
Molecular Responses of Vegetable, Ornamental Crops, and Model Plants to Salinity Stress



1
Department of Science Veterinary, Università degli Studi di Messina, 98168 Messina, Italy
2
Department of Agriculture, Food and Environment, Università degli Studi di Catania, 95131 Catania, Italy
3
Department of Agricultural and Environmental Sciences—Production, Landscape, Agroenergy, Università degli Studi di Milano, 20133 Milan, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(4), 3190; https://doi.org/10.3390/ijms24043190
Submission received: 11 January 2023
/
Revised: 2 February 2023
/
Accepted: 4 February 2023
/
Published: 6 February 2023
(This article belongs to the Special Issue Molecular Aspects of Plant Salinity Stress and Tolerance 2.0)
Abstract
:Vegetable and ornamental plants represent a very wide group of heterogeneous plants, both herbaceous and woody, generally without relevant salinity-tolerant mechanisms. The cultivation conditions—almost all are irrigated crops—and characteristics of the products, which must not present visual damage linked to salt stress, determine the necessity for a deep investigation of the response of these crops to salinity stress. Tolerance mechanisms are linked to the capacity of a plant to compartmentalize ions, produce compatible solutes, synthesize specific proteins and metabolites, and induce transcriptional factors. The present review critically evaluates advantages and disadvantages to study the molecular control of salt tolerance mechanisms in vegetable and ornamental plants, with the aim of distinguishing tools for the rapid and effective screening of salt tolerance levels in different plants. This information can not only help in suitable germplasm selection, which is very useful in consideration of the high biodiversity expressed by vegetable and ornamental plants, but also drive the further breeding activities.
1. Introduction
Salinity is a major abiotic stresses that reduces crop productivity by hampering physiological processes in plants [1,2,3]. It affects approximately 19.5% of agricultural lands covering more than 830 million hectares [4]. Problems associated with salinity stress in the near future will expand due to global climate changes, in particular with the increase in temperature and reduction in water availability [5]. High salinity levels are a problem for vegetable and flower crops—which are irrigated crops—because soil salinity is often the result of incorrect irrigation practices that cause an increase in the concentration of salt in topsoil layers [6]. Vegetable and flower crops are often glycophytes and cannot grow in the presence of high levels of salt; a NaCl concentration over 100–200 mM often results in plant death [7]. The aesthetical appearance—and hence the cash value—of these crops [8,9] is strictly linked to the absence of salinity-induced damage, which is one of the principal morphological indicators of salt stress in plants [10]. Nonetheless, for many vegetable plants, more than twenty years after the review on the subject by Shannon and Grive [8], information on the effect of salinity stress is not sufficient. This lack of information is even more apparent for flower and ornamental crops.
The negative effects of salinization on plants include a reduction in growth trends due to the alteration of numerous biochemical and physiological processes [11]. Plant species perceive saline conditions and respond through a complex signaling network generated by ions, osmotic potential changes, and the biosynthesis of plant hormones or reactive oxygen species (ROS) [12]. These signals reach their respective receptors and determine the activation of physiological mechanisms that allow the plant to adapt to stress conditions. In the case of abiotic stresses, such as saline stress, three types of signal transduction have been identified, namely the ion signaling pathway, the osmolyte regulatory pathway, and the gene regulatory pathway [13]. Understanding each tolerance mechanism that occurs at the cellular level is essential for understanding how the entire plant organism reacts, which will contribute to improving the quality and productivity of different crops (Table 1) [3].
Therefore, this review aims to analyze the most recent results reported with respect to the molecular control of salt tolerance in vegetable and flower crops, in particular the osmotic regulation mechanism, salinity-induced proteins, reactive oxygen metabolism, the mechanism of signal transduction, and the development of salinity stress. This will provide useful indications to improve our knowledge of the mechanisms of salt tolerance in these species and assist in the development of future programs for the evaluation and selection of germplasm and genetic improvement.
2. Osmotic Regulation Mechanism
Salinity negatively affects plant physiology through ionic toxicity as well as osmotic and oxidative stress. Osmotic adaptation is essential to sustain cell turgor. Plants react to the osmotic stress caused by high salt levels mainly with osmotic adjustments (Figure 1).
Osmotic regulation is essential to maintain cell turgor and plant metabolic activity and, therefore, plant growth and productivity [45]. Plant under stress biosynthesizes a large number of osmoprotectants, such as proline (Pro), glycine betaine (GB), sugars, and sugar alcohols, facilitating the antioxidant mechanism and ionic homeostasis [46,47]. Pro is an amino acid and osmoprotectant, which acts as an important signaling molecule that accumulates in the cytosol of plants and functions in the stabilization and protection of membrane, protein enzymes, and various proteins. Pro also plays an important role during salinity stress by increasing the production of membrane proteins and ROS scavengers and maintaining cellular solute homeostasis. Many researchers have reported that under salinity, Pro improves water absorption and the antioxidant mechanism. Furthermore, it reduces the accumulation of toxic ions [46,48,49]. The Pro content that a plant accumulates to relieve osmotic stress can, therefore, be used as a physiological indicator of its ability to tolerate stress [50]. This substance exists in plant cells in the free state and has a low molecular weight, high water solubility, and no net charge in the physiological pH range [51].
Soluble sugars include mainly glucose, sucrose, and trehalose. These sugars can stabilize the cell membrane and protoplast [52]. They are osmolytes that stabilize the integrity of the cell membrane, which protects proteins from aggregation and denaturation. Soluble sugars can also be used as a physiological indicator of salt tolerance due to their osmoregulation function [1]. Sugar alcohols, such as inositol, sorbitol, and mannitol, also induce salinity tolerance by regulating cellular osmotic levels. These compounds increase the water potential of cells, allowing plants to uptake more water from the soil (Figure 1). In particular, they mitigate stress by promoting growth, scavenging ROS, maintaining cell turgor, and aiding in the sequestration of Na+ from the cytosol into the vacuole.
Betaines, such as glycine betaine (GB), proline betaine, hydroxyproline betaine, and pipecolate betaine, also act as osmoprotectants [48]. Of these, GB acts as a strong and compatible osmoprotectant in mitigating salinity stress. Extensive research has shown that GB maintains osmotic adaptation by regulating the Na+ to K+ ratio and accumulating in the cell; in particular, this reduces the toxic effects of the ions [53].
Further investigations are necessary to better understand the role of different osmoprotectants and the changes in gene expression in plants that occur as a response to this stress. Different genes are involved at the transcriptional level; it will be useful to discriminate if this gene expression is a direct result of the stress conditions or injury responses.
3. Reactive Oxygen Species Metabolism
Under abiotic stress conditions, photosynthetic activity is reduced, and the excitation energy of light becomes greater than that used or required by photosynthesis, resulting in an accumulation of ROS in the chloroplasts [54]. Abiotic stresses that induce the production of ROS hinder plant growth and lessen crop yield via reduction in chlorophyll contents and photosynthetic efficiency; the degree of this damage depends on the severity, frequency, and duration of the abiotic stress [55,56,57]. In optimal conditions, ROS are neutralized by intracellular antioxidants, whereas under salt stress conditions, the extreme accumulation of ROS produces oxidative stress and strongly disturbs the normal metabolism, causing protein destruction and nucleic acid mutation [58], and the overproduction of ROS in the plant induces lipid peroxidation in the cell membrane. The loss of membrane integrity impairs the physiological and biochemical processes in the plants. Under saline stress conditions, plants activate the production of malondialdehyde (MDA), which is the main product of membrane lipid peroxidation; its content represents the degree of cell membrane damage [1]. An increase in this compound is considered an oxidative stress indicator and is used as a tool to evaluate the tolerance of plants to salt conditions [59]. In salt stress conditions, plants increase the production of ROS, such as H2O2 (hydrogen peroxide), O2− (superoxide), 1O2 (singlet oxygen), and ∙OH (hydroxyl radical). The overproduction of ROS then results in lipid peroxidation, protein degradation, and DNA mutation or damage [60]. To limit the oxidative damage caused by the excessive production of ROS, plants have implemented a complex antioxidant defense system which includes low-molecular-weight antioxidants (ascorbate, reduced glutathione, tocopherol, carotenoids, and flavonoids) and antioxidant enzymes, such as superoxide dismutase (SOD EC 1.15.1.1), peroxidase (POD EC 1.11.1.6), ascorbate peroxidase (APX EC 1.11.1.6), and catalase (CAT EC 1.11.1.6) [61].
The principal ROS-scavenging enzymes in plants include CAT, SOD, and POD. Catalase decomposes hydrogen peroxide into water and oxygen. In the same way, SOD converts superoxide radicals into oxygen and H2O2. POD and some other enzymes are involved in the degradation of H2O2 into innocuous products [62]. Ascorbate breakdown by APX involves the reduction of H2O2 to water [63].
SOD and catalase have been identified as the most effective enzymes in scavenging active oxygen species that cause oxidative stress. An alternative mode of H2O2 destruction is via peroxidases, which are found throughout the cell and have a much higher affinity for H2O2 than catalases [64]. Different studies have shown that salt stress treatment increases SOD activity [65]. Similarly, higher POD activity in the leaves of different plant species under salt stress conditions has also been found [66,67].
The principal site of ROS production under salinity stress conditions is the electron transport chain (ETC) in the chloroplast and mitochondria [68].
Controlling the production and action of ROS allows for a better understanding of the effects of various abiotic stresses on plants (Figure 2).
In various cultivated plants (rice, tomato, citrus, pea, and mustard), several studies have shown that the production of ROS is increased under salt stress conditions, and ROS-mediated membrane damage has been demonstrated as a major cause of cellular toxicity [69,70]. A significant increase in enzyme activities (SOD, POD, and CAT) was observed in the leaves of Asteraceae families of salt tolerant cv Wuxi [71]. Similarly, in sunflower leaves subjected to 200 mM NaCl, increases in enzyme activity and soluble protein content were observed [72]. In maize plants, Kaya et al. [73] observed that prolonged salinity reduced some physiological parameters (leaf relative water content and leaf water potential) and enhanced MDA and H2O2 concentration [73].
The overproduction of GSH and APX has been shown to improve oxidative stress tolerance, resulting in enhanced water stress in wheat [74]. One of the main causes of decreases in crop productivity is the production of ROS during abiotic stresses (salinity, water stress, and high and low temperatures). Hence, the regulation of ROS is a crucial process to avoid unwanted cellular cytotoxicity (DNA damage) and oxidative damage (Figure 2).
The metabolism of ROS under salt stress is complicated and includes mechanisms and interactions with other molecules and components that are yet to be identified. Thus, further studies are necessary to understand the process more clearly. Knowledge of ROS metabolism under salt stress will be helpful in mitigating salt-induced oxidative stress.
4. Mechanism of Signal Transduction and the Development of Salinity Stress
A high concentration of salts in soil or water induces several changes at the molecular level that allow crops to face the stress condition. Salinity is defined as the total amount of dissolved salts. The simplest method for salinity level determination is through electrical conductivity (EC). This index indirectly measures the salt concentration in soil or water. The most dangerous elements are sodium, chlorides, and sulphates. Plants have a very limited need for these elements; thus, they often accumulate in the soil near the roots and increase its salinity. However, high salinity can also be a result of excessive fertilizer application in cropping systems. The continuous application of mineral fertilizers to crops increases the salinity of the soil with time. Sodium (Na+) stress can be increased by irrigation water or infiltration of seawater along the coastal area. Seawater infiltration can reduce the number of crop species that can be grown as well as their productivity [75]. With climate change, the increase in global temperature in most Mediterranean areas will result in higher evapotranspiration rates in crops; this reduction in water availability can increase the salinity levels in soils and irrigation water. Crops can be exposed to salinity early in their life if they are planted in salty soil. In this condition, their germination can be inhibited by the salinity, and their signaling and activation systems can be slowed down [76]. On the contrary, crops can be exposed to a progressive salinity increase during development due to an increase in salts in the irrigation or underground water. In these cases, ion concentrations in the root zone gradually increase; plants can sense this salinity and activate signaling networks related to defense against excessive ion uptake and cellular accumulation.
The excessive uptake of ions can activate diverse protection mechanisms, and different plant species have varying defense strategies. Plants sense salinity through the cells of the roots that are in direct contact with salt ions [77].
Excessive salinity can induce osmotic stress in plants via the activation of salt-associated genes that function to mitigate the negative effects of Na+ accumulation. In this state, plants do not uptake water and reduce the leaf growth and cell elongation [78].
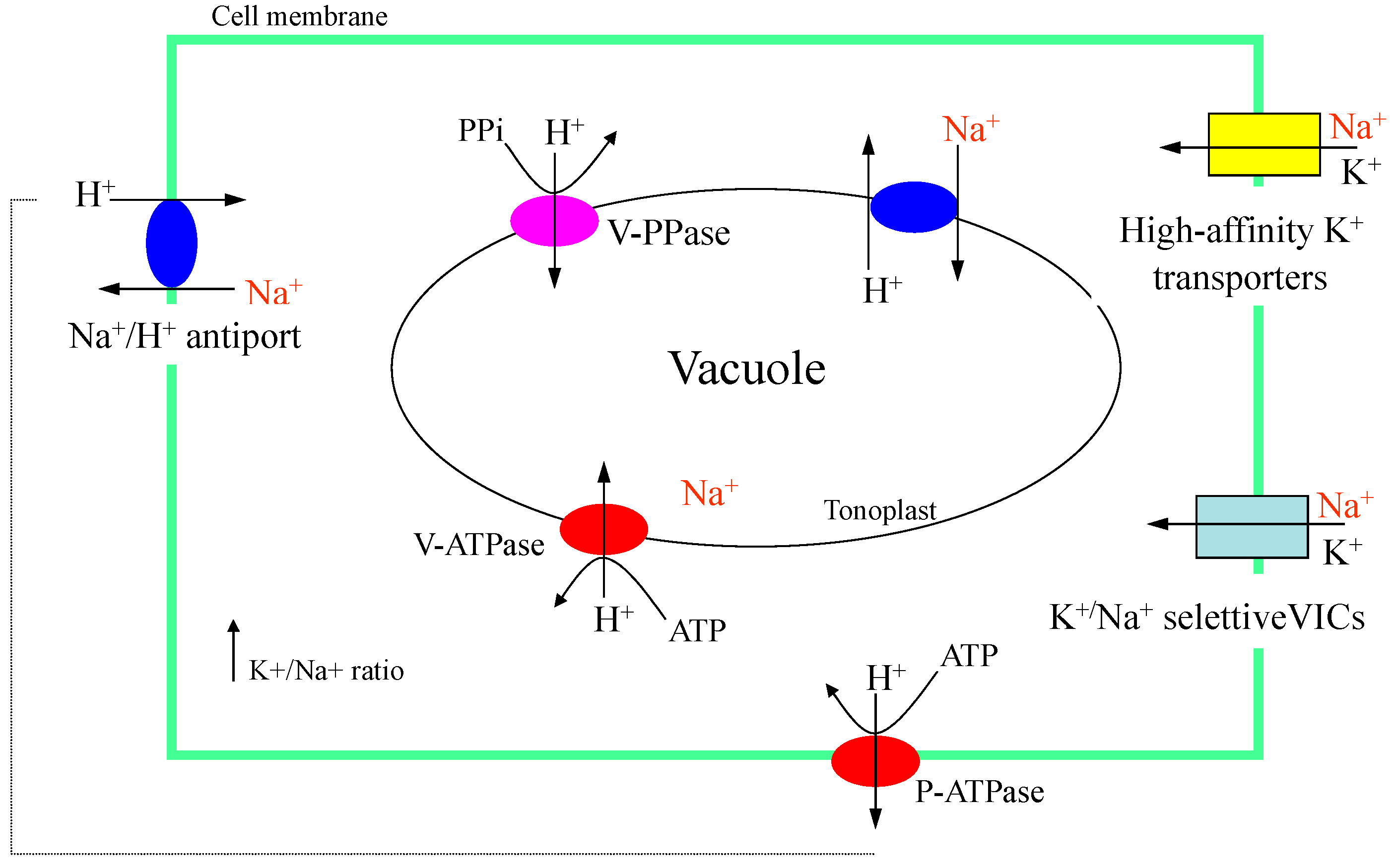
Na+ uptake can occur through ion transporters that have an affinity for monovalent cations, such as high-affinity potassium transporters (Figure 3). The accumulation of Na+ in the cytoplasm can inhibit the activities of enzymes, resulting in negative effects on cell metabolism. Decreasing the Na+ concentration in the cytoplasm can be achieved by storing this ion in the vacuole, a process carried out by Na+/H+ exchangers. If the Na+ concentration is also high in the vacuole, Na+/H+ exchangers can exclude the ion outside the cell (Figure 3). Several Na+/H+ transporter (NHX) genes have been identified in plants. These NHX genes are responsible for Na+/H+ uptake and they belong to the cation–proton antiporters 1 family. The translocation of the Na+ to the vacuole has the function of avoiding the inhibition of enzymatic reactions in the cytosol [79]. The ion homeostasis between Na+ and potassium (K+) in the cytoplasm is essential for maintaining vital physiological and biochemical processes. A low Na+ concentration in the cytoplasm can be achieved by the sequestration of the ion to the vacuole, while in the cytoplasm, K+ and calcium (Ca2+) increase in response to salinity [77].
Crops with high ionic uptake can adapt to salinity and maintain cell turgor by ionic compartmentalization in vacuoles and the accumulation of neutral solutes in the cytoplasm. Crops can avoid high ionic concentrations by direct salt extraction and controlling ionic uptake and translocation in growth zones. Crops can increase the volume of the areal part (succulence) as a dilution strategy [80]. Plants with excessive ionic uptake can experience negative effects due to ion accumulation in the cell wall and water deficits in the cytoplasm. The high accumulation of Na+ can impair metabolism and, in particular, photosynthesis.
Crops can adapt to high salinity, tolerate high ionic concentrations, and maintain turgor by compartmentalizing ions in the vacuoles and accumulating neutral solutes in the cytoplasm. Another strategy to avoid excessive ionic concentrations is the removal of salt from shoots by salt glands or phloem export or by the control of uptake and transport in growing areas.
Crops with reduced ionic uptake can adapt to salinity by avoiding drought stress. This strategy can be achieved by increasing the biosynthesis of organic solutes that maintain tissue turgor. In these crops, the cell wall acquires higher extensibility. Plant turgor can be preserved by increasing water permeability or leaf thickness to reduce transpiration. The use of water under high-salinity conditions is improved and crops show higher water use efficiency (WUE).
However, in crops that are sensitive to salinity stress, even low ion uptake can be damaged by high salinity as a consequence of a reduction in growth due to reduced turgor. Moreover, a reduction in CO2 fixation due to high stomatal resistance can limit plant development.
At the molecular level, specific genes have been shown to be associated with salinity and Na+ transport (Figure 4). Salt signaling is based on the plant’s sensitivity to salt. In Arabidopsis thaliana L., Salt Overly Sensitive (SOS) genes were isolated in mutants that were highly sensitive to salinity [81]. These mutants contributed to our understanding of how plants can defend themselves against excess salinity. SOS genes encode for specific proteins associated with Na+ transport. The SOS1 gene is a Na+/H+ antiporter integrated in the plasmalemma that transports Na+ from the cytoplasm to the apoplast. The SOS2 and SOS3 proteins are located in the cytoplasm and associated with SOS1 function. Under high salt concentration, Ca2+ regulates the SOS3–SOS2 protein complexes, which block Na+ transporters to avoid its accumulation in the cell [82]. Functional studies have revealed that SOS1 is the key regulator of Na+ tolerance, while SOS2 and SOS3 play supporting roles. These Na+ transporters function in equilibrating the ion concentration in the cell [77]. Cytosolic Ca2+ is involved in the regulation of SOSs and tolerance to salinity stress conditions [83]. Ca2+ acts on SOS2 and SOS3, which block the uptake of Na+ [84]. Ca2+ seems to be regulated by the reduced hyperosmolality-induced (Ca2+) increase1 (OSCA1) gene. A functional analysis carried out by studying osca1 mutants revealed that osmotic Ca2+ adjustment in the root and guard cells was disrupted, as was water flux regulation. Another type of channel for monovalent and bivalent cations is cyclic nucleotide-gated channels (CNGCs). In plants, 20 CNGCs have been found, and, in particular, CNGC18 is associated with Ca2+ activation [84].
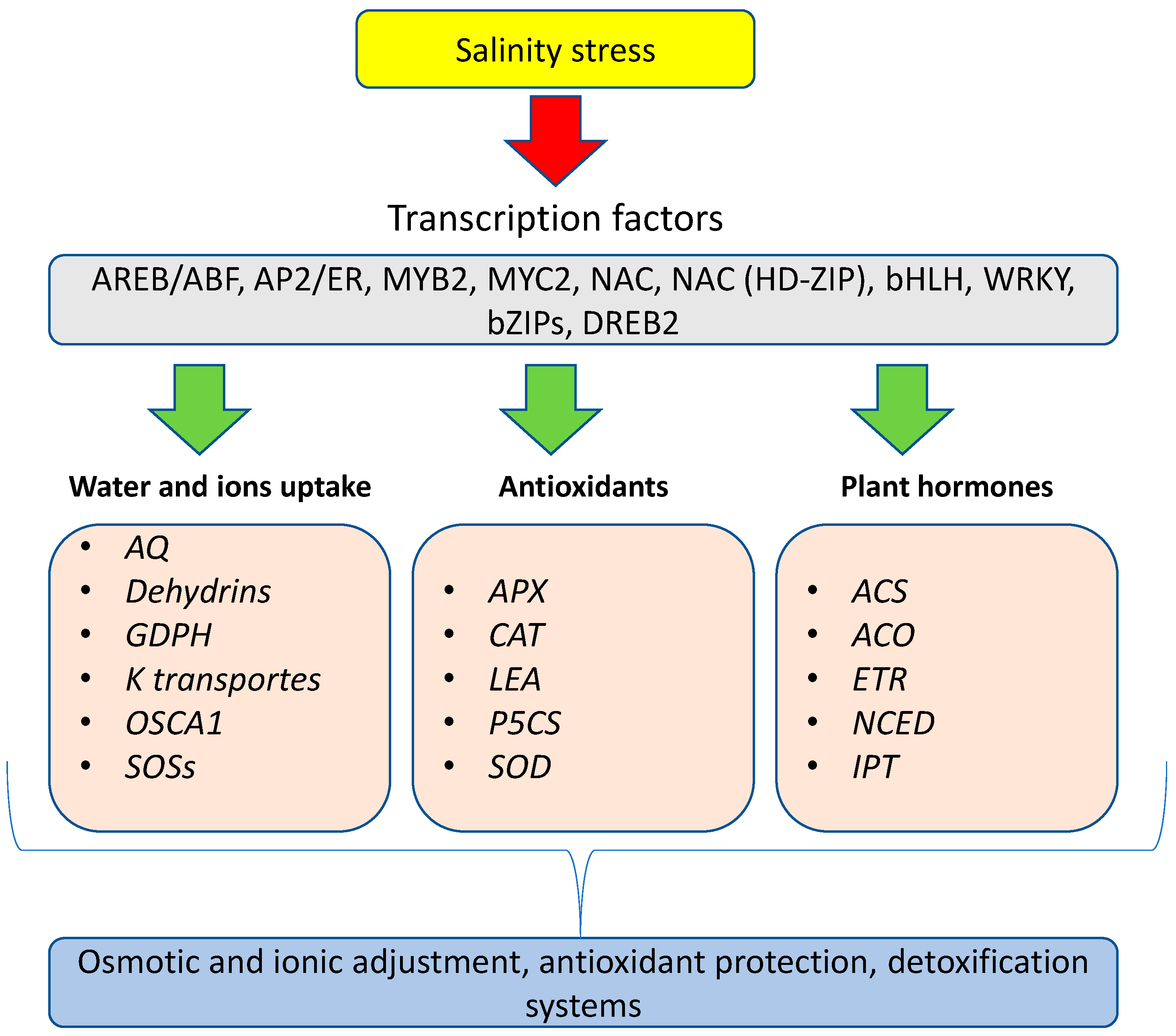
Transcriptional studies performed under salinity conditions have reported that many transcription factors are activated by salinity, including MYB2, MYC2, NAC, AREB/ABF, NAC (HD-ZIP), and DREB2 (Figure 4). These transcriptional changes regulate the expression of diverse genes, which leads to adaptation to salinity conditions. These genes are responsible for the activation of secondary genes that are associated with plant adaptation. From transcription to translation, the response can be very fast—from a few minutes or hours to days.
Transcriptional studies have elucidated the cluster of genes that is activated by salinity stress. In wheat, it was shown that the transcription factors induced by salinity included MYB, NAC, bHLH, WRKY, bZIPs, and AP2/ER (Figure 4). These genes regulate the expression of water channels (aquaporins (AQ)), late-embryogenesis-abundant (LEA) proteins, dehydrins, proline synthesis enzyme (P5CS), and proline oxidase (proline degradation enzyme). The activation of these genes increases the tolerance of wheat to salinity stress [85]. In addition, SOS1, K+ transporters, glycerol-3-phosphate dehydrogenase (GPDH), Calcium ATPase, and ABC transporters were also found to be activated under salt stress. An analog transcriptomic study performed on Solanum lycopersicum L. and S. chilense (Dunal) Reiche at the vegetative stage showed differentially expressed salt stress genes. The most important cluster of genes was associated with antioxidant defense enzymatic mechanisms, including SOD, CAT, and APX [86]. In a transcriptional study performed to investigate the effect of borage extract under salinity, the transcription factors DtRD29A and DtHB7 were highly expressed after 9 h under salinity conditions [87]. From a practical point of view, tolerance to salinity could be transiently activated by the application of calcium nitrate fertilizers. A similar effect can be obtained with potassium nitrate [88]. However, these two cations can act in different ways; Ca2+ plays a role in signaling and inhibiting Na+ accumulation, while K+ can reduce the Na+ concentration by competing for the same transport channels. There are also genes activated by salinity that are involved in membrane biosynthesis or repair, such as phosphatidylinositol-specific phospholipase C (PI-PLC). This gene activation is shared with different biosynthetic pathways.
Further investigations are required to identify the key genes that can induce tolerance to salinity stress in crops. Some salinity-associated genes have been found, and new undiscovered genes could be the master regulators of salinity tolerance in crops. The identification of these few genes could lead to the improved selection of new tolerant crops.
5. Salinity-Induced Proteins, Amino Acids, and Enzymes
Plants under abiotic stresses can accumulate a wide number of proteins that are shared among diverse stresses. Under salinity stress, plants can increase the accumulation of proteins as a storage form of nitrogen, which can be reused when the stress conditions are over. In particular, osmotins (26 kDa protein) are accumulated under salinity in diverse plant species [89]. Plants exposed to salinity also increase the accumulation of amino acids, the most important of which are proline, glutamine, and asparagine. Some of these are also associated with senescence, with the same function of translocating and storing nitrogen in the seeds, trunk, or branches.
Proline is one indicator of crop tolerance to abiotic stresses, in particular drought and salinity [90,91,92]. Proline plays a beneficial role in crops exposed to stress conditions. It is an excellent osmolyte, and its function can be summarized by its three major roles during abiotic stresses: it is a metal chelator, antioxidant compound, and a signaling molecule. The biosynthesis of proline from glutamic acid is carried out through the activities of the following enzymes: ∆’-pyrroline-5-carboxylate synthetase (P5CS, EC:2.7.2.11), which leads to ∆’-pyrroline-5-carboxylate (P5C) formation and the conversion to proline by ∆’-pyrroline-5-carboxylate reductase (P5CR, EC 1.5.1.2). An increase in biosynthetic proline enzymes in salinity conditions can help crops adapt to and tolerate stressful conditions.
In one study, overexpressing the Δ1-pyrroline-5-carboxylate synthetase gene in crops, such as soybean, induced an increase in tolerance to salinity, demonstrating the potential protection function of proline against salinity [93]. The endogenous proline concentration increased in plants exposed to salinity. Catharanthus roseus (L.) G. Don. ornamental plants under 80 mM NaCl stress also showed an increase in proline concentration [94]. Similar results were found in Silvinia natans (L.) All., which accumulated proline under high salinity stress [95]. On the basis of the observation of an increase in proline concentrations under salt stress, several studies have been performed to verify whether salinity tolerance can be achieved through exogenous applications for protecting crops. Exogenous treatments of proline can lead to a reduction in lipid peroxidation and programmed cell death under salinity stress [96]. In melon plants (Cucumis melo L.), the application of proline and potassium nitrate improved their tolerance to salinity (150 mM NaCl), avoiding yield reduction [97].
Salinity stress can activate several stress-related pathways, such as ethylene biosynthesis. Ethylene is a gaseous plant hormone that is biosynthesized via three enzymatic reaction steps. The first enzyme, S-adenosyl-methionine synthetase (S-AdoMet), converts the amino acid methionine into S-AdoMet, which is transformed into 1-Aminocyclopropane-1-carboxylate (ACC) by 1-aminocyclopropane-1-carboxylate synthase (ACC synthase, EC 4.4.1.14). ACC is directly converted into ethylene by an enzyme that oxidizes the ACC to form carbon dioxide and ethylene. This step is catalyzed by 1-aminocyclopropane-1-carboxylate oxidase (ACC oxidase, EC 1.14.17.4). Under salinity stress, the ethylene signaling network is activated with the involvement of its receptors, such as ethylene receptor 1 (ETR1); ethylene signaling messengers, such as constitutive triple response 1 (CTR1); and transcription factors, such as ethylene insensitive (EINs), ethylene insensitive lines (EILs), or ethylene response factors (ERFs). Plants under salinity stress increase ethylene production, and the amount of ethylene produced is associated with salinity stress intensity. Experiments with Arabidopsis etr1-1 mutants exhibiting reduced ethylene production showed a higher sensitivity to salinity. Therefore, treatments that alleviate salinity stress or increase salinity tolerance can be measured through the ethylene production.
Salinity tolerance or sensitivity in plants is also mediated by water balance, as previously described (Figure 5). At the biochemical level, it has been found that an increase in abscisic acid (ABA) in plants can help to counteract salinity stress. In tomato crops, an increase in ABA in rootstocks improved tomato tolerance. ABA is a plant hormone that is produced by carotenoid degradation (also known as the C40 indirect pathway). The key enzyme in this pathway is 9-cis-epoxycarotenoid dioxygenase (NCED, EC 1.13.11.51), which catalyzes the rate-limiting step in ABA biosynthesis. Transgenic tomato plants overexpressing the SlNCED gene exhibit a high ABA concentration, which helps the plants overcome transient stressful salinity conditions [98,99]. Several studies and review papers have suggested an important role of ABA in regulating crop tolerance to salinity [100]. Scientific evidence suggests that it can be used as a strategy in the selection of vegetable species or cultivars with higher ABA content or inducible salt-related ABA accumulation for growing vegetables under high-salinity conditions.
Another hormone involved in plant development is gibberellic acid (GA). This hormone has various different functions, but the most important is cell elongation. This effect can increase salinity tolerance in plants. One study performed on maize (Zea mays L.) demonstrated that treatment with GA3 (100 mg L−1) increased root and shoot biomass and PRO concentration and preserved membrane integrity in plants exposed to 100 mM NaCl [101]. Treatment with GA did not increase the activities of detoxification enzymes, indicating that the treated plants had lower stress. GA could act through the dilution effect on biomass development. Most investigations should be performed on the role of GA in protecting plants against salinity stress. Cytokinins are responsible for cell division and are leaf-yellowing inhibitors. In salinity stressed plants, the exogenous application of cytokinins can have direct or indirect effects. In marjoram (Majorana hortensis Moench) and spearmint (Mentha spicata L.) plants, the foliar application of cytokinins—in particular diphenylurea or kinetin—was able to counteract the negative effects of salinity on both plant growth and essential oil accumulation [102]. In rice (Oryza sativa L.), mutants that overaccumulated cytokinins showed a higher yield than wild-type stressed plants [103]. These results could be due to the effect of cytokinins on delaying senescence and increasing the concentrations of chlorophyll and the precursor of Pro. Brassinosteroids (BR) are involved in the plant hormones network (Figure 5) through their interaction with ethylene biosynthesis and can stimulate crop tolerance [104].
Metabolic profiles change under salinity stress, and further investigations should be performed to understand how biosynthesis or accumulation can be achieved using agronomic tools, such as plant growth regulators or biostimulants. Moreover, combined studies considering both transcriptomic and metabolomic data could help in the identification of the genes and metabolites that can be enhanced in new crops through specific breeding programs with the support of assisted selection of molecular markers.
6. Conclusions and Future Prospective
Salinity is strictly associated with irrigation and has a dramatic effect on crop productivity; hence, it plays an important role in food crises. This problem is particularly evident in vegetable and ornamental crops, which are irrigation crops that are required to be absent of salt damage due to the relevance of visual aspects in the quality of products. Moreover, many of them are glycophytic plants that are sensitive to salinity. The currently published data and scientific evidence suggest that crops have a predominant strategy that involves one or more pathways for salinity stress tolerance. Crops tolerant to salinity can have higher concentrations of antioxidant compounds or highly active enzymatic systems that regulate the biosynthesis of metabolites associated with osmotic adjustment.
A considerable number of questions and aspects need to be further investigated. Further information about the mechanisms of action of ROS, the role of osmoprotectants, and the mechanisms of signal transduction and salinity-induced substances are necessary. Omic approaches to these questions will be useful for improving our understanding of tolerance mechanisms and better orientating breeding activities by using new biotechnological solutions. This information can be exploited for selecting tolerant wild species that can be used as rootstock for grafting, as a source for tolerant traits, or for direct use in agricultural systems.
Author Contributions
Conceptualization, A.F. and D.R.; methodology, S.T., D.R and A.F.; formal analysis, S.T., D.R. and A.F.; writing—original draft preparation, S.T., D.R. and A.F.; writing—review and editing, S.T., D.R. and A.F.; supervision, D.R. and A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Malvankar, M.R.; Karle, S.B.; Kumar, K. Reactive nitrogen species: Paradigms of cellular signaling and regulation of salt stress in plants. Environ. Exp. Bot. 2019, 161, 86–97. [Google Scholar] [CrossRef]
- Chen, J.-T.; Aroca, R.; Romano, D. Molecular aspects of plant salinity stress and tolerance. Int. J. Mol. Sci 2021, 22, 4918. [Google Scholar] [CrossRef]
- Agarwal, P.; Agarwal, P.K.; Gohil, D. Transcription Factor-Based Genetic Engineering for Salinity Tolerance in Crops. In Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S., Suprasanna, P., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2018; Volume 1, pp. 185–211. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Baniasadi, F.; Saffari, V.R.; Moud, A.A.M. Physiological and growth responses of Calendula officinalis L. plants to the interaction effects of polyamines and salt stress. Sci. Hortic. 2018, 234, 312–317. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Cassaniti, C.; Romano, D.; Hop, M.E.C.M.; Flowers, T.J. Growing floricultural crops with brackish water. Environ. Exp. Bot. 2013, 92, 165–175. [Google Scholar] [CrossRef]
- Bañón, S.; Fernández, J.A.; Ochoa, J.; Sánchez-Blanco, M.J. Paclobutrazol as an aid to reduce some effects of salt stress in oleander seedlings. Eur. J. Hortic. Sci. 2005, 70, 43–49. [Google Scholar]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Stevens, R.M.; Pech, J.M.; Grigson, G.J. A short non-saline sprinkling increases the tuber weights of saline sprinkler irrigated potatoes. Agronomy 2017, 7, 4. [Google Scholar] [CrossRef]
- Nxele, X.; Klein, A.; Ndimba, B.K. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. Afr. J. Bot. 2017, 1, 261–266. [Google Scholar] [CrossRef]
- Chen, L.; Meng, Y.; Jiang, D.; Yang, F.; Zhou, Y. Physio-biochemical responses of three Aquilegia species seedlings to salt stress. Agronomy 2022, 12, 2841. [Google Scholar] [CrossRef]
- Fernandez-Garcia, N.; Hernandez, M.; Casado-Vela, J.; Bru, R.; Elortza, F.; Hedden, P.; Olmos, E. Changes to the proteome and targeted metabolites of xylem sap in Brassica oleracea in response to salt stress. Plant Cell Environ. 2011, 34, 821–836. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative stress and antioxidant defense mechanisms in plants under salt stress. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 191–205. [Google Scholar]
- Jia, J.; Liang, Y.; Gou, T.; Hu, Y.; Zhu, Y.; Huo, H.; Guo, J.; Gong, H. The expression response of plasma membrane aquaporins to salt stress in tomato plants. Environ. Exp. Bot. 2020, 178, 104190. [Google Scholar] [CrossRef]
- Lacramioara, O.; Grigore, M.N.; Vochita, G. Impact of saline stress on growth and biochemical indices of Calendula officinalis seedlings. Rom. Biotechnol. Lett. 2015, 20, 11007. [Google Scholar]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of sweet pepper with stress tolerance-inducing compounds alleviates salinity stress oxidative damage by mediating the physio-biochemical activities and antioxidant systems. Agronomy 2019, 10, 26. [Google Scholar] [CrossRef]
- Sarwar, M.; Anjum, S.; Alam, M.W.; Ali, Q.; Ayyub, C.M.; Haider, M.S.; Ashraf, M.I.; Mahboob, W. Triacontanol regulates morphological traits and enzymatic activities of salinity affected hot pepper plants. Sci. Rep. 2022, 12, 3736. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Ahmad, I.; Basit, A.; Shehata, W.F.; Hassan, U.; Shah, S.T.; Haleema, B.; Jalal, A.; Amin, R.; Khalid, M.A.; et al. Ascorbic acid enhances growth and yield of sweet peppers (Capsicum annum) by mitigating salinity stress. Gesunde Pflanzen 2022, 74, 423–433. [Google Scholar] [CrossRef]
- Gengmao, Z.; Yu, H.; Xing, S.; Shihui, L.; Quanmei, S.; Changhai, W. Salinity stress increases secondary metabolites and enzyme activity in safflower. Ind. Crops Prod. 2015, 64, 175–181. [Google Scholar] [CrossRef]
- Hassan, F.A.S.; Ali, E.; Gaber, A.; Fetouh, M.I.; Mazrou, R. Chitosan nanoparticles effectively combat salinity stress by enhancing antioxidant activity and alkaloid biosynthesis in Catharanthus roseus (L.) G. Don. Plant Physiol. Biochem. 2021, 162, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Vanlal, R.; Anand, P.; Kumar, G.; Tiwari, A.K. Effect of saline stress on growth and biochemical indices of chrysanthemum (Chrysanthemum morifolium) germplasm. Indian J. Agric. Sci. 2019, 89, 41–45. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, C.; Chen, C.; Zou, L.; Yin, S.; Liu, S.; Yuan, J.; Wu, N.; Liu, X. Comparative transcriptome analysis reveals variations of bioactive constituents in Lonicera japonica flowers under salt stress. Plant Physiol. Biochem. 2022, 173, 87–96. [Google Scholar] [CrossRef]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Ghashghaie, J. Genotypic differences in physiological and biochemical responses to salinity stress in melon (Cucumis melo L.) plants: Prospects for selection of salt tolerant landraces. Plant Physiol. Biochem. 2017, 119, 294–311. [Google Scholar] [CrossRef]
- Sarwar, M.; Anjum, S.; Ali, Q.; Alam, M.W.; Haider, M.S.; Mehboob, W. Triacontanol modulates salt stress tolerance in cucumber by altering the physiological and biochemical status of plant cells. Sci. Rep. 2021, 11, 24504. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Hegarat, E.; Jiménez-Lao, M.; Lao, M.T. Effects of exogenous application of osmotic adjustment substances on growth, pigment concentration, and physiological parameters of Dracaena sanderiana Sander under different levels of salinity. Agronomy 2020, 10, 125. [Google Scholar] [CrossRef]
- Samadi, S.; Habibi, G.; Vaziri, A. Exogenous trehalose alleviates the inhibitory effects of salt stress in strawberry plants. Acta Physiol. Plant. 2019, 41, 112. [Google Scholar] [CrossRef]
- Ahsan, M.; Zulfiqar, H.; Farooq, M.A.; Ali, S.; Tufail, A.; Kanwal, S.; Shaheen, M.R.; Sajid, M.; Gul, H.; Jamal, A.; et al. Strigolactone (GR24) Application positively regulates photosynthetic attributes, stress-related metabolites and antioxidant enzymatic activities of ornamental sunflower (Helianthus annuus cv. Vincent’s Choice) under salinity stress. Agriculture 2023, 13, 50. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Pestana, M.; Correia, P.J.; Lao, M.T. Lavandula multifida response to salinity: Growth, nutrient uptake, and physiological changes. J. Plant. Nutr. Soil Sci. 2017, 180, 96–104. [Google Scholar] [CrossRef]
- Shahzadi, A.K.; Bano, H.; Ogbaga, C.C.; Ayyaz, A.; Parveen, R.; Zafar, Z.U.; Ashraf, M. Coordinated impact of ion exclusion, antioxidants and photosynthetic potential on salt tolerance of ridge gourd [Luffa acutangula (L.) Roxb.]. Plant Physiol. Biochem. 2021, 167, 517–528. [Google Scholar] [CrossRef]
- Loko, B.; Montcho, K.D.H.; Agbossékpé, F.; Mensah, A.C.G.; Assogba Komlan, F.; Lutts, S.; Gandonou, C.B. Response of African Basil (Ocimum gratissimum L.) to salt stress under tropical conditions in the Republic of Benin: Growth, ions and organic solutes accumulation. Int. J. Soil Sci. 2022, 34, 47–60. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of salt stress on growth, physiological parameters, and ionic concentration of water dropwort (Oenanthe javanica) cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.; Shahbaz, M.; Maqsood, M.F.; Farhat, F.; Zulfiqar, U.; Javed, T.; Fraz Ali, M.; Alhomrani, M.; Alamri, A.S. Proline-Induced Modifications in Morpho-Physiological, Biochemical and Yield Attributes of Pea (Pisum sativum L.) Cultivars under Salt Stress. Sustainability 2022, 14, 13579. [Google Scholar] [CrossRef]
- Shahzad, S.; Ali, S.; Ahmad, R.; Ercisli, S.; Anjum, M.A. Foliar application of silicon enhances growth, flower yield, quality and postharvest life of tuberose (Polianthes tuberosa L.) under saline conditions by improving antioxidant defense mechanism. Silicon 2022, 14, 1511–1518. [Google Scholar] [CrossRef]
- Borsai, O.; Hassan, M.A.; Negrușier, C.; Raigón, M.D.; Boscaiu, M.; Sestraș, R.E.; Vicente, O. Responses to salt stress in Portulaca: Insight into its tolerance mechanisms. Plants 2020, 9, 1660. [Google Scholar] [CrossRef]
- Omidi, M.; Khandan-Mirkohi, A.; Kafi, M.; Zamani, Z.; Ajdanian, L.; Babaei, M. Biochemical and molecular responses of Rosa damascena Mill. cv. Kashan to salicylic acid under salinity stress. BMC Plant Biol. 2022, 22, 373. [Google Scholar] [CrossRef]
- Ahmad, P.; Abass Ahanger, M.; Nasser Alyemeni, M.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef]
- Mittova, V.; Theodoulou, F.L.; Kiddle, G.; Gómez, L.; Volokita, M.; Tal, M.; Foyer, C.H.; Guy, M. Coordinate induction of glutathione biosynthesis and glutathione-metabolizing enzymes is correlated with salt tolerance in tomato. FEBS Lett. 2003, 554, 417–421. [Google Scholar] [CrossRef]
- Ikuyinminu, E.; Goñi, O.; O’Connell, S. Enhancing irrigation salinity stress tolerance and increasing yield in tomato using a precision engineered protein hydrolysate and Ascophyllum nodosum-derived biostimulant. Agronomy 2022, 12, 809. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Naz, S.; Altaf, M.M.; Qadir, A.; Anwar, M.; Shakoor, A.; Hayat, F. Exogenous melatonin enhances salt stress tolerance in tomato seedlings. Biol. Plant. 2020, 64, 604–615. [Google Scholar] [CrossRef]
- Yin, Z.; Lu, J.; Meng, S.; Liu, Y.; Mostafa, I.; Qi, M.; Li, T. Exogenous melatonin improves salt tolerance in tomato by regulating photosynthetic electron flux and the ascorbate–glutathione cycle. J. Plant Interact. 2019, 14, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Parvin, K.; Nahar, K.; Hasanuzzaman, M.; Bhuyan, M.B.; Mohsin, S.M.; Fujita, M. Exogenous vanillic acid enhances salt tolerance of tomato: Insight into plant antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2020, 150, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.E.; Hsiao, T.C.; Silk, W.K. Growth of the maize primary root at low water potentials: II. Role of growth and deposition of hexose and potassium in osmotic adjustment. Plant Physiol. 1990, 93, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Dai, W. Plant response to salinity stress. In Stress Physiology of Woody Plants; Day, W., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 155–173. [Google Scholar]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, K.; Goto, Y.; Asatsuma, S.; Koizumi, M.; Mitsui, T.; Matsuoka, K. A mobile secretory vesicle cluster involved in mass transport from the Golgi to the plant cell exterior. Plant Cell 2009, 21, 1212–1229. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; de Almeida Viégas, R.; da Rocha, I.M.A.; de Oliveira Monteiro Moreira, A.C.; de Azevedo Moreira, R.; Oliveira, J.T.A. Proline accumulation and glutamine synthetase activity are increased by salt-induced proteolysis in cashew leaves. J. Plant Physiol. 2003, 160, 115–123. [Google Scholar] [CrossRef]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.H. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Toscano, S.; Trivellini, A.; Cocetta, G.; Bulgari, R.; Francini, A.; Romano, D.; Ferrante, A. Effect of preharvest abiotic stresses on the accumulation of bioactive compounds in horticultural produce. Front. Plant Sci. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naing, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant. 2021, 172, 1711–1723. [Google Scholar] [CrossRef]
- Ai, T.N.; Naing, A.H.; Yun, B.W.; Lim, S.H.; Kim, C.K. Overexpression of RsMYB1 enhances anthocyanin accumulation and heavy metal stress tolerance in transgenic petunia. Front. Plant Sci. 2018, 9, 1388. [Google Scholar] [CrossRef]
- Naing, A.H.; Park, K.I.; Ai, T.N.; Chung, M.Y.; Han, J.S.; Kang, Y.W.; Lim, K.B.; Kim, C.K. Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Czégény, G.; Wu, M.; Dér, A.; Eriksson, L.A.; Strid, Å.; Hideg, É. Hydrogen peroxide contributes to the ultraviolet-B (280–315 nm) induced oxidative stress of plant leaves through multiple pathways. FEBS Lett. 2014, 588, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, B.; Yaşar, F.; Ozpay, T.; Tuerkoezue, D.; Terziodlu, O.; Tamkoc, A. Variations in response to salt stress among field pea genotypes (Pisum sativum sp. arvense L.). J. Anim. Veter. Adv. 2008, 7, 907–910. Available online: https://medwelljournals.com/abstract/?doi=javaa.2008.907.910 (accessed on 2 December 2022).
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Signal. 2006, 8, 1757–1764. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Baek, K.H. Physiological and biochemical perspectives of non-salt tolerant plants during bacterial interaction against soil salinity. Plant Physiol. Biochem. 2017, 116, 116–126. [Google Scholar] [CrossRef]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Kahrizi, S.; Sedighi, M.; Sofalian, O. Effect of salt stress on proline and activity of antioxidant enzymes in ten durum wheat cultivars. Ann. Biol. Res. 2012, 3, 3870–3874. [Google Scholar]
- Chookhampaeng, S. The effect of salt stress on growth, chlorophyll content proline content and antioxidative enzymes of pepper (Capsicum annuum L.) seedling. Eur. J. Res. 2011, 49, 103–109. [Google Scholar]
- Sevengor, S.; Yasar, F.; Kusvuran, S.; Ellialtioglu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jeleel, C.A.; Azooz, M.M.; Nabi, G. Generation of ROS and non-enzymatic antioxidants during abiotic stress in plants. Bot. Res. Intern. 2009, 2, 11–20. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Sharma, S. Antioxidative defence system, lipid peroxidation, proline metabolizing enzymes and biochemical activity in two genotypes of Morus alba L. subjected to NaCl stress. Russ. J. Plant. Physiol. 2010, 57, 509–517. [Google Scholar] [CrossRef]
- Xue, Y.F.; Liu, Z.P. Antioxidant enzymes and physiological characteristics in two Jerusalem artichoke cultivars under salt stress. Russ. J. Plant Physiol. 2008, 55, 776–781. [Google Scholar] [CrossRef]
- Ebrahimian, E.; Bybordi, A. Exogenous silicium and zinc increase antioxidant enzyme activity and alleviate salt stress in leaves of sunflower. J. Food Agric. Environ. 2011, 9, 422–427. [Google Scholar]
- Kaya, C.; Akram, N.A.; Ashraf, M.; Sonmez, O. Exogenous application of humic acid mitigates salinity stress in maize (Zea mays L.) plants by improving some key physico-biochemical attributes. Cereal Res. Commun. 2018, 46, 67–78. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Saxena, D.C. Role of antioxidant systems in wheat genotypes tolerance to water stress. Biol. Plant. 1998, 41, 387–394. [Google Scholar] [CrossRef]
- Mariani, L.; Ferrante, A. Agronomic management for enhancing plant tolerance to abiotic stresses—Drought, salinity, hypoxia, and lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Shelake, R.M.; Kadam, U.S.; Kumar, R.; Pramanik, D.; Singh, A.K.; Kim, J.Y. Engineering drought and salinity tolerance traits in crops through CRISPR-mediated genome editing: Targets, tools, challenges, and perspectives. Plant Commun. 2022, 3, 100417. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Sci. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Role of vacuolar membrane transport systems in plant salinity tolerance. J. Plant Growth Regul. 2022, 1–38. [Google Scholar] [CrossRef]
- Grigore, M.N.; Toma, C. Histo-anatomical strategies of Chenopodiaceae halophytes: Adaptive, ecological and evolutionary implications. WSEAS Trans. Biol. Biomed. 2007, 4, 204–218. [Google Scholar]
- Qiu, Q.S.; Guo, Y.; Quintero, F.J.; Pardo, J.M.; Schumaker, K.S.; Zhu, J.K. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J. Biol. Chem. 2004, 279, 207–215. [Google Scholar] [CrossRef]
- Quintero, F.J.; Ohta, M.; Shi, H.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef]
- Ghosh, S.; Bheri, M.; Bisht, D.; Pandey, G.K. Calcium signaling and transport machinery: Potential for development of stress tolerance in plants. Curr. Plant Biol. 2022, 29, 100235. [Google Scholar] [CrossRef]
- Dong, Q.; Wallrad, L.; Almutairi, B.O.; Kudla, J. Ca2+ signaling in plant responses to abiotic stresses. J. Integr. Plant Biol. 2022, 64, 287–300. [Google Scholar] [CrossRef]
- Amirbakhtiar, N.; Ismaili, A.; Ghaffari, M.R.; Nazarian Firouzabadi, F.; Shobbar, Z.S. Transcriptome response of roots to salt stress in a salinity-tolerant bread wheat cultivar. PLoS ONE 2019, 14, e0213305. [Google Scholar] [CrossRef]
- Kashyap, S.P.; Kumari, N.; Mishra, P.; Moharana, D.P.; Aamir, M.; Singh, B.; Prasanna, H.C. Transcriptional regulation-mediating ROS homeostasis and physio-biochemical changes in wild tomato (Solanum chilense) and cultivated tomato (Solanum lycopersicum) under high salinity. Saudi J. Biol. Sci. 2020, 27, 1999–2009. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Trivellini, A.; Ferrante, A. Transcriptional regulation in rocket leaves as affected by salinity. Plants 2019, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ak, B.E.; Higgs, D. Response of salt-stressed strawberry plants to supplementary calcium nitrate and/or potassium nitrate. J. Plant Nutr. 2003, 26, 543–560. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Shannon, M.C.; Rhoades, J.D.; Draper, J.H.; Scardaci, S.C.; Spyres, M.D. Assessment of salt tolerance in rice cultivars in response to salinity problems in California. Crop Sci. 1998, 38, 394–398. [Google Scholar] [CrossRef]
- Arteaga, S.; Yabor, L.; Díez, M.J.; Prohens, J.; Boscaiu, M.; Vicente, O. The use of proline in screening for tolerance to drought and salinity in common bean (Phaseolus vulgaris L.) genotypes. Agronomy 2020, 10, 817. [Google Scholar] [CrossRef]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity responses and tolerance mechanisms in underground vegetable crops: An integrative review. Planta 2022, 255, 68. [Google Scholar] [CrossRef]
- Zhang, G.C.; Zhu, W.L.; Gai, J.Y.; Zhu, Y.L.; Yang, L.F. Enhanced salt tolerance of transgenic vegetable soybeans resulting from overexpression of a novel Δ1-pyrroline-5-carboxylate synthetase gene from Solanum torvum Swartz. Hortic. Environ. Biotechnol. 2015, 56, 94–104. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Panneerselvam, R. Calcium chloride effects on salinity-induced oxidative stress, proline metabolism and indole alkaloid accumulation in Catharanthus roseus. C. R. Biol. 2007, 330, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Jampeetong, A.; Brix, H. Effects of NaCl salinity on growth, morphology, photosynthesis and proline accumulation of Salvinia natans. Aquat. Bot. 2009, 91, 181–186. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Tuna, A.L.; Ashraf, M.; Altunlu, H. Improved salt tolerance of melon (Cucumis melo L.) by the addition of proline and potassium nitrate. Environ. Exp. Bot. 2007, 60, 397–403. [Google Scholar] [CrossRef]
- Martínez-Andújar, C.; Martínez-Pérez, A.; Ferrández-Ayela, A.; Albacete, A.; Martínez-Melgarejo, P.A.; Dodd, I.C.; Thompson, A.J.; Pérez-Pérez, J.M.; Pérez-Alfocea, F. Impact of overexpression of 9-cis-epoxycarotenoid dioxygenase on growth and gene expression under salinity stress. Plant Sci. 2020, 295, 110268. [Google Scholar] [CrossRef]
- Martínez-Andújar, C.; Martínez-Pérez, A.; Albacete, A.; Martínez-Melgarejo, P.A.; Dodd, I.C.; Thompson, A.J.; Mohareb, F.; Estelles-Lopez, L.; Kevei, Z.; Ferrández-Ayela, A.; et al. Overproduction of ABA in rootstocks alleviates salinity stress in tomato shoots. Plant Cell Environ. 2021, 44, 2966–2986. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Dikilitas, M.; Higgs, D. The combined effects of gibberellic acid and salinity on some antioxidant enzyme activities, plant growth parameters and nutritional status in maize plants. Environ. Exp. Bot. 2008, 62, 1–9. [Google Scholar] [CrossRef]
- El-Keltawi, N.E.; Croteau, R. Salinity depression of growth and essential oil formation in spearmint and marjoram and its reversal by foliar applied cytokinin. Phytochemistry 1987, 26, 1333–1334. [Google Scholar] [CrossRef]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflorescence meristem-specific cytokinin oxidase–OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Jiroutova, P.; Oklestkova, J.; Strnad, M. Crosstalk between brassinosteroids and ethylene during plant growth and under abiotic stress conditions. Int. J. Mol. Sci. 2018, 19, 3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
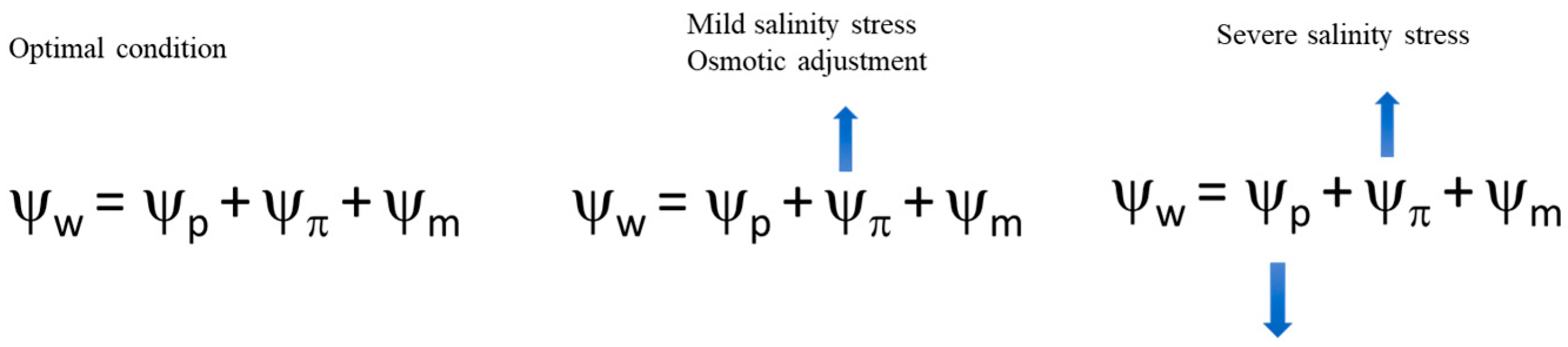
Figure 1.
Water balance of crops under normal conditions and under progressive stress conditions (from left to right). The increase in salinity stress induces the accumulation of osmolytes that increase the ability of root cells to uptake water (the cell osmotic potential becomes more negative). The progression of salinity and its severity cannot be counteracted with only osmolytes; the plant does not uptake water and loses its turgor. ΨW = water potential; Ψp = pressure potential; Ψπ = gravimetric potential; and Ψm = matric adsorption force.
Figure 1.
Water balance of crops under normal conditions and under progressive stress conditions (from left to right). The increase in salinity stress induces the accumulation of osmolytes that increase the ability of root cells to uptake water (the cell osmotic potential becomes more negative). The progression of salinity and its severity cannot be counteracted with only osmolytes; the plant does not uptake water and loses its turgor. ΨW = water potential; Ψp = pressure potential; Ψπ = gravimetric potential; and Ψm = matric adsorption force.

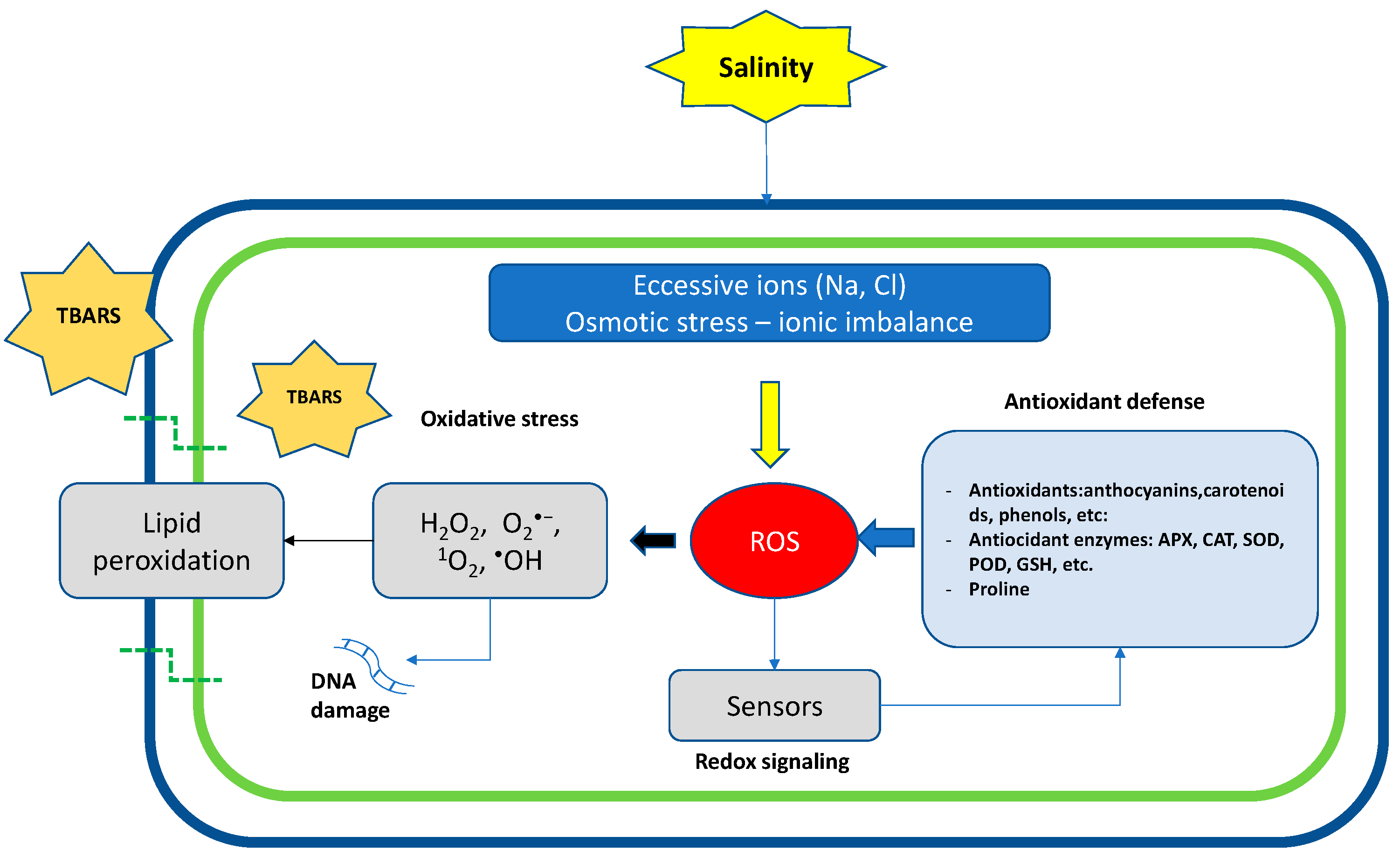
Figure 2.
At the cellular level, salinity induces osmotic stress, ion imbalance, ROS accumulation, DNA damage, and lipid peroxidation. Crops activate redox signaling that leads to the induction of antioxidant defenses, such as the biosynthesis of antioxidants or the activation of antioxidant enzymatic systems.
Figure 2.
At the cellular level, salinity induces osmotic stress, ion imbalance, ROS accumulation, DNA damage, and lipid peroxidation. Crops activate redox signaling that leads to the induction of antioxidant defenses, such as the biosynthesis of antioxidants or the activation of antioxidant enzymatic systems.

Figure 3.
Possible Na+ uptake or extrusion in plant cells, including accumulation in the vacuole, for the protection of biochemical processes at the cytoplasm level.
Figure 3.
Possible Na+ uptake or extrusion in plant cells, including accumulation in the vacuole, for the protection of biochemical processes at the cytoplasm level.

Figure 4.
The first crop salinity responses at the molecular level involve the expression of transcription factors that activate the regulatory genes responsible for the activation of secondary genes. These clusters of genes are involved in water and ion uptake, the production of antioxidant enzymes, and plant hormones biosynthesis.
Figure 4.
The first crop salinity responses at the molecular level involve the expression of transcription factors that activate the regulatory genes responsible for the activation of secondary genes. These clusters of genes are involved in water and ion uptake, the production of antioxidant enzymes, and plant hormones biosynthesis.

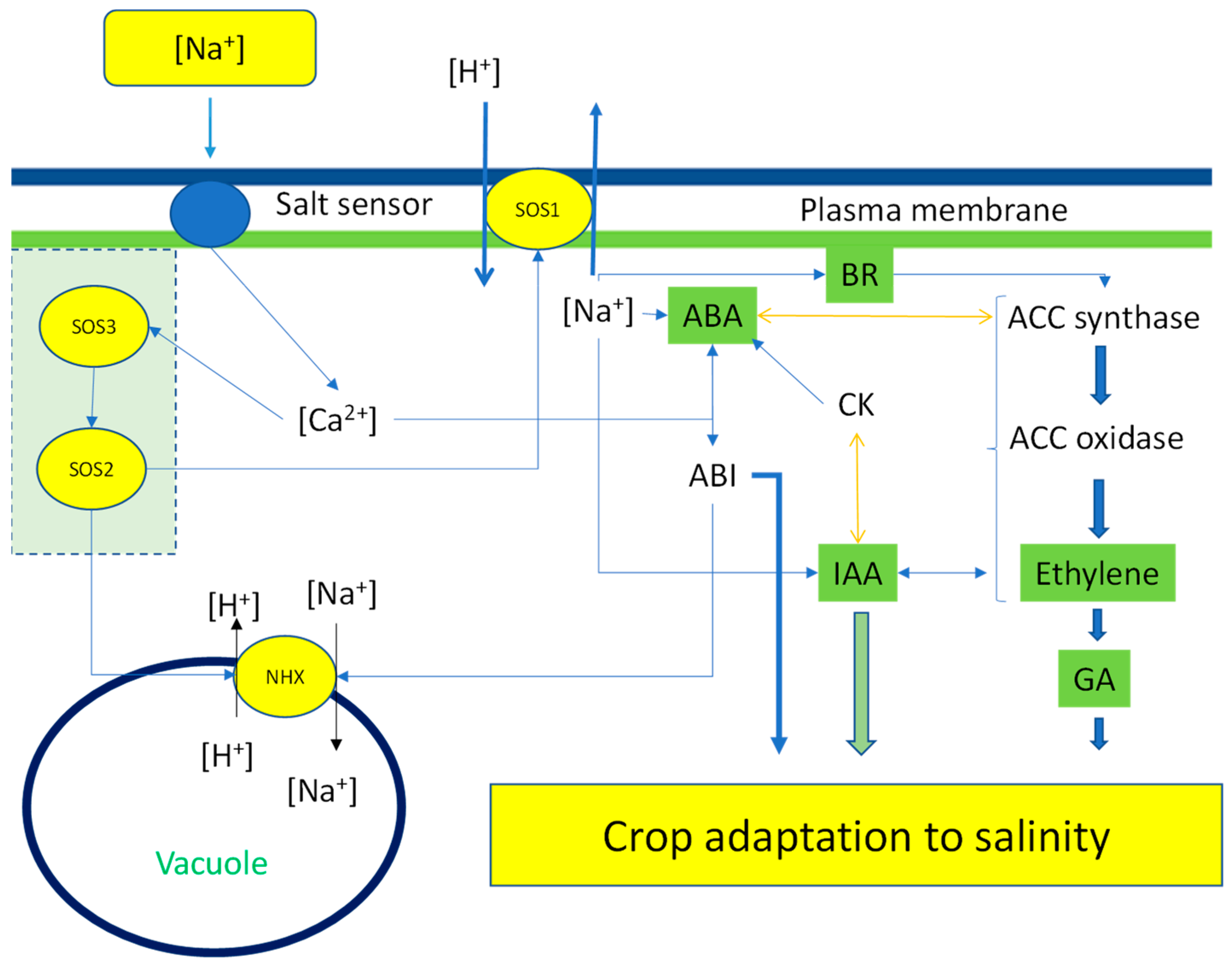
Figure 5.
Under salinity stress due to high sodium concentration, the cell activates several protective systems that lead to adaptation. The sodium sensor promotes an increase in calcium in the cytoplasm that regulates the SOS1 and SOS3. These complexes are responsible for sodium accumulation in the vacuole and extrusion of sodium in the apoplast out of the cell. Sodium and calcium regulate the plant hormone network and interact with each other to allow for crop adaptation to high-salinity conditions.
Figure 5.
Under salinity stress due to high sodium concentration, the cell activates several protective systems that lead to adaptation. The sodium sensor promotes an increase in calcium in the cytoplasm that regulates the SOS1 and SOS3. These complexes are responsible for sodium accumulation in the vacuole and extrusion of sodium in the apoplast out of the cell. Sodium and calcium regulate the plant hormone network and interact with each other to allow for crop adaptation to high-salinity conditions.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main molecular effects of salt stress in different vegetable and ornamental crops.
| Plant Species and Cultivar | Salt Stress Level | Effect of Salt Stress on Plants | References |
|---|---|---|---|
| Aquilegia oxysepala Trautv. & C.A.Mey., A. parviflora Ledeb., and A. viridiflora Pall. | 5.0 ± 0.2 dS m−1 and 10.0 ± 0.2 dS m−1 | Increase in MDA and proline; increased activity of SOD (5.0 dS m−1); A. parviflora POD increase; and A. viridiflora POD increase (10.0 dS m−1) | [14] |
| Brassica oleracea L. | 80 mM | POX increase | [15] |
| Brassica oleracea L. | 50, 100, 150, and 200 mM NaCl | Increase in CAT and POX activity, increase in proline | [16] |
| Brassica rapa L. subsp. rapa ‘Qiamagu’ | 50, 100, 150, and 200 mM NaCl | Increased activity of SOD (50, 100, 150, and 200 mM), POD (100, 150, and 200 mM), CAT (150 mM), and APX (200 mM); increase in MDA (100, 150, 200 mM) | [17] |
| Brassica rapa L. subsp. rapa ‘Wenzhoupancai’ | 50, 100, 150, and 200 mM NaCl | Increased activity in SOD and APX (200 mM), POD and CAT (100, 150, and 200 mM); increase in MDA (100, 150 mM) | [17] |
| Calendula officinalis L. | 50–100 mM NaCl, 36 d | Increase in proline | [18] |
| Calendula officinalis L. | 1, 5, and 9 dSm−1 | Increase in MDA in leaves and roots, increase in proline in leaves (9 dS m−1), and increase in CAT activity; decrease in POD activity | [6] |
| Capsicum annuum L. | 2000 and 4000 ppm NaCl | Increased activity of CAT and POX; increase in proline | [19] |
| Capsicum annuum L. | 75 mM NaCl | Increase in SOD, POX, and CAT activity; increase in MDA | [20] |
| Capsicum annuum L. ‘Candy Apple’ | 35, 70, and 105 mM | Increase in APX and PPO activity | [21] |
| Carthamus tinctorius L. | 50, 100, and 150 mM NaCl | Increased activity of CAT (50 mM), SOD (100 mM), and POD (50, 100, and 150 mM) | [22] |
| Catharanthus roseus (L.) G. Don | 150 mM NaCl | MDA increase in vegetative and flowering stage; increase in CAT, GPX, and GR activity in vegetative and flowering stage | [23] |
| Chrysanthemum L. cvs. (‘Garden Beauty’, ‘Shanti’, ‘Red Stone’, ‘Basanti’, ‘Yellow Reflex’, ‘Ravi Kiran’, ‘Anmol’, ‘Mother Teresa’, ‘Sweta Singar’, and ‘Jaya’) | 150 mM NaCl | Increase in proline | [24] |
| Cornus florida L. and C. hongkongensis subsp. elegans (Fang & Hsieh) Q.Y.Xiang | 0.2%, 0.3%, 0.4%, and 0.45% salt solution | Increase in MDA, SOD activity (0.2%, 0.3%, 0.4%, and 0.45% salt solution) and proline (0.3%, 0.4%, and 0.45% salt solution) | [25] |
| Cucumis melo L. | 30, 60, and 90 mM NaCl | Increase in proline, MDA, APX, CAT, SOD, and POD | [26] |
| Cucumis sativus L. (‘Green long’, ‘Marketmore’, ‘Summer green’, and ‘20252’) | NaCl 50 mM L−1 | Increase in proline | [27] |
| Dracaena braunii Engl. | 2.0 and 7.5 dS m−1 | Increase in proline | [28] |
| Fragaria ×ananassa (Duchesne ex Weston) Duchesne ex Rozier ‘Gaviota’ | 50 mM | Increase in MDA | [29] |
| Helianthus annuus L. (ornamental sunflower) | 150 mM NaCl | Increased activity of CAT and POD; increase of proline | [30] |
| Lavandula multifida L. | 10–200 mM NaCl, 60 d | Increase in soluble sugars concentration | [31] |
| Luffa acutangula Roxb. | 75 and 150 mM | Increase in proline | [32] |
| Ocimum gratissimum L. (African basil) | 30, 60, 90, 120 mM | Increase in proline in leaves (120 mM) and root (90 and 120 mM) | [33] |
| Oenanthe javanica DC. ‘V11E0022’ and ‘V11E0135’ | 50 and 100 mM NaCl | Increase in leaves and roots of MDA and proline | [34] |
| Pisum sativum L. ‘L-888’ and ‘Round’ | 150 mM | ‘L-888’ increase in CAT activity; ‘Round’ increase in proline and decrease in SOD activity | [35] |
| Polianthes tuberosa L. | 50 and 100 mM NaCl | Increase in SOD, POD (100 mM), GR, and APX; increase in proline (100 mM) | [36] |
| Portulaca oleracea subsp. oleracea L., P. grandiflora Hook., P. halimoides L., and P. oleracea ‘Toucan Scarlet Shades’ | 100, 200, and 400 mM | Increase in proline in leaves and roots | [37] |
| Rosa damascena Mill. ‘Kashan’ | 4, 8, and 12 dS m−1 | Increase in MDA (8 mM); increase in proline (8, 12 mM); and increase of CAT activity | [38] |
| Solanum lycopersicum L. | 50 μM S-nitroso-N-acetyl penicillamine (SNAP) 200 | Increased activity of APX, glutathione reductase (GR), peroxidase and rise in proline content | [39] |
| Solanum lycopersicum L. | 100 mM | Mn-SOD, MDHAR, and GR decrease | [40] |
| Solanum lycopersicum L. | 300 mM NaCl | Increase in proline | [41] |
| Solanum lycopersicum L. | 150 mM NaCl | Increase in MDA content; increase in SOD and CAT activity | [42] |
| Solanum lycopersicum L. ‘Liaoyuanduoli’ | 150 mM NaCl | Increase in MDA and SOD, APX, GPX, GR, MDHAR, and DHAR activity | [43] |
| Solanum lycopersicum L. ‘Pusa Ruby’ | 150 mM NaCl | Increase in proline and MDA; increase activity of APX, MDHAR, DHAR, GR, SOD, CAT, GPX, and GST | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Toscano, S.; Romano, D.; Ferrante, A. Molecular Responses of Vegetable, Ornamental Crops, and Model Plants to Salinity Stress. Int. J. Mol. Sci. 2023, 24, 3190. https://doi.org/10.3390/ijms24043190
AMA Style
Toscano S, Romano D, Ferrante A. Molecular Responses of Vegetable, Ornamental Crops, and Model Plants to Salinity Stress. International Journal of Molecular Sciences. 2023; 24(4):3190. https://doi.org/10.3390/ijms24043190
Chicago/Turabian StyleToscano, Stefania, Daniela Romano, and Antonio Ferrante. 2023. "Molecular Responses of Vegetable, Ornamental Crops, and Model Plants to Salinity Stress" International Journal of Molecular Sciences 24, no. 4: 3190. https://doi.org/10.3390/ijms24043190
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.