
Tryptophan and Substance Abuse: Mechanisms and Impact
by
 , and
, and
Majid Davidson
1,2 ,
,
Niloufar Rashidi
1,
Md Kamal Hossain
1,
Ali Raza
3,4,
Kulmira Nurgali
1,2,5,* and
Vasso Apostolopoulos
1,6,*
1
Institute for Health and Sport, Victoria University, Melbourne, VIC 3021, Australia
2
Regenerative Medicine and Stem Cells Program, Australian Institute of Musculoskeletal Science (AIMSS), Melbourne, VIC 3021, Australia
3
Fiona Elsey Cancer Research Institute, Ballarat, VIC 3353, Australia
4
Federation University, Ballarat, VIC 3353, Australia
5
Department of Medicine Western Health, Faculty of Medicine, Dentistry and Health Sciences, University of Melbourne, Melbourne, VIC 3021, Australia
6
Immunology Program, Australian Institute of Musculoskeletal Science (AIMSS), Melbourne, VIC 3021, Australia
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(3), 2737; https://doi.org/10.3390/ijms24032737
Submission received: 4 January 2023
/
Revised: 22 January 2023
/
Accepted: 28 January 2023
/
Published: 1 February 2023
(This article belongs to the Special Issue Mechanisms and Interventions for Neurological and Psychological Disorders 2.0)
Abstract
:Addiction, the continuous misuse of addictive material, causes long-term dysfunction in the neurological system. It substantially affects the control strength of reward, memory, and motivation. Addictive substances (alcohol, marijuana, caffeine, heroin, methamphetamine (METH), and nicotine) are highly active central nervous stimulants. Addiction leads to severe health issues, including cardiovascular diseases, serious infections, and pulmonary/dental diseases. Drug dependence may result in unfavorable cognitive impairments that can continue during abstinence and negatively influence recovery performance. Although addiction is a critical global health challenge with numerous consequences and complications, currently, there are no efficient options for treating drug addiction, particularly METH. Currently, novel treatment approaches such as psychological contingency management, cognitive behavioral therapy, and motivational enhancement strategies are of great interest. Herein, we evaluate the devastating impacts of different addictive substances/drugs on users′ mental health and the role of tryptophan in alleviating unfavorable side effects. The tryptophan metabolites in the mammalian brain and their potential to treat compulsive abuse of addictive substances are investigated by assessing the functional effects of addictive substances on tryptophan. Future perspectives on developing promising modalities to treat addiction and the role of tryptophan and its metabolites to alleviate drug dependency are discussed.
1. Introduction
Drug addiction is identified as a compulsive relapsing disorder that has devastating effects on the brain and behavior of users and results in a disability to control the drug-seeking tendency and use, notwithstanding unfavorable consequences [1]. DA is initiated via experimental utilization of a recreational drug in society or exposure to a prescribed medicine from a friend [2,3]. Substances including methamphetamine (METH), alcohol, marijuana, heroin, and nicotine are the most commonly used addictive drugs that may have disparate detrimental side effects on users. Numerous short-term and long-term side effects such as insomnia, drowsiness, increased blood pressure, uncontrolled movement, and rapid heart rate are the dire and unfavorable consequences of disparate addictive drug abuse [4,5].
In recent decades, alcohol addiction has been a global crisis that accounts for almost 5% of the disease burden [6]. Based on the statistics, alcohol is the most prevalently abused substance in the United States and is responsible for 3.5% of annual deaths due to its detrimental and deleterious effects, such as liver cirrhosis and neurotoxicity [7]. Therefore, developing cost-effective and promising drugs to treat alcohol addiction is crucial, as only a small proportion of patients can achieve effective remission. Marijuana (whose street names include dope, pot, grass, weed, and hashish) manifests as an addictive drug extracted from the cannabis plant. Tetrahydrocannabinol is the active ingredient in marijuana, which is the main reason for its potential for dependency. This addictive substance can significantly change perception due to its psychoactive characteristics. Depending on the mode of administration, marijuana can produce various neuropsychological disorders such as hallucinations, abnormal happiness, and difficulty concentrating [8]. Heroin, another commonly used addictive substance produced from morphine [9,10], is highly abused among people from various social cultures. Heroin targets the brain and receptors on cells, which increases the heart rate and causes breathing and sleeping issues [11,12]. In addition, dry mouth, drowsiness, constipation, and depression are common side effects [13].
The World Health Organization predicts that almost one-third of the global adult population smokes tobacco, accounting for many smoking-related deaths [14,15]. Nicotine (one of the foremost effective ingredients in tobacco) is a naturally manufactured alkaloid used as an anxiolytic stimulant [16]. Similar to other psychostimulant agents, nicotine increases extracellular dopamine concentrations in the nucleus accumbens, which can cause neuropsychological/behavioral alterations in users [17]. Table 1 provides information regarding common addictive substances.
Despite the availability of current treatment options, such as behavioral therapy and medication-assisted treatment, they are often ineffective for addressing the underlying mechanisms of addiction [37,38]. These treatments typically involve behavioral therapies, medications, and support groups, but they do not address the underlying neurobiological mechanisms of addiction. Understanding the mechanisms of addiction and its physiological impacts can provide an opportunity for developing a more efficient treatment for this global health issue. Tryptophan is an essential amino acid that plays vital roles in several critical biological processes, including the regulation of mood and behavior [39]. Addiction leads to an imbalance in the levels of tryptophan and its metabolites, which can result in serious health issues [40]. Understanding the connection between tryptophan and addiction may hold potential for developing novel therapeutic approaches for addiction treatment.
In this review article, the detrimental effects of addictive substances, specifically METH, on human mental and psychological dependency are evaluated. The role of METH and other addictive substances on mental and psychological dependency in humans is presented, along with a description of tryptophan metabolites in the mammalian brain and their potential as new treatment options for controlling compulsive substance abuse. The functional effects of METH on tryptophan and its disparate metabolites, including serotonin 5-HT (5-hydroxytryptophan), melatonin, kynurenine, and reactive oxygen and nitrogen species, as well as the formation and structure of each, are examined. The understanding of the connection between tryptophan metabolites and addiction provides a valuable opportunity to investigate future perspectives on the development of promising modalities for addiction treatment.
2. General Effects of Addiction on the Body
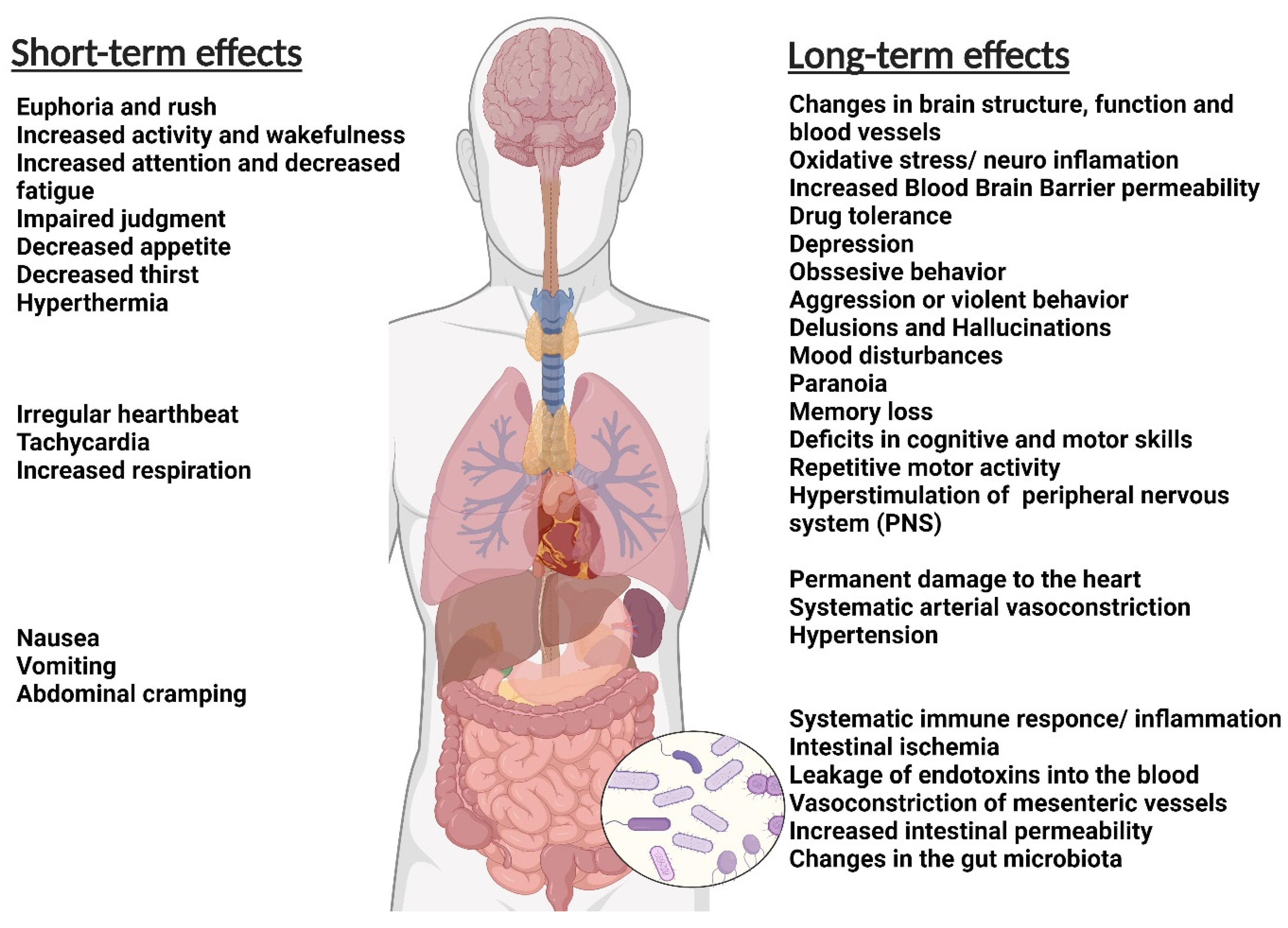
The abuser of disparate types of addictive materials usually suffers from one or more health-related challenges (Figure 1). Tobacco smoke may increase the risk of cancers, heart failure, brain stroke, lung diseases, and chronic obstructive pulmonary diseases such as emphysema and bronchitis. Smoking also increases the risk of tuberculosis, certain eye diseases, and autoimmune system diseases such as rheumatoid arthritis [41,42].
Marijuana is known as the most commonly abused addictive drug in the United States, and its use is expanding substantially in all adult age groups of both sexes. Abusing these addictive substances can significantly enhance the risk of permanent intelligence quotient (IQ) loss by eight points. Moreover, different studies have corroborated the existence of a close relationship between marijuana abuse and psychotic disorders such as depression, anxiety, and suicidal ideation [43,44].
Likewise, the unusual use of alcohol is increasing worldwide and is regarded as one of the main reasons for chronic liver diseases [45]. Long-term abuse of alcohol may also increase the risk of chronic diseases such as hypertension, decreased immune system functioning, digestive problems, and cancer.
Caffeine, a stimulant found in coffee, tea, energy drinks, and various medications, acts on the central nervous system by binding to adenosine receptors, leading to increased alertness and decreased fatigue [46]. While moderate caffeine consumption may provide some health benefits, excessive caffeine intake can lead to various adverse health outcomes, such as insomnia, anxiety, high blood pressure, and an increased risk of certain types of cancer [47]. Long-term heavy caffeine use can also lead to tolerance, withdrawal symptoms, and difficulty reducing or stopping caffeine consumption, ultimately leading to addiction.
Cocaine is one of the most addictive substances, and its abuse may cause the emergence of many unfavorable and dangerous side effects such as mood disorders, cardiovascular diseases, weight loss, an increase in the potential risk of human immunodeficiency virus infection, and memory loss [48].
METH is an amine drug that is abused due to its sympathomimetic effects and is taken via inhalation, ingestion, injection, or smoking [49,50]. METH is a weak lipophilic base that diffuses across the plasmalemma of the presynaptic neurons and vesicular membranes [51]. METH enters the monoaminergic terminals of neurons due to its structural similarities to monoamine hormones (serotonin, adrenaline, noradrenaline, dopamine, and melatonin). It then accumulates in the synaptic vesicles through the action of vesicular monoamine transporters. It causes an initial outward flow of hormones along with the prevention of the reuptake of neurotransmitters and their metabolism by enzymes such as monoamine oxidase [52,53,54]. Following the initial METH abuse, an increase in the levels of monoamines occurs [51], leading to an elevation in intra-synaptic levels of monoamine [55]. This increase results in hallucinations, euphoria, and anorexia [49], as well as risk-taking sexual behaviors, which raises serious public health and safety concerns [56]. Repetitive and escalating doses of METH result in decreased monoamine levels due to a reduction in monoamine transporter binding sites as well as in the activity of synthetic enzymes [57]. The abuse of addictive drugs such as METH confuses addicts on whether reward-specific circuits or adaptive natural reward circuits are activated in their brains. As such, addicted individuals feel that drug abuse is part of their natural biological needs [58]. However, studies have shown that multiple high-dose administrations of METH result in drug resistance in the body, a phenomenon known as drug tolerance [59]. Levels of the tryptophan metabolite, which is an amine precursor to serotonin, melatonin, kynurenine, and quinolinine pathways, have been noted to be affected by METH addiction [60]. Decreased levels of tryptophan have also been found to take place in suicidal adolescent patients [61]. Clinically, it is known that METH toxicity affects nearly every organ system in the body, with the most considerable damage occurring to the CNS [62]. This includes metabolic compromise, oxidative stress, and inflammation of neurons resulting in neuronal death [63].
3. Tryptophan and Its Metabolites
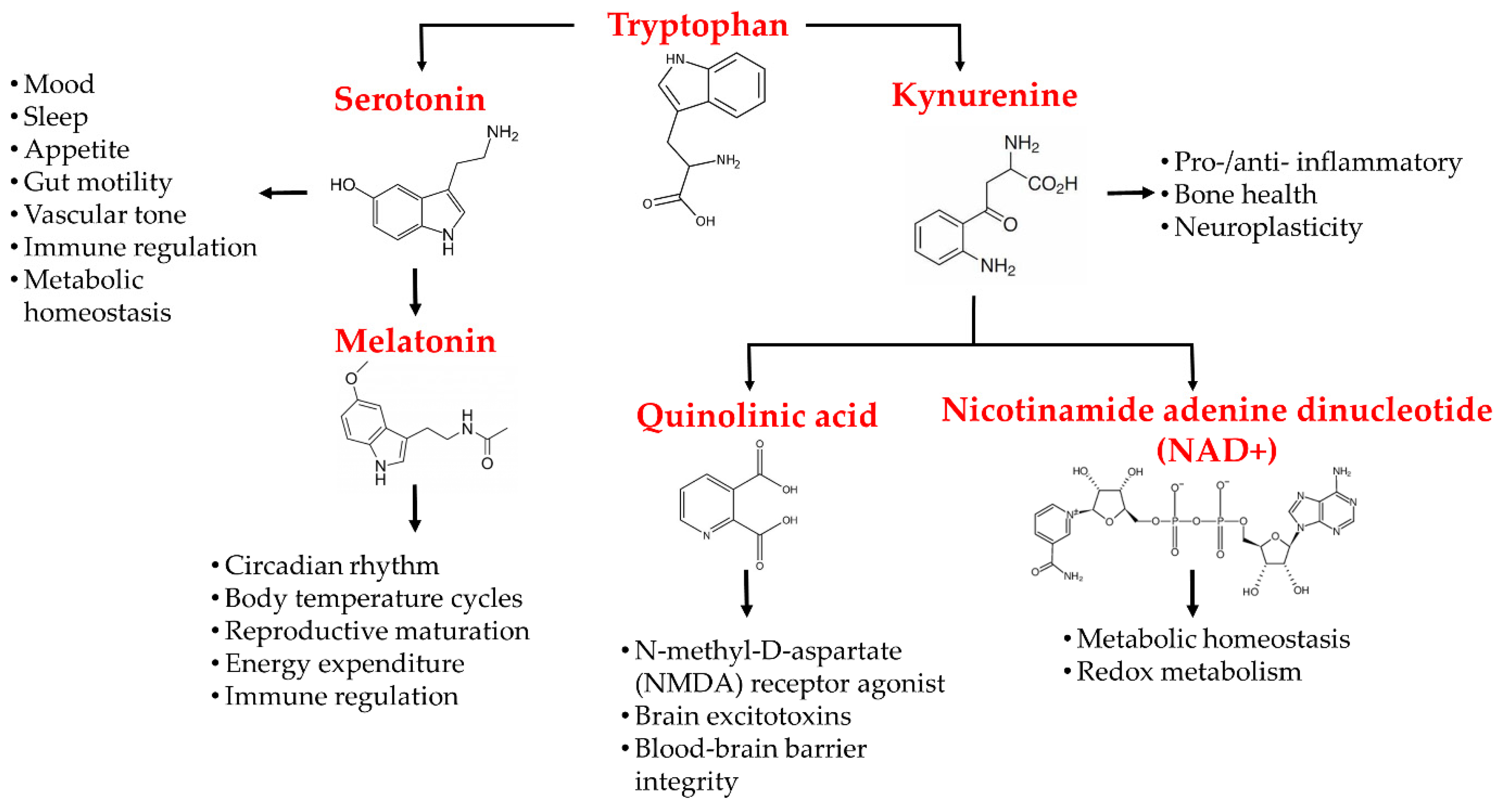
Tryptophan is an important amino acid that plays a crucial role in various biological processes, including in an infant′s growth, muscle development, enzyme function, and neurotransmitter regulation [64,65]. Tryptophan is a precursor to the metabolites serotonin, melatonin, and kynurenine [66]. Most of the daily intake of tryptophan oxidizes down the kynurenine pathway, whilst the rest degrades via serotonin pathways [67]. Tryptophan is metabolized into serotonin and melatonin via the generation of 5-hydroxytryptophan, which includes human peripheral tryptophan hydroxylase 1 (hpTrpH1) or human neural tryptophan hydroxylase 2 (hnTrpH2), depending on whether metabolism occurs in peripheral entero-chromatic cells or neural cells of the CNS. 5-hydroxytryptophan is then metabolized into serotonin and, further down, into melatonin via the action of different enzymes [66,67]. The kynurenine pathway either involves the generation of kynurenic acid (KA) or the formation of 3-hydroxy-anthranillic acid, further leading to quinolinic acid (QA). These components, QA and KA, are believed to have noteworthy impacts on CNS neurons. QA possesses the potential for application as a neurotoxin, while KA is a neuroprotectant. 3-hydroxykynurenine is the third kynurenine metabolite that may create free radicals and aggravate neuronal damage [68].
The kynurenine pathway plays a critical role in regulating immune responses and has the potential to be a target for various inflammatory and neurological disorders by modulating the immune response and regulating the production of kynurenine derivatives [69]. Due to tryptophan catabolism′s incredible impact on increasing immune suppression, disparate small-molecule inhibitors have been developed and tested in clinical trials [70,71,72]. Inflammatory cytokines and chemokines are key mediators of the immune response, and their release can lead to a cascade of effects that can disturb the immune system and cause a corresponding increase in kynurenine production [73,74]. An imbalance in kynurenine production can shift the balance of tryptophan metabolism, resulting in the accumulation of pro-inflammatory and neurotoxic kynurenine derivatives [75]. This can lead to a range of negative effects, including increased susceptibility to infections, chronic inflammation, and neurodegeneration. Furthermore, research has shown that the kynurenine pathway is closely linked to various psychiatric and neurological disorders, including depression, anxiety, schizophrenia, and Alzheimer′s disease [76].
The kynurenine pathway involves 95% of tryptophan degradation in the liver via two rate-limiting enzymes: indoleamine 2,3-dioxygenase (IDO) and tryptophan 2,3-deoxygenase (TDO) [77]. These enzymes, TDO and IDO, control tryptophan degradation through the kynurenine pathway. TDO is primarily expressed in the liver and is responsible for the initial degradation of tryptophan. This enzyme converts tryptophan into kynurenine, which is then further metabolized by IDO. IDO is expressed in various tissues, including the immune cells, and plays a key role in the regulation of immune responses. TDO usually leads to nicotinamide dinucleotide (NAD+) in the liver only. However, IDO is known to carry the same extra-hepatical reaction but in much smaller amounts than the enzyme TDO [67]. Additionally, the combination of IDO1 inhibitors together with immune checkpoint inhibitors, such as anti-programmed cell death 1 (PD1) and anti-programmed cell death ligand 1 (PD-L1), have opened new avenues towards increasing the efficacy of immuno-oncology in cancer treatment [78,79,80].
The mammalian brain contains from nanomolar to low micromolar concentrations of kynurenine pathway metabolites [81]. Acute stress causes an increase in tryptophan metabolism to kynurenine in mice brains and plasma [82]. In fact, increased IDO enzyme activity and IdoI-v1 expression have been noted in the hypothalamus and frontal cortex of the brain, specifically in astrocytes. Ferulic acid, a plant-derived phenol compound with anti-inflammatory effects, has been shown to have suppressing effects on lipopolysaccharide-induced IDO messenger RNA (mRNA) expression in mouse microglial cells [83]. In addition, ferulic acid suppresses the phosphorylation of p38 MAPK and the nuclear translocation of NF-κB [83]. Figure 2 schematically illustrates the tryptophan metabolites, their pathways, and their effects on the CNS.
4. Tryptophan and Addiction: METH Addiction as an Example
Tryptophan is an essential amino acid that can only be obtained via dietary intake, and a diet with low levels of tryptophan is related to malnutrition or poor diet habits [67]. Short-term and long-term abuse of alcohol and its associated withdrawal is closely related to tryptophan metabolism/disposition in humans and animals. Various studies have proved that the activity of the rate-limiting enzyme of tryptophan degradation (liver tryptophan pyrrolase) increases substantially during acute alcohol abuse and during withdrawal [84]. The occurrence of this alteration causes a significant change in the synthesis process of brain serotonin and the appropriate availability of tryptophan to the brain [84]. It has been reported that serotonin can be considered the prominent tryptophan metabolite related to psychiatric comorbidity in abstinent cocaine-addicted patients [40]. However, the non-existence of efficacious treatments following the great relapse rate among cocaine abusers remains a major health problem.
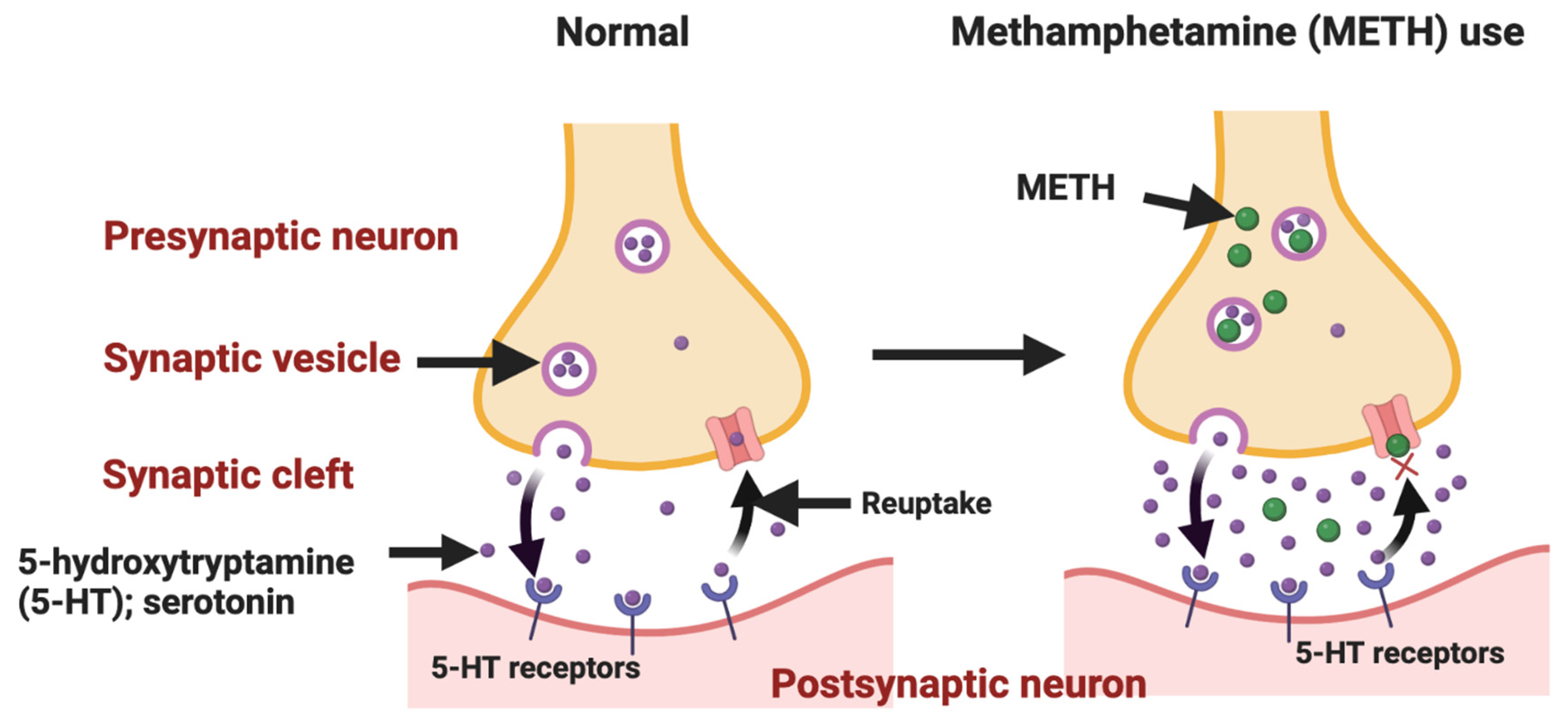
METH (under the street names ice, crystal, glass, or kryptonite) is a potent synthetic central nervous system (CNS) drug that has been placed second in rank of application in the United States, Asia, and Oceania after cannabis [85,86]. METH is often sold in powder or crystal form and is administered via intranasal sniffing, oral ingestion, pulmonary inhalation, and injection. METH addiction leads to serious side effects on disparate organs within the human body, such as subjective euphoria/arousal, chronic depression, and psychomotor activation [87,88]. Poor mental health and social disability (i.e., unemployment and economic hardship) can be considered the most immediate adverse effects of METH dependence, which significantly decreases the quality of life among users [89,90]. METH dependence is a major global public health challenge due to its medical, psychiatric, cognitive, and socioeconomic consequences [91,92]. Figure 3 represents the METH-induced serotonin pathway in the CNS.
Weekly stimulant use of recreational drugs such as METH, cocaine, etc., results in decreased levels of tryptophan. Suicidal adolescent patients suffering from major depressive disorder have decreased levels of circulating tryptophan [61], and suicidality/depression is considered one of the symptoms of METH addiction. Overall, research has shown that METH abuse affects the CNS, the spleen, the gut, and the liver [60]. Damage to these organ systems can also adversely affect tryptophan metabolism pathways, causing an imbalance in the levels of its metabolites. The effect of METH on tryptophan and its metabolites and the associated consequences are briefly discussed below.
4.1. Serotonin (5-hydroxytryptophan)
Hydroxylation of L-tryptophan leads to the formation of L-5-hydroxytryptophan (L-5HT) via the action of tryptophan hydroxylase. Aromatic amino acid de-carboxylate helps convert L-5HT to serotonin 5-HT. Tryptophan hydroxylase 1 (TPH1) is known to be composed of two types of TPH enzymes, and is responsible for 95% of the body’s 5-HT formation in the gut, which can travel to all body organs via the blood except for the brain, as it cannot pass through the blood–brain barrier. TPH2 helps the formation of 5-HT in the brain along with the action of aromatic l-amino acid decarboxylase (AADC) [93]. It has been shown that tryptophan hydroxylase activity increases in the globus pallidus, nucleus accumbens, and the caudate area within 1 h of a single bout of METH. However, long-term multiple METH doses only decreased TPH2 activity in the caudate area but not in the nucleus accumbens and globus pallidus [94]. However, recently, it was noted that TPH2 gene variants are not a factor in vulnerability to METH psychosis/dependence as no genotypic/allelic changes are observed between METH-dependent patients (n = 162) and controls (n = 243) [95].
Further experiments would aid in the understanding of why there is a lack of correlation between long-term and short-term METH use around the activity of TPH2 in the nucleus accumbens and globus pallidus regions of the brain. Despite this, a decrease in TPH2 levels is observed in the caudate area in both short- and long-term METH administration. These studies show that the decrease in TPH2 activity occurs in different brain regions, and it is plausible to argue that a decrease in TPH2 levels in the brain could be responsible for low 5-HT levels. It is essential to keep in mind that other factors/molecular mechanisms could also be responsible for low 5-HT levels in the CNS, such as tolerance-related 5-HT deficits in the brain [59].
4.2. Melatonin
Melatonin is a metabolite of serotonin that is formed via the serotonin pathway through the action of enzymes aralkylamine N-acetyltransferase (AA-NAT) and hydroxyindole-O-methyltransferase (HIOMT). After its formation in the pinealocytes of the pineal gland, it is transferred to the blood and cerebrospinal fluid of the brain, where second messengers of sleep are initiated in order to cause sleepiness [67]. Melatonin is formed through the metabolism of serotonin. Therefore, a decrement in serotonin levels would mean a decrease in melatonin levels in METH addicts [57,96]. As such, serotonin is also known to regulate sleep–wake cycles [67], which would explain the disturbance in the sleep cycles of METH addicts. Overall, amphetamine abuse is known to be linked to disturbances in sleep cycles, with chronic abuse being more detrimental than the usual acute abuse of drug addiction [97]. A study in rhesus macaques showed disrupted wake cycles and poor sleep efficiency after both low and high METH self-administration [98].
4.3. Kynurenine
The metabolism of tryptophan forms kynurenine via the action of the enzymes IDO and TDO [61]. Kynurenine is not known to be neuroactive but can cross the blood–brain barrier to aid in the formation of free radicals producing 3-hydroxyanthranillic acid, 3-hydroxykynurenine, or quinolinic acid. It has also been shown that depressed, suicidal patients have decreased kynurenine plasma levels [61], and suicidality is one of the major symptoms of METH addiction. Pro-inflammatory cytokines are known to further help kynurenine metabolism and cause an increase in the levels of its metabolites [66]. Alterations in the evel of the kynurenine pathway metabolites are known as a major cause of neurotoxicity [99]. Over-activation of the IDO and TDO enzymes is known to cause the degradation of most of the tryptophan along the kynurenine pathway. Thus, this leads to lower levels of serotonin via the upregulation of tryptophan along the kynurenine pathway, which is thought to be one of the reasons for the occurrence of depression [99].
4.4. Kynurenic Acid
The action of kynurenine aminotransferase on kynurenine leads to the formation of kynurenic acid, which takes place in the neurons, astrocytes, and oligodendrocytes of the CNS. Once it is synthesized, it is released into the extracellular spaces to act on its pre- and post-synaptic targets [67]. Physiologically, kynurenic acid has a neuroprotective role as it helps prevent pro-inflammatory cytokines, but its elevated levels are known to be linked to psychosis and cognitive deficits [66]. It also inhibits the alpha-7 nicotinic receptor because the receptor, when activated by its agonist, is known to have cognitive-enhancing effects [100]. Kynurenic acid is also known to stabilize the levels of free radicals within the CNS [66]. D-amphetamine, an enantiomer of amphetamine, when systematically administered to rats of different ages, causes a decrease in the extracellular levels and tissue content of kynurenic acid in the brain within 1 h following D-amphetamine administration [101]. Schizophrenic patients with psychosis and a history of suicide have lower cerebrospinal fluid kynurenic acid concentrations [102]. However, patients with schizophrenia only have relatively higher levels of kynurenic acid levels in cerebrospinal fluid when compared with healthy individuals [66]. This shows that depression/suicidality can lead to lower levels of kynurenic acid. Kynurenic acid is known to act as an antagonist for glutamatergic NMDA receptors. Trihexyphenidyl is known to antagonize the METH-induced reward pathway via the suppression of dopamine release from the mesolimbic area [103]. Present data show that high levels of kynurenic acid can prove to be beneficial in the CNS of METH addicts, but kynurenic acid is found to be low in suicidal depressive and psychotic patients, which are common major symptoms of METH addiction.
4.5. Xanthurenic Acid
The catabolism of the 3-hydroxykynurenine forms xanthurenic acid via the action of kynurenine aminotransferase. It is also considered an analog of kynurenic acid and is thus also known to have neuroprotective functions. It is thought to be localized in the cell body and dendrites of neurons but not in the axons, which is why there are debates on whether xanthurenic acid has a role in neurotransmission in the CNS [104]. However, the upregulation of xanthurenic acid in the CNS leads to a decreased synthesis of neurotransmitters such as serotonin and dopamine [105]. In addition, levels of depression and levels of xanthurenic acid positively correlate with each other in the urine of patients with depression [106]. Some initial studies also show that xanthurenic acid may act as an allosteric agonist for the metabotropic glutamate receptors type 2 and 3, but further research is required to support this finding. Along with xanthurenic acid, they are known to act as inhibitors for vesicular glutamatergic transporters whose immediate action is the prevention of glutamate reuptake from the synaptic vessels [107]. A similar mechanism takes place via METH in the neural synapses of a METH addict′s CNS neurons. The current data show that while xanthurenic acid may have neuroprotective characteristics, its upregulation has similar effects as those seen in METH addicts. Therefore, future research on xanthurenic acid in the context of METH addiction will aid in a better understanding of the neural actions of xanthurenic acid in the CNS.
4.6. 3-hydroxyanthranillic Acid
In the CNS, 3-hydroxyanthranillic acid is formed via the action of kynureninase on anthranilic acid, whereas in the periphery, it is formed from 3-hydroxykynurenine [66]. 3-hydroxyanthranillic acid is a very reactive compound that can either act as an antioxidant or pro-oxidant depending on the redox reactions in the brain. One study showed no notable differences in the plasma levels of 3-hydroxyanthranillic acid between suicidal depressed and healthy control groups [61]. However, a positive correlation between the levels of 3-hydroxyanthranillic acid and choline were noted; choline is responsible for cell membrane breakdown in melancholic depressive patients [66]. Inhibition of kynurenine in the kynurenic acid pathway leads to increased production of 3-hydroxyanthranillic acid [108], and since depressive patients have low levels of kynurenic acid in their brains [102], it is possible that depressive patients have high levels of 3-hydroxyanthranillic acid. Based on this, it can be hypothesized that depression can be caused by METH abuse, and 3-hydroxyanthranillic acid levels would be higher in depressive METH addicts. Further research, however, is required to better understand the relationships between METH addiction, depression, and 3-hydroxyanthranillic acid.
4.7. Quinolinic Acid
Quinolinic acid is formed from 3-hydroxyanthranillic acid via the action of enzyme 3-hydroxyanthranillic acid oxygenase. Quinolinic acid is mainly thought to be produced in the brain by microglial cells and by infiltrating macrophages and is thought to be neurotoxic via several different mechanisms [66]. Quinolinic acid is known to severely impair energy metabolism in the striatum of developing rats via the activation of NMDA receptors [109]. Quinolinic acid has also been known to cause an increase in the release of glutamate while preventing its reuptake from the synaptic vesicles in individuals [110]. Quinolinic levels have been found to be very high in the cerebrospinal fluid of depressed suicide attempters [111]. High levels of quinolinic acid correlate with interleukin (IL)-6, suggesting that high levels of quinolinic acid may be due to inflammatory responses [112]. In addition, quinolinic acid levels are high in the cortical and subcortical regions of psychiatric suicidal patients [113]. In fact, ketamine, an NMDA receptor antagonist, helps overcome suicidal thoughts [114]. However, even though ketamine can act as an antagonist for the NMDA receptor, a balanced ratio of quinolinic acid/kynurenic acid can be more effective in avoiding depression with respect to the NMDA receptor because both the acids can act as an agonist (quinolinic acid) and antagonist (kynurenic acid) for the receptor [111]. D-amphetamine, an enantiomer of amphetamine, is known to cause a decrease in levels of kynurenic acid [101]. As such, it is plausible to argue that a decrease in kynurenic acid would push tryptophan metabolism towards quinolinic acid and, thus, raise the concentration of quinolinic acid, which would support the fact that quinolinic acid is one of the factors responsible for depression/suicidality in METH addicts.
4.8. Picolinic Acid
When the enzyme 2-amino-3-carboxymuconic-6-semialdehyde decarboxylase (ACMSD) acts on ACMS, rather than forming quinolinic acid spontaneously, it instead forms picolinic acid [66]. It is not known whether picolinic acid is able to cross the blood–brain barrier or not, but it can be easily detected in human cerebrospinal fluid [115].
ACMSD is also present in glial and neuronal cells of the cortex and hippocampus [113]; hence, picolinic acid can be produced in the brain. A study showed that picolinic acid can antagonize the neurotoxic effects of quinolinic acid, as well as having neuroprotective properties [116]. In depressed suicide attempters, picolinic acid levels in the cerebrospinal fluid are known to significantly drop when compared with the levels in healthy individuals, which is linked to a drop in the actions of the ACMSD enzyme [117]. This means that an increase in quinolinic acid levels will occur because 2-amino-3-carboxymuconic-6-semialdehyde spontaneously converts to quinolinic acid in the absence of the ACMSD enzyme. Based on these findings, it can be concluded that lower levels of picolinic acid can be expected in the brains and bodies of people with METH addiction. However, the relationship between picolinic acid and METH addiction is not fully known, and further studies will provide a better understanding of this relationship.
4.9. Nicotinamide Adenine Dinucleotide (NAD+)
Nicotinamide adenine dinucleotide (NAD+) is formed from the metabolism of quinolinic acid via the action of quinolinic acid phosphoribosyl transferase [61]. The TDO enzyme of the tryptophan pathway, which is known to carry out most of the metabolism in the tryptophan pathway in the liver, also leads to the formation of NAD+. It is also considered to have a significant role in the maintenance of cell viability [67]. In one study in mice, which were forced to swim, it was noted that NAD+ had anti-depressant effects in mice after they were stressed [118]. NAD+ is also found to be an important co-enzyme in many oxidation–reduction reactions [67]. It is also responsible for cell homeostasis and energy metabolism, thus making it an important cellular molecule that is a regulator of the circadian cycle [119]. NAD+ can have positive effects in METH addicts who suffer from suicidal depression, but further research is required to explore the correlation between NAD+ and METH-induced neuronal damage.
4.10. Reactive Oxygen and Nitrogen Species
Reactive oxygen (ROS) and nitrogen species (RNS) are produced as by-products of the tryptophan metabolism pathway. Increases in ROS and RNS are known to be responsible for oxidative stress and nitrosative stress in the body, and this oxidative stress has been linked to anxiety, depression, and inflammatory responses. RNS and ROS usually include superoxide, nitric oxide, and hydrogen peroxide [120]. Based on this, increases in levels of RNS and ROS are very likely in the CNSs of METH addicts. It has also been noted that decreased levels of the antioxidative compound glutathione also occur in victims of major depressive disorder [121]. Due to this, antidepressants that target the production of RNS and ROS are currently being explored, which includes inhibiting the over-production of RNS and ROS along with suppressing their inflammatory effects [122]. METH affects the balance of tryptophan and its metabolites significantly, and further research on each of the pathways will be constructive towards the development of new therapeutics, which will aid in evading the detrimental effects of METH on the tryptophan metabolism pathways.
5. Tryptophan Metabolites and Treatment Options for Addiction
Acute/chronic addiction to alcohol has significant impacts on tryptophan metabolism in human and animal subjects. Tryptophan has been studied in relation to alcohol addiction because it is the precursor of indolylamine serotonin, which controls disparate functions in the CNS such as mood and depression [123,124]. It was corroborated in the mid-1980s that 5-HT had the ability to control the abnormal tendency to drink alcohol [84]. In animal studies, the presence of low central 5-HT concentration in various animal models was an interesting aspect of tryptophan metabolism related to alcohol addiction [125]. This deficiency is of great importance in alcohol-abuse behavior. The increasing trend of cocaine abuse has been a major health issue associated with millions of people′s dependency and high social costs [126,127]. Numerous studies have investigated the mechanism of action of cocaine, with no efficacious treatments for patients and compulsive users [128]. A small amount of tryptophan is used for 5-HT synthesis, while a greater proportion is metabolized via the kynurenine pathway manufacturing biologically active metabolites such as kynurenic acid, 3-hydroxykynurenine, and quinolinic acid. While 3-hydroxykynurenine and quinolinic acid are neurotoxic metabolites, kynurenic acid is thought to be involved in neuroprotection. Due to the great influence of 5-HT in the pathogenesis of mood disorders, the relationship between neurotoxic and neuroprotective metabolites of tryptophan is of great importance [129]. Currently, the pharmacological increase in kynurenic acid has been under extensive investigation as a new and promising option for treating those patients suffering from compulsive abuse of marijuana and nicotine dependency [40,130,131]. It has been corroborated that kynurenic acid can play an important role in relapse-like situations owing to its enhancement of an abrogating cocaine-seeking demeanor [131]. Table 2 presents information regarding the most commonly used addictive substances and their associated tryptophan metabolites.
6. Conclusions
This review evaluated the detrimental effects of addictive substances, specifically METH, on human mental and psychological dependency. The role of METH and other addictive substances in mental and psychological dependency in humans was presented, along with a description of tryptophan metabolites in the mammalian brain and their potential as new treatment options for controlling compulsive substance abuse. Tobacco smoke significantly increases the potential hazard of different chronic diseases such as lung diseases, heart failure, brain stroke, and chronic obstructive pulmonary diseases. The abuse of marijuana may result in various unfavorable side effects, such as the risk of permanent IQ loss, depression, and suicide planning. In the last five years, vaping has become popular, especially among the youth, and health issues and severe complications have already been reported; one awaits the addictive behaviors and health issues due to its use in the coming years. In the case of alcohol, it is worth mentioning that the long-term abuse of this addictive substance may cause the emergence of disparate chronic diseases such as high blood pressure, a decrease in the performance of the immune system, and digestive problems. Despite detrimental long-term results related to METH and other addictive substances, numerous people use them to alleviate short-term physical/emotional sensations. By eliminating these unfavorable states, drug-use behaviors are negatively reinforced. Abstinence from METH may cause a return to former undesirable emotional states that are associated with craving, usually leading to an elevated probability of relapse. The functional effects of METH on tryptophan and its disparate metabolites, including serotonin 5-HT (5-hydroxytryptophan), melatonin, kynurenine, and reactive oxygen and nitrogen species, as well as the formation and structure of each, were examined. Understanding the connections between tryptophan metabolites and addiction provides a valuable opportunity to investigate future perspectives on developing promising modalities for addiction treatment. The compulsive abuse of addictive substances as active CNS stimulants eventuates thse emergence of serious side effects such as poor mental/physical health and socio-economic challenges. Tryptophan metabolites show potential for application as new treatment options to manage compulsive abuse of addictive substances. Moreover, several studies have reported the development of vaccines against METH addiction [142,143,144], but none have been translated into human clinical trials yet. The identification of tryptophan metabolites as potential treatment options for addiction, in combination with new approaches such as vaccines, may lead to more effective methods for managing compulsive substance abuse in the future.
Author Contributions
Conceptualization, V.A. and K.N.; writing—original draft preparation, M.D. and N.R.; writing—review and editing, V.A., A.R., M.K.H., and M.D.; supervision, K.N. and V.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would also like to thank the Immunology and Translational Research Group and the Intestinal Neuropathy Group for their ongoing discussions. The Institute for Health and Sport, Victoria University Australia is also appreciated for its support. M.D., N.R., and M.K.H were supported by the Victoria University Postgraduate Scholarships.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goldstein, R.; Volkow, N. Dysfunktion av prefrontal cortex i missbruk: Neuroimaging fynd och kliniska konsekvenser. Nat. Rev. Neurosci. 2011, 12, 652–669. [Google Scholar] [CrossRef]
- Volkow, N.D.; Li, T.-K. Drug addiction: The neurobiology of behaviour gone awry. Nat. Rev. Neurosci. 2004, 5, 963–970. [Google Scholar] [CrossRef]
- Barnard, M. Drug Addiction and Families; Jessica Kingsley Publishers: London, UK, 2006. [Google Scholar]
- Ihezie, S.A.; Dafny, N. Prevention of Opioid Addiction. J. ISSN 2021, 2766, 2276. [Google Scholar] [CrossRef]
- Pantoni, M.M.; Kim, J.L.; Van Alstyne, K.R.; Anagnostaras, S.G. MDMA and memory, addiction, and depression: Dose-effect analysis. Psychopharmacology 2022, 239, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Heilig, M.; Augier, E.; Pfarr, S.; Sommer, W.H. Developing neuroscience-based treatments for alcohol addiction: A matter of choice? Transl. Psychiatry 2019, 9, 255. [Google Scholar] [CrossRef]
- Adams, M.K.; Chong, E.W.; Williamson, E.; Aung, K.Z.; Makeyeva, G.A.; Giles, G.G.; English, D.R.; Hopper, J.; Guymer, R.H.; Baird, P.N. 20/20—Alcohol and age-related macular degeneration: The Melbourne Collaborative Cohort Study. Am. J. Epidemiol. 2012, 176, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Nwokike, M.; Anusiem, C.; Arinze, C.; Ogbonna, A. The Abuse of Central Nervous System Stimulants and its Impact on the Youth of Eastern Nigeria. Int. Neuropsychiatr. Dis. J. 2021, 15, 20–33. [Google Scholar] [CrossRef]
- White, E.; Comiskey, C. Heroin epidemics, treatment and ODE modelling. Math. Biosci. 2007, 208, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Carmack, S.A.; Keeley, R.J.; Vendruscolo, J.C.; Lowery-Gionta, E.G.; Lu, H.; Koob, G.F.; Stein, E.A.; Vendruscolo, L.F. Heroin addiction engages negative emotional learning brain circuits in rats. J. Clin. Investig. 2019, 129, 2480–2484. [Google Scholar] [CrossRef]
- Ritter, A.; Cao, V.L.; Shanahan, M.; Shukla, N.; Perez, P.; Farrell, M. Modelling heroin careers over 40 years: Social costs. Drug Alcohol Depend. 2017, 100, e177. [Google Scholar] [CrossRef]
- Walsh, J.M. Are we there yet? Measuring Progress in the US War on Drugs in Latin America. WOLA Drug War Monit. 2004, 8. [Google Scholar]
- Hser, Y.-I. Predicting long-term stable recovery from heroin addiction: Findings from a 33-year follow-up study. J. Addict. Dis. 2007, 26, 51–60. [Google Scholar] [CrossRef]
- Dani, J.A.; Harris, R.A. Nicotine addiction and comorbidity with alcohol abuse and mental illness. Nat. Neurosci. 2005, 8, 1465–1470. [Google Scholar] [CrossRef] [PubMed]
- Scarlata, M.; Keeley, R.; Stein, E. Nicotine addiction: Translational insights from circuit neuroscience. Pharmacol. Biochem. Behav. 2021, 204, 173171. [Google Scholar] [CrossRef] [PubMed]
- Fagerström, K. Nicotine: Pharmacology, toxicity and therapeutic use. J. Smok. Cessat. 2014, 9, 53–59. [Google Scholar] [CrossRef]
- Jetton, J.A.; Ding, K.; Kim, Y.; Stone, D.U. Effects of tobacco smoking on human corneal wound healing. Cornea 2014, 33, 453–456. [Google Scholar] [CrossRef]
- Antonelli, M.; Ferrulli, A.; Sestito, L.; Vassallo, G.A.; Tarli, C.; Mosoni, C.; Rando, M.M.; Mirijello, A.; Gasbarrini, A.; Addolorato, G. Alcohol addiction-the safety of available approved treatment options. Expert Opin. Drug Saf. 2018, 17, 169–177. [Google Scholar] [CrossRef]
- Gibbins, R.J. Chronic Alcoholism and Alcohol Addiction; University of Toronto Press: Toronto, ON, USA, 2019. [Google Scholar]
- Behling, M.; Winters, B. Methods to Stop Caffeine Use and Minimize Caffeine Withdrawal Symptoms in the State of Caffeine Dependence: A Literature Review. Stud. Works 2021. Available online: https://scholarsarchive.byu.edu/cgi/viewcontent.cgi?article=1363&context=studentpub (accessed on 3 January 2023).
- Favrod-Coune, T.; Broers, B. Addiction to caffeine and other xanthines. In Textbook of Addiction Treatment; Springer: Berlin/Heidelberg, Germany, 2021; pp. 215–228. [Google Scholar]
- Alasmari, F. Caffeine induces neurobehavioral effects through modulating neurotransmitters. Saudi Pharm. J. 2020, 28, 445–451. [Google Scholar] [CrossRef]
- Chandio, Z.A.; Sidiqua, A.; Khaskheli, M.I.; Waghani, A.; Metlo, W.A. Review Effect of Caffeine Overdose. RADS J. Biol. Res. Appl. Sci. 2020, 11, 152–156. [Google Scholar] [CrossRef]
- Riezzo, I.; Fiore, C.; De Carlo, D.; Pascale, N.; Neri, M.; Turillazzi, E.; Fineschi, V. Side effects of cocaine abuse: Multiorgan toxicity and pathological consequences. Curr. Med. Chem. 2012, 19, 5624–5646. [Google Scholar] [CrossRef]
- Pierce, R.C.; Fant, B.; Swinford-Jackson, S.E.; Heller, E.A.; Berrettini, W.H.; Wimmer, M.E. Environmental, genetic and epigenetic contributions to cocaine addiction. Neuropsychopharmacology 2018, 43, 1471–1480. [Google Scholar] [CrossRef] [Green Version]
- Valencia-Alfonso, C.-E.; Luigjes, J.; Smolders, R.; Cohen, M.X.; Levar, N.; Mazaheri, A.; van den Munckhof, P.; Schuurman, P.R.; van den Brink, W.; Denys, D. Effective deep brain stimulation in heroin addiction: A case report with complementary intracranial electroencephalogram. Biol. Psychiatry 2012, 71, e35–e37. [Google Scholar] [CrossRef] [PubMed]
- Sarajlija, M.; Raketic, D.; Nesic, N. Heroin addiction in Serbian patients with Tourette syndrome. J. Psychiatr. Pract. 2018, 24, 424–427. [Google Scholar] [CrossRef]
- Sabet, K. Marijuana and legalization impacts. Berkeley J. Crim. L. 2018, 23, 84. [Google Scholar]
- Ethan, X.; Logan, A.; Liam, M.; Leonard, J. Impact of Marijuana (Cannabis) on Health, Safety and Economy. Int. Digit. Organ. Sci. Res. (IDOSR) J. Exp. Sci. 2020, 5, 43–52. [Google Scholar]
- Sun, X.; Xu, C.S.; Chadha, N.; Chen, A.; Liu, J. Focus: Addiction: Marijuana for Glaucoma: A Recipe for Disaster or Treatment? Yale J. Biol. Med. 2015, 88, 265–269. [Google Scholar]
- Temmingh, H.S.; van den Brink, W.; Howells, F.; Sibeko, G.; Stein, D.J. Methamphetamine use and antipsychotic-related extrapyramidal side-effects in patients with psychotic disorders. J. Dual Diagn. 2020, 16, 208–217. [Google Scholar] [CrossRef]
- Dhein, S.; Schmelmer, K.; Guenther, J.; Salameh, A. Aspects of methamphetamine abuse in adolescents and young adults in a Thuringian County. Eur. Addict. Res. 2018, 24, 98–105. [Google Scholar] [CrossRef]
- Naji, L.; Dennis, B.; Rosic, T.; Wiercioch, W.; Paul, J.; Worster, A.; Thabane, L.; Samaan, Z. Mirtazapine for the treatment of amphetamine and methamphetamine use disorder: A systematic review and meta-analysis. Drug Alcohol Depend. 2022, 232, 109295. [Google Scholar] [CrossRef]
- Escobar-Chávez, J.J.; Domínguez-Delgado, C.L.; Rodríguez-Cruz, I.M. Targeting nicotine addiction: The possibility of a therapeutic vaccine. Drug Des. Dev. Ther. 2011, 5, 211–224. [Google Scholar] [CrossRef]
- Sliwińska-Mossoń, M.; Zieleń, I.; Milnerowicz, H. New trends in the treatment of nicotine addiction. Acta Pol. Pharm. 2014, 71, 525–530. [Google Scholar] [PubMed]
- Jamali, Q. Galantamine as a Treatment Option for Nicotine Addiction. J. Smok. Cessat. 2021, 2021, 9975811. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.H.; Walitzer, K.S.; Blanco, J.; Swiatek, D.; Paine Hughes, L.; Quiñones-Lombraña, A.; Shyhalla, K. Medication-enhanced behavior therapy for alcohol use disorder: Naltrexone, Alcoholics Anonymous Facilitation, and OPRM1 genetic variation. J. Subst. Abus. Treat. 2019, 104, 7–14. [Google Scholar] [CrossRef]
- Saunders, E.C.; McGovern, M.P.; Lambert-Harris, C.; Meier, A.; McLeman, B.; Xie, H. The impact of addiction medications on treatment outcomes for persons with co-occurring PTSD and opioid use disorders. Am. J. Addict. 2015, 24, 722–731. [Google Scholar] [CrossRef] [Green Version]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chirumbolo, S.; Chartrand, M.S.; Bjørklund, G. How important is tryptophan in human health? Crit. Rev. Food Sci. Nutr. 2019, 59, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Araos, P.; Vidal, R.; O’Shea, E.; Pedraz, M.; García-Marchena, N.; Serrano, A.; Suárez, J.; Castilla-Ortega, E.; Ruiz, J.J.; Campos-Cloute, R.; et al. Serotonin is the main tryptophan metabolite associated with psychiatric comorbidity in abstinent cocaine-addicted patients. Sci. Rep. 2019, 9, 16842. [Google Scholar] [CrossRef]
- Bamia, C.; Trichopoulou, A.; Lenas, D.; Trichopoulos, D. Tobacco smoking in relation to body fat mass and distribution in a general population sample. Int. J. Obes. 2004, 28, 1091–1096. [Google Scholar] [CrossRef]
- Harris, K.K.; Zopey, M.; Friedman, T.C. Metabolic effects of smoking cessation. Nat. Rev. Endocrinol. 2016, 12, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Arterberry, B.J.; Treloar, H.R.; Smith, A.E.; Martens, M.P.; Pedersen, S.L.; McCarthy, D.M. Marijuana use, driving, and related cognitions. Psychol. Addict. Behav. 2013, 27, 854–860. [Google Scholar] [CrossRef]
- Gottlieb, E. Cannabis: A Danger to the Adolescent Brain—How Pediatricians Can Address Marijuana Use; Citeseer: University Park, PA, USA, 2012. [Google Scholar]
- Roerecke, M.; Vafaei, A.; Hasan, O.S.M.; Chrystoja, B.R.; Cruz, M.; Lee, R.; Neuman, M.G.; Rehm, J. Alcohol Consumption and Risk of Liver Cirrhosis: A Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2019, 114, 1574–1586. [Google Scholar] [CrossRef]
- Nawrot, P.; Jordan, S.; Eastwood, J.; Rotstein, J.; Hugenholtz, A.; Feeley, M. Effects of caffeine on human health. Food Addit. Contam. 2003, 20, 1–30. [Google Scholar] [CrossRef]
- Saimaiti, A.; Zhou, D.D.; Li, J.; Xiong, R.G.; Gan, R.Y.; Huang, S.Y.; Shang, A.; Zhao, C.N.; Li, H.Y.; Li, H.B. Dietary sources, health benefits, and risks of caffeine. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hswen, Y.; Zhang, A.; Brownstein, J.S. Estimating the incidence of cocaine use and mortality with music lyrics about cocaine. NPJ Digit. Med. 2021, 4, 100. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Hong, R.A.; Shohet, R.V.; Seto, T.B.; Parikh, N.I. Methamphetamine-Associated Cardiomyopathy. Clin. Cardiol. 2013, 36, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.; Mayer, M.; Habib, A.; Rashidi, N.; Filippone, R.T.; Fraser, S.; Prakash, M.D.; Sinnayah, P.; Tangalakis, K.; Mathai, M.L.; et al. Methamphetamine Induces Systemic Inflammation and Anxiety: The Role of the Gut–Immune–Brain Axis. Int. J. Mol. Sci. 2022, 23, 11224. [Google Scholar] [CrossRef]
- Sulzer, D.; Sonders, M.S.; Poulsen, N.W.; Galli, A. Mechanisms of neurotransmitter release by amphetamines: A review. Prog. Neurobiol. 2005, 75, 406–433. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Wang, C.; Zhang, X.; Pestell, R.G. Acetylation of nuclear receptors in cellular growth and apoptosis. Biochem. Pharmacol. 2004, 68, 1199–1208. [Google Scholar] [CrossRef]
- Prakash, M.D.; Tangalakis, K.; Antonipillai, J.; Stojanovska, L.; Nurgali, K.; Apostolopoulos, V. Methamphetamine: Effects on the brain, gut and immune system. Pharmacol. Res. 2017, 120, 60–67. [Google Scholar] [CrossRef]
- Papageorgiou, M.; Raza, A.; Fraser, S.; Nurgali, K.; Apostolopoulos, V. Methamphetamine and its immune-modulating effects. Maturitas 2019, 121, 13–21. [Google Scholar] [CrossRef]
- Surratt, C.K.; Ukairo, O.T.; Ramanujapuram, S. Recognition of psychostimulants, antidepressants, and other inhibitors of synaptic neurotransmitter uptake by the plasma membrane monoamine transporters. AAPS J. 2005, 7, E739–E751. [Google Scholar] [CrossRef]
- Darke, S.; Kaye, S.; McKetin, R.; Duflou, J. Major physical and psychological harms of methamphetamine use. Drug Alcohol Rev. 2009, 27, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Teixeira-Gomes, A.; Costa, V.M.; Feio-Azevedo, R.; de Lourdes Bastos, M.; Carvalho, F.; Capela, J.P. The neurotoxicity of amphetamines during the adolescent period. Int. J. Dev. Neurosci. 2015, 41, 44–62. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.; Rose Childress, A.; Thomas Mclellan, A.; Ehrman, R. Classical Conditioning in Drug-Dependent Humans. Ann. N. Y. Acad. Sci. 1992, 654, 400–415. [Google Scholar] [CrossRef]
- Danaceau, J.P.; Deering, C.E.; Day, J.E.; Smeal, S.J.; Johnson-Davis, K.L.; Fleckenstein, A.E.; Wilkins, D.G. Persistence of tolerance to methamphetamine-induced monoamine deficits. Eur. J. Pharmacol. 2007, 559, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Carrico, A.; Johnson, M.; Morin, S.F.; Remien, R.; Riley, E.D.; Hecht, F.; Fuchs, D. Stimulant Use is Associated with Immune Activation and Depleted Tryptophan Among HIV-Positive Persons on Anti-Retroviral Therapy. Brain Behav. Immun. 2008, 22, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Bradley, K.; Case, J.A.C.; Khan, O.; Ricart, T.; Hanna, A.; Alonso, C.; Gabbay, V. The role of the kynurenine pathway in suicidality in adolescent major depressive disorder. Psychiatry Res. 2015, 227, 206–212. [Google Scholar] [CrossRef]
- Fleckenstein, A.E.; Volz, T.J.; Riddle, E.L.; Gibb, J.W.; Hanson, G.R. New Insights into the Mechanism of Action of Amphetamines. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 681–698. [Google Scholar] [CrossRef]
- Halpin, L.E.; Collins, S.A.; Yamamoto, B.K. Neurotoxicity of methamphetamine and 3,4-methylenedioxymethamphetamine. Life Sci. 2014, 97, 37–44. [Google Scholar] [CrossRef]
- Fiore, A.; Murray, P.J. Tryptophan and indole metabolism in immune regulation. Curr. Opin. Immunol. 2021, 70, 7–14. [Google Scholar] [CrossRef]
- Davidson, M.; Rashidi, N.; Nurgali, K.; Apostolopoulos, V. The Role of Tryptophan Metabolites in Neuropsychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 9968. [Google Scholar] [CrossRef]
- Bryleva, E.; Brundin, L. Kynurenine Pathway Metabolites and Suicidality. Neuropharmacology 2017, 112, 324–330. [Google Scholar] [CrossRef]
- Engin, A.; Engin, A. Tryptophan Metabolism: Implications for Biological Processes, Health and Disease; Humana Press: Totowa, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Petit, A.; Karila, L.; Chalmin, F.; Lejoyeux, M. Methamphetamine addiction: A review of the literature. J. Addict. Res. Ther. S 2012, 1, 1–6. [Google Scholar] [CrossRef]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Nollen, E.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Cheong, J.E.; Ekkati, A.; Sun, L. A patent review of IDO1 inhibitors for cancer. Expert Opin. Ther. Pat. 2018, 28, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.H. The targeting of tumor-associated macrophages by vaccination. Cell Stress 2019, 3, 139. [Google Scholar] [CrossRef] [PubMed]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef] [PubMed]
- Van der Meide, P.H.; Schellekens, H. Cytokines and the immune response. Biotherapy 1996, 8, 243–249. [Google Scholar] [CrossRef]
- Li, Y.; Hu, N.; Yang, D.; Oxenkrug, G.; Yang, Q. Regulating the balance between the kynurenine and serotonin pathways of tryptophan metabolism. FEBS J. 2017, 284, 948–966. [Google Scholar] [CrossRef]
- Maddison, D.C.; Giorgini, F. The kynurenine pathway and neurodegenerative disease. Semin. Cell Dev. Biol. 2015, 40, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Boyen, G.; Steinkamp, M.; Reinshagen, M.; Schäfer, K.-H.; Adler, G.; Kirsch, J. Proinflammatory cytokines increase glial fibrillary acidic protein expression in enteric glia. Gut 2004, 53, 222–228. [Google Scholar] [CrossRef]
- Gibney, G.T.; Hamid, O.; Lutzky, J.; Olszanski, A.J.; Mitchell, T.C.; Gajewski, T.F.; Chmielowski, B.; Hanks, B.A.; Zhao, Y.; Newton, R.C. Phase 1/2 study of epacadostat in combination with ipilimumab in patients with unresectable or metastatic melanoma. J. Immunother. Cancer 2019, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Driessens, G.; Bartlett, D.; Cai, D.; Cauwenberghs, S.; Crosignani, S.; Dalvie, D.; Denies, S.; Dillon, C.P.; Fantin, V.R. Characterization of the selective indoleamine 2, 3-dioxygenase-1 (IDO1) catalytic inhibitor EOS200271/PF-06840003 supports IDO1 as a critical resistance mechanism to PD-(L) 1 blockade therapy. Mol. Cancer Ther. 2018, 17, 2530–2542. [Google Scholar] [CrossRef] [Green Version]
- Hassanzadeganroudsari, M.; Soltani, M.; Heydarinasab, A.; Nakhjiri, A.T.; Hossain, M.D.K.; Khiyavi, A.A. Mathematical modeling and simulation of molecular mass transfer across blood brain barrier in brain capillary. J. Mol. Liq. 2020, 310, 113254. [Google Scholar] [CrossRef]
- Gobaille, S.; Kemmel, V.; Brumaru, D.; Dugave, C.; Aunis, D.; Maitre, M. Xanthurenic acid distribution, transport, accumulation and release in the rat brain. J. Neurochem. 2008, 105, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Dostal, C.; Carson Sulzer, M.; Kelley, K.W.; Freund, G.; McCusker, R.H. Glial and tissue-specific regulation of Kynurenine Pathway dioxygenases by acute stress of mice. Neurobiol. Stress 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Koshiguchi, M.; Komazaki, H.; Hirai, S.; Egashira, Y. Ferulic acid suppresses expression of tryptophan metabolic key enzyme indoleamine 2, 3-dioxygenase via NFκB and p38 MAPK in lipopolysaccharide-stimulated microglial cells. Biosci. Biotechnol. Biochem. 2017, 81, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B. Tryptophan metabolism in alcoholism. Nutr. Res. Rev. 2002, 15, 123–152. [Google Scholar] [CrossRef]
- Merz, F. United Nations Office on Drugs and Crime: World Drug Report 2017. 2017. SIRIUS-Z. Strateg. Anal. 2018, 2, 85–86. [Google Scholar] [CrossRef]
- Azmi, A.; Jai, J.; Zamanhuri, N.; Yahya, A. Precious metals recovery from electroplating wastewater: A review. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2018; p. 012024. [Google Scholar]
- Vrajová, M.; Šlamberová, R.; Hoschl, C.; Ovsepian, S.V. Methamphetamine and sleep impairments: Neurobehavioral correlates and molecular mechanisms. Sleep 2021, 44, zsab001. [Google Scholar] [CrossRef] [PubMed]
- Rawson, R.A.; Gonzales, R.; Marinelli-Casey, P.; Ang, A. Methamphetamine dependence: A closer look at treatment response and clinical characteristics associated with route of administration in outpatient treatment. Am. J. Addict. 2007, 16, 291–299. [Google Scholar] [CrossRef]
- McKetin, R.; Voce, A.; Burns, R.; Shanahan, M. Health-related quality of life among people who use methamphetamine. Drug Alcohol Rev. 2019, 38, 503–509. [Google Scholar] [CrossRef] [PubMed]
- McKetin, R.; Lubman, D.I.; Lee, N.M.; Ross, J.E.; Slade, T.N. Major depression among methamphetamine users entering drug treatment programs. Med. J. Aust. 2011, 195, S51–S55. [Google Scholar] [CrossRef]
- Hoffman, W.F.; Jacobs, M.B.; Dennis, L.E.; McCready, H.D.; Hickok, A.W.; Smith, S.B.; Kohno, M. Psychopathy and corticostriatal connectivity: The link to criminal behavior in methamphetamine dependence. Front. Psychiatry 2020, 11, 90. [Google Scholar] [CrossRef]
- Newton, T.F.; De La Garza, R.; Kalechstein, A.D.; Tziortzis, D.; Jacobsen, C.A. Theories of addiction: Methamphetamine users’ explanations for continuing drug use and relapse. Am. J. Addict. 2009, 18, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Filip, M.; Bader, M. Overview on 5-HT receptors and their role in physiology and pathology of the central nervous system. Pharmacol. Rep. 2009, 61, 761–777. [Google Scholar] [CrossRef]
- Haughey, H.; Fleckenstein, A.E.; Hanson, G.R.; Haughey, H.M.; Fleckenstein, A.E.; Hanson, G.R. Differential regional effects of methampetamine on the activities of tryptophan and tyrosine hydroxylase. J. Neurochem. 1999, 72, 661–668. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ujike, H.; Iwata, N.; Inada, T.; Yamada, M.; Sekine, Y.; Uchimura, N.; Iyo, M.; Ozaki, N.; Itokawa, M.; et al. Association Analysis of the Tryptophan Hydroxylase 2 Gene Polymorphisms in Patients with Methamphetamine Dependence/Psychosis. Curr. Neuropharmacol. 2011, 9, 176–182. [Google Scholar] [CrossRef]
- Sitte, H.; Freissmuth, M. Amphetamines, new psychoactive drugs and the monoamine transporter cycle. Trends Pharmacol. Sci. 2015, 36, 41–50. [Google Scholar] [CrossRef]
- Hasler, B.; Smith, L.J.; Cousins, J.; Bootzin, R. Circadian Rhythms, Sleep, and Substance Abuse. Sleep Med. Rev. 2012, 16, 67–81. [Google Scholar] [CrossRef]
- Andersen, M.; Perez Diaz, M.; Murnane, K.; Howell, L.L. Effects of methamphetamine self-administration on actigraphy-based sleep parameters in rhesus monkeys. Psychopharmacology 2013, 227, 101–107. [Google Scholar] [CrossRef]
- Kuwano, N.; Kato, T.; Setoyama, D.; Sato-Kasai, M.; Shimokawa, N.; Hayakawa, K.; Ohgidani, M.; Sagata, N.; Kubo, H.; Kishimoto, J.; et al. Tryptophan-kynurenine and lipid related metabolites as blood biomarkers for first-episode drug-naïve patients with major depressive disorder: An exploratory pilot case-control study. J. Affect. Disord. 2018, 231, 74–82. [Google Scholar] [CrossRef]
- Stone, T.; Stoy, N.; Gail Darlington, L. An expanding range of targets for kynurenine metabolites of tryptophan. Trends Pharmacol. Sci. 2013, 34, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Rassoulpour, A.; Wu, H.Q.; Poeggeler, B.; Schwarcz, R. Systemic d-amphetamine administration causes a reduction of kynurenic acid levels in rat brain. Brain Res. 1998, 802, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Carlborg, A.; Jokinen, J.; Jönsson, E.; Erhardt, S.; Nordstrom, P. CSF kynurenic acid and suicide risk in schizophrenia spectrum psychosis. Psychiatry Res. 2013, 205, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Shimosato, K.; Nagao, N.; Watanabe, S.; Kitayama, S. Suppressive effects of trihexyphenidyl on methamphetamine-induced DA release as measured by in vivo microdialysis. Synapse 2003, 49, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Scofield, M. Similitude in Methamphetamine-Induced Neuroadaptations Across Susceptibility and Chronic Drug Exposure Paradigms. Biol. Psychiatry 2017, 81, e83–e84. [Google Scholar] [CrossRef]
- Haruki, H.; Hovius, R.; Grønlund Pedersen, M.; Johnsson, K. Tetrahydrobiopterin Biosynthesis as a Potential Target of the Kynurenine Pathway Metabolite Xanthurenic Acid. J. Biol. Chem. 2016, 291, 652–657. [Google Scholar] [CrossRef]
- Stone, T. Kynurenines in the CNS: From endogenous obscurity to therapeutic importance. Prog. Neurobiol. 2001, 64, 185–218. [Google Scholar]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Copeland, C.S.; Neale, S.A.; Bruno, V.; Battaglia, G.; Salt, T.; Nicoletti, F. Cinnabarinic acid and xanthurenic acid: Two kynurenine metabolites that interact with metabotropic glutamate receptors. Neuropharmacology 2017, 112, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Stone, T. Inhibitors of the Kynurenine Pathway. Eur. J. Med. Chem. 2000, 35, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Grando, V.; Dutra Filho, C.S.; Wannmacher, C.; Wajner, M. Evidence that quinolinic acid severely impairs energy metabolism through activation of NMDA receptors in striatum from developing rats. J. Neurochem. 2006, 99, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G. Quinolinic acid: Neurotoxicity. FEBS J. 2012, 279, 1355. [Google Scholar] [CrossRef]
- Erhardt, S.; Lim, C.; Linderholm, K.; Janelidze, S.; Lindqvist, D.; Samuelsson, M.; Lundberg, K.; Postolache, T.; Träskman-Bendz, L.; Guillemin, G.; et al. Connecting Inflammation with Glutamate Agonism in Suicidality. Neuropsychopharmacology 2012, 38, 743–752. [Google Scholar] [CrossRef]
- Bay-Richter, C.; Linderholm, K.; Lim, C.; Samuelsson, M.; Träskman-Bendz, L.; Guillemin, G.; Erhardt, S.; Brundin, L. A role for inflammatory metabolites as modulators of the glutamate N-methyl-D-aspartate receptor in depression and suicidality. Brain Behav. Immun. 2015, 43, 110–117. [Google Scholar] [CrossRef]
- Van Heeringen, K.; Bijttebier, S.; Desmyter, S.; Vervaet, M.; Baeken, C. Is there a neuroanatomical basis of the vulnerability to suicidal behavior? A coordinate-based meta-analysis of structural and functional MRI studies. Front. Hum. Neurosci. 2014, 8, 824. [Google Scholar] [CrossRef]
- Zarate, C.; Brutsche, N.; Ibrahim, L.; Franco-Chaves, J.; Diazgranados, N.; Cravchik, A.; Selter, J.; Marquardt, C.; Liberty, V.; Luckenbaugh, D. Replication of Ketamine’s Antidepressant Efficacy in Bipolar Depression: A Randomized Controlled Add-On Trial. Biol. Psychiatry 2012, 71, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chen, T.; Wang, J.; Jia, Y.; Ren, H.; Wu, F.; Hu, M.; Chen, Y. Methamphetamine modulates the production of interleukin-6 and tumor necrosis factor-alpha via the cAMP/PKA/CREB signaling pathway in lipopolysaccharide-activated microglia. Int. Immunopharmacol. 2018, 56, 168–178. [Google Scholar] [CrossRef]
- Grant, R.; Coggan, S.; Smythe, G. The Physiological Action of Picolinic Acid in the Human Brain. Int. J. Tryptophan Res. 2009, 2, 71–79. [Google Scholar] [CrossRef]
- Davidson, J.; Abraham, K.; Connor, K.; McLeod, M.N. Effectiveness of Chromium in atypical depression: A placebo-controlled trial. Biol. Psychiatry 2003, 53, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Rex, A.; Schickert, R.; Fink, H. Antidepressant-like effect of nicotinamide adenine dinucleotide in the forced swim test in rats. Pharmacol. Biochem. Behav. 2004, 77, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.-I.; Hsu, C.-P.; Sadoshima, J. Regulation of Cell Survival and Death by Pyridine Nucleotides. Circ. Res. 2012, 111, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative Stress and Psychological Disorders. Curr. Neuropharmacol. 2014, 12, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Gawryluk, J.; Wang, J.-F.; Andreazza, A.C.; Shao, L.; Young, L.T. Decreased levels of glutathione, the major brain antioxidant, in post-mortem prefrontal cortex from patients with psychiatric disorders. Int. J. Neuropsychopharmacol. 2011, 14, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-Y.; Lee, S.-J.; Han, C.; A Patkar, A.; S Masand, P.; Pae, C.-U. Oxidative/nitrosative stress and antidepressants: Targets for novel antidepressants. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 46, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.-B. Tryptophan metabolism: A versatile area providing multiple targets for pharmacological intervention. Egypt. J. Basic Clin. Pharmacol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Saberi, B. Alcoholism and Serotonin. Psychopharmacology 2019, 171, 340–348. [Google Scholar]
- Kelley, K.W.; Dantzer, R. Alcoholism and inflammation: Neuroimmunology of behavioral and mood disorders. Brain Behav. Immun. 2011, 25, S13–S20. [Google Scholar] [CrossRef]
- Degenhardt, L.; Baxter, A.J.; Lee, Y.Y.; Hall, W.; Sara, G.E.; Johns, N.; Flaxman, A.; Whiteford, H.A.; Vos, T. The global epidemiology and burden of psychostimulant dependence: Findings from the Global Burden of Disease Study 2010. Drug Alcohol Depend. 2014, 137, 36–47. [Google Scholar] [CrossRef]
- Araos, P.; Vergara-Moragues, E.; González-Saiz, F.; Pedraz, M.; García-Marchena, N.; Romero-Sanchiz, P.; Ruiz, J.J.; Campos-Cloute, R.; Serrano, A.; Pavón, F.J. Differences in the rates of drug polyconsumption and psychiatric comorbidity among patients with cocaine use disorders according to the mental health service. J. Psychoact. Drugs 2017, 49, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Shorter, D.; Domingo, C.B.; Kosten, T.R. Emerging drugs for the treatment of cocaine use disorder: A review of neurobiological targets and pharmacotherapy. Expert Opin. Emerg. Drugs 2015, 20, 15–29. [Google Scholar] [CrossRef]
- Myint, A.M.; Kim, Y.K. Cytokine–serotonin interaction through IDO: A neurodegeneration hypothesis of depression. Med. Hypotheses 2003, 61, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Justinova, Z.; Mascia, P.; Wu, H.-Q.; Secci, M.E.; Redhi, G.H.; Panlilio, L.V.; Scherma, M.; Barnes, C.; Parashos, A.; Zara, T. Reducing cannabinoid abuse and preventing relapse by enhancing endogenous brain levels of kynurenic acid. Nat. Neurosci. 2013, 16, 1652–1661. [Google Scholar] [CrossRef]
- Vengeliene, V.; Cannella, N.; Takahashi, T.; Spanagel, R. Metabolic shift of the kynurenine pathway impairs alcohol and cocaine seeking and relapse. Psychopharmacology 2016, 233, 3449–3459. [Google Scholar] [CrossRef]
- Wrona, M.Z.; Dryhurst, G. A putative metabolite of serotonin, tryptamine-4, 5-dione, is an irreversible inhibitor of tryptophan hydroxylase: Possible relevance to the serotonergic neurotoxicity of methamphetamine. Chem. Res. Toxicol. 2001, 14, 1184–1192. [Google Scholar] [CrossRef]
- Zheng, T.; Liu, L.; Shi, J.; Yu, X.; Xiao, W.; Sun, R.; Zhou, Y.; Aa, J.; Wang, G. The metabolic impact of methamphetamine on the systemic metabolism of rats and potential markers of methamphetamine abuse. Mol. BioSyst. 2014, 10, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Jenny, M.; Schröcksnadel, S.; Überall, F.; Fuchs, D. The potential role of cannabinoids in modulating serotonergic signaling by their influence on tryptophan metabolism. Pharmaceuticals 2010, 3, 2647–2660. [Google Scholar] [CrossRef]
- McDew-White, M.; Lee, E.; Alvarez, X.; Sestak, K.; Ling, B.J.; Byrareddy, S.N.; Okeoma, C.M.; Mohan, M. Cannabinoid control of gingival immune activation in chronically SIV-infected rhesus macaques involves modulation of the indoleamine-2,3-dioxygenase-1 pathway and salivary microbiome. EBioMedicine 2022, 75, 103769. [Google Scholar] [CrossRef]
- Ishak, N.; Ahmad, A.H.; Noor, S.A.M.; Ahmad, A. Detection of heroin metabolites at different developmental stages of Lucilia cuprina (Diptera: Calliphoridae) reared in heroin-treated meat: A preliminary analysis. Egypt. J. Forensic Sci. 2019, 9, 65. [Google Scholar] [CrossRef]
- Gaynor, C.M.; Handley, S.L. Effects of nicotine on head-shakes and tryptophan metabolites. Psychopharmacology 2001, 153, 327–333. [Google Scholar] [CrossRef]
- Fang, C.; Hayashi, S.; Du, X.; Cai, X.; Deng, B.; Zheng, H.; Ishido, S.; Tsutsui, H.; Sheng, J. Caffeine protects against stress-induced murine depression through activation of PPARγC1α-mediated restoration of the kynurenine pathway in the skeletal muscle. Sci. Rep. 2021, 11, 7287. [Google Scholar] [CrossRef]
- Varma, S.D.; Hegde, K.R. Kynurenine-induced photo oxidative damage to lens in vitro: Protective effect of caffeine. Mol. Cell. Biochem. 2010, 340, 49–54. [Google Scholar] [CrossRef]
- Giménez-Gómez, P.; Ballestín, R.; de Biedma-Elduayen, L.G.; Vidal, R.; Ferrer-Pérez, C.; Reguilón, M.D.; O’Shea, E.; Miñarro, J.; Colado, M.I.; Rodríguez-Arias, M. Decreased kynurenine pathway potentiate resilience to social defeat effect on cocaine reward. Neuropharmacology 2021, 197, 108753. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.; Mestre-Pintó, J.-I.; Gómez-Gómez, À.; Martinez-Sanvisens, D.; Rodríguez-Minguela, R.; Papaseit, E.; Pérez-Mañá, C.; Langohr, K.; Valverde, O.; Pozo, Ó.J. The tryptophan system in cocaine-induced depression. J. Clin. Med. 2020, 9, 4103. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Davidson, M.; Kypreos, E.; Feehan, J.; Muir, J.A.; Nurgali, K.; Apostolopoulos, V. Immunotherapies for the Treatment of Drug Addiction. Vaccines 2022, 10, 1778. [Google Scholar] [CrossRef]
- Kamal Hossain, M.; Davidson, M.; Feehan, J.; Deraos, G.; Nurgali, K.; Matsoukas, J.; Apostolopoulos, V. Development and characterization of a novel conjugated methamphetamine vaccine. Vaccine 2022, 40, 5882–5891. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Hassanzadeganroudsari, M.; Kypreos, E.; Feehan, J.; Apostolopoulos, V. Immune to addiction: How immunotherapies can be used to combat methamphetamine addiction. Expert Rev. Vaccines 2021, 20, 707–715. [Google Scholar] [CrossRef]
Figure 1.
The general short-term and long-term effects of addiction on the body (Figure created with BioRender.com).
Figure 1.
The general short-term and long-term effects of addiction on the body (Figure created with BioRender.com).

Figure 2.
Schematic demonstration of tryptophan metabolites, its related disparate pathways, and its metabolites′ effects on the central nervous system.
Figure 2.
Schematic demonstration of tryptophan metabolites, its related disparate pathways, and its metabolites′ effects on the central nervous system.

Figure 3.
Methamphetamine (METH) administration leads to a detrimental effect on the serotonin pathway in the central nervous system [Figure created with BioRender.com].
Figure 3.
Methamphetamine (METH) administration leads to a detrimental effect on the serotonin pathway in the central nervous system [Figure created with BioRender.com].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Some of the common addictive substances, mechanisms of action, and side effects.
| Drug | Mechanism of Action | Chemical Structure | Mode of Administration | Side Effects | Ref. |
|---|---|---|---|---|---|
| Alcohol | Promotion of inhibitory neurotransmission |  | Oral | Liver cirrhosis Neurotoxicity Drowsiness Loss of consciousness Gaps in memory | [6,18,19] |
| Caffeine | Release of dopamine, serotonin, and noradrenaline |  | Oral | Increased vigilance and metabolic activity Insomnia Nervousness Restlessness Nausea | [20,21,22,23] |
| Cocaine | Potentiation of dopamine and norepinephrine |  | Injection Inhalation | Euphoria Increased concentration Pupillary dilation | [24,25] |
| Heroin | Stimulation of opioid receptor |  | Injection Inhalation | Analgesia Depression Pupillary constriction Pneumonia Increased heart rate Difficulty sleeping Difficulty breathing Dry mouth | [10,26,27] |
| Marijuana | Stimulation of cannabinoid receptor type 1 and cannabinoid receptor type 2 |  | Oral Smoke | Tachycardia High blood pressure Decrease in saccadic accuracy Difficulty concentrating Hallucinations Abnormal happiness Depression | [28,29,30] |
| Methamphetamine | Catecholamine potentiation |  | Injection Inhalation Oral Intranasal sniffing | Depression Euphoria Pupillary dilation Cardiovascular diseases | [31,32,33] |
| Nicotine | Nicotinic acetylcholine agonist |  | Oral Smoke | Delayed coronary healing Hypertension Tachycardia Smoke-associated dryness | [34,35,36] |
Table 2.
Summary of common addictive substances and their associated tryptophan metabolites.
| Drug | Tryptophan Metabolite | Ref. |
|---|---|---|
| METH | 5-HT, KA, and QA | [132,133] |
| Alcohol | 5-HT, KA, 3-HK, and 3-HAA | [84,123] |
| Marijuana | IDO 1, KA, and QA | [134,135] |
| Heroin | QA and 5-HIAA | [136] |
| Nicotine | KA, 5-HT, KYN, HKY, and 3-HAA | [137] |
| Caffeine | KA, 5-HT, 5-HIAA, and QA | [138,139] |
| Cocaine | KA, KYN, 5-HT, and 5-HIAA | [140,141] |
Abbreviations: IDO, indoleamine 2,3-deoxygenase; KA, kynurenic acid; KYN, kynurenine; L-5HT, L-5-hydroxytryptophan; METH, methamphetamine; QA, Quinolinic acid; 3-HAA, 3-hydroxyanthranillic acid; 5-HIAA, 5-hydroxyindoleacetic acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Davidson, M.; Rashidi, N.; Hossain, M.K.; Raza, A.; Nurgali, K.; Apostolopoulos, V. Tryptophan and Substance Abuse: Mechanisms and Impact. Int. J. Mol. Sci. 2023, 24, 2737. https://doi.org/10.3390/ijms24032737
AMA Style
Davidson M, Rashidi N, Hossain MK, Raza A, Nurgali K, Apostolopoulos V. Tryptophan and Substance Abuse: Mechanisms and Impact. International Journal of Molecular Sciences. 2023; 24(3):2737. https://doi.org/10.3390/ijms24032737
Chicago/Turabian StyleDavidson, Majid, Niloufar Rashidi, Md Kamal Hossain, Ali Raza, Kulmira Nurgali, and Vasso Apostolopoulos. 2023. "Tryptophan and Substance Abuse: Mechanisms and Impact" International Journal of Molecular Sciences 24, no. 3: 2737. https://doi.org/10.3390/ijms24032737
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.