
Advanced Glycation End-Products and Diabetic Neuropathy of the Retina

1
Department of Ophthalmology and Visual Science, Chiba University Graduate School of Medicine, Inohana 1-8-1, Chuo-ku, Chiba 260-8670, Japan
2
Department of Ophthalmology, School of Medicine, International University of Health and Welfare, 4-3 Kozunomori, Narita 286-8686, Japan
Int. J. Mol. Sci. 2023, 24(3), 2927; https://doi.org/10.3390/ijms24032927
Submission received: 17 December 2022
/
Revised: 29 January 2023
/
Accepted: 31 January 2023
/
Published: 2 February 2023
(This article belongs to the Special Issue Current Insights on Neuroprotection)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Diabetic retinopathy is a tissue-specific neurovascular impairment of the retina in patients with both type 1 and type 2 diabetes. Several pathological factors are involved in the progressive impairment of the interdependence between cells that consist of the neurovascular units (NVUs). The advanced glycation end-products (AGEs) are one of the major pathological factors that cause the impairments of neurovascular coupling in diabetic retinopathy. Although the exact mechanisms for the toxicities of the AGEs in diabetic retinopathy have not been definitively determined, the AGE-receptor of the AGE (RAGE) axis, production of reactive oxygen species, inflammatory reactions, and the activation of the cell death pathways are associated with the impairment of the NVUs in diabetic retinopathy. More specifically, neuronal cell death is an irreversible change that is directly associated with vision reduction in diabetic patients. Thus, neuroprotective therapies must be established for diabetic retinopathy. The AGEs are one of the therapeutic targets to examine to ameliorate the pathological changes in the NVUs in diabetic retinopathy. This review focuses on the basic and pathological findings of AGE-induced neurovascular abnormalities and the potential therapeutic approaches, including the use of anti-glycated drugs to protect the AGE-induced impairments of the NVUs in diabetic retinopathy.
1. Introduction
Diabetes is a metabolic disease characterized by chronic hyperglycemia, and the prevalence of diabetic patients has increased to approximately 600 million and will reach 700 million worldwide by 2045 [1]. Diabetic retinopathy is a major complication in diabetic patients, and it is defined as a tissue-specific neurovascular impairment of the interdependence between multiple cells, which consists of the neurovascular units (NVUs) in both type 1 and type 2 diabetes [2]. Twenty years after the onset of diabetes, all patients with type 1 and 60% of patients with type 2 diabetes will have progressed to diabetic retinopathy [3]. As a result, 95 million diabetic patients have diabetic retinopathy in the world [4], and 10% of the patients with diabetes have progressed to the vision-threatening stages of diabetic retinopathy. This includes clinically significant diabetic macular edema and proliferative diabetic retinopathy [5].
The NVUs in the retina is composed of the vascular cells (endothelial cells and pericytes), the glial cells (Müller cells, astrocytes, and microglia), and the neuronal cells (ganglion cells, amacrine cells, bipolar cells, and horizontal cells) [6,7,8,9]. The photoreceptor cells are mainly nourished by nutrients that pass through the choroidal membrane and retinal pigment epithelium. Thus, the interdependence between photoreceptors and the NVUs may be weaker than other neurons in the retina. The interdependence of the cells constituting the NVUs maintains retinal homeostasis and function under healthy conditions.
Chronic hyperglycemia is a trigger for developing an impairment of the interdependence of cells which consists of the NVUs, because chronic hyperglycemia causes an increase in several stress factors, including advanced glycation end-products (AGEs), inflammation, oxidative stress, activation of the polyol pathway and protein kinase C (PKC). The increase in these stress factors leads to microvascular impairments and glial dysfunction, which then results in increasing the expression of vascular endothelial growth factor (VEGF) and inflammatory cytokines in the eye. This is followed by a breakdown of the blood–retinal barrier [6,7,8,9]. During the course of these changes, there is also excitotoxicity and a decrease in neurotrophic factors resulting in neuronal cell death and axon degeneration, which leads directly to a reduction in vision [6,7,8,9].
Among these stress factors, oxidative stress causes differences in post-transcriptional RNA editing events and gene expression, which may be related to pathological changes in diabetic retinopathy, including retinal degeneration and neovascularization [10,11,12]. The main targets of oxidative stress are glucose, amino acids, and lipids, which result in generating reactive carbonyl compounds (RCOs) [13]. These increasing RCOs, which cause carbonyl stresses, facilitate the synthesis of AGEs under diabetic conditions [13].
The AGEs are pathological factors involved in the onset and progression of diabetic retinopathy and may be one of the therapeutic targets to block to ameliorate the impairment of the NVUs in diabetic retinopathy [14]. This review focuses on the basic and pathological findings of AGEs and potential therapeutic approaches, including the identification of anti-glycated drugs to protect the AGE-induced neurovascular impairments.
2. AGE Production and Accumulation
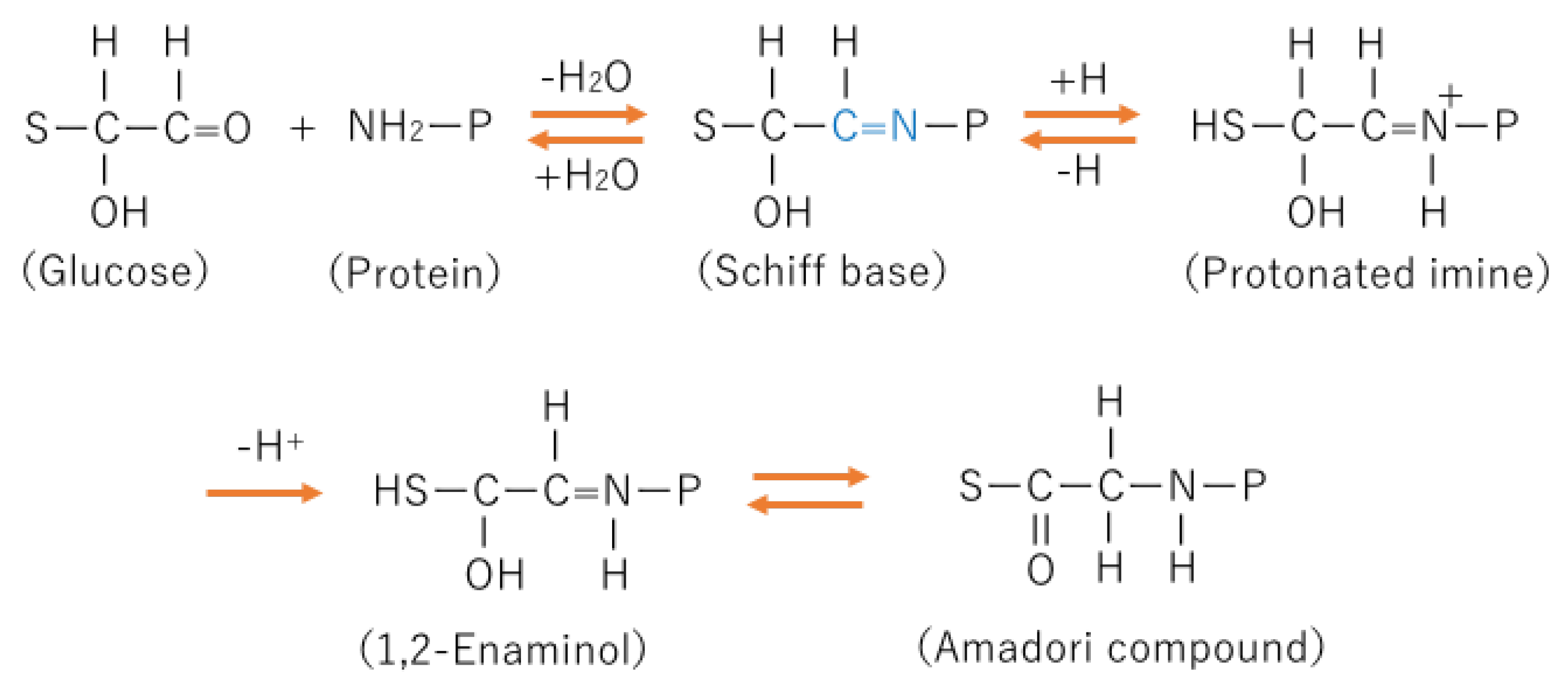
The process for the formation of AGE in the body is complicated and still not completely determined, but the early stage of AGE formation is known to be the Maillard reaction. The Maillard reaction is a nonenzymic reaction (Figure 1).
The intermediate product of the process of AGE formation is an Amadori compound. Amadori compounds are relatively stable and can be precursors of AGEs [15]. The process from a Shiff base to an Amadori compound is thought to be irreversible. Glycohemoglobin A1c (HbA1c) is one of Amadori compounds, which is a clinical marker of glucose control before 1–2 months in diabetic patients.
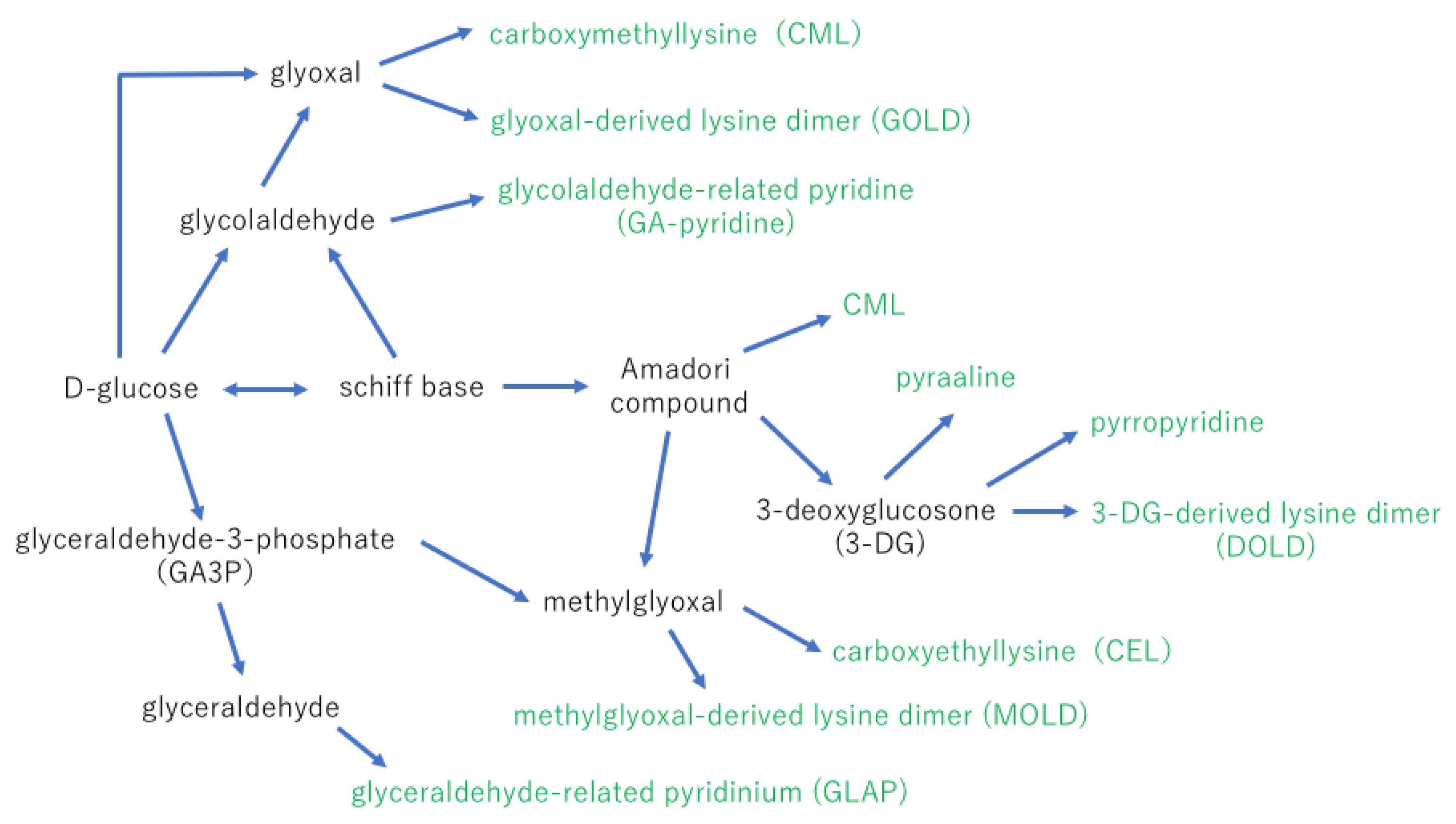
Amadori products are sources of the α-dicarbonyl compounds via metal ions catalyzed autoxidation of glucose or the nonoxidative hydrolysis and fragmentation. 3-deoxy-D-erythro-hexos-2-ulose (3-deoxyglucosone; 3-DG) is a 10,000 times stronger reactive compound for AGE formation than glucose. 3-DG is known to be synthesized from fructose or fructose-3-phosphate via the polyol pathway [16].
The final step of AGE formation is very complex, but a possible process is displayed in Figure 2. Other AGEs that have been identified in the body are carboxymethyl arginine (CMA) [17], imidazolone [18], formyl threosyl pyrrole (FTP) [19], argpyrimidine [20], pentosidine [21], and crossline [22].
The sources of AGEs, including chronic hyperglycemia, are not only endogenous but also exogenous, including foods and tobacco. For example, the smoke of tobacco contains glyoxal and methylglyoxal, and their presence results in the accumulation of AGEs in the body [26]. After intake of AGEs from foods, 10–30% of AGEs are absorbed, and one-third of the AGEs intake is excreted. Thus, 6–7% of AGEs in foods are expected to accumulate in the body. A recent review summarizes the high- and low-AGE food products. High AGE levels of foods are foods rich in protein and fat, baked and grilled food, frying products, and animal products. On the other hand, foods low in AGE are low-fat products, high-carbohydrates products, raw products, and products cooked at low temperatures [26]. Therefore, eating low-level AGE foods may be one method of preventing the progression of diabetic conditions.
The polyol pathway is one of the main sources of AGE formation. The polyol pathway is activated under chronic hyperglycemic conditions (Figure 3). Briefly, under high glucose conditions, excess glucose taken in by retinal cells is metabolized into sorbitol by aldose reductase, and sorbitol is metabolized into fructose by sorbitol dehydrogenase [27,28]. The fructose is converted into fructose-3-phosphate and 3-DG. Because 3-DG is a 10,000 times stronger reactive compound for AGE production than glucose, AGE formation is accelerated by increasing 3-DG by activating the polyol pathway (Figure 3) [28]. During the activation of the polyol pathway, the level of an antioxidant molecule, nicotinamide adenine dinucleotide phosphate (NADPH), is decreased because of its excessive consumption. In addition, nicotinamide adenine dinucleotide (NADH) is increased, which results in PKC activation. These changes contribute to the increased production of reactive oxygen species (ROS) [27,28]. The excess ROS production causes inflammation and cell death in diabetic retinopathy [29]. At the same time, sorbitol is a hyperosmotic factor that increases osmotic stress. The increase in sorbitol production may be due to Müller cell swelling in early diabetic retinopathy (Figure 3) [30]. Oxidative stress induces the alterations of several transcription factors, such as the activator protein-1, p53, and nuclear factor-kappa B (NF-κB) [31]. However, the alteration of gene expression induced by oxidative stress is beyond the scope of this review.
3. Pathophysiology of AGEs
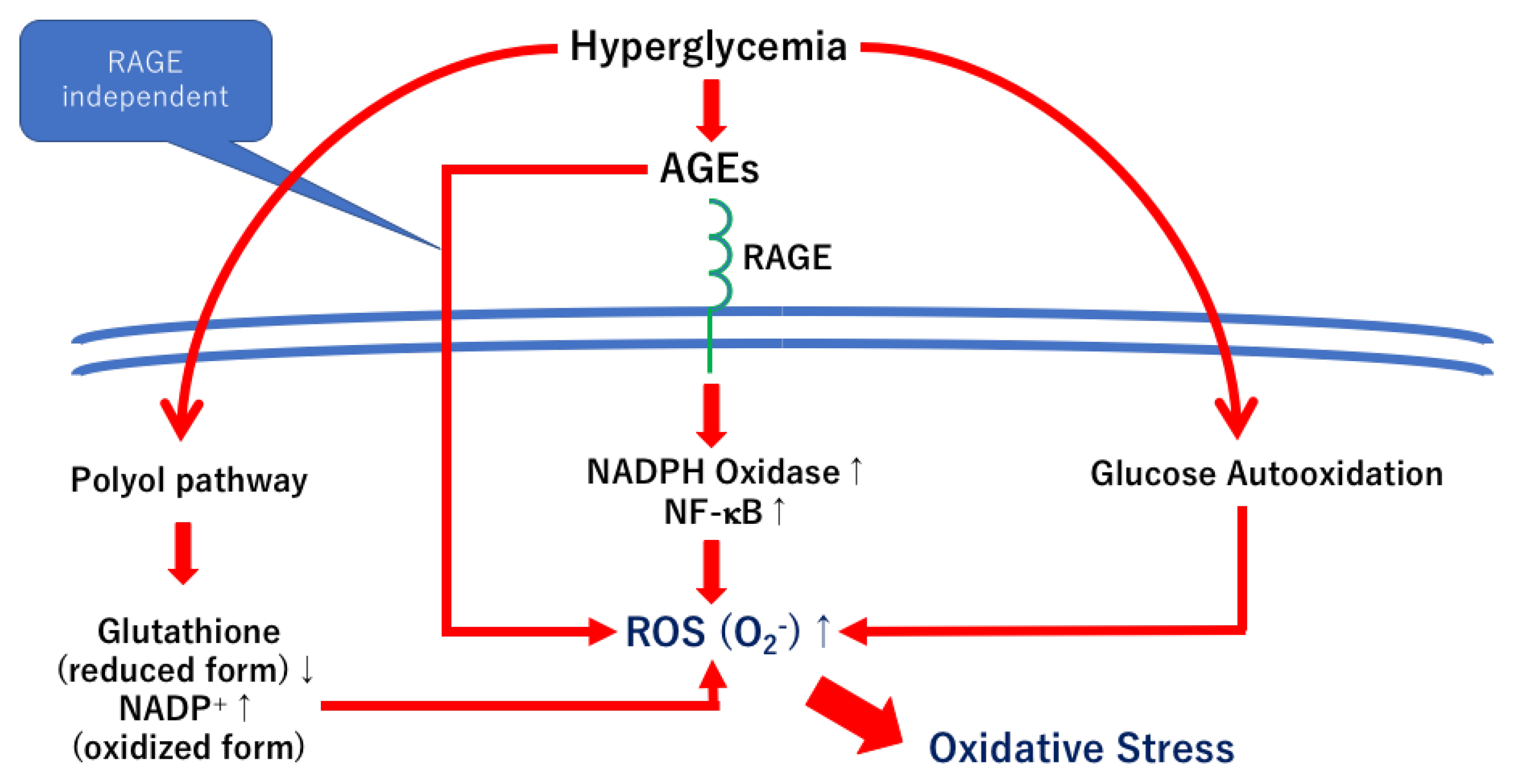
An increase in the synthesis of AGEs leads to biochemical damage to the retinal tissue. The biochemical damages are induced by both the receptors of AGE (RAGE)-dependent and RAGE-independent mechanisms [9]. Under physiological conditions, RAGE is expressed in many tissues and cells, including neurons, glial cells, smooth muscle cells, fibroblasts, monocytes, and endothelial cells [32,33,34]. Not only AGEs but also other ligands such as S100, amphoterin, macrophage-1 antigen, and amyloid-β can bind to the RAGEs, which results in pathophysiological events, including inflammation, ROS generation, and cell death [35].
Although not all cellular mechanisms are activated under the AGE-RAGE axis [36], the AGE/RAGE interaction is thought to activate several cellular pathways and to be associated with the pathophysiology of diabetic retinopathy [9]. For example, the NADPH- NF-κB axis, the RAS-extracellular signal-regulated kinase 1/2 (ERK1/2) axis, and the Ras-mitogen-activated protein kinase (MAPK) axis are involved in inflammatory reactions [36,37,38]. The Ras-c-Jun N terminal kinase (JNK) axis is related to apoptosis.
An increase in the synthesis of the AGEs leads to cross-link formations with proteins, which result in reducing the protein turnover and energy production, activation of endoplasmic reticulum (ER) stress, and macrophage activation [9,39,40]. In addition, the Amadori compounds can become the source of ROS. These cellular events are involved in the AGE-RAGE independent mechanisms (Figure 4).
Ying et al. used the non-invasive skin autofluorescence method [41] to examine the association between the AGE level in the skin and the stages of diabetic retinopathy [42]. They concluded that the AGE level in the skin was associated with the prevalence and severity of diabetic retinopathy and that the AGE level in the skin may be a more suitable indicator than HbA1c for chronic hyperglycemic controls [42]. Similarly, Zhang et al. suggest that the AGE level in the skin is significantly associated with the DR stage in type 2 diabetes mellitus [43]. Takayanagi et al. examined the level of skin autofluorescence to assess the AGE level in the body, and they reported that it was higher in diabetic patients than in non-diabetic patients. They concluded that the higher AGE scores were independently associated with the progression of diabetic retinopathy [44]. In addition, higher AGE scores were risk factors for proliferative diabetic retinopathy [44]. They also concluded that the AGE scores in the skin could be obtained non-invasively, and it was a reliable marker of patients developing sight-threatening diabetic retinopathy [44].
AGE accumulation was found in the endothelial cells of the retina [45], which resulted in capillaries occlusion through increased levels of intracellular cell adhesion molecules followed by culminating retinal ischemic injury [46]. Furthermore, the AGEs in pericytes facilitate pericyte apoptosis by the activation of caspase-3 due to a reduction in the Bcl/Bax ratio [47]. Pericyte loss can trigger microvascular abnormalities, including endothelial cell proliferation and dysfunction in diabetic retinopathy [9,48]. A recent study showed that the higher AGEs score in the skin was a risk factor for diabetic retinopathy, but the score of the carotenoids, major antioxidants, was significantly and negatively correlated with the AGE score, which reflected the vegetable uptake score [49]. Oxidative stress is associated with the pathogenesis of AGEs in diabetic retinas (Figure 4), and carotenoids are plant-derived antioxidants. Shalini et al. suggested that decreased plasma levels and lower dietary intake of carotenoids were found in patients with diabetic retinopathy compared to those without diabetic retinopathy [50].
4. AGEs and Retinal Neuronal Abnormalities
One of the best-known AGEs in the body is CML [23]. However, the results of earlier studies indicated that CML is not strongly associated with the severity of diabetic complications [51,52]. Thus, non-CML AGEs are more important for the pathogenesis of diabetic complications. On the other hand, glycolaldehyde- and glyceraldehyde-derived AGEs induce pericyte loss and increased expression of VEGF, and they promote endothelial cell proliferation. These changes result in the progression of diabetic retinopathy [53,54]. Furthermore, glyceraldehyde-derived AGEs are more toxic than glucose-derived AGEs, and they induce neuronal cell death in cortical neurons [55,56]. However, no studies have demonstrated the toxic effects of glycolaldehyde- and glyceraldehyde-derived AGEs on retinal neurons. Thus, we have investigated the toxic effects of aldehyde-derived AGEs on neuronal cell death and regeneration of retinal ganglion cells in retinal cultures [57,58]. Three-dimensional collagen gel culture systems were helpful for examining the neurotoxic, neuroprotective, and regenerative effects of factors or reagents of interest [59,60,61,62,63,64]. First, we incubated 10 μg/mL of the standard glucose-derived AGEs, glycolaldehyde-derived AGEs, and glyceraldehyde-derived AGEs in serum-free media. In diabetic patients, between 1 and 120 μg/mL of AGEs (approximately 4–480 μg/mL of glycated bovine serum albumin) are circulating in the body [65]. Previously, 100 μg/mL and 250 μg/mL glucose-derived AGEs were shown to have neurotoxic effects [66]. The results of our study indicated that even a low concentration of AGEs induced neuronal apoptosis and decreased neurite regeneration in cultured retinas [57]. Although there were no differences in the toxic effects among the three types of AGEs, the neuroprotective and regenerative effects of neurotrophin-4 (NT-4) in glycolaldehyde- and glyceraldehyde-derived AGEs incubated retinas were less than those in glucose-derived AGEs incubated retinas [57]. The results of our other study suggested that glycolaldehyde- and glyceraldehyde-derived AGEs reduced the neuroprotective and regenerative effects of neurotrophic factors, NT-4, glial cell line-derived neurotrophic factor, hepatocyte growth factor, and tauroursodeoxycholic acid (TUDCA) more than the glucose-derived AGEs [58]. Taken together, aldehyde-derived AGEs may reduce the therapeutic effects of neurotrophic factors in diabetic retinas.
The AGEs–RAGE axis is associated with the pathogenesis of diabetic complications, including diabetic retinopathy [67,68]. However, in cultured retinas, the neuroprotective and regenerative effects of the RAGE inhibitors were significantly weaker than the other neurotrophic factors, NT-4, TUDCA, and citicoline [64]. Thus, not only the RAGE-dependent pathways but also the RAGE-independent pathways should be involved in the pathogenesis of neuronal abnormalities in diabetic retinas (Figure 4). On the other hand, the AGE/RAGE signaling upregulates NF-κB and VEGF, which then results in vascular abnormalities in diabetic retinas [68]. The results of our previous study indicated that the expression of NF-κB was increased in AGEs incubated retinas, and its expression was correlated with an increase in neuronal cell death in cultured retinas [58] (Figure 4). Thus, inhibition of the AGE/RAGE signaling may be one of the therapeutic options to prevent the progression of diabetic retinopathy.
5. Potential Anti-AGEs Therapies for Diabetic Retinopathy
Anti-glycation or anti-AGEs therapies are becoming areas of great interest for the treatment of diabetic complications, including diabetic retinopathy. The levels of exogenous AGEs can be reduced by saving AGEs-rich foods and stopping smoking. However, chronic hyperglycemia leads to the accumulation of endogenous AGEs as described, and thus, anti-glycation or anti-AGEs therapies are needed to reduce the endogenous AGEs in the body. The ideal anti-AGE drugs should have multiple effects, including inhibition of AGEs formation, breakers of cross-links, antioxidants, RAGE antagonists, metal chelating, and carbonyl compound scavengers. Although no ideal anti-AGEs drug has been developed, there are several anti-AGEs drugs, such as chemical drugs, natural drugs, or clinical drugs and natural compounds that have already been used.
5.1. AGE Inhibitors
The ideal AGE inhibitor should contain all of the effective factors, e.g., antioxidants, metal chelators, and carbonyl scavengers. Because the Fenton reaction is involved in the late stage of AGE formation, metal chelating reagents are candidates for AGE inhibitors [69]. Although there are no ideal AGE inhibitors, some candidates, both chemical and natural AGE inhibitors, have been suggested.
Aminoguanidine is one of the old and well-known chemical AGE inhibitors. Aminoguanidine acts as a dicarbonyl scavenger, a free radical scavenger, and a metal chelator [70,71]. Two clinical trials have been performed that examined the effects of aminoguanidine on diabetic nephropathy in patients with type 1 (ACTION I) [72] and type 2 (ACTION II) [73]. The ACTION II study was suspended because of severe side effects, including infection, liver dysfunction, lupus-like symptoms, and ANCA-associated vasculitis. The ACTION I group completed the study, but similar side effects were observed. However, the primary end point, which was the time to double the creatinine level in the serum, was not reached [72]. On the other hand, one of the secondary end points, the evaluation of retinopathy progression, was achieved, i.e., aminoguanidine delayed the progression determined by the Early Treatment of Diabetic Retinopathy Study score [72]. However, the effects of aminoguanidine on delaying the progression of diabetic retinopathy were not repeated. An AGE inhibitor, OPB-9195, was developed in Japan to reduce the progression of nephropathy by inhibiting the synthesis of tumor growth factor-β. However, the trial was stopped for unknown reasons [74]. LR-90 is a new AGE inhibitor whose inhibitory effects are stronger than aminoguanidine and pyridoxamine in vitro [75]. LR-90 is thought to be a dicarbonyl scavenger as well as a metal chelator. A recent study showed that LR-90 reduced the acellular capillary numbers and pericyte loss in diabetic animal models, and the authors concluded that LR-90 could be a drug for preventing retinopathy in diabetic patients [76].
Pyridoxamine, an amine of vitamin B6, is a well-known natural AGE inhibitor. It acts as a dicarbonyl scavenger [77], an antioxidant [77,78], and a metal chelator [79,80]. Stitt et al. demonstrated that pyridoxamine reduced capillary drop-out and the expression of extracellular matrix genes in the retinal vessels of diabetic rats. They concluded that pyridoxamine might be useful for treating diabetic retinopathy [81]. Although the primary end points could not be reached, pyridoxamine was shown to be safe and tolerable in two clinical trials in patients with diabetic nephropathy [82,83]. Pramanik et al. examined the effects of vitamin B, C, and E supplementation on the progression of diabetic retinopathy [84]. The clinical trial included 175 patients with type 2 diabetes, and the follow-up period was five years. The results indicated that the supplementation delayed the development of diabetic retinopathy and reduced pathological biomarkers such as ROS, malondialdehyde, AGEs, and VEGF [84]. Recent studies have focused on the neuroprotective effects of pyridoxamine on diabetic retinopathy. Ren et al. showed that pyridoxamine had a protective effect on the photoreceptors of diabetic mice [85]. This protection was accomplished by an ERK /nuclear factor erythroid 2-related factor 2/apoptosis signal-regulating kinase 1 signaling pathway [85]. The results of another study showed that the glyceraldehyde-derived AGEs reduced the neurite outgrowth in human neuroblastoma SH-SY5Y cells, but aminoguanidine and pyridoxamine ameliorated the suppression of neurite outgrowth induced by glyceraldehyde-derived AGEs [86]. Thus, aminoguanidine and pyridoxamine may prevent the development of neuronal abnormalities in diabetic retinopathy. Further studies are needed to determine more conclusively the neuroprotective and regenerative effects of aminoguanidine and pyridoxamine in diabetic retinas. Pyridoxamine is a natural compound, and Japanese sushi toppings include abundant amounts of pyridoxamine [87].
Benfotiamine is a lipophilic derivative of thiamine (vitamin B1) which is absorbed in the body more easily than thiamine. Benfotiamine inhibits the hexosamine pathway, AGE formation, and the DAG-PKC pathway by activating transketolase [88]. In addition, the number of acellular vessels was reduced by inhibiting these three pathways and also by the activation of NF-κB in the retinas of diabetic animals [88]. Clinical trials for examining the effects of benfotiamine on diabetic neuropathy showed positive results in which the neuropathy scores were significantly improved without any severe side effects. Unfortunately, the followed-up periods were short [89,90]. A long-term clinical trial that assesses the length of the corneal nerve fibers by corneal confocal microscopy is ongoing (DRKS00014832) [91]. Clinical trials on the effects of benfotiamine on diabetic retinopathy have not been performed. However, the results of recent studies showed that patients with diabetic polyneuropathy had significantly thinner retinal nerve fiber layers than patients without diabetic polyneuropathy. In addition, the parameters of the ganglion cell complex in early diabetic retinopathy were distinctly abnormal in patients with diabetic polyneuropathy [92,93,94,95]. These results suggested that the pathogenesis of diabetic polyneuropathy is, in part, similar to that of diabetic neuropathy in the retina. Thus, drugs effective for diabetic polyneuropathy may be effective in preventing or slowing the development of diabetic neuropathy in the retina. Further studies are needed to examine the effect of benfotiamine on early diabetic retinopathy.
Trapa bispinosa roxb., the water chestnut, is a well-known small herb that contains a high quantity of minerals, ions, and vitamins [96]. Previous in vitro studies showed that trapa bispinosa roxb. had significant antioxidative effects against free radicals [97], and it could inhibit the synthesis of CML and the cross-linking of AGEs [98]. It can also degrade the α-dicarbonyl compounds [98]. Recent in vivo studies showed that the systemic administration of trapa bispinosa roxb. and lutein reduced the accumulation of AGEs in the retina of streptozotocin-induced diabetic rats [99]. It also improved the abnormal regulation of retinal blood flow in type 2 diabetic mice [100]. In addition, trapa bispinosa roxb. and lutein reduced the expression of the glial fibrillary acid protein in the endfeet of Müller cells and also reduced the expression of VEGF in the retinas of diabetic mice [100]. The authors concluded that trapa bispinosa roxb. and lutein improved the abnormal neurovascular coupling in the retinas of diabetic mice [100]. Extracts of trapa bispinosa roxb. mixed with lutein and zeaxanthin have become commercially available in Japan, but no clinical trials have been performed to evaluate their effects on preventing or slowing the progression of diabetic retinopathy. Thus, clinical studies are needed to determine the effectiveness of these supplements for the early treatment of patients with diabetic retinopathy.
5.2. AGE Cross-Link Breakers
Once the AGEs are synthesized, the AGE inhibitors cannot reduce the level of accumulated AGEs in the body. AGE breakers are one of the options for reducing the level of accumulated AGEs. Alagebrium chloride (ALT-711) is a well-known AGE breaker, and it breaks down the existing AGE-protein cross-links and ameliorates the pathological changes in diabetic complications [101]. AGE-protein cross-links are associated with the pathogenesis of atherosclerosis and cardiovascular diseases [102]. A recent population-based study showed that the accumulation of AGEs was independently associated with heart failure, and this association was more prominent in patients with diabetes [103]. However, an earlier clinical trial reported that alagebrium did not have a beneficial effect on the heart [104].
Another AGE cross-link breaker, TRC4186, had good tolerance and was safe in the phase I clinical trial [105], but no further studies were found.
The exact mechanism of AGE breakers has not been completely determined, and further studies are needed to understand the pharmacological mechanism of AGE cross-link breakers. There are few studies concerning the effects of AGE breakers on diabetic retinopathy. However, a recent study showed that a major dietary flavonoid, epicatechin, broke down the existing glycated albumin in vitro and reduced the accumulated AGE in the retinas in vivo [106]. Thus, epicatechin may be a potential AGE breaker and should be considered for treating diabetic retinopathy.
Curcumin (diferuloylmethane) is a representative flavonoid, and tetrahydrocurcumin, one of the major metabolites of curcumin, is a potential AGE breaker [107]. Tetrahydrocurcumin has a higher antioxidant activity than curcumin [108] and may be a stronger AGE breaker than curcumin [107]. Curcumin ameliorated both vascular and neuronal abnormalities in diabetic retinas in vivo by reducing the levels of VEGF and tumor necrosis factor-α, protecting the inner retinal barrier, reducing oxidative stress, and capillary basement membrane thickening [109,110,111]. Filippelli et al. reported that in patients with proliferative diabetic retinopathy, curcumin slightly reduced the pro-inflammatory cytokines and soluble mediators in the vitreous [112]. One of the weak points of curcumin and tetrahydrocurcumin is their low aqueous solubility which results in their poor bioavailability in the eye. However, Maharjan et al. recently developed a drug delivery system, the encapsulation of curcumin or tetrahydrocurcumin into the hydroxypropyl-cyclodextrins, which led to an increase in its solubility and enhanced the retinal epithelial permeability [113]. This system increased the antioxidant activity of the ocular epithelial cells and may become an ocular medication in the future [113].
5.3. RAGE Blockers and Inhibitors
AGE/RAGE signaling activates several pathways, including NADPH, Ras-ERK, Ras-MAPK, Ras-JNK, and endothelial nitric oxide synthase (eNOS). Activation of these pathways results in ROS formation, inflammatory reaction, apoptosis, and NO synthesis [9,114]. Most signals can directly or indirectly upregulate NF-κB [114] (Figure 4). Although the RAGE-independent axis cannot be blocked, RAGE blockers and inhibitors can be, in part, anti-AGE therapies for diabetic retinopathy (Figure 4).
For example, azeliragon (TTP488 or PF-4494700) is known to be a RAGE inhibitor by binding with a RAGE binding site and ameliorates Alzheimer’s disease via the Janus tyrosine kinase (JAK) and signal transducer and activator of transcription (STAT) pathway [115]. The JAK-STAT pathway is activated under the AGE/RAGE signaling and related to vascular remodeling and cellular proliferation leading to renal hypertrophy. Because there are pathophysiological pharmacotherapeutics links between Alzheimer’s disease and diabetes [116], azeliragon is one of the therapeutic options for diabetic complications. In fact, azeliragon ameliorates diabetic neuropathy in diabetic animal models [117].
5.4. Existing Drugs and Natural Compounds for Various Diseases
There are several existing drugs, such as glitazones, angiotensin-converting enzyme inhibitors (ACEIs), statins, or metformin, that have some effects on the AGE inhibitors. These drugs have some effects on AGE inhibitors, and thus, their anti-AGE effects may be limited, but the merit of these drugs is that they are already in use as clinical medications and their safety and tolerance have been determined.
One of the oral anti-diabetic drugs, pioglitazone, is an insulin sensitizer and is used in Japan as an additional anti-diabetic drug for patients with diabetic retinopathy. A clinical randomized controlled trial showed that pioglitazone increased the level of soluble RAGE and endogenous secretory RAGE, which are AGE/RAGE antagonists. It also decreased the expression of RAGE [118]. In addition, these anti-AGE effects were independent of the insulin resistance level [118]. Alpha-glucosidase inhibitors, another anti-diabetic drug, decreased the level of AGEs in the serum [119]. However, recent studies showed that several natural compounds inhibit α-glucosidase and AGE formation, such as Vernonia amygdalina extracts [120], pregnane glycosides, and the flavonoids isolated from Carelluma hexagone [121]; Nymphoides indica rhizome extracts [122], flavonoids, and proanthocyanidins-rich fractions extracted from Eugenia dysenterica fruits and leaves [123]; isoflavonoids isolated from Masclura tricuspidate [124]; isolated active constituents from Osmanthus fragrans flowers and structural analogs [125]; phenolics isolated from Eugenia jambolana [126]; or phenolics isolated from Chrozophora oblongifolia aerial parts [127]. Alpha-glucosidase and AGE formation is a much-discussed topic, and thus, some natural compounds that inhibit α-glucosidase and AGE formation may become commercially available as medications or supplementations in the near future.
Angiotensin II type 1 receptor blocker (ARB) and ACEI are used as major anti-hypertension drugs worldwide. Both ARB and ACEI reduce AGE formation as metal chelators and antioxidants in vitro [128] and in vivo [129,130]. The effects of anti-AGE accumulation of these drugs were shown in patients with diabetes mellitus [131,132]. A recent meta-analysis reported that the renin-angiotensin system inhibitors reduced the risk of diabetic retinopathy [133]. In addition, ACEI might be more effective than ARBs for the regression of diabetic retinopathy (registered number of International Prospective Register of Systematic Reviews, CRD42013004548) [133]. Sun et al. recently demonstrated that the ACE-mediated transforming growth factor-β1/Smad signaling pathway was associated with the blood–retinal barrier breakdown in diabetic retinopathy [134].
Other existing drugs for various diseases that have anti-AGE effects are calcium channel blockers, amlodipine and azelinidipine [135], an anti-hyperlipidemia drug, statin [136,137], and anti-diabetic drug metformin [138]. Recently, metformin was considered to be a potential therapeutic drug for neurological diseases because metformin enhances mitochondrial function by its activation of AMP-activated protein kinase (AMPK) [139]. The neuroprotective effect of metformin for patients with acute stroke and type 2 diabetes was associated with the AMPK/mammalian target of the rapamycin signaling pathway [140]. Furthermore, metformin can reduce the risk of dementia in patients with diabetes [141]. Metformin had protective effects on diabetic rat retinas by suppressing the toll-like receptor 4/NF-κB and had antioxidative stress [142]. A recent population-based cohort study reported that metformin reduced the risk of developing non-proliferative diabetic retinopathy and potentially reduced the risk of sight-threatening diabetic retinopathy [143].
Many natural compounds are known to have anti-glycation properties, but their bioactivities are relatively weak. Although there are safety profiles for natural compounds, a continuous intake as a supplement is required to prevent the development and progression of diabetic retinopathy. Several well-known natural compounds that have anti-AGE effects have been introduced in this review. For example, cinnamon bark extract inhibits AGE formation by trapping reactive carbonyls [144]. Cinnamon bark proanthocyanidins have high antioxidant and carbonyl scavenging capacity [145]. These anti-glycemic effects may be responsible for cinnamon bark’s constituents, such as catechin, epicatechin, and procyanidin B2, in improving the retinal functional abnormalities as assessed by electroretinography. They also protect the photoreceptor cells by suppressing glial activity, angiogenesis, and oxidative stress [146].
Garlic belongs to the genus Allium, and garlic extracts have antioxidative and free-radical scavenging effects in diabetic rats [147]. Another in vivo diabetic animal study reported that garlic reduced the total oxidative status, NO levels, and TNF-α protein levels. Therefore, garlic extracts have hypoglycemic, antioxidant, and anti-inflammatory properties [148]. The results of a recent double-blind, randomized clinical trial showed that garlic tablets improved visual acuity and decreased central macular thickness. Thus, garlic tablets can be used as a complementary treatment for diabetic retinopathy [149].
Mate tea is prepared from the dry leaves of Ilex paraguariensis, and it contains caffeic acid and chlorogenic acid, which are responsible for its antiglycation properties [150]. Although Ilex paraguariensis extracts have more anti-glycemic activities than green tea extracts [151], there are no publications regarding the effect of mate tea on diabetic retinopathy.
Green tea is made from the leaves of the Camellia sinensis L plant, and several polyphenol antioxidants are part of its constituents. The antioxidative effects of green tea are associated with its neuroprotective effect on diabetic retinopathy in vivo [152]. A clinic-based case-control study showed that regular Chinese green tea consumption significantly reduced the risk of developing diabetic retinopathy [153]. Similarly, a rural community-based, cross-sectional survey (n = 5281) demonstrated that long-term tea consumption reduced the risk of diabetic retinopathy compared to non-tea consumption [154].
AGEs are associated with neuronal cell death in several diseases, such as Alzheimer’s disease [155,156], brain ischemia [157], Parkinson’s disease [158], and diabetic retinopathy [159]. Similar to cultured retinal neurons [57,58], glyceraldehyde-derived AGEs are associated with neuronal cell death in Alzheimer’s disease [155]. A citrus flavonoid, hesperetin inhibited the ER stress-mediated neuronal apoptosis in AGEs-induced neuroblastoma cells [160]. The ER stress-mediated cell death was similarly observed in AGEs-exposed cultured retinal neurons [58,64]. Thus, hesperetin may be a potential agent for treating Alzheimer’s disease as well as diabetic retinopathy.
A bioflavonoid compound in citrus fruits, diosmetin, protected AGEs-induced neuroblastoma cell death by suppressing the ER stress-related cell death pathways [161]. Rosemary extracts are known to have comparable anti-glycation activity to aminoguanidine [162]. The results of a recent study indicated that rosmarinic acid inhibited DNA glycation, which could lead to hippocampal neuron death in diabetic animals, and it normalized the Akt1 and Akt3 expression [163]. Akt1 and Akt3 are inhibitors of apoptosis, and their expressions were downregulated in diabetic animals. Recently, Vieira et al. developed a rosmarinic acid-loaded poly lactic-co-glycolic acid intraocular implant [164]. The delivery device showed no toxic effects and reduced new vessel formation in rabbit eyes [164]. Thus, the intravitreal implant can prolong the release of rosmarinic acid and potentially can be used to prevent neovascularization in ophthalmic diseases, including diabetic retinopathy.
6. Conclusions
Increased AGE accumulation induces not only vascular abnormalities but also neuronal abnormalities, and it is involved in several retinal and optic nerve diseases, including diabetic retinopathy, age-related macular degeneration, and glaucoma. In addition, AGEs are associated with neuronal diseases such as Alzheimer’s disease, Parkinson’s disease, and strokes. Anti-AGE therapies are one of the options for preventing the dysfunction of the NVUs of diabetic retinopathy. The first-line anti-AGE therapy is reducing the intake of exogenous AGEs. Thus, low-AGE food products are practically selected for patients with diabetes. However, chronic hyperglycemia leads to endogenous AGE accumulation in the body of diabetic patients, which results in the development of diabetic complications. Therefore, anti-AGE drugs should be required to reduce endogenous accumulated AGEs. The anti-AGE drugs should have the effects of antioxidants, metal chelators, and carbonyl scavengers. In addition, cross-link breakers and RAGE inhibitors are also potential drugs for anti-AGE therapies. However, there are no ideal anti-AGE drugs with significant therapeutic effects and without any side effects. Natural compounds have lower bioactivities for anti-AGE effects than chemical compounds, but they have the merits of their safety profiles. Some natural compounds for anti-AGE therapies may be useful for the treatment of patients with early diabetic retinopathy. Further basic and clinical studies should be performed to establish the effectiveness and safety of anti-AGE drugs for diabetic retinopathy.
Funding
This research was supported by a Grant-in Aid from the Ministry of Education, Science, Sports, and Culture of the Japanese Government (grant number 22K09829).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thanks Duco Hamasaki of the Bascom Palmer Institute for editing the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.D.; Chew, E.; Duh, E.J.; Sobrin, L.; Sun, J.K.; VanderBeek, B.L.; Wykoff, C.C.; Gardner, T.W. Diabetic retinopathy: A position statement by the American Diabetes Association. Diabetes Care 2017, 40, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Fong, D.S.; Aiello, L.; Gardner, T.W.; King, G.L.; Blankenship, G.; Cavallerano, J.D.; Ferris, F.L., 3rd; Klein, R.; American Diabetes Association. Diabetic retinopathy. Diabetes Care 2003, 26, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Leasher, J.L.; Bourne, R.R.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Naidoo, K.; Pesudovs, K.; Price, H.; White, R.A.; Wong, T.Y.; et al. Global estimates on the number of people blind or visually impaired by diabetic retinopathy: A meta-analysis from 1990 to 2010. Diabetes Care 2016, 39, 1643–1649. [Google Scholar] [CrossRef]
- Yau, J.W.; Rogers, S.L.; Kawasaki, R.; Lamoureux, E.L.; Kowalski, J.W.; Bek, T.; Chen, S.J.; Dekker, J.M.; Fletcher, A.; Grauslund, J.; et al. Meta-Analysis for Eye Disease (META-EYE) Study Group. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care 2012, 35, 556–564. [Google Scholar] [CrossRef]
- Oshitari, T. Diabetic retinopathy: Neurovascular disease requiring neuroprotective and regenerative therapies. Neural Regen. Res. 2022, 17, 795–796. [Google Scholar] [CrossRef]
- Oshitari, T. The pathogenesis and therapeutic approaches of diabetic neuropathy in the retina. Int. J. Mol. Sci. 2021, 22, 9050. [Google Scholar] [CrossRef]
- Antonetti, D.A.; Klein, R.; Gardner, T.W. Diabetic retinopathy. N. Engl. J. Med. 2012, 366, 1227–1239. [Google Scholar] [CrossRef]
- Oshitari, T. Neurovascular impairment and therapeutic strategies in diabetic retinopathy. Int. J. Environ. Res. Public Health 2021, 19, 439. [Google Scholar] [CrossRef]
- Donato, L.; Scimone, C.; Alibrandi, S.; Scalinci, S.Z.; Rinaldi, C.; D’Angelo, R.; Sidoti, A. Epitranscriptome Analysis of Oxidative Stressed Retinal Epithelial Cells Depicted a Possible RNA Editing Landscape of Retinal Degeneration. Antioxidants 2022, 11, 1967. [Google Scholar] [CrossRef]
- Scimone, C.; Donato, L.; Alibrandi, S.; Esposito, T.; Alafaci, C.; D’Angelo, R.; Sidoti, A. Transcriptome analysis provides new molecular signatures in sporadic Cerebral Cavernous Malformation endothelial cells. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165956. [Google Scholar] [CrossRef]
- Donato, L.; Alibrandi, S.; Scimone, C.; Rinaldi, C.; Dascola, A.; Calamuneri, A.; D’Angelo, R.; Sidoti, A. The impact of modifier genes on cone-rod dystrophy heterogeneity: An explorative familial pilot study and a hypothesis on neurotransmission impairment. PLoS ONE 2022, 17, e0278857. [Google Scholar] [CrossRef]
- Miyata, T.; Kurokawa, K.; van Ypersele, D.E.; Strihou, C. Advanced glycation and lipoxidation end products: Role of reactive carbonyl compounds generated during carbohydrate and lipid metabolism. J. Am. Soc. Nephrol. 2000, 11, 1744–1752. [Google Scholar] [CrossRef]
- Mengstie, M.A.; Chekol Abebe, E.; Behaile Teklemariam, A.; Tilahun Mulu, A.; Agidew, M.M.; Teshome Azezew, M.; Zewde, E.A.; Agegnehu Teshome, A. Endogenous advanced glycation end products in the pathogenesis of chronic diabetic complications. Front. Mol. Biosci. 2022, 9, 1002710. [Google Scholar] [CrossRef]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef]
- Szwergold, B.S.; Kappler, F.; Brown, T.R. Identification of fructose 3-phosphate in the lens of diabetic rats. Science 1990, 247, 451–454. [Google Scholar] [CrossRef]
- Odani, H.; Iijima, K.; Nakata, M.; Miyata, S.; Kusunoki, H.; Yasuda, Y.; Hiki, Y.; Irie, S.; Maeda, K.; Fujimoto, D. Identification of N(omega)-carboxymethylarginine, a new advanced glycation endproduct in serum proteins of diabetic patients: Possibility of a new marker of aging and diabetes. Biochem. Biophys. Res. Commun. 2001, 285, 1232–1236. [Google Scholar] [CrossRef]
- Ahmed, N.; Argirov, O.K.; Minhas, H.S.; Cordeiro, C.A.; Thornalley, P.J. Assay of advanced glycation endproducts (AGEs): Surveying AGEs by chromatographic assay with derivatization by 6-aminoquinolyl-N-hydroxysuccinimidyl-carbamate and application to Nepsilon-carboxymethyl-lysine- and Nepsilon-(1-carboxyethyl)lysine-modified albumin. Biochem. J. 2002, 364, 1–14. [Google Scholar]
- Nagaraj, R.H.; Monnier, V.M. Protein modification by the degradation products of ascorbate: Formation of a novel pyrrole from the Maillard reaction of L-threose with proteins. Biochim. Biophys. Acta 1995, 1253, 75–84. [Google Scholar] [CrossRef]
- Shipanova, I.N.; Glomb, M.A.; Nagaraj, R.H. Protein modification by methylglyoxal: Chemical nature and synthetic mechanism of a major fluorescent adduct. Arch. Biochem. Bophys. 1997, 344, 29–36. [Google Scholar] [CrossRef]
- Sell, D.R.; Monnier, V.M. Structure elucidation of a senescence cross-link from human extracellular matrix. Implication of pentoses in the aging process. J. Biol. Chem. 1989, 264, 21597–21602. [Google Scholar] [CrossRef]
- Obayashi, H.; Nakano, K.; Shigeta, H.; Yamaguchi, M.; Yoshimori, K.; Fukui, M.; Fujii, M.; Kitagawa, Y.; Nakamura, N.; Nakamura, K.; et al. Formation of crossline as a fluorescent advanced glycation end product in vitro and in vivo. Biochem. Biophys. Res. Commun. 1996, 226, 37–41. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Thorpe, S.R.; Baynes, J.W. Identification of N epsilon-carboxymethyllysine as a degradation product of fructoselysine in glycated protein. J. Biol. Chem. 1986, 261, 4889–4894. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.U.; Brinkmann Frye, E.; Degenhardt, T.P.; Thorpe, S.R.; Baynes, J.W. N-epsilon-(carboxyethyl)lysine, a product of the chemical modification of proteins by methylglyoxal, increases with age in human lens proteins. Biochem. J. 1997, 324, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, R.H.; Shipanova, I.N.; Faust, F.M. Protein cross-linking by the Maillard reaction. Isolation, characterization, and in vivo detection of a lysine-lysine cross-link derived from methylglyoxal. J. Biol. Chem. 1996, 271, 19338–19345. [Google Scholar] [CrossRef] [PubMed]
- Zawada, A.; Machowiak, A.; Rychter, A.M.; Ratajczak, A.E.; Szymczak-Tomczak, A.; Dobrowolska, A.; Krela-Kaźmierczak, I. Accumulation of Advanced Glycation End-Products in the Body and Dietary Habits. Nutrients 2022, 14, 3982. [Google Scholar] [CrossRef]
- Luo, X.; Wu, J.; Jing, S.; Yan, L.J. Hyperglycemic stress and carbon stress in diabetic glucotoxicity. Aging Dis. 2016, 7, 90–110. [Google Scholar] [CrossRef]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef]
- Pannicke, T.; Iandiev, I.; Wurm, A.; Uckermann, O.; vom Hagen, F.; Reichenbach, A.; Wiedemann, P.; Hammes, H.P.; Bringmann, A. Diabetes alters osmotic swelling characteristics and membrane conductance of glial cells in rat retina. Diabetes 2006, 55, 633–639. [Google Scholar] [CrossRef]
- Rinaldi, C.; Donato, L.; Alibrandi, S.; Scimone, C.; D’Angelo, R.; Sidoti, A. Oxidative stress and neurovascular unit. Life 2021, 11, 767. [Google Scholar] [CrossRef]
- Wong, A.; Lüth, H.J.; Deuther-Conrad, W.; Dukic-Stefanovic, S.; Gasic-Milenkovic, J.; Arendt, T.; Münch, G. Advanced glycation endproducts co-localize with inducible nitric oxide synthase in Alzheimer’s disease. Brain Res. 2001, 920, 32–40. [Google Scholar] [CrossRef]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid. Med. Cell Longev. 2020, 2020, 3818196. [Google Scholar] [CrossRef]
- Takeuchi, M.; Watai, T.; Sasaki, N.; Choei, H.; Iwaki, M.; Ashizawa, T.; Inagaki, Y.; Yamagishi, S.; Kikuchi, S.; Riederer, P.; et al. Neurotoxicity of acetaldehyde-derived advanced glycation end products for cultured cortical neurons. J. Neuropathol. Exp. Neurol. 2003, 62, 486–496. [Google Scholar] [CrossRef]
- Bierhaus, A.; Stern, D.M.; Nawroth, P.P. RAGE in inflammation: A new therapeutic target? Curr. Opin. Investig. Drugs 2006, 7, 985–991. [Google Scholar]
- Sergi, D.; Boulestin, H.; Campbell, F.M.; Williams, L.M. The Role of Dietary Advanced Glycation End Products in Metabolic Dysfunction. Mol. Nutr. Food Res. 2021, 65, e1900934. [Google Scholar] [CrossRef]
- Kandarakis, S.A.; Piperi, C.; Topouzis, F.; Papavassiliou, A.G. Emerging role of advanced glycation-end products (AGEs) in the pathobiology of eye diseases. Prog. Retin. Eye Res. 2014, 42, 85–102. [Google Scholar] [CrossRef]
- Gąsiorowski, K.; Brokos, B.; Echeverria, V.; Barreto, G.E.; Leszek, J. RAGE-TLR Crosstalk Sustains Chronic Inflammation in Neurodegeneration. Mol. Neurobiol. 2018, 55, 1463–1476. [Google Scholar] [CrossRef]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Chung, Y.R.; Choi, J.A.; Koh, J.Y.; Yoon, Y.H. Ursodeoxycholic acid attenuates endoplasmic reticulum stress-related retinal pericyte loss in streptozotocin-induced diabetic mice. J. Diabetes Res. 2017, 2017, 1763292. [Google Scholar] [CrossRef]
- Wang, Y.K.; Zhu, L.; Zhang, L.; Zhang, G.; Liu, Y.; Wang, A. A portable system for noninvasive assessment of advanced glycation end-products using skin fluorescence and reflectance spectrum. J. Appl. Spectrosc. 2012, 79, 431–436. [Google Scholar] [CrossRef]
- Ying, L.; Shen, Y.; Zhang, Y.; Wang, Y.; Liu, Y.; Yin, J.; Wang, Y.; Yin, J.; Zhu, W.; Bao, Y.; et al. Association of advanced glycation end products with diabetic retinopathy in type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2021, 177, 108880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ma, P.; Chen, Q. The Correlation between the Level of Skin Advanced Glycation End Products in Type 2 Diabetes Mellitus and the Stages of Diabetic Retinopathy and the Types of Traditional Chinese Medicine Syndrome. Evid. Based Complement Alternat. Med. 2022, 2022, 5193944. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, Y.; Yamanaka, M.; Fujihara, J.; Matsuoka, Y.; Gohto, Y.; Obana, A.; Tanito, M. Evaluation of Relevance between Advanced Glycation End Products and Diabetic Retinopathy Stages Using Skin Autofluorescence. Antioxidants 2020, 9, 1100. [Google Scholar] [CrossRef]
- Murata, T.; Nagai, R.; Ishibashi, T.; Inomuta, H.; Ikeda, K.; Horiuchi, S. The relationship between accumulation of advanced glycation end products and expression of vascular endothelial growth factor in human diabetic retinas. Diabetologia 1997, 40, 764–769. [Google Scholar] [CrossRef]
- Moore, T.C.; Moore, J.E.; Kaji, Y.; Frizzell, N.; Usui, T.; Poulaki, V.; Campbell, I.L.; Stitt, A.W.; Gardiner, T.A.; Archer, D.B.; et al. The role of advanced glycation end products in retinal microvascular leukostasis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4457–4464. [Google Scholar] [CrossRef]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S. Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. Biomed Res. Int. 2014, 2014, 801269. [Google Scholar] [CrossRef]
- Huang, H. Pericyte-Endothelial Interactions in the Retinal Microvasculature. Int. J. Mol. Sci. 2020, 21, 7413. [Google Scholar] [CrossRef]
- Sasaki, J.; Takayanagi, Y.; Kadoh, Y.; Tanito, M. Relevance of Diabetic Retinopathy with AGEs and Carotenoid Levels Assessed by Skin Sensors. Antioxidants 2022, 11, 1370. [Google Scholar] [CrossRef]
- Shalini, T.; Jose, S.S.; Prasanthi, P.S.; Balakrishna, N.; Viswanath, K.; Reddy, G.B. Carotenoid status in type 2 diabetes patients with and without retinopathy. Food Funct. 2021, 12, 4402–4410. [Google Scholar] [CrossRef]
- Miura, J.; Yamagishi, S.I.; Uchigata, Y.; Takeuchi, M.; Yamamoto, H.; Makita, Z.; Iwamoto, Y. Serum levels of non-carboxymethyllysine advanced glycation endproducts are correlated to severity of microvascular complications in patients with Type 1 diabetes. J. Diabetes Complicat. 2003, 17, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Makita, Z.; Yanagisawa, K.; Kameda, Y.; Koike, T. Detection of noncarboxymethyllysine and carboxymethyllysine advanced glycation end products (AGE) in serum of diabetic patients. Mol. Med. 1999, 5, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagishi, S.; Amano, S.; Inagaki, Y.; Okamoto, T.; Koga, K.; Sasaki, N.; Yamamoto, H.; Takeuchi, M.; Makita, Z. Advanced glycation end products-induced apoptosis and overexpression of vascular endothelial growth factor in bovine retinal pericytes. Biochem. Biophys. Res. Commun. 2002, 290, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Yamagishi, S.; Inagaki, Y.; Amano, S.; Koga, K.; Abe, R.; Takeuchi, M.; Ohno, S.; Yoshimura, A.; Makita, Z. Angiogenesis induced by advanced glycation end products and its prevention by cerivastatin. FASEB J. 2002, 16, 1928–1930. [Google Scholar] [CrossRef]
- Takeuchi, M.; Bucala, R.; Suzuki, T.; Ohkubo, T.; Yamazaki, M.; Koike, T.; Kameda, Y.; Makita, Z. Neurotoxicity of advanced glycation end-products for cultured cortical neurons. J. Neuropathol. Exp. Neurol. 2000, 59, 1094–1105. [Google Scholar] [CrossRef]
- Choei, H.; Sasaki, N.; Takeuchi, M.; Yoshida, T.; Ukai, W.; Yamagishi, S.; Kikuchi, S.; Saito, T. Glyceraldehyde-derived advanced glycation end products in Alzheimer’s disease. Acta Neuropathol. 2004, 108, 189–193. [Google Scholar] [CrossRef]
- Bikbova, G.; Oshitari, T.; Yamamoto, S. Neurite regeneration in adult rat retinas exposed to advanced glycation end-products and regenerative effects of neurotrophin-4. Brain Res. 2013, 1534, 33–45. [Google Scholar] [CrossRef]
- Bikbova, G.; Oshitari, T.; Baba, T.; Yamamoto, S. Altered Expression of NF- κ B and SP1 after Exposure to Advanced Glycation End-Products and Effects of Neurotrophic Factors in AGEs Exposed Rat Retinas. J. Diabetes Res. 2015, 2015, 543818. [Google Scholar] [CrossRef]
- Oshitari, T.; Dezawa, M.; Okada, S.; Takano, M.; Negishi, H.; Horie, H.; Sawada, H.; Tokuhisa, T.; Adachi-Usami, E. The role of c-fos in cell death and regeneration of retinal ganglion cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2442–2449. [Google Scholar]
- Oshitari, T.; Roy, S. Diabetes: A potential enhancer of retinal injury in rat retinas. Neurosci. Lett. 2005, 390, 25–30. [Google Scholar] [CrossRef]
- Oshitari, T.; Yoshida-Hata, N.; Yamamoto, S. Effect of neurotrophic factors on neuronal apoptosis and neurite regeneration in cultured rat retinas exposed to high glucose. Brain Res. 2010, 1346, 43–51. [Google Scholar] [CrossRef]
- Oshitari, T.; Bikbova, G.; Yamamoto, S. Increased expression of phosphorylated c-Jun and phosphorylated c-Jun N-terminal kinase associated with neuronal cell death in diabetic and high glucose exposed rat retinas. Brain Res. Bull. 2014, 101, 18–25. [Google Scholar] [CrossRef]
- Bikbova, G.; Oshitari, T.; Baba, T.; Yamamoto, S. Mechanisms of neuronal cell death in AGE-exposed retinas—Research and literature review. Curr. Diabetes Rev. 2017, 13, 280–288. [Google Scholar] [CrossRef]
- Bikbova, G.; Oshitari, T.; Baba, T.; Yamamoto, S. Combination of neuroprotective and regenerative agents for AGE-induced retinal degeneration: In vitro study. Biomed Res. Int. 2017, 2017, 8604723. [Google Scholar] [CrossRef]
- Ono, Y.; Aoki, S.; Ohnishi, K.; Yasuda, T.; Kawano, K.; Tsukada, Y. Increased serum levels of advanced glycation end-products and diabetic complications. Diabetes Res. Clin. Pract. 1998, 41, 131–137. [Google Scholar] [CrossRef]
- Lecleire-Collet, A.; Tessier, L.H.; Massin, P.; Forster, V.; Brasseur, G.; Sahel, J.A.; Picaud, S. Advanced glycation end products can induce glial reaction and neuronal degeneration in retinal explants. Br. J. Ophthalmol. 2005, 89, 1631–1633. [Google Scholar] [CrossRef]
- Zong, H.; Ward, M.; Stitt, A.W. AGEs, RAGE, and diabetic retinopathy. Curr. Diab. Rep. 2011, 11, 244–252. [Google Scholar] [CrossRef]
- Xu, J.; Chen, L.J.; Yu, J.; Wang, H.J.; Zhang, F.; Liu, Q.; Wu, J. Involvement of Advanced Glycation End Products in the Pathogenesis of Diabetic Retinopathy. Cell Physiol. Biochem. 2018, 48, 705–717. [Google Scholar] [CrossRef]
- Bavkar, L.N.; Patil, R.S.; Rooge, S.B.; Nalawade, M.L.; Arvindekar, A.U. Acceleration of protein glycation by oxidative stress and comparative role of antioxidant and protein glycation inhibitor. Mol. Cell Biochem. 2019, 459, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Price, D.L.; Rhett, P.M.; Thorpe, S.R.; Baynes, J.W. Chelating activity of advanced glycation end-product inhibitors. J. Biol. Chem. 2001, 276, 48967–48972. [Google Scholar] [CrossRef]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Bolton, W.K.; Cattran, D.C.; Williams, M.E.; Adler, S.G.; Appel, G.B.; Cartwright, K.; Foiles, P.G.; Freedman, B.I.; Raskin, P.; Ratner, R.E.; et al. ACTION I Investigator Group. Randomized trial of an inhibitor of formation of advanced glycation end products in diabetic nephropathy. Am. J. Nephrol. 2004, 24, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.I.; Wuerth, J.P.; Cartwright, K.; Bain, R.P.; Dippe, S.; Hershon, K.; Mooradian, A.D.; Spinowitz, B.S. Design and baseline characteristics for the aminoguanidine Clinical Trial in Overt Type 2 Diabetic Nephropathy (ACTION II). Control Clin. Trials 1999, 20, 493–510. [Google Scholar] [CrossRef]
- Takeuchi, M.; Yamagishi, S.; Makita, Z. OPB-9195. Nihon Rinsho 2002, 60, 606–610. (In Japanese) [Google Scholar]
- Rahbar, S.; Yerneni, K.K.; Scott, S.; Gonzales, N.; Lalezari, I. Novel inhibitors of advanced glycation endproducts (part II). Mol. Cell Biol. Res. Commun. 2000, 3, 360–366. [Google Scholar] [CrossRef]
- Bhatwadekar, A.; Glenn, J.V.; Figarola, J.L.; Scott, S.; Gardiner, T.A.; Rahbar, S.; Stitt, A.W. A new advanced glycation inhibitor, LR-90, prevents experimental diabetic retinopathy in rats. Br. J. Ophthalmol. 2008, 92, 545–547. [Google Scholar] [CrossRef]
- Nagaraj, R.H.; Sarkar, P.; Mally, A.; Biemel, K.M.; Lederer, M.O.; Padayatti, P.S. Effect of pyridoxamine on chemical modification of proteins by carbonyls in diabetic rats: Characterization of a major product from the reaction of pyridoxamine and methylglyoxal. Arch. Biochem. Biophys. 2002, 402, 110–119. [Google Scholar] [CrossRef]
- Ramis, R.; Ortega-Castro, J.; Caballero, C.; Casasnovas, R.; Cerrillo, A.; Vilanova, B.; Adrover, M.; Frau, J. How Does Pyridoxamine Inhibit the Formation of Advanced Glycation End Products? The Role of Its Primary Antioxidant Activity. Antioxidants 2019, 8, 344. [Google Scholar] [CrossRef]
- Wondrak, G.T.; Jacobson, E.L. Vitamin B6: Beyond coenzyme functions. Subcell. Biochem. 2012, 56, 291–300. [Google Scholar]
- Voziyan, P.A.; Khalifah, R.G.; Thibaudeau, C.; Yildiz, A.; Jacob, J.; Serianni, A.S.; Hudson, B.G. Modification of proteins in vitro by physiological levels of glucose: Pyridoxamine inhibits conversion of Amadori intermediate to advanced glycation end-products through binding of redox metal ions. J. Biol. Chem. 2003, 278, 46616–46624. [Google Scholar] [CrossRef]
- Stitt, A.; Gardiner, T.A.; Alderson, N.L.; Canning, P.; Frizzell, N.; Duffy, N.; Boyle, C.; Januszewski, A.S.; Chachich, M.; Baynes, J.W.; et al. The AGE inhibitor pyridoxamine inhibits development of retinopathy in experimental diabetes. Diabetes 2002, 51, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.E.; Bolton, W.K.; Khalifah, R.G.; Degenhardt, T.P.; Schotzinger, R.J.; McGill, J.B. Effects of pyridoxamine in combined phase 2 studies of patients with type 1 and type 2 diabetes and overt nephropathy. Am. J. Nephrol. 2007, 27, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.J.; Greene, T.; Spitalewiz, S.; Blumenthal, S.; Berl, T.; Hunsicker, L.G.; Pohl, M.A.; Rohde, R.D.; Raz, I.; Yerushalmy, Y.; et al. Pyridorin in type 2 diabetic nephropathy. J. Am. Soc. Nephrol. 2012, 23, 131–136. [Google Scholar] [CrossRef]
- Pramanik, S.; Banerjee, K.; Mondal, L.K. The Amelioration of Detrimental Biochemical Anomalies by Supplementing B, C, and E Vitamins in Subjects with Type 2 Diabetes Mellitus May Reduce the Rate of Development of Diabetic Retinopathy. J. Diabetes Res. 2022, 2022, 3886710. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Sun, H.; Zhang, C.; Li, C.; Wang, J.; Shen, J.; Yu, D.; Kong, L. Protective function of pyridoxamine on retinal photoreceptor cells via activation of the p-Erk1/2/Nrf2/Trx/ASK1 signalling pathway in diabetic mice. Mol. Med. Rep. 2016, 14, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Ooi, H.; Nasu, R.; Furukawa, A.; Takeuchi, M.; Koriyama, Y. Pyridoxamine and Aminoguanidine Attenuate the Abnormal Aggregation of β-Tubulin and Suppression of Neurite Outgrowth by Glyceraldehyde-Derived Toxic Advanced Glycation End-Products. Front. Pharmacol. 2022, 13, 921611. [Google Scholar] [CrossRef]
- Do, H.T.; Yagi, T. Individual vitamin B6 contents in selected Japanese sushi toppings. Int. J. Food Sci. Nutr. 2012, 63, 246–249. [Google Scholar] [CrossRef]
- Hammes, H.P.; Du, X.; Edelstein, D.; Taguchi, T.; Matsumura, T.; Ju, Q.; Lin, J.; Bierhaus, A.; Nawroth, P.; Hannak, D.; et al. Benfotiamine blocks three major pathways of hyperglycemic damage and prevents experimental diabetic retinopathy. Nat. Med. 2003, 9, 294–299. [Google Scholar] [CrossRef]
- Haupt, E.; Ledermann, H.; Köpcke, W. Benfotiamine in the treatment of diabetic polyneuropathy--a three-week randomized, controlled pilot study (BEDIP study). Int. J. Clin. Pharmacol. Ther. 2005, 43, 71–77. [Google Scholar] [CrossRef]
- Stracke, H.; Gaus, W.; Achenbach, U.; Federlin, K.; Bretzel, R.G. Benfotiamine in diabetic polyneuropathy (BENDIP): Results of a randomised, double blind, placebo-controlled clinical study. Exp. Clin. Endocrinol. Diabetes 2008, 116, 600–605. [Google Scholar] [CrossRef]
- Bönhof, G.J.; Sipola, G.; Strom, A.; Herder, C.; Strassburger, K.; Knebel, B.; Reule, C.; Wollmann, J.C.; Icks, A.; Al-Hasani, H.; et al. BOND study: A randomised double-blind, placebo-controlled trial over 12 months to assess the effects of benfotiamine on morphometric, neurophysiological and clinical measures in patients with type 2 diabetes with symptomatic polyneuropathy. BMJ Open 2022, 12, e057142. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, M.W.; Byeon, S.H.; Kim, S.S.; Koh, H.J.; Lee, S.C.; Kim, M. Associations between individual retinal layer thicknesses and diabetic peripheral neuropathy using retinal layer segmentation analysis. Retina 2018, 38, 2190–2196. [Google Scholar] [CrossRef]
- Srinivasan, S.; Dehghani, C.; Pritchard, N.; Edwards, K.; Russell, A.W.; Malik, R.A.; Efron, N. Optical coherence tomography predicts 4-year incident diabetic neuropathy. Ophthalmic. Physiol. Opt. 2017, 37, 451–459. [Google Scholar] [CrossRef]
- Salvi, L.; Plateroti, P.; Balducci, S.; Bollanti, L.; Conti, F.G.; Vitale, M.; Recupero, S.M.; Enrici, M.M.; Fenicia, V.; Pugliese, G. Abnormalities of retinal ganglion cell complex at optical coherence tomography in patients with type 2 diabetes: A sign of diabetic polyneuropathy, not retinopathy. J. Diabetes Complicat. 2016, 30, 469–476. [Google Scholar] [CrossRef]
- Srinivasan, S.; Pritchard, N.; Sampson, G.P.; Edwards, K.; Vagenas, D.; Russell, A.W.; Malik, R.A.; Efron, N. Diagnostic capability of retinal thickness measures in diabetic peripheral neuropathy. J. Optom. 2017, 10, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Adkar, P.; Dongare, A.; Ambavade, S.; Bhaskar, V.H. Trapa bispinosa Roxb: A Review on Nutritional and Pharmacological Aspects. Adv. Pharmacol. Sci. 2014, 2014, 959830. [Google Scholar]
- Malviya, N.; Jain, S.; Jain, A.; Jain, S.; Gurjar, R. Evaluation of in vitro antioxidant potential of aqueous extract of Trapa natans L. fruits. Acta Pol. Pharm. 2010, 67, 391–396. [Google Scholar]
- Takeshita, S.; Yagi, M.; Uemura, T.; Yamada, M.; Yonei, Y. Peel extract of water chestnut (Trapa bispinosa Roxb.) inhibits glycation, degradesα-dicarbonyl compound, and breaks advanced glycation end product crosslinks. Glycative Stress Res. 2015, 2, 72–79. [Google Scholar]
- Kinoshita, S.; Sugawa, H.; Nanri, T.; Ohno, R.I.; Shirakawa, J.I.; Sato, H.; Katsuta, N.; Sakake, S.; Nagai, R. Trapa bispinosa Roxb. and lutein ameliorate cataract in type 1 diabetic rats. J. Clin. Biochem. Nutr. 2020, 66, 8–14. [Google Scholar] [CrossRef]
- Hanaguri, J.; Yokota, H.; Kushiyama, A.; Kushiyama, S.; Watanabe, M.; Yamagami, S.; Nagaoka, T. Beneficial Effect of Long-Term Administration of Supplement With Trapa Bispinosa Roxb. and Lutein on Retinal Neurovascular Coupling in Type 2 Diabetic Mice. Front. Physiol. 2022, 13, 788034. [Google Scholar] [CrossRef]
- Toprak, C.; Yigitaslan, S. Alagebrium and Complications of Diabetes Mellitus. Eurasian J. Med. 2019, 51, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.E. Importance of advanced glycation end products in diabetes-associated cardiovascular and renal disease. Am. J. Hypertens. 2004, 17, 31S–38S. [Google Scholar] [CrossRef] [PubMed]
- Arshi, B.; Chen, J.; Ikram, M.A.; Zillikens, M.C.; Kavousi, M. Advanced glycation end-products, cardiac function and heart failure in the general population: The Rotterdam Study. Diabetologia 2023, 66, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Hartog, J.W.; Willemsen, S.; van Veldhuisen, D.J.; Posma, J.L.; van Wijk, L.M.; Hummel, Y.M.; Hillege, H.L.; Voors, A.A.; BENEFICIAL investigators. Effects of alagebrium, an advanced glycation endproduct breaker, on exercise tolerance and cardiac function in patients with chronic heart failure. Eur. J. Heart Fail. 2011, 13, 899–908. [Google Scholar] [CrossRef]
- Chandra, K.P.; Shiwalkar, A.; Kotecha, J.; Thakkar, P.; Srivastava, A.; Chauthaiwale, V.; Sharma, S.K.; Cross, M.R.; Dutt, C. Phase I clinical studies of the advanced glycation end-product (AGE)-breaker TRC4186: Safety, tolerability and pharmacokinetics in healthy subjects. Clin. Drug Investig. 2009, 29, 559–575. [Google Scholar] [CrossRef]
- Kim, J.; Kim, C.S.; Moon, M.K.; Kim, J.S. Epicatechin breaks preformed glycated serum albumin and reverses the retinal accumulation of advanced glycation end products. Eur. J. Pharmacol. 2015, 748, 108–114. [Google Scholar] [CrossRef]
- Pari, L.; Murugan, P. Influence of tetrahydrocurcumin on tail tendon collagen contents and its properties in rats with streptozotocin-nicotinamide-induced type 2 diabetes. Fundam. Clin. Pharmacol. 2007, 21, 665–671. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Deb, L.; Prasad, S. Curcumin differs from tetrahydrocurcumin for molecular targets, signaling pathways and cellular responses. Molecules 2014, 20, 185–205. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Kumar, B.; Nag, T.C.; Agrawal, S.S.; Agrawal, R.; Agrawal, P.; Saxena, R.; Srivastava, S. Curcumin prevents experimental diabetic retinopathy in rats through its hypoglycemic, antioxidant, and anti-inflammatory mechanisms. J. Ocul. Pharmacol. Ther. 2011, 27, 123–130. [Google Scholar] [CrossRef]
- Yang, F.; Yu, J.; Ke, F.; Lan, M.; Li, D.; Tan, K.; Ling, J.; Wang, Y.; Wu, K.; Li, D. Curcumin alleviates diabetic retinopathy in experimental diabetic rats. Ophthalmic Res. 2018, 60, 43–54. [Google Scholar] [CrossRef]
- Zuo, Z.F.; Zhang, Q.; Liu, X.Z. Protective effects of curcumin on retinal Müller cell in early diabetic rats. Int. J. Ophthalmol. 2013, 6, 422–424. [Google Scholar]
- Filippelli, M.; Campagna, G.; Vito, P.; Zotti, T.; Ventre, L.; Rinaldi, M.; Bartollino, S.; dell’Omo, R.; Costagliola, C. Anti-inflammatory Effect of curcumin, homotaurine, and vitamin D3 on human vitreous in patients with diabetic retinopathy. Front. Neurol. 2021, 11, 592274. [Google Scholar] [CrossRef]
- Maharjan, P.; Jin, M.; Kim, D.; Yang, J.; Maharjan, A.; Shin, M.C.; Cho, K.H.; Kim, M.S.; Min, K.A. Evaluation of epithelial transport and oxidative stress protection of nanoengineered curcumin derivative-cyclodextrin formulation for ocular delivery. Arch. Pharm. Res. 2019, 42, 909–925. [Google Scholar] [CrossRef]
- Barlovic, D.P.; Soro-Paavonen, A.; Jandeleit-Dahm, K.A. RAGE biology, atherosclerosis and diabetes. Clin. Sci. 2011, 121, 43–55. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Wang, Y.; Li, J.; Liu, N. Azeliragon ameliorates Alzheimer’s disease via the Janus tyrosine kinase and signal transducer and activator of transcription signaling pathway. Clinics 2021, 76, e2348. [Google Scholar] [CrossRef]
- Rojas, M.; Chávez-Castillo, M.; Bautista, J.; Ortega, Á.; Nava, M.; Salazar, J.; Díaz-Camargo, E.; Medina, O.; Rojas-Quintero, J.; Bermúdez, V. Alzheimer’s disease and type 2 diabetes mellitus: Pathophysiologic and pharmacotherapeutics links. World J. Diabetes 2021, 12, 745–766. [Google Scholar] [CrossRef]
- Ma, S.; Nakamura, Y.; Hisaoka-Nakashima, K.; Morioka, N. Blockade of receptor for advanced glycation end-products with azeliragon ameliorates streptozotocin-induced diabetic neuropathy. Neurochem. Int. 2022, 163, 105470. [Google Scholar] [CrossRef]
- Koyama, H.; Tanaka, S.; Monden, M.; Shoji, T.; Morioka, T.; Fukumoto, S.; Mori, K.; Emoto, M.; Shoji, T.; Fukui, M.; et al. Comparison of effects of pioglitazone and glimepiride on plasma soluble RAGE and RAGE expression in peripheral mononuclear cells in type 2 diabetes: Randomized controlled trial (PioRAGE). Atherosclerosis 2014, 234, 329–334. [Google Scholar] [CrossRef]
- Tsunosue, M.; Mashiko, N.; Ohta, Y.; Matsuo, Y.; Ueda, K.; Ninomiya, M.; Tanaka, S.; Hoshiko, M.; Yoshiyama, Y.; Takeuchi, M.; et al. An alpha-glucosidase inhibitor, acarbose treatment decreases serum levels of glyceraldehyde-derived advanced glycation end products (AGEs) in patients with type 2 diabetes. Clin. Exp. Med. 2010, 10, 139–141. [Google Scholar] [CrossRef]
- Medjiofack Djeujo, F.; Cusinato, F.; Ragazzi, E.; Froldi, G. α-Glucosidase and advanced glycation end products inhibition with Vernonia amygdalina root and leaf extracts: New data supporting the antidiabetic properties. J. Pharm. Pharmacol. 2021, 73, 1240–1249. [Google Scholar] [CrossRef]
- Choucry, M.A.; Shalabi, A.A.; El Halawany, A.M.; El-Sakhawy, F.S.; Zaiter, A.; Morita, H.; Chaimbault, P.; Abdel-Sattar, E. New Pregnane Glycosides Isolated from Caralluma hexagona Lavranos as Inhibitors of α-Glucosidase, Pancreatic Lipase, and Advanced Glycation End Products Formation. ACS Omega 2021, 6, 18881–18889. [Google Scholar] [CrossRef]
- Hanif, M.; Khan, M.A.; Rafey, M.R.; Zaman, A.; Aziz, I.; Amanullah Amin, A. Inhibitive efficacy of Nymphoides indica rhizome extract on α-glucosidase, and cross-link formation of advanced glycation end product. J. Tradit. Chin. Med. 2021, 41, 376–380. [Google Scholar] [PubMed]
- Justino, A.B.; Guerra Silva, H.C.; Franco, R.R.; de Oliveira Cavalcante Pimentel, I.; Silva, N.F.; Saraiva, A.L.; Espindola, F.S. Flavonoids and proanthocyanidins-rich fractions from Eugenia dysenterica fruits and leaves inhibit the formation of advanced glycation end-products and the activities of α-amylase and α-glucosidase. J. Ethnopharmacol. 2022, 285, 114902. [Google Scholar] [CrossRef]
- Jo, Y.H.; Lee, S.; Yeon, S.W.; Turk, A.; Lee, J.H.; Hong, S.M.; Han, Y.K.; Lee, K.Y.; Hwang, B.Y.; Kim, S.Y.; et al. Anti-diabetic potential of Masclura tricuspidata leaves: Prenylated isoflavonoids with α-glucosidase inhibitory and anti-glycation activity. Bioorg. Chem. 2021, 114, 105098. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Park, J.H.; Chung, N.; Lee, H.S. Inhibitory Potential of Constituents from Osmanthus fragrans and Structural Analogues against Advanced Glycation End Products, α-Amylase, α-Glucosidase, and Oxidative Stress. Sci. Rep. 2017, 7, 45746. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, H.; Wang, G.; Liu, W.; Seeram, N.P.; Mu, Y.; Xu, Y.; Huang, X.; Li, L. Phenolics from Eugenia jambolana seeds with advanced glycation endproduct formation and alpha-glucosidase inhibitory activities. Food Funct. 2018, 9, 4246–4254. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Kashegari, A.T.; Shalabi, A.A.; Darwish, K.M.; El-Halawany, A.M.; Algandaby, M.M.; Ibrahim, S.R.M.; Mohamed, G.A.; Abdel-Naim, A.B.; Koshak, A.E.; et al. Phenolics from Chrozophora oblongifolia Aerial Parts as Inhibitors of α-Glucosidases and Advanced Glycation End Products: In-Vitro Assessment, Molecular Docking and Dynamics Studies. Biology 2022, 11, 762. [Google Scholar] [CrossRef]
- Miyata, T.; van Ypersele de Strihou, C.; Ueda, Y.; Ichimori, K.; Inagi, R.; Onogi, H.; Ishikawa, N.; Nangaku, M.; Kurokawa, K. Angiotensin II receptor antagonists and angiotensin-converting enzyme inhibitors lower in vitro the formation of advanced glycation end products: Biochemical mechanisms. J. Am. Soc. Nephrol. 2002, 13, 2478–2487. [Google Scholar] [CrossRef]
- Nangaku, M.; Miyata, T.; Sada, T.; Mizuno, M.; Inagi, R.; Ueda, Y.; Ishikawa, N.; Yuzawa, H.; Koike, H.; van Ypersele de Strihou, C.; et al. Anti-hypertensive agents inhibit in vivo the formation of advanced glycation end products and improve renal damage in a type 2 diabetic nephropathy rat model. J. Am. Soc. Nephrol. 2003, 14, 1212–1222. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.M.; Cooper, M.E.; Thallas, V.; Burns, W.C.; Thomas, M.C.; Brammar, G.C.; Lee, F.; Grant, S.L.; Burrell, L.M.; Jerums, G.; et al. Reduction of the accumulation of advanced glycation end products by ACE inhibition in experimental diabetic nephropathy. Diabetes 2002, 51, 3274–3782. [Google Scholar] [CrossRef]
- Forbes, J.M.; Thorpe, S.R.; Thallas-Bonke, V.; Pete, J.; Thomas, M.C.; Deemer, E.R.; Bassal, S.; El-Osta, A.; Long, D.M.; Panagiotopoulos, S.; et al. Modulation of soluble receptor for advanced glycation end products by angiotensin-converting enzyme-1 inhibition in diabetic nephropathy. J. Am. Soc. Nephrol. 2005, 16, 2363–2372. [Google Scholar] [CrossRef]
- Komiya, N.; Hirose, H.; Saisho, Y.; Saito, I.; Itoh, H. Effects of 12-month valsartan therapy on glycation and oxidative stress markers in type 2 diabetic subjects with hypertension. Int. Heart J. 2008, 49, 681–689. [Google Scholar] [CrossRef]
- Wangm, B.; Wang, F.; Zhang, Y.; Zhao, S.H.; Zhao, W.J.; Yan, S.L.; Wang, Y.G. Effects of RAS inhibitors on diabetic retinopathy: A systematic review and meta-analysis. Lancet Diabetes Endocrinol. 2015, 3, 263–274. [Google Scholar] [CrossRef]
- Sun, P.; Xu, N.; Li, Y.; Han, Y. Destruction of the blood-retina barrier in diabetic retinopathy depends on angiotensin-converting enzyme-mediated TGF-β1/Smad signaling pathway activation. Int. Immunopharmacol. 2020, 85, 106686. [Google Scholar] [CrossRef]
- Nakamura, T.; Sato, E.; Fujiwara, N.; Kawagoe, Y.; Koide, H.; Ueda, Y.; Takeuchi, M.; Yamagishi, S. Calcium channel blocker inhibition of AGE and RAGE axis limits renal injury in nondiabetic patients with stage I or II chronic kidney disease. Clin. Cardiol. 2011, 34, 372–377. [Google Scholar] [CrossRef]
- Santilli, F.; Bucciarelli, L.; Noto, D.; Cefalù, A.B.; Davì, V.; Ferrante, E.; Pettinella, C.; Averna, M.R.; Ciabattoni, G.; Davì, G. Decreased plasma soluble RAGE in patients with hypercholesterolemia: Effects of statins. Free Radic. Biol. Med. 2007, 43, 1255–1262. [Google Scholar] [CrossRef]
- Melin, E.O.; Dereke, J.; Hillman, M. Higher levels of the soluble receptor for advanced glycation end products and lower levels of the extracellular newly identified receptor for advanced glycation end products were associated with lipid-lowering drugs in patients with type 1 diabetes: A comparative cross-sectional study. Lipids Health Dis. 2020, 19, 223. [Google Scholar]
- Tanaka, Y.; Uchino, H.; Shimizu, T.; Yoshii, H.; Niwa, M.; Ohmura, C.; Mitsuhashi, N.; Onuma, T.; Kawamori, R. Effect of metformin on advanced glycation endproduct formation and peripheral nerve function in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 1999, 376, 17–22. [Google Scholar] [CrossRef]
- Demaré, S.; Kothari, A.; Calcutt, N.A.; Fernyhough, P. Metformin as a potential therapeutic for neurological disease: Mobilizing AMPK to repair the nervous system. Expert Rev. Neurother. 2021, 21, 45–63. [Google Scholar] [CrossRef]
- Zhao, M.; Li, X.W.; Chen, Z.; Hao, F.; Tao, S.X.; Yu, H.Y.; Cheng, R.; Liu, H. Neuro-Protective Role of Metformin in Patients with Acute Stroke and Type 2 Diabetes Mellitus via AMPK/Mammalian Target of Rapamycin (mTOR) Signaling Pathway and Oxidative Stress. Med. Sci. Monit. 2019, 25, 2186–2194. [Google Scholar] [CrossRef]
- Campbell, J.M.; Stephenson, M.D.; de Courten, B.; Chapman, I.; Bellman, S.M.; Aromataris, E. Metformin Use Associated with Reduced Risk of Dementia in Patients with Diabetes: A Systematic Review and Meta-Analysis. J. Alzheimers Dis. 2018, 65, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Alomar, S.Y.; Barakat, B.M.; Eldosoky, M.; Atef, H.; Mohamed, A.S.; Elhawary, R.; El-Shafey, M.; Youssef, A.M.; Elkazaz, A.Y.; Gabr, A.M.; et al. Protective effect of metformin on rat diabetic retinopathy involves suppression of toll-like receptor 4/nuclear factor-k B expression and glutamate excitotoxicity. Int. Immunopharmacol. 2021, 90, 107193. [Google Scholar] [CrossRef]
- Fan, Y.P.; Wu, C.T.; Lin, J.L.; Hsiung, C.A.; Liu, H.Y.; Lai, J.N.; Yang, C.C. Metformin Treatment Is Associated with a Decreased Risk of Nonproliferative Diabetic Retinopathy in Patients with Type 2 Diabetes Mellitus: A Population-Based Cohort Study. J. Diabetes Res. 2020, 2020, 9161039. [Google Scholar] [CrossRef]
- Peng, X.; Cheng, K.W.; Ma, J.; Chen, B.; Ho, C.T.; Lo, C.; Chen, F.; Wang, M. Cinnamon bark proanthocyanidins as reactive carbonyl scavengers to prevent the formation of advanced glycation endproducts. J. Agric. Food Chem. 2008, 56, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Ma, J.; Chao, J.; Sun, Z.; Chang, R.C.; Tse, I.; Li, E.T.; Chen, F.; Wang, M. Beneficial effects of cinnamon proanthocyanidins on the formation of specific advanced glycation endproducts and methylglyoxal-induced impairment on glucose consumption. J. Agric. Food Chem. 2010, 58, 6692–6696. [Google Scholar] [CrossRef] [PubMed]
- Kommula, S.R.; Vadakattu, S.S.; Myadara, S.; Putcha, U.K.; Tamanam, R.R.; Palla, S. Cinnamon Attenuated Long-Term IGT-Induced Retinal Abnormalities via Regulation of Glucose Homeostasis in Neonatal Streptozotocin Induced Rat Model. Indian J. Clin. Biochem. 2020, 35, 442–450. [Google Scholar] [CrossRef]
- Thomson, M.; Al-Qattan, K.K.; Js, D.; Ali, M. Anti-diabetic and anti-oxidant potential of aged garlic extract (AGE) in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2016, 16, 17. [Google Scholar] [CrossRef]
- Ziamajidi, N.; Nasiri, A.; Abbasalipourkabir, R.; Sadeghi Moheb, S. Effects of garlic extract on TNF-α expression and oxidative stress status in the kidneys of rats with STZ + nicotinamide-induced diabetes. Pharm. Biol. 2017, 55, 526–531. [Google Scholar] [CrossRef]
- Afarid, M.; Sadeghi, E.; Johari, M.; Namvar, E.; Sanie-Jahromi, F. Evaluation of the Effect of Garlic Tablet as a Complementary Treatment for Patients with Diabetic Retinopathy. J. Diabetes Res. 2022, 2022, 6620661. [Google Scholar] [CrossRef]
- Gugliucci, A.; Bastos, D.H.; Schulze, J.; Souza, M.F. Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of AGE generation by methylglyoxal in model proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef]
- Lunceford, N.; Gugliucci, A. Ilex paraguariensis extracts inhibit AGE formation more efficiently than green tea. Fitoterapia 2005, 76, 419–427. [Google Scholar] [CrossRef]
- Silva, K.C.; Rosales, M.A.; Hamassaki, D.E.; Saito, K.C.; Faria, A.M.; Ribeiro, P.A.; Faria, J.B.; Faria, J.M. Green tea is neuroprotective in diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1325–1336. [Google Scholar] [CrossRef]
- Ma, Q.; Chen, D.; Sun, H.P.; Yan, N.; Xu, Y.; Pan, C.W. Regular Chinese Green Tea Consumption is Protective for Diabetic Retinopathy: A Clinic-Based Case-Control Study. J. Diabetes Res. 2015, 2015, 231570. [Google Scholar] [CrossRef]
- Xu, C.; Bi, M.; Jin, X.; Zhu, M.; Wang, G.; Zhao, P.; Qin, X.; Xu, X.; Sun, X.; Ji, N.; et al. Long-Term Tea Consumption Is Associated with Reduced Risk of Diabetic Retinopathy: A Cross-Sectional Survey among Elderly Chinese from Rural Communities. J. Diabetes Res. 2020, 2020, 1860452. [Google Scholar] [CrossRef]
- Takeuchi, M.; Yamagishi, S. Possible involvement of advanced glycation end-products (AGEs) in the pathogenesis of Alzheimer’s disease. Curr. Pharm. Des. 2008, 14, 973–978. [Google Scholar] [CrossRef]
- Ko, S.Y.; Ko, H.A.; Chu, K.H.; Shieh, T.M.; Chi, T.C.; Chen, H.I.; Chang, W.C.; Chang, S.S. The Possible Mechanism of Advanced Glycation End Products (AGEs) for Alzheimer’s Disease. PLoS ONE 2015, 10, e0143345. [Google Scholar] [CrossRef]
- Shimizu, Y.; Harashima, A.; Munesue, S.; Oishi, M.; Hattori, T.; Hori, O.; Kitao, Y.; Yamamoto, H.; Leerach, N.; Nakada, M.; et al. Neuroprotective Effects of Endogenous Secretory Receptor for Advanced Glycation End-products in Brain Ischemia. Aging Dis. 2020, 11, 547–558. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, X.; Tuo, M.; Ma, J.; Xie, A. RAGE and its emerging role in the pathogenesis of Parkinson’s disease. Neurosci. Lett. 2018, 672, 65–69. [Google Scholar] [CrossRef]
- Pillar, S.; Moisseiev, E.; Sokolovska, J.; Grzybowski, A. Recent Developments in Diabetic Retinal Neurodegeneration: A Literature Review. J. Diabetes Res. 2020, 2020, 5728674. [Google Scholar] [CrossRef]
- Lai, M.C.; Liu, W.Y.; Liou, S.S.; Liu, I.M. The Citrus Flavonoid Hesperetin Encounters Diabetes-Mediated Alzheimer-Type Neuropathologic Changes through Relieving Advanced Glycation End-Products Inducing Endoplasmic Reticulum Stress. Nutrients 2022, 14, 745. [Google Scholar] [CrossRef]
- Lai, M.C.; Liu, W.Y.; Liou, S.S.; Liu, I.M. Diosmetin Targeted at Peroxisome Proliferator-Activated Receptor Gamma Alleviates Advanced Glycation End Products Induced Neuronal Injury. Nutrients 2022, 14, 2248. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Peng, C.H.; Chyau, C.C.; Lin, Y.C.; Wang, H.E.; Peng, R.Y. Low-density lipoprotein, collagen, and thrombin models reveal that Rosemarinus officinalis L. exhibits potent antiglycative effects. J. Agric. Food Chem. 2007, 55, 2884–2891. [Google Scholar] [CrossRef] [PubMed]
- Alrubaye, A.; Motovali-Bashi, M.; Miroliaei, M. Rosmarinic acid inhibits DNA glycation and modulates the expression of Akt1 and Akt3 partially in the hippocampus of diabetic rats. Sci. Rep. 2021, 11, 20605. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.C.; Moreira, C.P.S.; Castro, B.F.M.; Cotta, O.A.L.; Silva, L.M.; Fulgêncio, G.O.; Silva-Cunha, A.; Fialho, S.L. Rosmarinic Acid Intravitreal Implants: A New Therapeutic Approach for Ocular Neovascularization. Planta Med. 2020, 86, 1286–1297. [Google Scholar] [CrossRef]
Figure 1.
Initial step of the Maillard reaction. In the first step, a protein reacts with an amino acid, which results in the production of a Schiff base. The Schiff base is converted into an Amadori product by the Amadori rearrangement through a protonated imine and 1,2-enaminol formation. S: reducing sugar, P: protein.
Figure 1.
Initial step of the Maillard reaction. In the first step, a protein reacts with an amino acid, which results in the production of a Schiff base. The Schiff base is converted into an Amadori product by the Amadori rearrangement through a protonated imine and 1,2-enaminol formation. S: reducing sugar, P: protein.

Figure 2.
Hypothetical schemes of the final step of AGE formation. Carboxymethyllysine (CML) is synthesized from an Amadori compound or glyoxal [23]. Carboxyethyllysine (CEL) is made from methylglyoxal [24]. Glyoxal-derived lysine dimer (GOLD), methylglyoxal-derived lysine dimer (MOLD), and 3-DG-derived lysine dimer (DOLD) have cross-links between lysine residues and glyoxal, methylglyoxal or 3-DG [25]. Only a small percentage of AGEs in the body have been identified. Glyceraldehyde- and glycolaldehyde-derived AGEs may be synthesized faster than Amadori compound-derived AGEs and more toxic than other types of AGEs.
Figure 2.
Hypothetical schemes of the final step of AGE formation. Carboxymethyllysine (CML) is synthesized from an Amadori compound or glyoxal [23]. Carboxyethyllysine (CEL) is made from methylglyoxal [24]. Glyoxal-derived lysine dimer (GOLD), methylglyoxal-derived lysine dimer (MOLD), and 3-DG-derived lysine dimer (DOLD) have cross-links between lysine residues and glyoxal, methylglyoxal or 3-DG [25]. Only a small percentage of AGEs in the body have been identified. Glyceraldehyde- and glycolaldehyde-derived AGEs may be synthesized faster than Amadori compound-derived AGEs and more toxic than other types of AGEs.

Figure 3.
Scheme of the polyol pathway. Under normal conditions (upper figure), glucose is taken in by the retinal cells via sodium-glucose cotransporter (SGLT) or glucose transporter (GLUT). Glucose is converted to pyruvate by the metabolic glycolysis pathway. This process releases free energy to synthesize the high-energy molecule adenosine triphosphate (ATP) in the final step. In the diabetic condition, excessive uptake of glucose activates a minor cellular pathway, the polyol pathway (lower figure). The activated polyol pathway causes an increase in oxidative stress, osmotic stress, and AGE formation in diabetic retinas. G: glucose, G6P: glucose-6-phosphate, F6P: fructose-6-phosphate, GA3P: glyceraldehyde-3-P, NADP: nicotinamide adenine dinucleotide phosphate (NADP+; oxidized form, NADPH; reduced form), DAG: diacylglycerol, PKC: protein kinase C, AGEs: advanced glycation end products.
Figure 3.
Scheme of the polyol pathway. Under normal conditions (upper figure), glucose is taken in by the retinal cells via sodium-glucose cotransporter (SGLT) or glucose transporter (GLUT). Glucose is converted to pyruvate by the metabolic glycolysis pathway. This process releases free energy to synthesize the high-energy molecule adenosine triphosphate (ATP) in the final step. In the diabetic condition, excessive uptake of glucose activates a minor cellular pathway, the polyol pathway (lower figure). The activated polyol pathway causes an increase in oxidative stress, osmotic stress, and AGE formation in diabetic retinas. G: glucose, G6P: glucose-6-phosphate, F6P: fructose-6-phosphate, GA3P: glyceraldehyde-3-P, NADP: nicotinamide adenine dinucleotide phosphate (NADP+; oxidized form, NADPH; reduced form), DAG: diacylglycerol, PKC: protein kinase C, AGEs: advanced glycation end products.

Figure 4.
Hypothetic scheme of the pathophysiology of AGEs in diabetic retinas. Chronic hyperglycemia facilitates AGE production. Oxidative stress is induced by the AGE-RAGE axis, but Amadori compounds become the source of ROS production. Thus, the RAGE-independent pathways can be involved in increasing oxidative stress in diabetic retinas.
Figure 4.
Hypothetic scheme of the pathophysiology of AGEs in diabetic retinas. Chronic hyperglycemia facilitates AGE production. Oxidative stress is induced by the AGE-RAGE axis, but Amadori compounds become the source of ROS production. Thus, the RAGE-independent pathways can be involved in increasing oxidative stress in diabetic retinas.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Oshitari, T. Advanced Glycation End-Products and Diabetic Neuropathy of the Retina. Int. J. Mol. Sci. 2023, 24, 2927. https://doi.org/10.3390/ijms24032927
AMA Style
Oshitari T. Advanced Glycation End-Products and Diabetic Neuropathy of the Retina. International Journal of Molecular Sciences. 2023; 24(3):2927. https://doi.org/10.3390/ijms24032927
Chicago/Turabian StyleOshitari, Toshiyuki. 2023. "Advanced Glycation End-Products and Diabetic Neuropathy of the Retina" International Journal of Molecular Sciences 24, no. 3: 2927. https://doi.org/10.3390/ijms24032927
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.