
Monocytic HLA-DR Expression in Immune Responses of Acute Pancreatitis and COVID-19
 , ,
, ,
Abstract
:1. Introduction
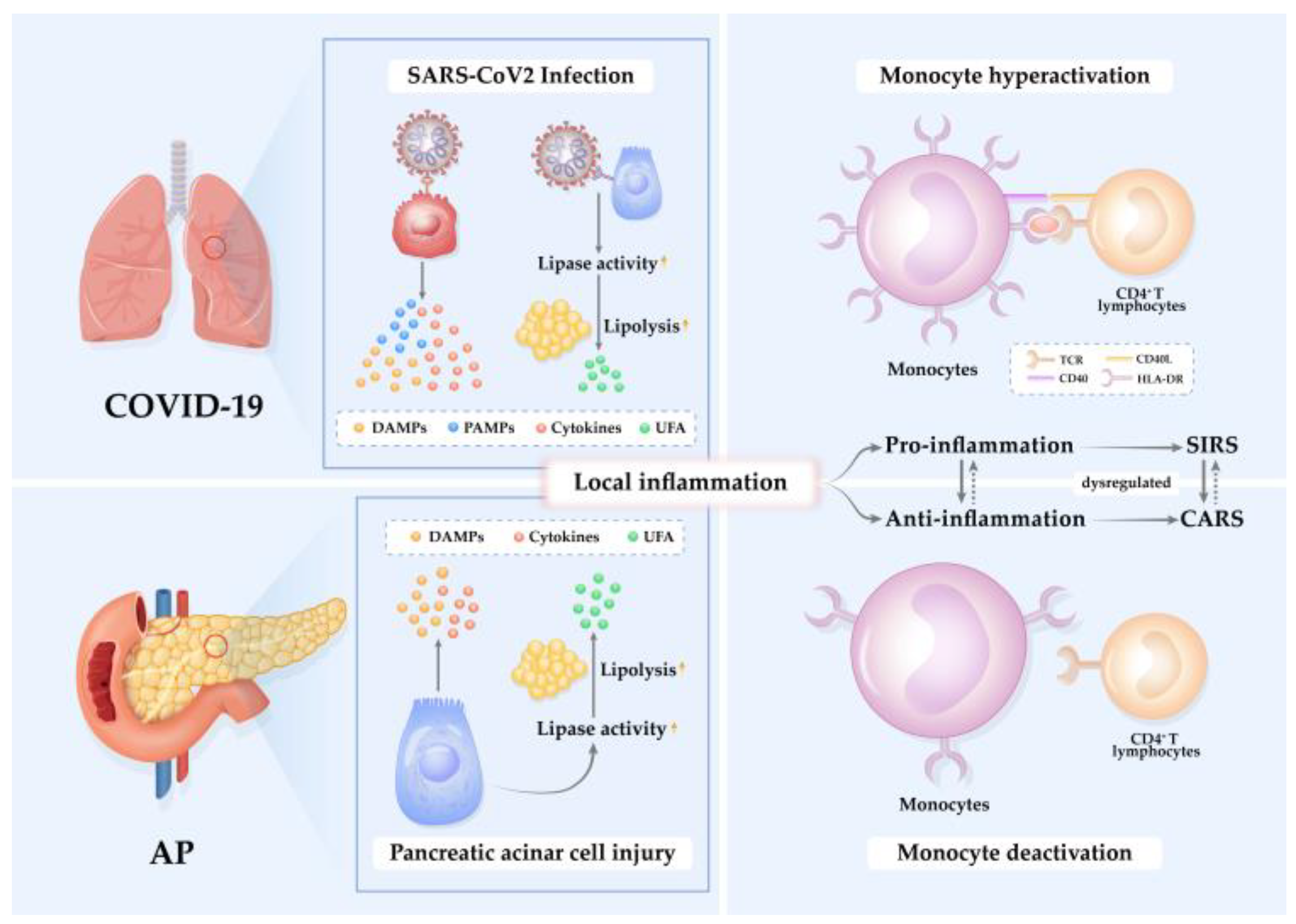
2. Pathogenesis and Immunopathology in AP and COVID-19
2.1. Pathophysiological Mechanisms in AP and COVID-19
2.2. Altered Immune Responses in AP and COVID-19
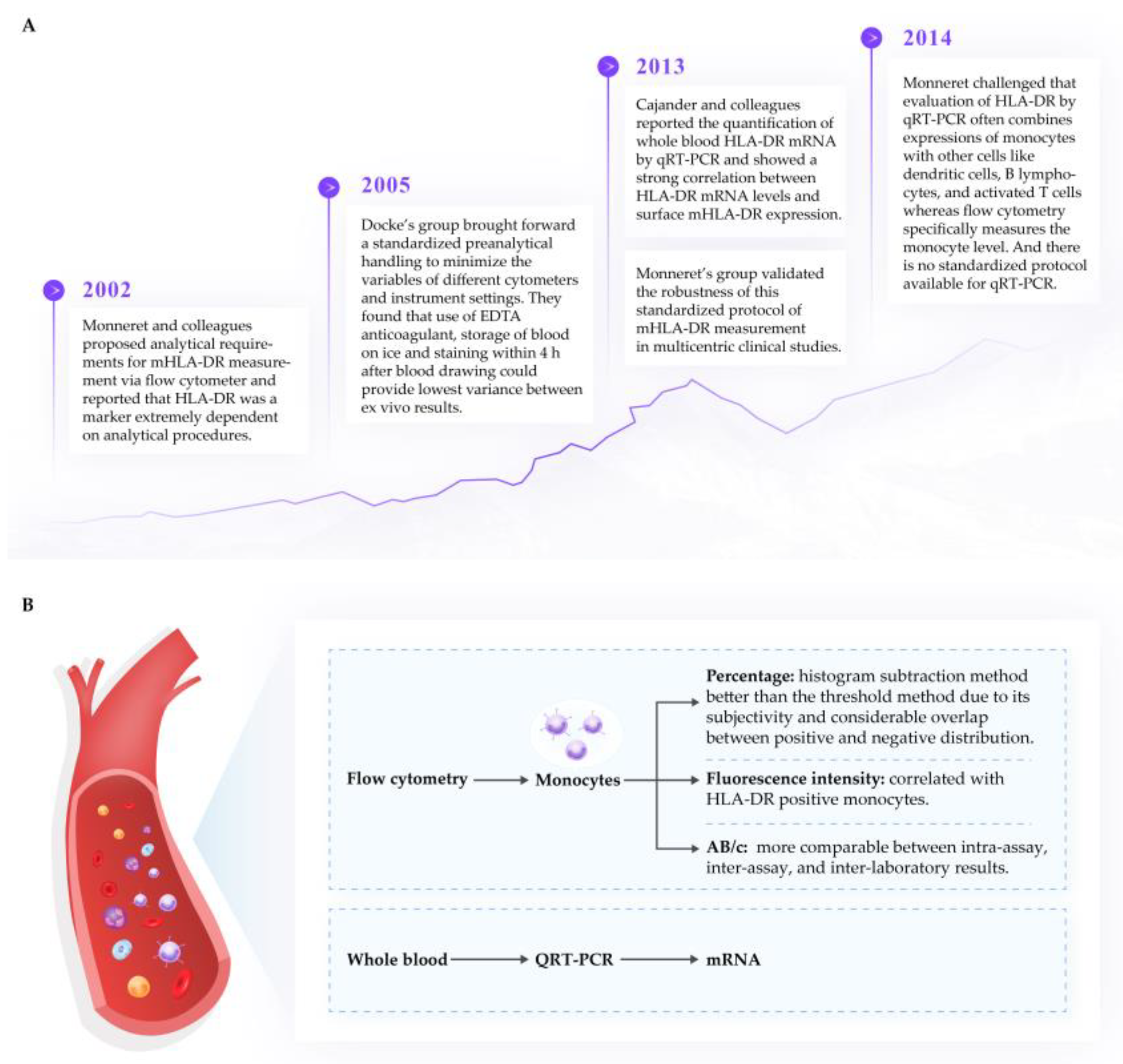
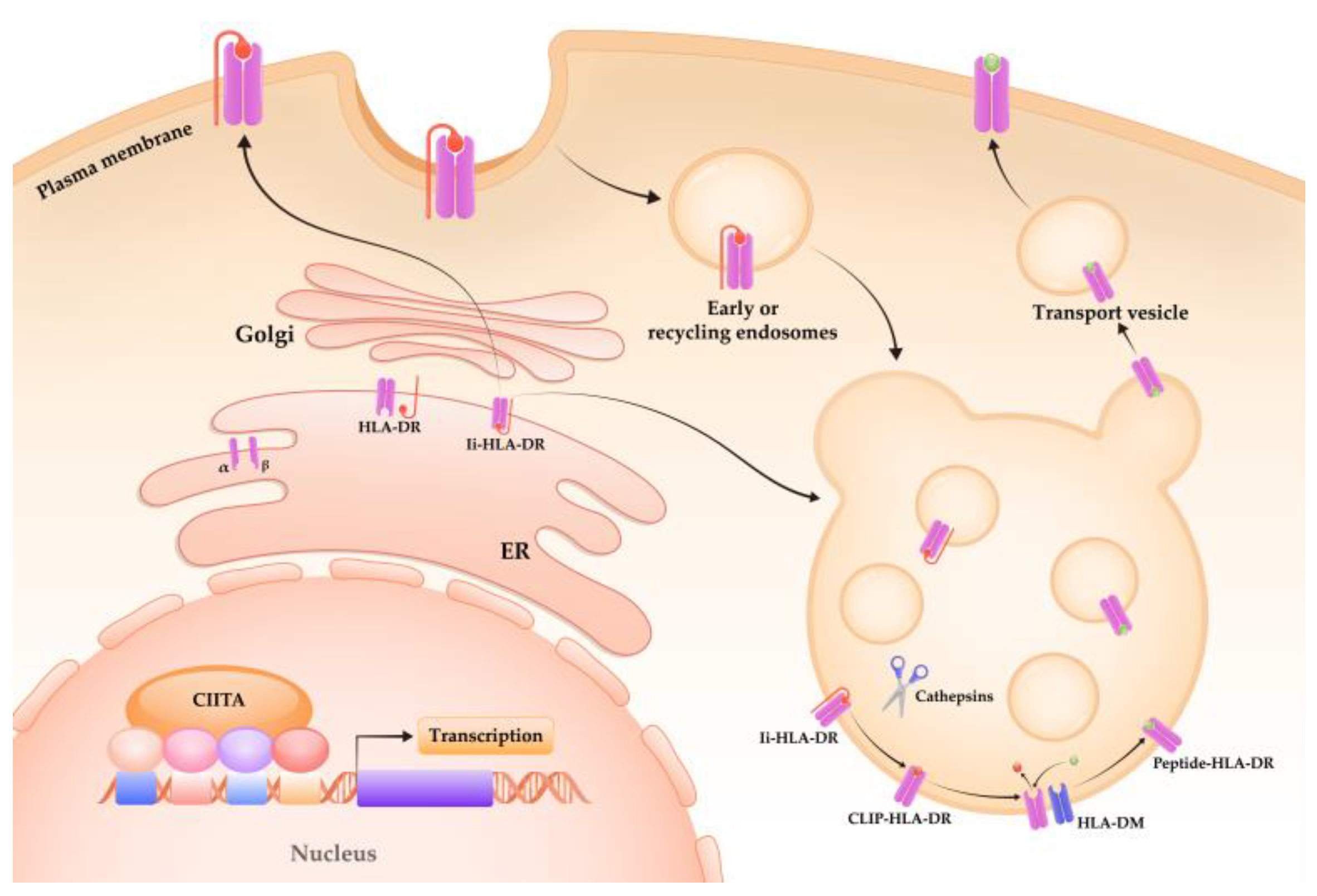
3. Structure and Expression of mHLA-DR
3.1. Measurement of mHLA-DR
3.2. Regulation of mHLA-DR Expression
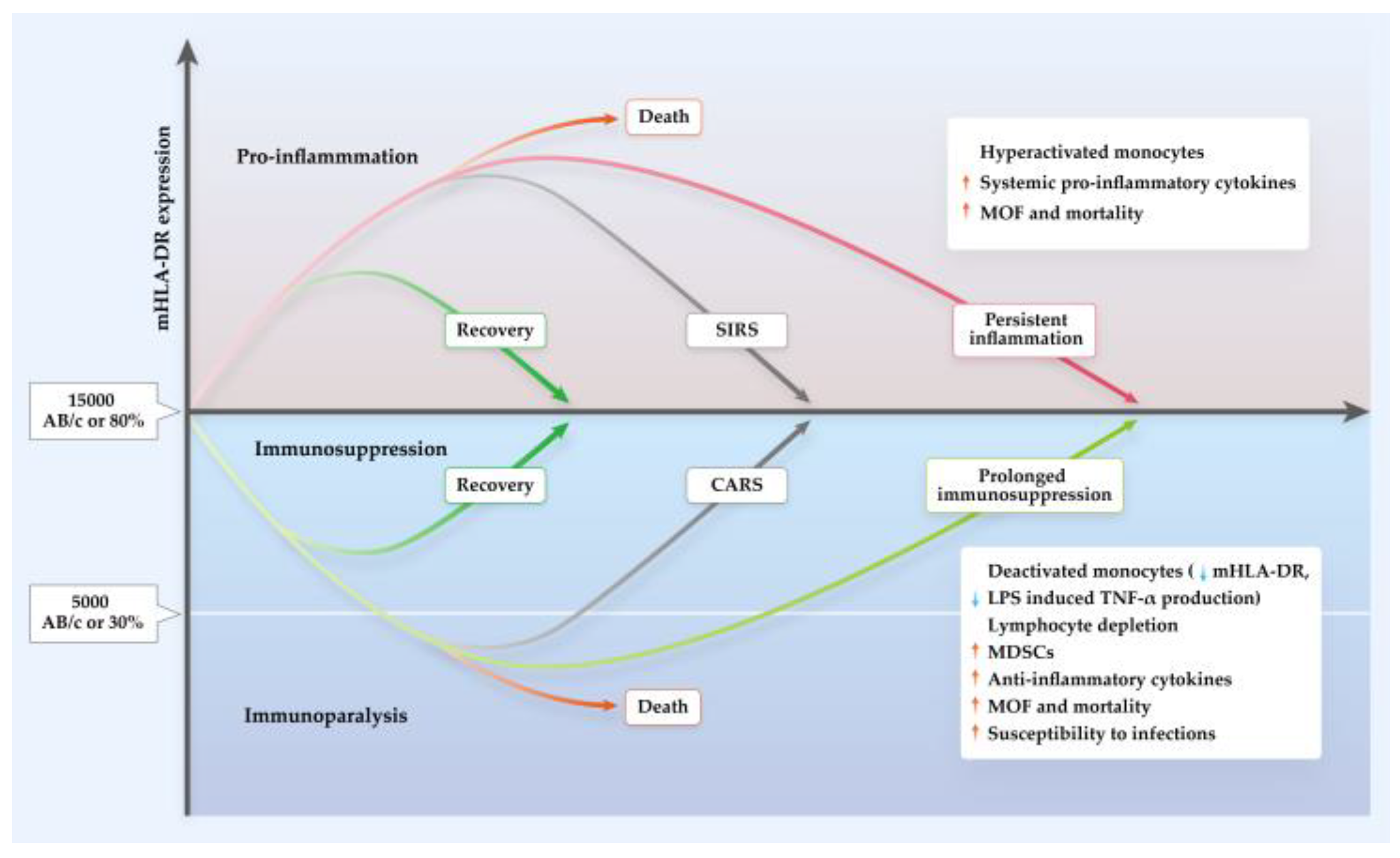
4. The Role of mHLA-DR in AP and COVID-19
4.1. Severity Prediction Using mHLA-DR in AP and COVID-19
4.2. Prediction of Infectious Complications using mHLA-DR
4.3. Regulation of mHLA-DR Expression in AP and COVID-19
4.4. Monocytic Myeloid-Derived Suppressor Cells
5. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iannuzzi, J.P.; King, J.A.; Leong, J.H.; Quan, J.; Windsor, J.W.; Tanyingoh, D.; Coward, S.; Forbes, N.; Heitman, S.J.; Shaheen, A.A.; et al. Global incidence of acute pancreatitis is increasing over time: A systematic review and meta-analysis. Gastroenterology 2022, 162, 122–134. [Google Scholar] [CrossRef]
- Li, C.L.; Jiang, M.; Pan, C.Q.; Li, J.; Xu, L.G. The global, regional, and national burden of acute pancreatitis in 204 countries and territories, 1990–2019. BMC Gastroenterol. 2021, 21, 332. [Google Scholar] [CrossRef] [PubMed]
- Szatmary, P.; Grammatikopoulos, T.; Cai, W.; Huang, W.; Mukherjee, R.; Halloran, C.; Beyer, G.; Sutton, R. Acute pancreatitis: Diagnosis and treatment. Drugs 2022, 82, 1251–1276. [Google Scholar] [CrossRef] [PubMed]
- Banks, P.A.; Bollen, T.L.; Dervenis, C.; Gooszen, H.G.; Johnson, C.D.; Sarr, M.G.; Tsiotos, G.G.; Vege, S.S.; Acute Pancreatitis Classification Working Group. Classification of acute pancreatitis--2012: Revision of the Atlanta classification and definitions by international consensus. Gut 2013, 62, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Linkermann, A.; Stockwell, B.R.; Krautwald, S.; Anders, H.J. Regulated cell death and inflammation: An auto-amplification loop causes organ failure. Nat. Rev. Immunol. 2014, 14, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Barreto, S.G.; Habtezion, A.; Gukovskaya, A.; Lugea, A.; Jeon, C.; Yadav, D.; Hegyi, P.; Venglovecz, V.; Sutton, R.; Pandol, S.J. Critical thresholds: Key to unlocking the door to the prevention and specific treatments for acute pancreatitis. Gut 2021, 70, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Li, A.; Xia, Q.; Liu, X.; Tian, B.; Mai, G.; Huang, Z.; Chen, G.; Tang, W.; Jin, X.; et al. The role of organ failure and infection in necrotizing pancreatitis: A prospective study. Ann. Surg. 2014, 259, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Sternby, H.; Bolado, F.; Canaval-Zuleta, H.J.; Marra-López, C.; Hernando-Alonso, A.I.; Del-Val-Antoñana, A.; García-Rayado, G.; Rivera-Irigoin, R.; Grau-García, F.J.; Oms, L.; et al. Determinants of severity in acute pancreatitis: A nation-wide multicenter prospective cohort study. Ann. Surg. 2019, 270, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Schepers, N.J.; Bakker, O.J.; Besselink, M.G.; Ahmed Ali, U.; Bollen, T.L.; Gooszen, H.G.; van Santvoort, H.C.; Bruno, M.J. Impact of characteristics of organ failure and infected necrosis on mortality in necrotising pancreatitis. Gut 2019, 68, 1044–1051. [Google Scholar] [CrossRef]
- Shi, N.; Liu, T.; de la Iglesia-Garcia, D.; Deng, L.; Jin, T.; Lan, L.; Zhu, P.; Hu, W.; Zhou, Z.; Singh, V.; et al. Duration of organ failure impacts mortality in acute pancreatitis. Gut 2020, 69, 604–605. [Google Scholar] [CrossRef] [Green Version]
- Petrov, M.S.; Shanbhag, S.; Chakraborty, M.; Phillips, A.R.; Windsor, J.A. Organ failure and infection of pancreatic necrosis as determinants of mortality in patients with acute pancreatitis. Gastroenterology 2010, 139, 813–820. [Google Scholar] [CrossRef]
- Werge, M.; Novovic, S.; Schmidt, P.N.; Gluud, L.L. Infection increases mortality in necrotizing pancreatitis: A systematic review and meta-analysis. Pancreatology 2016, 16, 698–707. [Google Scholar] [CrossRef]
- Moggia, E.; Koti, R.; Belgaumkar, A.P.; Fazio, F.; Pereira, S.P.; Davidson, B.R.; Gurusamy, K.S. Pharmacological interventions for acute pancreatitis. Cochrane Database Syst. Rev. 2017, 4, Cd011384. [Google Scholar] [CrossRef] [PubMed]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 20 September 2022).
- Coronavirus Disease 2019 (COVID-19) Treatment Guidelines; National Institutes of Health (US): Bethesda, MD, USA, 2021.
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef]
- Carvalho, T.; Krammer, F.; Iwasaki, A. The first 12 months of COVID-19: A timeline of immunological insights. Nat. Rev. Immunol. 2021, 21, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Thone, M.N.; Kwon, Y.J. COVID-19 vaccines: The status and perspectives in delivery points of view. Adv. Drug Deliv. Rev. 2021, 170, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Long, X.; Zhang, B.; Zhang, W.; Chen, X.; Zhang, Z. ACE2 expression in pancreas may cause pancreatic damage after SARS-CoV-2 infection. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2020, 18, 2128–2130.e2. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xu, Y.; Dong, Y.; Huang, Y.; Fu, Y.; Li, T.; Sun, C.; Pandanaboyana, S.; Windsor, J.A.; Fu, D. Prevalence and prognosis of increased pancreatic enzymes in patients with COVID-19: A systematic review and meta-analysis. Pancreatology 2022, 22, 539–546. [Google Scholar] [CrossRef]
- Goyal, H.; Sachdeva, S.; Perisetti, A.; Mann, R.; Inamdar, S.; Tharian, B. Hyperlipasemia and potential pancreatic injury patterns in COVID-19: A marker of severity or innocent bystander? Gastroenterology 2021, 160, 946–948.e2. [Google Scholar] [CrossRef]
- Kiyak, M.; Düzenli, T. Lipase elevation on admission predicts worse clinical outcomes in patients with COVID-19. Pancreatology 2022, 22, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Barkoudah, E.; Souza, D.A.T.; Jin, D.X.; McNabb-Baltar, J. Clinical course and outcome among patients with acute pancreatitis and COVID-19. Eur. J. Gastroenterol. Hepatol. 2021, 33, 695–700. [Google Scholar] [CrossRef]
- Szatmary, P.; Arora, A.; Thomas Raraty, M.G.; Joseph Dunne, D.F.; Baron, R.D.; Halloran, C.M. Emerging phenotype of severe acute respiratory syndrome-coronavirus 2-associated pancreatitis. Gastroenterology 2020, 159, 1551–1554. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, A.; Verellen, S.; Bruckers, L.; Geebelen, L.; Callebaut, I.; De Pauw, I.; Stessel, B.; Dubois, J. Secondary infection in COVID-19 critically ill patients: A retrospective single-center evaluation. BMC Infect. Dis. 2022, 22, 207. [Google Scholar] [CrossRef] [PubMed]
- Ripa, M.; Galli, L.; Poli, A.; Oltolini, C.; Spagnuolo, V.; Mastrangelo, A.; Muccini, C.; Monti, G.; De Luca, G.; Landoni, G.; et al. Secondary infections in patients hospitalized with COVID-19: Incidence and predictive factors. Clin. Microbiol. Infect. 2021, 27, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Gallo, C.G.; Fiorino, S.; Posabella, G.; Antonacci, D.; Tropeano, A.; Pausini, E.; Pausini, C.; Guarniero, T.; Hong, W.; Giampieri, E.; et al. COVID-19, what could sepsis, severe acute pancreatitis, gender differences, and aging teach us? Cytokine 2021, 148, 155628. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.Z.; Sidell, M.A.; Wu, B.U.; Setiawan, V.W.; Chen, Z.; Xiang, A.H. Pre-existing pancreatitis and elevated risks of COVID-19 severity and mortality. Gastroenterology 2022, 162, 1758–1760.e3. [Google Scholar] [CrossRef]
- Karki, R.; Kanneganti, T.D. The ‘cytokine storm’: Molecular mechanisms and therapeutic prospects. Trends Immunol. 2021, 42, 681–705. [Google Scholar] [CrossRef]
- Guéant, J.L.; Guéant-Rodriguez, R.M.; Fromonot, J.; Oussalah, A.; Louis, H.; Chery, C.; Gette, M.; Gleye, S.; Callet, J.; Raso, J.; et al. Elastase and exacerbation of neutrophil innate immunity are involved in multi-visceral manifestations of COVID-19. Allergy 2021, 76, 1846–1858. [Google Scholar] [CrossRef]
- de Vries, F.; Huckriede, J.; Wichapong, K.; Reutelingsperger, C.; Nicolaes, G.A.F. The role of extracellular histones in COVID-19. J. Intern. Med. 2022, 293, 275–292. [Google Scholar] [CrossRef]
- Lee, P.J.; Papachristou, G.L. New insights into acute pancreatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 479–496. [Google Scholar] [CrossRef]
- Szatmary, P.; Huang, W.; Criddle, D.; Tepikin, A.; Sutton, R. Biology, role and therapeutic potential of circulating histones in acute inflammatory disorders. J. Cell Mol. Med. 2018, 22, 4617–4629. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.M.; Lieberman, J.; Wu, H. Inflammasome activation at the crux of severe COVID-19. Nat. Rev. Immunol. 2021, 21, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, I.; Giroux, N.; Olson, L.; Morrison, S.A.; Llanga, T.; Akinade, T.O.; Zhu, Y.; Zhong, Y.; Bose, S.; Arvai, S.; et al. DAMPs/PAMPs induce monocytic TLR activation and tolerance in COVID-19 patients; nucleic acid binding scavengers can counteract such TLR agonists. Biomaterials 2022, 283, 121393. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Xue, J.; Jaffee, E.M.; Habtezion, A. Role of immune cells and immune-based therapies in pancreatitis and pancreatic ductal adenocarcinoma. Gastroenterology 2013, 144, 1230–1240. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Z.; Wang, Z.; Gutiérrez-Castrellón, P.; Shi, H. Cell deaths: Involvement in the pathogenesis and intervention therapy of COVID-19. Signal Transduct. Target. Ther. 2022, 7, 186. [Google Scholar] [CrossRef]
- Yamada, T.; Takaoka, A. Innate immune recognition against SARS-CoV-2. Inflamm. Regen. 2023, 43, 7. [Google Scholar] [CrossRef] [PubMed]
- Habtezion, A.; Gukovskaya, A.S.; Pandol, S.J. Acute pancreatitis: A multifaceted set of organelle and cellular interactions. Gastroenterology 2019, 156, 1941–1950. [Google Scholar] [CrossRef]
- Garg, P.K.; Singh, V.P. Organ failure due to systemic injury in acute pancreatitis. Gastroenterology 2019, 156, 2008–2023. [Google Scholar] [CrossRef]
- Xue, J.; Sharma, V.; Habtezion, A. Immune cells and immune-based therapy in pancreatitis. Immunol. Res. 2014, 58, 378–386. [Google Scholar] [CrossRef]
- Liu, S.; Szatmary, P.; Lin, J.W.; Wang, Q.; Sutton, R.; Chen, L.; Liu, T.; Huang, W.; Xia, Q. Circulating monocytes in acute pancreatitis. Front. Immunol. 2022, 13, 1062849. [Google Scholar] [CrossRef]
- Felsenstein, S.; Herbert, J.A.; McNamara, P.S.; Hedrich, C.M. COVID-19: Immunology and treatment options. Clin. Immunol. 2020, 215, 108448. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, P.; Smith, M.; Brand, M. Adaptive immune cell dysregulation and role in acute pancreatitis disease progression and treatment. Arch. Immunol. Et Ther. Exp. 2018, 66, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus infections and immune responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Anka, A.U.; Tahir, M.I.; Abubakar, S.D.; Alsabbagh, M.; Zian, Z.; Hamedifar, H.; Sabzevari, A.; Azizi, G. Coronavirus disease 2019 (COVID-19): An overview of the immunopathology, serological diagnosis and management. Scand. J. Immunol. 2021, 93, e12998. [Google Scholar] [CrossRef]
- Pietruczuk, M.; Dabrowska, M.I.; Wereszczynska-Siemiatkowska, U.; Dabrowski, A. Alteration of peripheral blood lymphocyte subsets in acute pancreatitis. World J. Gastroenterol. 2006, 12, 5344–5351. [Google Scholar] [CrossRef] [PubMed]
- Norman, J. The role of cytokines in the pathogenesis of acute pancreatitis. Am. J. Surg. 1998, 175, 76–83. [Google Scholar] [CrossRef]
- Makhija, R.; Kingsnorth, A.N. Cytokine storm in acute pancreatitis. J. Hepato-Biliary-Pancreat. Surg. 2002, 9, 401–410. [Google Scholar] [CrossRef]
- To, K.K.; Sridhar, S.; Chiu, K.H.; Hung, D.L.; Li, X.; Hung, I.F.; Tam, A.R.; Chung, T.W.; Chan, J.F.; Zhang, A.J.; et al. Lessons learned 1 year after SARS-CoV-2 emergence leading to COVID-19 pandemic. Emerg. Microbes Infect. 2021, 10, 507–535. [Google Scholar] [CrossRef]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef]
- Hegyi, P.; Szakács, Z.; Sahin-Tóth, M. Lipotoxicity and cytokine storm in severe acute pancreatitis and COVID-19. Gastroenterology 2020, 159, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.L.B.; Oliveira, R.A.; Severo, R.G.; Simões, E.S.A.C. COVID-19 related coagulopathy: What is Known Up to Now. Curr. Med. Chem. 2021, 28, 4207–4225. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Kothari, H.; Bosmann, M. Tissue factor in COVID-19-associated coagulopathy. Thromb. Res. 2022, 220, 35–47. [Google Scholar] [CrossRef]
- Bettac, L.; Denk, S.; Seufferlein, T.; Huber-Lang, M. Complement in pancreatic disease-perpetrator or savior? Front. Immunol. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Dumnicka, P.; Maduzia, D.; Ceranowicz, P.; Olszanecki, R.; Drożdż, R.; Kuśnierz-Cabala, B. The interplay between inflammation, coagulation and endothelial injury in the early phase of acute pancreatitis: Clinical implications. Int. J. Mol. Sci. 2017, 18, 354. [Google Scholar] [CrossRef]
- Kakafika, A.; Papadopoulos, V.; Mimidis, K.; Mikhailidis, D.P. Coagulation, platelets, and acute pancreatitis. Pancreas 2007, 34, 15–20. [Google Scholar] [CrossRef] [PubMed]
- El-Kurdi, B.; Khatua, B.; Rood, C.; Snozek, C.; Cartin-Ceba, R.; Singh, V.P. Mortality from coronavirus disease 2019 increases with unsaturated fat and may be reduced by early calcium and albumin supplementation. Gastroenterology 2020, 159, 1015–1018.e4. [Google Scholar] [CrossRef] [PubMed]
- Saccon, T.D.; Mousovich-Neto, F.; Ludwig, R.G.; Carregari, V.C.; Dos Anjos Souza, A.B.; Dos Passos, A.S.C.; Martini, M.C.; Barbosa, P.P.; de Souza, G.F.; Muraro, S.P.; et al. SARS-CoV-2 infects adipose tissue in a fat depot- and viral lineage-dependent manner. Nat. Commun. 2022, 13, 5722. [Google Scholar] [CrossRef]
- Patel, K.; Trivedi, R.N.; Durgampudi, C.; Noel, P.; Cline, R.A.; DeLany, J.P.; Navina, S.; Singh, V.P. Lipolysis of visceral adipocyte triglyceride by pancreatic lipases converts mild acute pancreatitis to severe pancreatitis independent of necrosis and inflammation. Am. J. Pathol. 2015, 185, 808–819. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.; Khatua, B.; Noel, P.; Kostenko, S.; Bag, A.; Balakrishnan, B.; Patel, K.S.; Guerra, A.A.; Martinez, M.N.; Trivedi, S.; et al. Pancreatic triglyceride lipase mediates lipotoxic systemic inflammation. J. Clin. Investig. 2020, 130, 1931–1947. [Google Scholar] [CrossRef] [Green Version]
- Navina, S.; Acharya, C.; DeLany, J.P.; Orlichenko, L.S.; Baty, C.J.; Shiva, S.S.; Durgampudi, C.; Karlsson, J.M.; Lee, K.; Bae, K.T.; et al. Lipotoxicity causes multisystem organ failure and exacerbates acute pancreatitis in obesity. Sci. Transl. Med. 2011, 3, 107ra110. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.B.; Kwon, H.J.; Sharif, M.; Lim, J.; Lee, I.C.; Ryu, Y.B.; Lee, J.I.; Kim, J.S.; Lee, Y.S.; Kim, D.H.; et al. Therapeutic strategy targeting host lipolysis limits infection by SARS-CoV-2 and influenza A virus. Signal Transduct. Target. Ther. 2022, 7, 367. [Google Scholar] [CrossRef] [PubMed]
- Meakins, J.L.; Pietsch, J.B.; Bubenick, O.; Kelly, R.; Rode, H.; Gordon, J.; MacLean, L.D. Delayed hypersensitivity: Indicator of acquired failure of host defenses in sepsis and trauma. Ann. Surg. 1977, 186, 241–250. [Google Scholar] [CrossRef]
- MacLean, L.D.; Meakins, J.L.; Taguchi, K.; Duignan, J.P.; Dhillon, K.S.; Gordon, J. Host resistance in sepsis and trauma. Ann. Surg. 1975, 182, 207–217. [Google Scholar] [CrossRef]
- Monneret, G.; Gossez, M.; Aghaeepour, N.; Gaudilliere, B.; Venet, F. How clinical flow cytometry rebooted sepsis immunology. Cytom. Part A J. Int. Soc. Anal. Cytol. 2019, 95, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sabrido, J.L.; Valdecantos, E.; Bastida, E.; Tellado, J.M. The anergic state as a predictor of pancreatic sepsis. Zent. Fur Chir. 1989, 114, 114–120. [Google Scholar]
- Mayerle, J.; Dummer, A.; Sendler, M.; Malla, S.R.; van den Brandt, C.; Teller, S.; Aghdassi, A.; Nitsche, C.; Lerch, M.M. Differential roles of inflammatory cells in pancreatitis. J. Gastroenterol. Hepatol. 2012, 27 (Suppl. 2), 47–51. [Google Scholar] [CrossRef]
- Jain, S.; Midha, S.; Mahapatra, S.J.; Gupta, S.; Sharma, M.K.; Nayak, B.; Jacob, T.G.; Shalimar; Garg, P.K. Interleukin-6 significantly improves predictive value of systemic inflammatory response syndrome for predicting severe acute pancreatitis. Pancreatology 2018, 18, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M. Acute pancreatitis as a model of SIRS. Front. Biosci. (Landmark Ed.) 2009, 14, 2042–2050. [Google Scholar] [CrossRef]
- Bone, R.C. Sir Isaac Newton, sepsis, SIRS, and CARS. Crit. Care Med. 1996, 24, 1125–1128. [Google Scholar] [CrossRef]
- Zhulai, G.A.; Oleinik, E.K.; Ostrovskii, K.A.; Oleinik, V.M.; Kravchenko, P.N.; Churov, A.V. Alterations of lymphocyte subsets and indicators of immune suppression in patients with acute pancreatitis. Eksp. Klin. Gastroenterol. 2014, 9, 21–25. [Google Scholar]
- Döcke, W.D.; Randow, F.; Syrbe, U.; Krausch, D.; Asadullah, K.; Reinke, P.; Volk, H.D.; Kox, W. Monocyte deactivation in septic patients: Restoration by IFN-gamma treatment. Nat. Med. 1997, 3, 678–681. [Google Scholar] [CrossRef]
- Volk, H.D.; Reinke, P.; Krausch, D.; Zuckermann, H.; Asadullah, K.; Müller, J.M.; Döcke, W.D.; Kox, W.J. Monocyte deactivation--rationale for a new therapeutic strategy in sepsis. Intensive Care Med. 1996, 22 (Suppl. 4), S474–S481. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Lepape, A.; Monneret, G. Clinical review: Flow cytometry perspectives in the ICU-from diagnosis of infection to monitoring of injury-induced immune dysfunctions. Crit. Care 2011, 15, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in sepsis: A novel understanding of the disorder and a new therapeutic approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Leentjens, J.; Kox, M.; van der Hoeven, J.G.; Netea, M.G.; Pickkers, P. Immunotherapy for the adjunctive treatment of sepsis: From immunosuppression to immunostimulation. Time for a paradigm change? Am. J. Respir. Crit. Care Med. 2013, 187, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J. What is the pathophysiology of the septic host upon admission? Int. J. Antimicrob. Agents 2010, 36 (Suppl. 2), S2–S5. [Google Scholar] [CrossRef] [PubMed]
- Minkov, G.A.; Halacheva, K.S.; Yovtchev, Y.P.; Gulubova, M.V. Pathophysiological mechanisms of acute pancreatitis define inflammatory markers of clinical prognosis. Pancreas 2015, 44, 713–717. [Google Scholar] [CrossRef]
- Zhuang, Y.G.; Peng, H.; Chen, Y.Z.; Zhou, S.Q.; Chen, Y.Q. Dynamic monitoring of monocyte HLA-DR expression for the diagnosis, prognosis, and prediction of sepsis. Front. Biosci.-Landmrk 2017, 22, 1344–1354. [Google Scholar]
- Li, J.; Yang, W.-J.; Huang, L.-M.; Tang, C.-W. Immunomodulatory therapies for acute pancreatitis. World J. Gastroenterol. 2014, 20, 16935–16947. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.K.; Levy, M.M.; Ward, N.S. Biomarkers of Immunosuppression. Crit. Care Clin. 2020, 36, 167–176. [Google Scholar] [CrossRef]
- Benlyamani, I.; Venet, F.; Coudereau, R.; Gossez, M.; Monneret, G. Monocyte HLA-DR measurement by flow cytometry in COVID-19 patients: An interim review. Cytom. Part A J. Int. Soc. Anal. Cytol. 2020, 97, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Jones, E.K.; Rubin, S.J.S.; Subrahmanyam, P.B.; Swaminathan, G.; Mikhail, D.; Bai, L.; Singh, G.; Wei, Y.; Sharma, V.; et al. Novel circulating and tissue monocytes as well as macrophages in pancreatitis and recovery. Gastroenterology 2021, 161, 2014–2029.e14. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Benincasa, G.; Criscuolo, C.; Faenza, M.; Liberato, C.; Rusciano, M. Immune reactivity during COVID-19: Implications for treatment. Immunol. Lett. 2021, 231, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Volk, H.D.; Thieme, M.; Heym, S.; Döcke, W.D.; Ruppe, U.; Tausch, W.; Manger, D.; Zuckermann, S.; Golosubow, A.; Nieter, B.; et al. Alterations in function and phenotype of monocytes from patients with septic disease--predictive value and new therapeutic strategies. Behring. Inst. Mitt. 1991, 88, 208–215. [Google Scholar]
- Lin, R.Y.; Astiz, M.E.; Saxon, J.C.; Rackow, E.C. Altered leukocyte immunophenotypes in septic shock. Studies of HLA-DR, CD11b, CD14, and IL-2R expression. Chest 1993, 104, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Ditschkowski, M.; Kreuzfelder, E.; Rebmann, V.; Ferencik, S.; Majetschak, M.; Schmid, E.N.; Obertacke, U.; Hirche, H.; Schade, U.F.; Grosse-Wilde, H. HLA-DR expression and soluble HLA-DR levels in septic patients after trauma. Ann. Surg. 1999, 229, 246–254. [Google Scholar] [CrossRef]
- Wakefield, C.H.; Carey, P.D.; Foulds, S.; Monson, J.R.; Guillou, P.J. Changes in major histocompatibility complex class II expression in monocytes and T cells of patients developing infection after surgery. Br. J. Surg. 1993, 80, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Sachse, C.; Prigge, M.; Cramer, G.; Pallua, N.; Henkel, E. Association between reduced human leukocyte antigen (HLA)-DR expression on blood monocytes and increased plasma level of interleukin-10 in patients with severe burns. Clin. Chem. Lab. Med. 1999, 37, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.F.; Auffray, C.; Korman, A.J.; Shackelford, D.A.; Strominger, J. The class II molecules of the human and murine major histocompatibility complex. Cell 1984, 36, 1–13. [Google Scholar] [CrossRef]
- Unanue, E.R.; Turk, V.; Neefjes, J. Variations in MHC Class II antigen processing and presentation in health and disease. Annu. Rev. Immunol. 2016, 34, 265–297. [Google Scholar] [CrossRef]
- Crux, N.B.; Elahi, S. Human leukocyte antigen (HLA) and immune regulation: How do classical and non-classical HLA alleles modulate immune response to human immunodeficiency virus and hepatitis C virus infections? Front. Immunol. 2017, 8, 832. [Google Scholar] [CrossRef]
- Mosaad, Y.M. Clinical role of human leukocyte antigen in health and disease. Scand. J. Immunol. 2015, 82, 283–306. [Google Scholar] [CrossRef]
- Monneret, G.; Venet, F. Sepsis-induced immune alterations monitoring by flow cytometry as a promising tool for individualized therapy. Cytom. B Clin. Cytom. 2016, 90, 376–386. [Google Scholar] [CrossRef]
- Monneret, G.; Venet, F. Monocyte HLA-DR in sepsis: Shall we stop following the flow? Crit. Care 2014, 18, 102. [Google Scholar] [CrossRef]
- Monneret, G.; Elmenkouri, N.; Bohe, J.; Debard, A.L.; Gutowski, M.C.; Bienvenu, J.; Lepape, A. Analytical requirements for measuring monocytic human lymphocyte antigen DR by flow cytometry: Application to the monitoring of patients with septic shock. Clin. Chem. 2002, 48, 1589–1592. [Google Scholar] [CrossRef] [PubMed]
- Docke, W.D.; Hoflich, C.; Davis, K.A.; Rottgers, K.; Meisel, C.; Kiefer, P.; Weber, S.U.; Hedwig-Geissing, M.; Kreuzfelder, E.; Tschentscher, P.; et al. Monitoring temporary immunodepression by flow cytometric measurement of monocytic HLA-DR expression: A multicenter standardized study. Clin. Chem. 2005, 51, 2341–2347. [Google Scholar] [CrossRef] [PubMed]
- Demaret, J.; Walencik, A.; Jacob, M.C.; Timsit, J.F.; Venet, F.; Lepape, A.; Monneret, G. Inter-laboratory assessment of flow cytometric monocyte HLA-DR expression in clinical samples. Cytom. B Clin. Cytom. 2013, 84, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Cajander, S.; Backman, A.; Tina, E.; Stralin, K.; Soderquist, B.; Kallman, J. Preliminary results in quantitation of HLA-DRA by real-time PCR: A promising approach to identify immunosuppression in sepsis. Crit. Care 2013, 17, R223. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, P.J. Expression regulation of major histocompatibility complex class I and class II encoding genes. Front. Immunol. 2011, 2, 48. [Google Scholar] [CrossRef]
- Steimle, V.; Otten, L.A.; Zufferey, M.; Mach, B. Complementation cloning of an MHC class II transactivator mutated in hereditary MHC class II deficiency (or bare lymphocyte syndrome). Cell 1993, 75, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Miatello, J.; Lukaszewicz, A.C.; Carter, M.J.; Faivre, V.; Hua, S.; Martinet, K.Z.; Bourgeois, C.; Quintana-Murci, L.; Payen, D.; Boniotto, M.; et al. CIITA promoter polymorphism impairs monocytes HLA-DR expression in patients with septic shock. iScience 2022, 25, 105291. [Google Scholar] [CrossRef] [PubMed]
- Pishesha, N.; Harmand, T.J.; Ploegh, H.L. A guide to antigen processing and presentation. Nat. Rev. Immunol. 2022, 22, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Viville, S.; Neefjes, J.; Lotteau, V.; Dierich, A.; Lemeur, M.; Ploegh, H.; Benoist, C.; Mathis, D. Mice lacking the MHC class II-associated invariant chain. Cell 1993, 72, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Kunz, S.; Crompton, N.E.; Volk, H.D.; Sabat, R. Multiple mechanisms of reduced major histocompatibility complex class II expression in endotoxin tolerance. J. Biol. Chem. 2003, 278, 18030–18036. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, C.; Karlsson, L. Nonclassical MHC class II molecules. Annu. Rev. Immunol. 2000, 18, 113–142. [Google Scholar] [CrossRef] [PubMed]
- Fung-Leung, W.P.; Surh, C.D.; Liljedahl, M.; Pang, J.; Leturcq, D.; Peterson, P.A.; Webb, S.R.; Karlsson, L. Antigen presentation and T cell development in H2-M-deficient mice. Science 1996, 271, 1278–1281. [Google Scholar] [CrossRef]
- Pinet, V.; Vergelli, M.; Martin, R.; Bakke, O.; Long, E.O. Antigen presentation mediated by recycling of surface HLA-DR molecules. Nature 1995, 375, 603–606. [Google Scholar] [CrossRef]
- Neefjes, J.; Jongsma, M.L.M.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef]
- Koppelman, B.; Neefjes, J.J.; de Vries, J.E.; de Waal Malefyt, R. Interleukin-10 down-regulates MHC class II alphabeta peptide complexes at the plasma membrane of monocytes by affecting arrival and recycling. Immunity 1997, 7, 861–871. [Google Scholar] [CrossRef]
- Ting, J.P.; Trowsdale, J. Genetic control of MHC class II expression. Cell 2002, 109 (Suppl. 1), S21–S33. [Google Scholar] [CrossRef]
- Czarniecki, C.W.; Chiu, H.H.; Wong, G.H.; McCabe, S.M.; Palladino, M.A. Transforming growth factor-beta 1 modulates the expression of class II histocompatibility antigens on human cells. J. Immunol. 1988, 140, 4217–4223. [Google Scholar] [CrossRef]
- Piskurich, J.F.; Wang, Y.; Linhoff, M.W.; White, L.C.; Ting, J.P. Identification of distinct regions of 5’ flanking DNA that mediate constitutive, IFN-gamma, STAT1, and TGF-beta-regulated expression of the class II transactivator gene. J. Immunol. 1998, 160, 233–240. [Google Scholar] [CrossRef]
- Lu, H.T.; Riley, J.L.; Babcock, G.T.; Huston, M.; Stark, G.R.; Boss, J.M.; Ransohoff, R.M. Interferon (IFN) beta acts downstream of IFN-gamma-induced class II transactivator messenger RNA accumulation to block major histocompatibility complex class II gene expression and requires the 48-kD DNA-binding protein, ISGF3-gamma. J. Exp. Med. 1995, 182, 1517–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershman, M.J.; Appel, S.H.; Wellhausen, S.R.; Sonnenfeld, G.; Polk, H.C., Jr. Interferon-gamma treatment increases HLA-DR expression on monocytes in severely injured patients. Clin Exp Immunol 1989, 77, 67–70. [Google Scholar] [PubMed]
- Blanar, M.A.; Boettger, E.C.; Flavell, R.A. Transcriptional activation of HLA-DR alpha by interferon gamma requires a trans-acting protein. Proc. Natl. Acad. Sci. USA 1988, 85, 4672–4676. [Google Scholar] [CrossRef] [PubMed]
- Pierre, P.; Turley, S.J.; Gatti, E.; Hull, M.; Meltzer, J.; Mirza, A.; Inaba, K.; Steinman, R.M.; Mellman, I. Developmental regulation of MHC class II transport in mouse dendritic cells. Nature 1997, 388, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Engering, A.; Pinet, V.; Pieters, J.; Lanzavecchia, A. Inflammatory stimuli induce accumulation of MHC class II complexes on dendritic cells. Nature 1997, 388, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Sepsis-induced immunosuppression: From cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013, 13, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Yu, M.; Chai, Y. Pathological alteration and therapeutic implications of sepsis-induced immune cell apoptosis. Cell Death Dis. 2019, 10, 782. [Google Scholar] [CrossRef] [PubMed]
- Kylanpaa-Back, M.L.; Takala, A.; Kemppainen, E.; Puolakkainen, P.; Kautiainen, H.; Jansson, S.E.; Haapiainen, R.; Repo, H. Cellular markers of systemic inflammation and immune suppression in patients with organ failure due to severe acute pancreatitis. Scand. J. Gastroenterol. 2001, 36, 1100–1107. [Google Scholar]
- Richter, A.; Nebe, T.; Wendl, K.; Schuster, K.; Klaebisch, G.; Quintel, M.; Lorenz, D.; Post, S.; Trede, M. HLA-DR expression in acute pancreatitis. Eur. J. Surg. 1999, 165, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Turunen, A.; Kuuliala, A.; Penttilä, A.; Kaukonen, K.M.; Mustonen, H.; Pettilä, V.; Puolakkainen, P.; Kylänpää, L.; Kuuliala, K. Time course of signaling profiles of blood leukocytes in acute pancreatitis and sepsis. Scand. J. Clin. Lab. Investig. 2020, 80, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Li, J.P.; Yang, J.; Huang, J.R.; Jiang, D.L.; Zhang, F.; Liu, M.F.; Qiang, Y.; Gu, Y.L. Immunosuppression and the infection caused by gut mucosal barrier dysfunction in patients with early severe acute pancreatitis. Front. Biosci. (Landmark Ed.) 2013, 18, 892–900. [Google Scholar] [PubMed]
- Mentula, P.; Kylanpaa, M.L.; Kemppainen, E.; Jansson, S.E.; Sarna, S.; Puolakkainen, P.; Haapiainen, R.; Repo, H. Plasma anti-inflammatory cytokines and monocyte human leucocyte antigen-DR expression in patients with acute pancreatitis. Scand. J. Gastroenterol. 2004, 39, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.K.; Li, W.Q.; Li, N.; Li, J.S. Mononuclear histocompatibility leukocyte antigen-DR expression in the early phase of acute pancreatitis. Pancreatology 2004, 4, 233–243. [Google Scholar] [CrossRef]
- Walter, L.O.; Cardoso, C.C.; Santos-Pirath, Í.M.; Costa, H.Z.; Gartner, R.; Werle, I.; Mohr, E.T.B.; da Rosa, J.S.; Felisberto, M.; Kretzer, I.F.; et al. The relationship between peripheral immune response and disease severity in SARS-CoV-2-infected subjects: A cross-sectional study. Immunology 2022, 165, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wen, Y.; Qi, F.; Gao, X.; Chen, W.; Xu, G.; Wei, C.; Wang, H.; Tang, X.; Lin, J.; et al. Suppressive monocytes impair MAIT cells response via IL-10 in patients with severe COVID-19. J. Immunol. 2021, 207, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Gatti, A.; Radrizzani, D.; Viganò, P.; Mazzone, A.; Brando, B. Decrease of non-classical and intermediate monocyte subsets in severe acute SARS-CoV-2 infection. Cytom. Part A J. Int. Soc. Anal. Cytol. 2020, 97, 887–890. [Google Scholar] [CrossRef]
- Mudd, P.A.; Crawford, J.C.; Turner, J.S.; Souquette, A.; Reynolds, D.; Bender, D.; Bosanquet, J.P.; Anand, N.J.; Striker, D.A.; Martin, R.S.; et al. Distinct inflammatory profiles distinguish COVID-19 from influenza with limited contributions from cytokine storm. Sci. Adv. 2020, 6, eabe3024. [Google Scholar] [CrossRef]
- Xu, G.; Qi, F.; Li, H.; Yang, Q.; Wang, H.; Wang, X.; Liu, X.; Zhao, J.; Liao, X.; Liu, Y.; et al. The differential immune responses to COVID-19 in peripheral and lung revealed by single-cell RNA sequencing. Cell Discov. 2020, 6, 73. [Google Scholar] [CrossRef]
- Bonnet, B.; Cosme, J.; Dupuis, C.; Coupez, E.; Adda, M.; Calvet, L.; Fabre, L.; Saint-Sardos, P.; Bereiziat, M.; Vidal, M.; et al. Severe COVID-19 is characterized by the co-occurrence of moderate cytokine inflammation and severe monocyte dysregulation. EBioMedicine 2021, 73, 103622. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.E.; Jørgensen, M.J.; Nore, K.G.; Dahl, T.B.; Yang, K.; Ranheim, T.; Huse, C.; Lind, A.; Nur, S.; Stiksrud, B.; et al. Critical COVID-19 is associated with distinct leukocyte phenotypes and transcriptome patterns. J. Intern. Med. 2021, 290, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Jiang, Y.; Wei, X.; Liu, X.; Guan, J.; Chen, Y.; Lu, H.; Qian, J.; Wang, Z.; Lin, X. Dynamic changes in monocytes subsets in COVID-19 patients. Hum. Immunol. 2021, 82, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Zenarruzabeitia, O.; Astarloa-Pando, G.; Terrén, I.; Orrantia, A.; Pérez-Garay, R.; Seijas-Betolaza, I.; Nieto-Arana, J.; Imaz-Ayo, N.; Pérez-Fernández, S.; Arana-Arri, E.; et al. T cell activation, highly armed cytotoxic cells and a shift in monocytes CD300 receptors expression is characteristic of patients with severe COVID-19. Front. Immunol. 2021, 12, 655934. [Google Scholar] [CrossRef] [PubMed]
- Cizmecioglu, A.; Emsen, A.; Sumer, S.; Ergun, D.; Akay Cizmecioglu, H.; Turk Dagi, H.; Artac, H. Reduced monocyte subsets, their HLA-DR expressions, and relations to acute phase reactants in severe COVID-19 cases. Viral Immunol. 2022, 35, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Hammad, R.; Kotb, H.G.; Eldesoky, G.A.; Mosaad, A.M.; El-Nasser, A.M.; Abd El Hakam, F.E.; Eldesoky, N.A.; Mashaal, A.; Farhoud, H. Utility of monocyte expression of HLA-DR versus T lymphocyte frequency in the assessment of COVID-19 outcome. Int. J. Gen. Med. 2022, 15, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Gossez, M.; Bidar, F.; Bodinier, M.; Coudereau, R.; Lukaszewicz, A.C.; Tardiveau, C.; Brengel-Pesce, K.; Cheynet, V.; Cazalis, M.A.; et al. T cell response against SARS-CoV-2 persists after one year in patients surviving severe COVID-19. EBioMedicine 2022, 78, 103967. [Google Scholar] [CrossRef]
- Marais, C.; Claude, C.; Semaan, N.; Charbel, R.; Barreault, S.; Travert, B.; Piloquet, J.E.; Demailly, Z.; Morin, L.; Merchaoui, Z.; et al. Myeloid phenotypes in severe COVID-19 predict secondary infection and mortality: A pilot study. Ann. Intensive Care 2021, 11, 111. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex immune dysregulation in COVID-19 patients with severe respiratory failure. Cell Host Microbe 2020, 27, 992–1000.e3. [Google Scholar] [CrossRef]
- Moser, D.; Biere, K.; Han, B.; Hoerl, M.; Schelling, G.; Choukér, A.; Woehrle, T. COVID-19 impairs immune response to candida albicans. Front. Immunol. 2021, 12, 640644. [Google Scholar] [CrossRef]
- Utrero-Rico, A.; González-Cuadrado, C.; Chivite-Lacaba, M.; Cabrera-Marante, O.; Laguna-Goya, R.; Almendro-Vazquez, P.; Díaz-Pedroche, C.; Ruiz-Ruigómez, M.; Lalueza, A.; Folgueira, M.D.; et al. Alterations in circulating monocytes predict COVID-19 severity and include chromatin modifications still detectable six months after recovery. Biomedicines 2021, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Mairpady Shambat, S.; Gómez-Mejia, A.; Schweizer, T.A.; Huemer, M.; Chang, C.C.; Acevedo, C.; Bergada-Pijuan, J.; Vulin, C.; Hofmaenner, D.A.; Scheier, T.C.; et al. Hyperinflammatory environment drives dysfunctional myeloid cell effector response to bacterial challenge in COVID-19. PLoS Pathog. 2022, 18, e1010176. [Google Scholar] [CrossRef] [PubMed]
- Boumaza, A.; Gay, L.; Mezouar, S.; Bestion, E.; Diallo, A.B.; Michel, M.; Desnues, B.; Raoult, D.; La Scola, B.; Halfon, P.; et al. Monocytes and macrophages, targets of severe acute respiratory syndrome coronavirus 2: The clue for coronavirus disease 2019 immunoparalysis. J. Infect. Dis. 2021, 224, 395–406. [Google Scholar] [CrossRef]
- Laing, A.G.; Lorenc, A.; Del Molino Del Barrio, I.; Das, A.; Fish, M.; Monin, L.; Muñoz-Ruiz, M.; McKenzie, D.R.; Hayday, T.S.; Francos-Quijorna, I.; et al. A dynamic COVID-19 immune signature includes associations with poor prognosis. Nat. Med. 2020, 26, 1623–1635. [Google Scholar] [CrossRef] [PubMed]
- Penttilä, P.A.; Van Gassen, S.; Panovska, D.; Vanderbeke, L.; Van Herck, Y.; Quintelier, K.; Emmaneel, A.; Filtjens, J.; Malengier-Devlies, B.; Ahmadzadeh, K.; et al. High dimensional profiling identifies specific immune types along the recovery trajectories of critically ill COVID19 patients. Cell Mol. Life Sci. 2021, 78, 3987–4002. [Google Scholar] [CrossRef]
- Zhang, R.; Shi, J.; Zhang, R.; Ni, J.; Habtezion, A.; Wang, X.; Hu, G.; Xue, J. Expanded CD14(hi)CD16(-) Immunosuppressive monocytes predict disease severity in patients with acute pancreatitis. J. Immunol. 2019, 202, 2578–2584. [Google Scholar] [CrossRef]
- Minkov, G.; Dimitrov, E.; Yovtchev, Y.; Enchev, E.; Lokova, R.; Halacheva, K. Prognostic value of peripheral blood CD14+HLA-DR+ monocytes in patients with acute pancreatitis. J. Immunoass. Immunochem. 2021, 42, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Mentula, P.; Kylanpaa-Back, M.L.; Kemppainen, E.; Takala, A.; Jansson, S.E.; Kautiainen, H.; Puolakkainen, P.; Haapiainen, R.; Repo, H. Decreased HLA (human leucocyte antigen)-DR expression on peripheral blood monocytes predicts the development of organ failure in patients with acute pancreatitis. Clin. Sci. 2003, 105, 409–417. [Google Scholar] [CrossRef]
- Ho, Y.P.; Sheen, I.S.; Chiu, C.T.; Wu, C.S.; Lin, C.Y. A strong association between down-regulation of HLA-DR expression and the late mortality in patients with severe acute pancreatitis. Am. J. Gastroenterol. 2006, 101, 1117–1124. [Google Scholar] [CrossRef]
- Yang, M.; Liu, Z.; Kang, K.; Yu, K.; Wang, C. Letter to the Editor: CD14(+)HLA-DR(+) Cells in patients may be a biomarker reflecting the progression of COVID-19. Viral Immunol. 2021, 34, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Satoh, A.; Miura, T.; Satoh, K.; Masamune, A.; Yamagiwa, T.; Sakai, Y.; Shibuya, K.; Takeda, K.; Kaku, M.; Shimosegawa, T. Human leukocyte antigen-DR expression on peripheral monocytes as a predictive marker of sepsis during acute pancreatitis. Pancreas 2002, 25, 245–250. [Google Scholar] [CrossRef]
- Kylanpaa, M.L.; Mentula, P.; Kemppainen, E.; Puolakkainen, P.; Aittomaki, S.; Silvennoinen, O.; Haapiainen, R.; Repo, H. Monocyte anergy is present in patients with severe acute pancreatitis and is significantly alleviated by granulocyte-macrophage colony-stimulating factor and interferon-gamma in vitro. Pancreas 2005, 31, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.P.; Chiu, C.T.; Sheen, I.S.; Tseng, S.C.; Lai, P.C.; Ho, S.Y.; Chen, W.T.; Lin, T.N.; Lin, C.Y. Tumor necrosis factor-alpha and interleukin-10 contribute to immunoparalysis in patients with acute pancreatitis. Hum. Immunol. 2011, 72, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.Q.; Guo, J.; Xia, Q.; Yang, X.N.; Huang, W.; Huang, Z.W.; Xue, P. Human leukocyte antigen-DR expression on peripheral monocytes may be an early marker for secondary infection in severe acute pancreatitis. Hepatogastroenterology 2013, 60, 1896–1902. [Google Scholar] [PubMed]
- Gotzinger, P.; Sautner, T.; Spittler, A.; Barlan, M.; Wamser, P.; Roth, E.; Jakesz, R.; Fugger, R. Severe acute pancreatitis causes alterations in HLA-DR and CD14 expression on peripheral blood monocytes independently of surgical treatment. Eur. J. Surg. 2000, 166, 628–632. [Google Scholar] [PubMed]
- Wolk, K.; Hoflich, C.; Zuckermann-Becker, H.; Docke, W.D.; Volk, H.D.; Sabat, R. Reduced monocyte CD86 expression in postinflammatory immunodeficiency. Crit. Care Med. 2007, 35, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, C.; Yu, M.; Zhao, X.; Du, J.; Li, Y.; Jing, H.; Dong, Z.; Kou, J.; Bi, Y.; et al. Neutrophil extracellular traps induced by activated platelets contribute to procoagulant activity in patients with colorectal cancer. Thromb. Res. 2019, 180, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Ferat-Osorio, E.; Wong-Baeza, I.; Esquivel-Callejas, N.; Figueroa-Figueroa, S.; Duarte-Rojo, A.; Guzman-Valdivia-Gomez, G.; Rodea-Rosas, H.; Torres-Gonzalez, R.; Sanchez-Fernandez, P.; Arriaga-Pizano, L.; et al. Triggering receptor expressed on myeloid cells-1 expression on monocytes is associated with inflammation but not with infection in acute pancreatitis. Crit. Care 2009, 13, R69. [Google Scholar] [CrossRef] [PubMed]
- Hue, S.; Beldi-Ferchiou, A.; Bendib, I.; Surenaud, M.; Fourati, S.; Frapard, T.; Rivoal, S.; Razazi, K.; Carteaux, G.; Delfau-Larue, M.H.; et al. Uncontrolled innate and impaired adaptive immune responses in patients with COVID-19 acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Peruzzi, B.; Bencini, S.; Capone, M.; Mazzoni, A.; Maggi, L.; Salvati, L.; Vanni, A.; Orazzini, C.; Nozzoli, C.; Morettini, A.; et al. Quantitative and qualitative alterations of circulating myeloid cells and plasmacytoid DC in SARS-CoV-2 infection. Immunology 2020, 161, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Hou, H.; Yao, Y.; Wu, S.; Huang, M.; Ran, X.; Zhou, H.; Liu, Z.; Sun, Z. Systemically comparing host immunity between survived and deceased COVID-19 patients. Cell Mol. Immunol. 2020, 17, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Pfortmueller, C.A.; Meisel, C.; Fux, M.; Schefold, J.C. Assessment of immune organ dysfunction in critical illness: Utility of innate immune response markers. Intensive Care Med. Exp. 2017, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Boomer, J.S.; To, K.; Chang, K.C.; Takasu, O.; Osborne, D.F.; Walton, A.H.; Bricker, T.L.; Jarman, S.D., 2nd; Kreisel, D.; Krupnick, A.S.; et al. Immunosuppression in patients who die of sepsis and multiple organ failure. JAMA 2011, 306, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Monneret, G.; Lepape, A.; Venet, F. A dynamic view of mHLA-DR expression in management of severe septic patients. Crit. Care 2011, 15, 198. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.M.; Sankaran, S.J.; Plank, L.D.; Windsor, J.A.; Petrov, M.S. Meta-analysis of gut barrier dysfunction in patients with acute pancreatitis. Br. J. Surg. 2014, 101, 1644–1656. [Google Scholar] [CrossRef]
- Qin, Y.; Pinhu, L.; You, Y.; Sooranna, S.; Huang, Z.; Zhou, X.; Yin, Y.; Song, S. The role of Fas expression on the occurrence of immunosuppression in severe acute pancreatitis. Dig. Dis. Sci. 2013, 58, 3300–3307. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Jakkampudi, A.; Reddy, R.; Reddy, P.B.; Patil, A.; Murthy, H.V.V.; Rao, G.V.; Reddy, D.N.; Talukdar, R. Association of systemic inflammatory and anti-inflammatory responses with adverse outcomes in acute pancreatitis: Preliminary results of an ongoing study. Dig. Dis. Sci. 2017, 62, 3468–3478. [Google Scholar] [CrossRef]
- Yu, Z.X.; Chen, X.C.; Zhang, B.Y.; Liu, N.; Gu, Q. Association between HLA-DR expression and multidrug-resistant infection in patients with severe acute pancreatitis. Curr. Med. Sci. 2018, 38, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J. Complex immune deregulation in severe COVID-19: More than a mechanism of pathogenesis. EBioMedicine 2021, 73, 103673. [Google Scholar] [CrossRef]
- Loftus, T.J.; Ungaro, R.; Dirain, M.; Efron, P.A.; Mazer, M.B.; Remy, K.E.; Hotchkiss, R.S.; Zhong, L.; Bacher, R.; Starostik, P.; et al. Overlapping but disparate inflammatory and immunosuppressive responses to SARS-CoV-2 and bacterial sepsis: An immunological time course analysis. Front. Immunol. 2021, 12, 792448. [Google Scholar] [CrossRef]
- Ding, L.; Wan, M.; Wang, D.; Cao, H.; Wang, H.; Gao, P. Myeloid-derived suppressor cells in patients with acute pancreatitis with increased inhibitory function. Front. Immunol. 2022, 13, 840620. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Notari, S.; Gili, S.; Bordoni, V.; Casetti, R.; Cimini, E.; Tartaglia, E.; Mariotti, D.; Agrati, C.; Sacchi, A. Myeloid-derived suppressor cells in COVID-19: The paradox of good. Front. Immunol. 2022, 13, 842949. [Google Scholar] [CrossRef]
- Falck-Jones, S.; Österberg, B.; Smed-Sörensen, A. Respiratory and systemic monocytes, dendritic cells and myeloid-derived suppressor cells in COVID-19: Implications for disease severity. J. Intern. Med. 2022, 293, 130–143. [Google Scholar] [CrossRef]
- Kylänpää, M.L.; Repo, H.; Puolakkainen, P.A. Inflammation and immunosuppression in severe acute pancreatitis. World J. Gastroenterol. 2010, 16, 2867–2872. [Google Scholar] [CrossRef] [PubMed]
- Monneret, G.; Venet, F.; Meisel, C.; Schefold, J.C. Assessment of monocytic HLA-DR expression in ICU patients: Analytical issues for multicentric flow cytometry studies. Crit. Care 2010, 14, 432. [Google Scholar] [CrossRef] [PubMed]
- De Waele, J.; Vogelaers, D.; Decruyenaere, J.; De Vos, M.; Colardyn, F. Infectious complications of acute pancreatitis. Acta Clin. Belg. 2004, 59, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Leppäniemi, A.; Tolonen, M.; Tarasconi, A.; Segovia-Lohse, H.; Gamberini, E.; Kirkpatrick, A.W.; Ball, C.G.; Parry, N.; Sartelli, M.; Wolbrink, D.; et al. 2019 WSES guidelines for the management of severe acute pancreatitis. World J. Emerg. Surg. 2019, 14, 27. [Google Scholar] [CrossRef]
- Estella, Á.; Vidal-Cortés, P.; Rodríguez, A.; Andaluz Ojeda, D.; Martín-Loeches, I.; Díaz, E.; Suberviola, B.; Gracia Arnillas, M.P.; Catalán González, M.; Álvarez-Lerma, F.; et al. Management of infectious complications associated with coronavirus infection in severe patients admitted to ICU. Med. Intensiv. 2021, 45, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Dickel, S.; Grimm, C.; Amschler, K.; Schnitzler, S.U.; Schanz, J.; Moerer, O.; Payen, D.; Tampe, B.; Winkler, M.S. Case report: Interferon-γ restores monocytic human leukocyte antigen receptor (mHLA-DR) in severe COVID-19 with acquired immunosuppression syndrome. Front. Immunol. 2021, 12, 645124. [Google Scholar] [CrossRef]
- van Laarhoven, A.; Kurver, L.; Overheul, G.J.; Kooistra, E.J.; Abdo, W.F.; van Crevel, R.; Duivenvoorden, R.; Kox, M.; Ten Oever, J.; Schouten, J.; et al. Interferon gamma immunotherapy in five critically ill COVID-19 patients with impaired cellular immunity: A case series. Med (N. Y.) 2021, 2, 1163–1170.e2. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.S.; Ait Hamou, Z.; Gastli, N.; Chapuis, N.; Pène, F. Potential role for interferon gamma in the treatment of recurrent ventilator-acquired pneumonia in patients with COVID-19: A hypothesis. Intensive Care Med. 2021, 47, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Laterre, P.F.; François, B.; Collienne, C.; Hantson, P.; Jeannet, R.; Remy, K.E.; Hotchkiss, R.S. Association of interleukin 7 immunotherapy with lymphocyte counts among patients with severe coronavirus disease 2019 (COVID-19). JAMA Netw. Open 2020, 3, e2016485. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Zhou, J.; Mao, W.; Chen, T.; Zhu, Y.; Pan, X.; Mei, H.; Singh, V.; Buxbaum, J.; Doig, G.; et al. Immune enhancement in patients with predicted severe acute necrotising pancreatitis: A multicentre double-blind randomised controlled trial. Intensive Care Med. 2022, 48, 899–909. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokines | HLA-DR Expression | Regulatory Mechanisms | References |
|---|---|---|---|
| IL-10 | ↓ | Downregulation of CIITA; Altering vesicular traffic of HLA-DR in exocytosis and recycling | [112,113] |
| TGF-β | ↓ | Inhibition of CIITA and downregulation of HLA-DR transcription | [114,115] |
| IFN-β | ↓ | Downregulation of CIITA | [116] |
| IFN-γ | ↑ | Promotion of HLA-DR and CD74 transcription | [117,118] |
| GM-CSF | ↑ | Promoting exocytosis and reducing internalization | [119] |
| TNF-α, IL-1 | ↑ | Boosting biosynthesis and stability of HLA-DR increasing half-life from about 10 h to over 100 h | [120] |
| IL-4 | ↑ | Upregulation of CIITA | [113] |
| Disease | Prediction | Sample Size (Incidents/Total) | Measuring Time | Cut-Off Value | AUC | Sensitivity | Specificity | Others |
|---|---|---|---|---|---|---|---|---|
| AP [149] | MAP from MSAP/SAP | 27/50 | Admission | <2274 MFI | 0.805 | 70.4% | 82.6% | Combined with classical monocyte proportions (AUC, 0.862) |
| SAP from MSAP | 9/23 | <1094.5 MFI | 0.690 | 85.7% | 55.6% | - | ||
| AP [150] | SAP | 19/58 | Admission | <50.8% | 0.728 | 72% | 72% | - |
| Day 2 | <43.35% | 0.800 | 84% | 80% | - | |||
| Day 5 | <60.8% | 0.877 | 82% | 78% | - | |||
| AP [151] | Organ failure | 29/310 | Admission | <78% | 0.78 | 83% | 72% | - |
| <38 RFU | 0.81 | 69% | 84% | - | ||||
| AP [152] | Mortality | 7/25 | Day 10 | <52.3% | 0.944 | 94.4% | 85.7% | - |
| COVID-19 [136] | Critical COVID-19 | 9/32 | Days 0–3 | <81.55% | 0.961 | 100.00% | 80.00% | - |
| COVID-19 [139] | Severe COVID-19 | 48/97 | Admission | <143 MFI | 0.9 | 89.6% | 81.6% | Independent predictor of COVID-19 severity (OR = 0.976, 95% CI: 0.955–0.997) |
| COVID-19 [134] | Mortality | 35/124 | Days 0–3 | <11,312 AB/c | 0.64 | 74% | 54% | - |
| Days 7–10 | <4672 AB/c | 0.85 | 75% | 86% | - | |||
| COVID-19 [153] | Mortality | 1/12 | Admission | <270.56 cells/mL | 0.875 | 100.0% | 87.5% | - |
| Disease | Prediction | Sample Size (Incidents/Total) | Measuring Time | Cut-Off Value | AUC | Sensitivity | Specificity | Others |
|---|---|---|---|---|---|---|---|---|
| AP [154] | Sepsis | 6/64 | Admission | <60% | - | 100% | 91.3% | Superior to Ranson’s score and APACHE II score |
| Day 7 | - | 100% | 93.2% | - | ||||
| Day 14 | - | 100% | 98.2% | - | ||||
| AP [128] | Septic complications | 11/74 | Day 7 | <40% | - | 73% | 94% | - |
| Day 10 | - | 82% | 98% | - | ||||
| Day 14 | - | 100% | 100% | - | ||||
| AP [152] | Septic complications | 6/25 | Day 10 | <58% | 0.926 | 76.5% | 100% | Comparable to Ranson and APACHE II scores and better than CRP levels (AUC, 0.841, 0.869, and 0.460, respectively) |
| AP [157] | Secondary infection | 11/40 | Admission | <35.8% | 0.837 | 81.8% | 82.8% | - |
| COVID-19 [134] | Secondary infection | 38/124 | Days 0–3 | <10,523 AB/c | 0.70 | 76% | 60% | - |
| Days 7–10 | <6804 AB/c | 0.62 | 77% | 52% | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Luo, W.; Szatmary, P.; Zhang, X.; Lin, J.-W.; Chen, L.; Liu, D.; Sutton, R.; Xia, Q.; Jin, T.; et al. Monocytic HLA-DR Expression in Immune Responses of Acute Pancreatitis and COVID-19. Int. J. Mol. Sci. 2023, 24, 3246. https://doi.org/10.3390/ijms24043246
Liu S, Luo W, Szatmary P, Zhang X, Lin J-W, Chen L, Liu D, Sutton R, Xia Q, Jin T, et al. Monocytic HLA-DR Expression in Immune Responses of Acute Pancreatitis and COVID-19. International Journal of Molecular Sciences. 2023; 24(4):3246. https://doi.org/10.3390/ijms24043246
Chicago/Turabian StyleLiu, Shiyu, Wenjuan Luo, Peter Szatmary, Xiaoying Zhang, Jing-Wen Lin, Lu Chen, Dan Liu, Robert Sutton, Qing Xia, Tao Jin, and et al. 2023. "Monocytic HLA-DR Expression in Immune Responses of Acute Pancreatitis and COVID-19" International Journal of Molecular Sciences 24, no. 4: 3246. https://doi.org/10.3390/ijms24043246