller, S.L.; Lim, R.; Jenkin, G.; Gunn, A.J.; Bennet, L. Lack of Neuroprotection with a Single Intravenous Infusion of Human Amnion Epithelial Cells after Severe Hypoxia–Ischemia in Near-Term Fetal Sheep. Int. J. Mol. Sci. 2022, 23, 8393. [Google Scholar] [CrossRef]
- Guan, J.; Bennet, L.; George, S.; Waldvogel, H.; Faull, R.; Gluckman, P.; Keunen, H.; Gunn, A. Selective neuroprotective effects with insulin-like growth factor-1 in phenotypic striatal neurons following ischemic brain injury in fetal sheep. Neuroscience 1999, 95, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Gunn, A.J.; Gunn, T.R.; Gunning, M.I.; Williams, C.E.; Gluckman, P.D. Neuroprotection With Prolonged Head Cooling Started Before Postischemic Seizures in Fetal Sheep. Pediatrics 1998, 102, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Wassink, G.; Davidson, J.O.; Fraser, M.; Yuill, C.A.; Bennet, L.; Gunn, A.J. Non-additive effects of adjunct erythropoietin therapy with therapeutic hypothermia after global cerebral ischaemia in near-term fetal sheep. J. Physiol. 2020, 598, 999–1015. [Google Scholar] [CrossRef]
- Davidson, J.; Green, C.R.; Nicholson, L.; O’Carroll, S.J.; Fraser, M.; Bennet, L.; Gunn, A.J. Connexin hemichannel blockade improves outcomes in a model of fetal ischemia. Ann. Neurol. 2012, 71, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.O.; Yuill, C.A.; Zhang, F.G.; Wassink, G.; Bennet, L.; Gunn, A.J. Extending the duration of hypothermia does not further improve white matter protection after ischemia in term-equivalent fetal sheep. Sci. Rep. 2016, 6, 25178. [Google Scholar] [CrossRef]
- Shankaran, S.; Laptook, A.R.; Pappas, A.; McDonald, S.A.; Das, A.; Tyson, J.E.; Poindexter, B.B.; Schibler, K.; Bell, E.F.; Heyne, R.J.; et al. Effect of depth and duration of cooling on deaths in the NICU among neonates with hypoxic ischemic encephalopathy: A randomized clinical trial. JAMA 2014, 312, 2629–2639. [Google Scholar] [CrossRef]
- Salman, M.M.; Kitchen, P.; Woodroofe, M.N.; Brown, J.E.; Bill, R.M.; Conner, A.C.; Conner, M.T. Hypothermia increases aquaporin 4 (AQP4) plasma membrane abundance in human primary cortical astrocytes via a calcium/transient receptor potential vanilloid 4 (TRPV4)- and calmodulin-mediated mechanism. Eur. J. Neurosci. 2017, 46, 2542–2547. [Google Scholar] [CrossRef]
- Han, H.S.; Park, J.; Kim, J.-H.; Suk, K. Molecular and Cellular Pathways as a Target of Therapeutic Hypothermia: Pharmacological Aspect. Curr. Neuropharmacol. 2012, 10, 80–87. [Google Scholar] [CrossRef]
- González-Ibarra, F.P.; Varon, J.; López-Meza, E.G. Therapeutic Hypothermia: Critical Review of the Molecular Mechanisms of Action. Front. Neurol. 2011, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.M.; Kitchen, P.; Woodroofe, M.N.; Bill, R.M.; Conner, A.C.; Heath, P.R.; Conner, M.T. Transcriptome Analysis of Gene Expression Provides New Insights into the Effect of Mild Therapeutic Hypothermia on Primary Human Cortical Astrocytes Cultured under Hypoxia. Front. Cell. Neurosci. 2017, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Michinaga, S.; Koyama, Y. Pathophysiological Responses and Roles of Astrocytes in Traumatic Brain Injury. Int. J. Mol. Sci. 2021, 22, 6418. [Google Scholar] [CrossRef] [PubMed]
- Wendimu, M.Y.; Hooks, S.B. Microglia Phenotypes in Aging and Neurodegenerative Diseases. Cells 2022, 11, 2091. [Google Scholar] [CrossRef]
- Schmidt, K.M.; Repine, M.J.; Hicks, S.D.; DeFranco, D.B.; Callaway, C.W. Regional changes in glial cell line-derived neurotrophic factor after cardiac arrest and hypothermia in rats. Neurosci. Lett. 2004, 368, 135–139. [Google Scholar] [CrossRef]
- Zhao, J.-F.; Ren, T.; Li, X.-Y.; Guo, T.-L.; Liu, C.-H.; Wang, X. Research Progress on the Role of Microglia Membrane Proteins or Receptors in Neuroinflammation and Degeneration. Front. Cell. Neurosci. 2022, 16, 831977. [Google Scholar] [CrossRef]
- Benmamar-Badel, A.; Owens, T.; Wlodarczyk, A. Protective Microglial Subset in Development, Aging, and Disease: Lessons From Transcriptomic Studies. Front. Immunol. 2020, 11, 430. [Google Scholar] [CrossRef]
- Fiumelli, H.; Kiraly, M.; Ambrus, A.; Magistretti, P.J.; Martin, J.-L. Opposite Regulation of Calbindin and Calretinin Expression by Brain-Derived Neurotrophic Factor in Cortical Neurons. J. Neurochem. 2000, 74, 1870–1877. [Google Scholar] [CrossRef]
- Meyer, H.S.; Schwarz, D.; Wimmer, V.C.; Schmitt, A.C.; Kerr, J.N.D.; Sakmann, B.; Helmstaedter, M. Inhibitory interneurons in a cortical column form hot zones of inhibition in layers 2 and 5A. Proc. Natl. Acad. Sci. USA 2011, 108, 16807–16812. [Google Scholar] [CrossRef]
- Benes, F.M.; Berretta, S. GABAergic Interneurons Implications for Understanding Schizophrenia and Bipolar Disorder. Neuropsychopharmacology 2001, 25, 1–27. [Google Scholar] [CrossRef]
- Gao, W.J.; Wormington, A.B.; Newman, D.E.; Pallas, S.L. Development of inhibitory circuitry in visual and auditory cortex of postnatal ferrets: Immunocytochemical localization of calbindin- and parvalbumin-containing neurons. J. Comp. Neurol. 2000, 422, 140–157. [Google Scholar] [CrossRef]
- Hendry, S.; Jones, E. GABA neuronal subpopulations in cat primary auditory cortex: Co-localization with calcium binding proteins. Brain Res. 1991, 543, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Bjerke, I.E.; Yates, S.C.; Laja, A.; Witter, M.P.; Puchades, M.A.; Bjaalie, J.G.; Leergaard, T.B. Densities and numbers of calbindin and parvalbumin positive neurons across the rat and mouse brain. iScience 2021, 24, 101906. [Google Scholar] [CrossRef] [PubMed]
- Gabbott, P.L.; Bacon, S. Local circuit neurons in the medial prefrontal cortex (areas 24a,b,c, 25 and 32) in the monkey: I. Cell morphology and morphometrics. J. Comp. Neurol. 1996, 364, 567–608. [Google Scholar] [CrossRef]
- Demeulemeester, H.; Vandesande, F.; Orban, G.; Heizmann, C.; Pochet, R. Calbindin D-28K and parvalbumin immunoreactivity is confined to two separate neuronal subpopulations in the cat visual cortex, whereas partial coexistence is shown in the dorsal lateral geniculate nucleus. Neurosci. Lett. 1989, 99, 6–11. [Google Scholar] [CrossRef]
- Kosaka, T.; Kosaka, K.; Nakayama, T.; Hunziker, W.; Heizmann, C.W. Axons and axon terminals of cerebellar Purkinje cells and basket cells have higher levels of parvalbumin immunoreactivity than somata and dendrites: Quantitative analysis by immunogold labeling. Exp. Brain Res. 1993, 93, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Hartwich, K.; Pollak, T.; Klausberger, T. Distinct Firing Patterns of Identified Basket and Dendrite-Targeting Interneurons in the Prefrontal Cortex during Hippocampal Theta and Local Spindle Oscillations. J. Neurosci. 2009, 29, 9563–9574. [Google Scholar] [CrossRef]
- Yan, Y.-H.; Van Brederode, J.F.M.; Hendrickson, A.E. Transient co-localization of calretinin, parvalbumin, and calbindin-D28k in developing visual cortex of monkey. J. Neurocytol. 1995, 24, 825–837. [Google Scholar] [CrossRef]
- Rudy, B.; Fishell, G.; Lee, S.; Hjerling-Leffler, J. Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 2011, 71, 45–61. [Google Scholar] [CrossRef]
- Nahar, L.; Delacroix, B.M.; Nam, H.W. The Role of Parvalbumin Interneurons in Neurotransmitter Balance and Neurological Disease. Front. Psychiatry 2021, 12, 679960. [Google Scholar] [CrossRef]
- Gonchar, Y.; Wang, Q.; Burkhalter, A. Multiple distinct subtypes of GABAergic neurons in mouse visual cortex identified by triple immunostaining. Front. Neuroanat. 2007, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Rupert, D.D.; Shea, S.D. Parvalbumin-Positive Interneurons Regulate Cortical Sensory Plasticity in Adulthood and Development Through Shared Mechanisms. Front. Neural Circuits 2022, 16, 886629. [Google Scholar] [CrossRef] [PubMed]
- Karnani, M.M.; Agetsuma, M.; Yuste, R. A blanket of inhibition: Functional inferences from dense inhibitory connectivity. Curr. Opin. Neurobiol. 2014, 26, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Gan, J.; Jonas, P. Interneurons. Fast-spiking, parvalbumin(+) GABAergic interneurons: From cellular design to microcircuit function. Science 2014, 345, 1255263. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal Oscillations in Cortical Networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Carlen, M.; Meletis, K.; Siegle, J.H.; Cardin, J.A.; Futai, K.; Vierling-Claassen, D.; Rühlmann, C.; Jones, S.R.; Deisseroth, K.; Sheng, M.; et al. A critical role for NMDA receptors in parvalbumin interneurons for gamma rhythm induction and behavior. Mol. Psychiatry 2012, 17, 537–548. [Google Scholar] [CrossRef]
- Herrera, T.I.; Edwards, L.; Malcolm, W.F.; Smith, P.B.; Fisher, K.A.; Pizoli, C.; Gustafson, K.E.; Goldstein, R.F.; Cotten, C.M.; Goldberg, R.N.; et al. Outcomes of preterm infants treated with hypothermia for hypoxic-ischemic encephalopathy. Early Hum. Dev. 2018, 125, 1–7. [Google Scholar] [CrossRef]
- Harris, E.; Abel, J.M.; Tejada, L.D.; Rissman, E.F. Calbindin Knockout Alters Sex-Specific Regulation of Behavior and Gene Expression in Amygdala and Prefrontal Cortex. Endocrinology 2016, 157, 1967–1979. [Google Scholar] [CrossRef]
- Ferrer, I.; Soriano, E. Postnatal development of parvalbumin and calbindin D28K immunoreactivities in the cerebral cortex of the rat. Anat. Embryol. 1993, 188, 63–73. [Google Scholar]
- Letinic, K.; Kostovic, I. Postnatal development of calcium-binding proteins calbindin and parvalbumin in human visual cortex. Cereb. Cortex 1998, 8, 660–669. [Google Scholar] [CrossRef]
- Zaitsev, A.; Gonzalez-Burgos, G.; Povysheva, N.; Kröner, S.; Lewis, D.; Krimer, L. Localization of Calcium-binding Proteins in Physiologically and Morphologically Characterized Interneurons of Monkey Dorsolateral Prefrontal Cortex. Cereb. Cortex 2005, 15, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-Z.; Takahashi, S.; Cui, C.; Inoue, M.; Fukui, Y. Distribution of calbindin-D28K immunoreactive neurons in rat primary motor cortex. J. Med. Investig. 2002, 49, 35–39. [Google Scholar]
- Fazzari, P.; Mortimer, N.; Yabut, O.; Vogt, D.; Pla, R. Cortical distribution of GABAergic interneurons is determined by migration time and brain size. Development 2020, 147, dev185033. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. Vet.-Clin. Pathol. 2012, 41, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Parsons, Y. Stereotaxic method and atlas for the ovine fetal forebrain. J. Dev. Physiol. 1983, 5, 101–128. [Google Scholar]
- Galinsky, R.; Davidson, J.O.; Lear, C.A.; Bennet, L.; Green, C.R.; Gunn, A.J. Connexin hemichannel blockade improves survival of striatal GABA-ergic neurons after global cerebral ischaemia in term-equivalent fetal sheep. Sci. Rep. 2017, 7, 6304. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution | Target | Source |
|---|---|---|---|
| Primary Antibodies | |||
| Gamma-aminobutyric acid 65/67 | 1:200 | GABAergic interneurons | Abcam, Melbourne, VIC, Australia |
| Parvalbumin | 1:200 | Parvalbumin interneurons | Swant Ltd., Marly, Switzerland |
| Calbindin | 1:200 | Calbindin interneurons | Swant Ltd. |
| Secondary Antibodies | |||
| Biotinylated goat anti-mouse IgG | 1:200 | Vector Laboratories, Burlingame, CA, USA | |
| Biotinylated goat anti-rabbit IgG | 1:200 | Vector Laboratories |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Davidson, J.O.; Zhou, K.Q.; Wilson, R.; Wassink, G.; Prasad, J.D.; Bennet, L.; Gunn, A.J.; Dean, J.M. Therapeutic Hypothermia Attenuates Cortical Interneuron Loss after Cerebral Ischemia in Near-Term Fetal Sheep. Int. J. Mol. Sci. 2023, 24, 3706. https://doi.org/10.3390/ijms24043706
Yang P, Davidson JO, Zhou KQ, Wilson R, Wassink G, Prasad JD, Bennet L, Gunn AJ, Dean JM. Therapeutic Hypothermia Attenuates Cortical Interneuron Loss after Cerebral Ischemia in Near-Term Fetal Sheep. International Journal of Molecular Sciences. 2023; 24(4):3706. https://doi.org/10.3390/ijms24043706
Chicago/Turabian StyleYang, Panzao, Joanne O. Davidson, Kelly Q. Zhou, Rani Wilson, Guido Wassink, Jaya D. Prasad, Laura Bennet, Alistair J. Gunn, and Justin M. Dean. 2023. "Therapeutic Hypothermia Attenuates Cortical Interneuron Loss after Cerebral Ischemia in Near-Term Fetal Sheep" International Journal of Molecular Sciences 24, no. 4: 3706. https://doi.org/10.3390/ijms24043706