

Mechanobiology of the Human Intervertebral Disc: Systematic Review of the Literature and Future Perspectives
 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
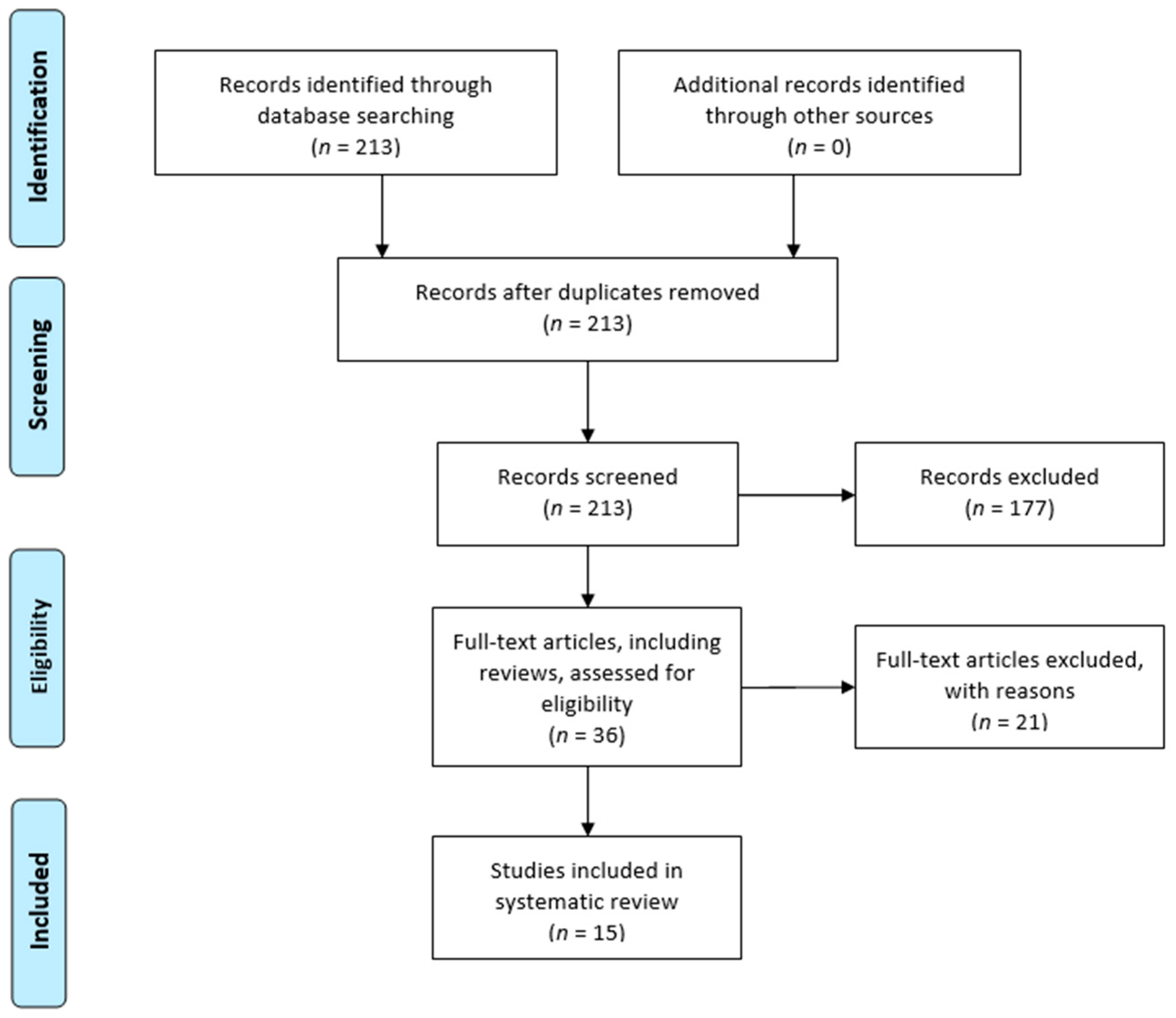
2.1. Review Design
2.2. Search Strategy
2.3. Study Selection
2.4. Data Extraction
3. Results
3.1. Included Studies
3.2. Cohort Characteristics
3.3. Evaluated Cell Types and Molecular Pathways
3.4. Applied Mechanical Stimuli and Cell Environmental Conditions
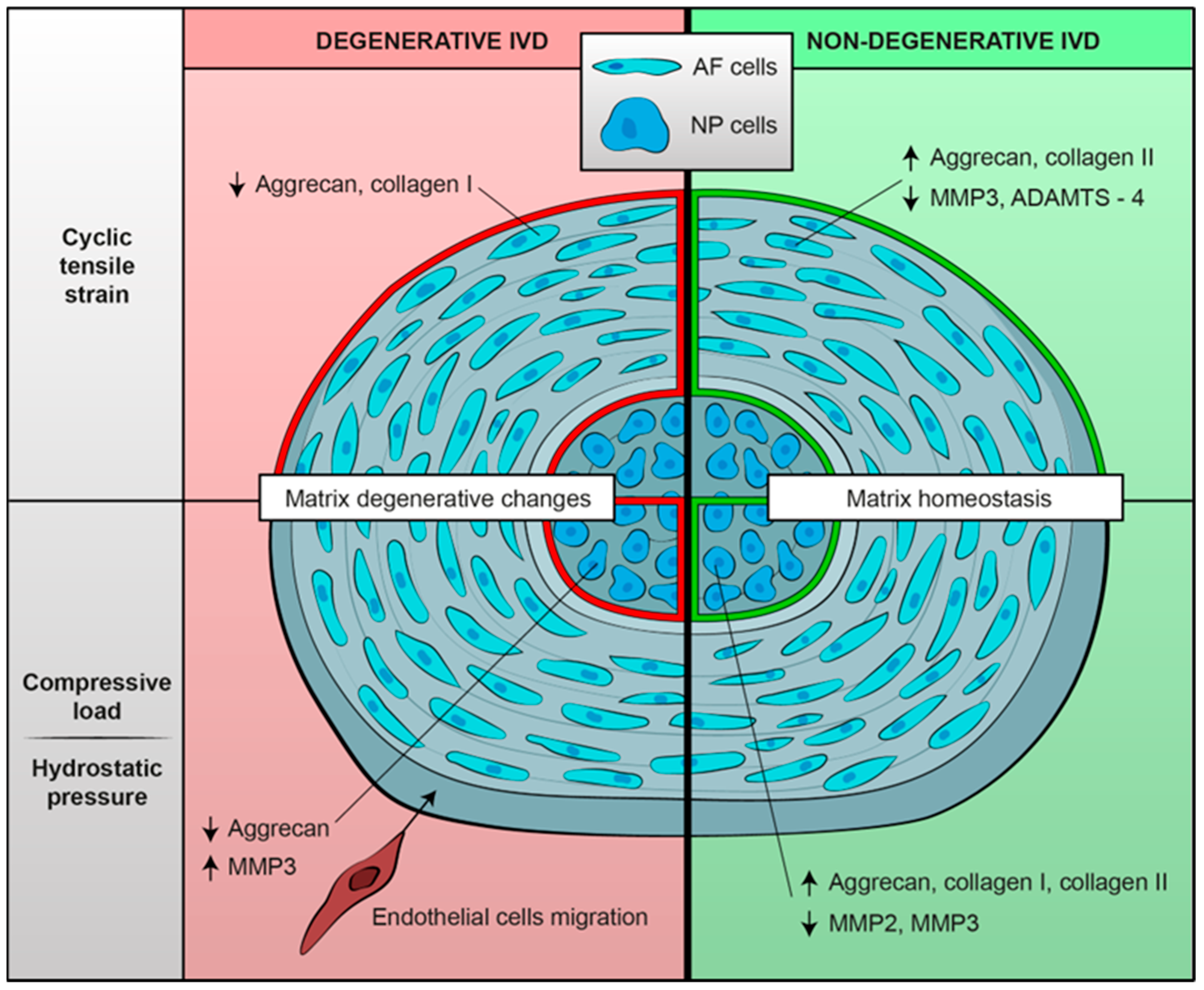
3.4.1. Cyclic Tensile Strain (CTS)
3.4.2. Compressive Load
3.4.3. Hydrostatic Pressure
3.4.4. Microgravity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ehrlich, G.E. Low back pain. Bull. World Health Organ. 2003, 81, 671. [Google Scholar] [PubMed]
- Dagenais, S.; Caro, J.; Haldeman, S. A systematic review of low back pain cost of illness studies in the United States and internationally. Spine J. 2008, 8, 8–20. [Google Scholar] [CrossRef] [PubMed]
- DePalma, M.J.; Ketchum, J.M.; Saullo, T. What Is the Source of Chronic Low Back Pain and Does Age Play a Role? Pain Med. 2011, 12, 224–233. [Google Scholar] [CrossRef]
- Guiot, B.H.; Fessler, R.G.; Benzel, E.C.; Parsa, A.T.; McCormick, P.C.; Sonntag, V.K.H. Molecular Biology of Degenerative Disc Disease. Neurosurgery 2000, 47, 1034–1040. [Google Scholar] [CrossRef]
- Eyre, D.; Buckwalter, J.; Caterson, B.; Heinegard, D.; Oegema, T.; Pearce, R.; Pope, M.; Urban, J. Basic Sciences Perspectives: Part B—Intervertebral Discs; American Academy of Orthopaedic Surgeons: Park Ridge, IL, USA, 1989. [Google Scholar]
- Diamant, B.; Karlsson, J.; Nachemson, A. Correlation between lactate levels and pH in discs of patients with lumbar rhizopathies. Cell Mol. Life Sci. 1968, 24, 1195–1196. [Google Scholar] [CrossRef]
- Nachemson, A. Intradiscal Measurements of pH in Patients with Lumbar Rhizopathies. Acta Orthop. 1969, 40, 23–42. [Google Scholar] [CrossRef]
- Wilke, H.J.; Neef, P.; Caimi, M.; Hoogland, T.; Claes, L.E. New In-vivo measurements of pressures in disc in daily. Spine 1999, 8, 755–762. [Google Scholar] [CrossRef]
- Costi, J.; Stokes, I.; Gardner-Morse, M.; Laible, J.; Scoffone, H.; Iatridis, J. Direct measurement of intervertebral disc maximum shear strain in six degrees of freedom: Motions that place disc tissue at risk of injury. J. Biomech. 2007, 40, 2457–2466. [Google Scholar] [CrossRef] [PubMed]
- Ambard, D.; Cherblanc, F. Mechanical Behavior of Annulus Fibrosus: A Microstructural Model of Fibers Reorientation. Ann. Biomed. Eng. 2009, 37, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Freemont, A.; Peacock, T.; Goupille, P.; Hoyland, J.; O’Brien, J.; Jayson, M. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef] [PubMed]
- David, G.; Ciurea, A.V.; Iencean, S.M.; Mohan, A. Angiogenesis in the degeneration of the lumbar intervertebral disc. J. Med. Life 2010, 3, 154–161. [Google Scholar] [PubMed]
- La Binch, A.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Cross, A.K.; Le Maitre, C.L. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Thromb. Haemost. 2014, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-Q.; Jiang, L.-S.; Dai, L.-Y. Programmed cell death in intervertebral disc degeneration. Apoptosis 2006, 11, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.; Freemont, A.; Hoyland, J. Accelerated cellular senescence in degenerate intervertebral discs: A possible role in the pathogenesis of intervertebral disc degeneration. Thromb. Haemost. 2007, 9, R45. [Google Scholar] [CrossRef] [PubMed]
- Hastreiter, D.; Ozuna, R.M.; Spector, M. Regional variations in certain cellular characteristics in human lumbar intervertebral discs, including the presence of α-smooth muscle actin. J. Orthop. Res. Publ. Orthop. Res. Soc. 2001, 19, 597–604. [Google Scholar] [CrossRef]
- Hunter, C.; Matyas, J.; Duncan, N. The Notochordal Cell in the Nucleus Pulposus: A Review in the Context of Tissue Engineering. Tissue Eng. 2003, 9, 667–677. [Google Scholar] [CrossRef]
- Le Maitre, C.; Pockert, A.; Buttle, D.; Freemont, T.; Hoyland, J. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35, 652–655. [Google Scholar] [CrossRef]
- Akyol, S.; Eraslan, B.S.; Etyemez, H.; Tanriverdi, T.; Hanci, M. Catabolic cytokine expressions in patients with degenerative disc disease. Turk. Neurosurg. 2010, 20, 492–499. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Hoyland, J.A.; Freemont, A.J. Catabolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1β and TNFα expression profile. Thromb. Haemost. 2007, 9, R77. [Google Scholar] [CrossRef]
- Kang, J.D.; Georgescu, H.I.; McIntyre-Larkin, L.; Stefanovic-Racic, M.; Evans, C.H. Herniated Cervical Intervertebral Discs Spontaneously Produce Matrix Metalloproteinases, Nitric Oxide, Interleukin-6, and Prostaglandin E2. Spine 1995, 20, 2373–2378. [Google Scholar] [CrossRef]
- Kang, J.D.; Georgescu, H.I.; McIntyre-Larkin, L.; Stefanovic-Racic, M.; Donaldson, W.F., III; Evans, C.H. Herniated Lumbar Intervertebral Discs Spontaneously Produce Matrix Metalloproteinases, Nitric Oxide, Interleukin-6, and Prostaglandin E2. Spine 1996, 21, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M.; Uchida, K.; Inoue, S.; Takano, S.; Nakawaki, M.; Kawakubo, A.; Sekiguchi, H.; Nakazawa, T.; Imura, T.; Saito, W.; et al. A High Body Mass Index and the Vacuum Phenomenon Upregulate Pain-Related Molecules in Human Degenerated Intervertebral Discs. Int. J. Mol. Sci. 2022, 23, 2973. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.-J.; Cui, H.; Pan, H.; MC Cheung, K.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful intervertebral disc degeneration and inflammation: From laboratory evidence to clinical interventions. Bone Res. 2021, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Galbusera, F.; Pratsinis, H.; Mavrogonatou, E.; Mietsch, A.; Kletsas, D.; Wilke, H.-J. Mechanical loading of the intervertebral disc: From the macroscopic to the cellular level. Eur. Spine J. 2014, 23, 333–343. [Google Scholar] [CrossRef]
- Setton, L.A.; Chen, J. Cell Mechanics and Mechanobiology in the Intervertebral Disc. Spine 2004, 29, 2710–2723. [Google Scholar] [CrossRef]
- Setton, L.A.; Chen, J. Mechanobiology of the Intervertebral Disc and Relevance to Disc Degeneration. J. Bone Jt. Surg. 2006, 88, 52–57. [Google Scholar] [CrossRef]
- Steele, J.; Bruce-Low, S.; Smith, D.; Osborne, N.; Thorkeldsen, A. Can specific loading through exercise impart healing or regeneration of the intervertebral disc? Spine J. 2015, 15, 2117–2121. [Google Scholar] [CrossRef]
- Hsieh, A.H.; Twomey, J.D. Cellular mechanobiology of the intervertebral disc: New directions and approaches. J. Biomech. 2010, 43, 137–145. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Sideri, S.; Papageorgiou, S.N.; Eliades, T. Registration in the international prospective register of systematic reviews (PROSPERO) of systematic review protocols was associated with increased review quality. J. Clin. Epidemiology 2018, 100, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Zhang, Y.; Sun, Y.; Kong, M.; Han, S.; Wang, C.; Wang, Y.; Xu, D.; Tu, Q.; Zhu, K.; et al. Inhibition of RhoA/MRTF-A signaling alleviates nucleus pulposus fibrosis induced by mechanical stress overload. Connect. Tissue Res. 2022, 63, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, B.; Liu, Z.-H.; Song, W.; Wang, D.; Chen, B.-Y.; Fan, J.; Xu, Z.; Geng, D.; Luo, Z.-J. Notochordal-Cell-Derived Exosomes Induced by Compressive Load Inhibit Angiogenesis via the miR-140-5p/Wnt/β-Catenin Axis. Mol. Ther. Nucl. Acids 2020, 22, 1092–1106. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Würtz, K.; Liedert, A.; Schmidt, C.; Börm, W.; Ignatius, A.; Wilke, H.-J.; Claes, L. A three-dimensional collagen matrix as a suitable culture system for the comparison of cyclic strain and hydrostatic pressure effects on intervertebral disc cells. J. Neurosurgery: Spine 2005, 2, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Frain, J.; Fotheringham, A.P.; Freemont, A.J.; Hoyland, J.A. Human cells derived from degenerate intervertebral discs respond differently to those derived from non-degenerate intervertebral discs following application of dynamic hydrostatic pressure. Biorheology 2008, 45, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Frain, J.; Millward-Sadler, J.; Fotheringham, A.P.; Freemont, A.J.; Hoyland, J.A. Altered integrin mechanotransduction in human nucleus pulposus cells derived from degenerated discs. Arthritis Rheum. 2009, 60, 460–469. [Google Scholar] [CrossRef]
- Aladin, D.M.K.; Cheung, K.; Chan, D.; Yee, A.F.Y.; Jim, J.J.T.; Luk, K.D.K.; Lu, W.W. Expression of the Trp2 Allele of COL9A2 is Associated with Alterations in the Mechanical Properties of Human Intervertebral Discs. Spine 2007, 32, 2820–2826. [Google Scholar] [CrossRef]
- Franco-Obregón, A.; Cambria, E.; Greutert, H.; Wernas, T.; Hitzl, W.; Egli, M.; Sekiguchi, M.; Boos, N.; Hausmann, O.; Ferguson, S.J.; et al. TRPC6 in simulated microgravity of intervertebral disc cells. Eur Spine J. 2018, 27, 2621–2630. [Google Scholar] [CrossRef]
- Cambria, E.; Arlt, M.; Wandel, S.; Krupkova, O.; Hitzl, W.; Passini, F.; Hausmann, O.; Snedeker, J.; Ferguson, S.; Wuertz-Kozak, K. TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells. Cells 2020, 9, 1736. [Google Scholar] [CrossRef]
- Likhitpanichkul, M.; Torre, O.M.; Gruen, J.; Walter, B.A.; Hecht, A.C.; Iatridis, J.C. Do mechanical strain and TNF-α interact to amplify pro-inflammatory cytokine production in human annulus fibrosus cells? J. Biomech. 2016, 49, 1214–1220. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Nagra, N.S.; Freemont, A.J.; Millward-Sadler, S.J.; Hoyland, J.A. Integrin—Dependent Mechanotransduction in Mechanically Stimulated Human Annulus Fibrosus Cells: Evidence for an Alternative Mechanotransduction Pathway Operating with Degeneration. PLoS ONE 2013, 8, e72994. [Google Scholar] [CrossRef]
- Gilbert, H.T.; Hoyland, J.A.; Freemont, A.J.; Millward-Sadler, S.J. The involvement of interleukin-1 and interleukin-4 in the response of human annulus fibrosus cells to cyclic tensile strain: An altered mechanotransduction pathway with degeneration. Thromb. Haemost. Res. Ther. 2011, 13, R8-12. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, H.T.J.; Hoyland, J.A.; Millward-Sadler, S.J. The response of human anulus fibrosus cells to cyclic tensile strain is frequency-dependent and altered with disc degeneration. Arthritis Rheum. 2010, 62, 3385–3394. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, H.-Q.; Zhang, Q.; Yan, X.-D.; Hao, M.; Luo, Z.-J. Alterations of ADAMTSs and TIMP-3 in human nucleus pulposus cells subjected to compressive load: Implications in the pathogenesis of human intervertebral disc degeneration. J. Orthop. Res.Off. Publ. Orthop. Res. Soc. 2012, 30, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Liedert, A.; Wuertz, K.; Buser, Z.; Rinkler, C.; Käfer, W.; Ignatius, A.; Claes, L.; Roberts, S.; Johnson, W.E.B. Mechanical Stimulation Alters Pleiotrophin and Aggrecan Expression by Human Intervertebral Disc Cells and Influences Their Capacity to Stimulate Endothelial Cell Migration. Spine 2009, 34, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Hodson, N.W.; Patel, S.; Richardson, S.M.; Hoyland, J.A.; Gilbert, H.T.J. Degenerate intervertebral disc-like pH induces a catabolic mechanoresponse in human nucleus pulposus cells. JOR Spine 2018, 1, e1004. [Google Scholar] [CrossRef] [PubMed]
- Fearing, B.V.; Hernandez, P.A.; Setton, L.A.; Chahine, N.O. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef]
- Chan, S.C.W.; Ferguson, S.J.; Gantenbein-Ritter, B. The effects of dynamic loading on the intervertebral disc. Eur. Spine J. 2011, 20, 1796–1812. [Google Scholar] [CrossRef]
- Kadow, T.; Sowa, G.; Vo, N.; Kang, J.D. Molecular Basis of Intervertebral Disc Degeneration and Herniations: What Are the Important Translational Questions? Clin. Orthop. Relat. Res. 2015, 473, 1903–1912. [Google Scholar] [CrossRef]
- Richardson, S.M.; Doyle, P.; Minogue, B.M.; Gnanalingham, K.; Hoyland, J.A. Increased expression of matrix metalloproteinase-10, nerve growth factor and substance P in the painful degenerate intervertebral disc. Thromb. Haemost. Res. Ther. 2009, 11, R126-8. [Google Scholar] [CrossRef]
- Purmessur, D.; Walter, B.; Roughley, P.; Laudier, D.; Hecht, A.; Iatridis, J. A role for TNFα in intervertebral disc degeneration: A non-recoverable catabolic shift. Biochem. Biophys. Res. Commun. 2013, 433, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Daly, C.; Ghosh, P.; Jenkin, G.; Oehme, D.; Goldschlager, T. A Review of Animal Models of Intervertebral Disc Degeneration: Pathophysiology, Regeneration, and Translation to the Clinic. BioMed Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Author | Study Aim | Samples | Age (Mean ± sd) Gender | Cell Type | Experimental Model | Molecular Pathway Involved | Mechanical Stimulus Applied | Observed Response | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Song et al. 2022 [32] | Analysis of MMP-12, CTGF and collagen I, III transcriptional regulation to assess the role of RhoA/MRTF-A signaling pathway in the development of IDD | NP tissues from lumbar spine of 25 human patients 15 patients with IVD 10 control patients with spinal cord injury | IVD patients: 55.5 ± 5.1 years -5 males -10 females Control group: 39.2 ± 7.6 years -4 males -6 females | NP | Human NP cells cultured on 2D flexible silicone Elastomer substrate and stretched on a Flexcell apparatus | RhoA/ROCK2– MRTF-A | CASES: 15% cyclic strain at a frequency of 6 cycles/min (cycle consist of 3 s of stretching alternating with 3 s of relaxation). Cyclic stretching was performed for 24 h. (Hyperphysiological mechanical loading) CONTROLS: NO mechanical strain | Cyclic tensile strain (CTS) of NP cells ↑ Nuclear translocation of MRTF-A ↑ RhoA, ROCK2, MRTF-A, SRF, MMP-12, and CTGF ↓ Aggrecan and collagen II | ||
| CTS in NP cells + MRTF-A inhibitor (CCG-1423) ⊗ RhoA signaling inhibited ⊗ MRTF-A transcription inhibited ⊗ Aggrecan and collagen II downregulation inhibited ↓ SRF, MMP-12, and CTGF | ||||||||||
| Sun et al. 2020 [33] | To evaluate the role of NCs produced exosomes in the homeostasis of intervertebral disc and their link with mechanical load and disc angiogenesis | NP tissues from 20 human patients 10 IDD patients 10 AIS patients | IDD patients: 46.2 years (34–68) AIS patients: 19.3 years (16–24) | NC and NP cells | NC cell 2D culture samples subjected to controllable compressive stress at 0 MPa, 0.5 MPa, and 1.0 MPa for 24 and 48 h | miR-140-5p– Wnt/b-catenin axis | 0 MPa, 0.5 MPa, and 1.0 MPa for 24 and 48 h | / | ||
| Cambria et al. 2020 [34] | To investigate the role of TRPV4 in human AF cells stretch-induced inflammation | 9 human IVD biopsies retrieved from patients undergoing spinal surgery for disc herniation or DDD | 56.1 ± 20.5 years -4 males -5 females | AF | 2D cultures in chambers mounted on commercial stretching bioreactor (STB-140-10, STREX) | TRPV4 ion channels | CASES: 20% cyclic sinusoidal uniaxial strain, frequency 1 Hz, cells were stretched for 1,2,4,8,12 and 24 h. (Hyperphysiological mechanical loading) CONTROLS: same condition without strain | CTS in AF cells ↑ IL-6, IL-8, COX2 and MMP-1. PGE2 release ↑ p38 and ERK phosphorylation (phosphorylated MAPKs) ↓ Aggrecan and collagen II Activation of p38 | ||
| CTS in AF cells + pharmacological inhibition of TRPV4 ⊗ p38 stretch-induced phosphorylation is inhibited | ||||||||||
| CTS in AF cells + CRISPR-Cas9 Knock-Out of TRPV4 ⊗ IL-8, IL-6 and MMP-1 increase inhibited No changes in gene expression of COX2 | ||||||||||
| Franco-Obregon et al. 2018 [35] | To evaluate the effects of simulated microgravity and pharmacological inhibition of TRPC channel on human IVD cells | 16 human IVD cells retrieved from surgical biopsies composed of both AF and NP 15 patients affected by DDD in the lumbar spine 1 patient affect by DDD in the cervical spine | 51.5 ± 10.5 years -5 males -8 females -3 unknown | Both NP and AF | 2D Culture chambers mounted on gimbal frames that simulated microgravity | TRPC6 | Simulated microgravity (Hypophysiological mechanical loading) | ↓ Cell proliferation ↑ Cell senescence and accumulation of cells in G2/M | ||
| Hodson et al. 2018. [36] | To evaluate the role of mechanical and chemical IVD microenvironment on disc cells | NP cells from 3 human patients 2 human cadavers (IVD) 1 patient who underwent spinal surgery following trauma of the spine | Cadavers: 2 males aged 36 and 63 Spinal surgery patient: female aged 47 | NP | Precultured NP cells embedded in agarose constructs and compressed using a Flexcell FX4000 Compression System (Flexcell International) | Catabolic and anabolic genes (multiple genes, mainly aggrecan and MMP-3) | CASES: 0.004 MPa, 1.0 Hz and 1 h of compression under pH conditions representative of nondegenerate (pH 7.1) and degenerate (pH 6.5) IVDs (Physiological mechanical loading) CONTROLS: uncompressed | Non-degenerative IVD conditions–pH (7.1) ↑ AGC, TIMP1, MMP-3, ADAMTS5 (increase in both anabolic and catabolic gene expression) AGC expression is modulated by integrin receptors, the presence of RGD peptides prevented the AGC expression increasing | ||
| Degenerative IVD conditions–pH (6.5) ↑ MMP-3 (loss of mechanoregulation of anabolic gene expression) AGC gene expression was not mechanoresponsive at pH 6.5. The increase in MMP-3 expression did not appear to be RGD-integrin dependent | ||||||||||
| Likhitpanichkul et al. 2016 [37] | To assess the interaction between TNF-α and tensile strain levels on AF cells in vitro. To test the effectiveness of anti-TNF-α drug and anti-IL-6 drug on AF cells | AF cells from 5 human IVD derived from spinal surgeries and autopsies | 50 ± 16 years | AF | AF cells 2D cultures in silicone-membrane chambers (STREX, B-Bridge, Cupertino, CA) | Pro-inflammatory cytokines (TNF-α, IL-1β, IL-8 and IL-6) and cytoskeletal changes (F-actin, vinculin, and α-tubulin) | CASES: TNF-α + physiological (5%) and pathological (15%) cyclic tensile uniaxial strain at 0.5 hz for 24 h CONTROLS: unstrained | ↑ IL-1β and IL-8 at 15% compared to 5% strain. ↑ F-actin ↑ Cell roundedness ↓ Microtubule network (α-tubulin) Infliximab non- significantly decreased IL-1β and IL-8 levels compared to TNF-α stimulated AF cells undergoing 15% strain, and significantly decreased IL-1β and IL 8 levels compared to anti-IL-6 treatment | ||
| Gilbert et al. 2013 [38] | To investigate the role of integrins in the mechanoresponse of human AF cells | 6 human AF samples derived from surgical procedures for DDD and autopsies 3 classified as degenerative 3 classified as non-degenerative | Degenerative samples: 41 years (29–49) Non-degenerative samples: 47 years (37–57) | AF | 2D cultures on Silicone membranes stretched by using the FX4000 Flexercell Tension System (Flexcell International, Hillsborough, NC, USA)) | MMP-3, ADAMTS-4, aggrecan and collagen I | CASES: cyclic tensile strain with and without function blocking RGD –peptides with 10% strain, 1.0 Hz for 20 min (Physiological mechanical loading) CONTROLS: unstrained | RAD or RGD peptide treatment in unstrained cells had no effect on gene expression | ||
| CTS in non-degenerative AF cells + RAD peptides ↓ ADAMTS-4 ↑ FAK phosphorylation No effect on collagen I gene expression | CTS in degenerative AF cells + RAD peptides ↓ Collagen I No effect on ADAMTS-4 gene expression No effect on levels of FAK phosphorylation | |||||||||
| CTS in non-degenerative AF cells + RGD peptides ↑ ADAMTS-4 ⊗ CTS induced increase in FAK phosphorylation is inhibited No effect on collagen I gene expression | CTS in degenerative AF cells + RGD peptides ↓ Collagen I No effect on ADAMTS-4 gene expression No effect on levels of FAK phosphorylation | |||||||||
| Gilbert et al. 2011 [39] | To investigate the role of IL-1 and IL-4 in mechanotransduction. of human AF cell To evaluate the role of proinflammatory cytokines in the different cellular responses of non-degenerative and degenerative AF cells. | 6 human AF samples 3 classified as degenerative (2 from patients who underwent spinal surgeries for DDD and 1 from cadaver, respectively) 3 postmortem non-degenerative | Degenerative samples: 50 years (29–66) Non-degenerative samples: 47 years (37–57) | AF | Silicone membranes within the Bioflex culture plates using the FX4000 Flexercell Tension System (Flexcell International) | MMP-3, ADAMTS-4, aggrecan and collagen I | CASES: cyclic tensile strain with and without IL-1Ra or IL-4RAb with 10% strain, 1.0 Hz for 20 min (Physiological mechanical loading) CONTROLS: unstrained | CTS in non-degenerative AF cells + IL-1Ra ↑ ↑ MMP-3 | CTS in degenerative AF cells + IL-1Ra ↓↓ Collagen I | |
| CTS in non-degenerative AF cells + IL-4RAb ↓ MMP-3 | CTS in degenerative AF cells + IL-4RAb ↓↓ Collagen I | |||||||||
| CTS in non-degenerative AF cells without IL-1Ra or IL-4Rab ↓ ADAMTS-4 and MMP-3 No change in the relative gene expression of aggrecan or collagen I | CTS in degenerative AF cells without IL-1Ra or IL-4RAb ↓ Aggrecan and collagen I No change in the relative gene expression of MMP-3 or ADAMTS-4 | |||||||||
| Huang et al. 2011 [40] | To investigate the role of ADAMTSs and TIMP-3 in NP cells under mechanical stress. | 30 human NP samples 15 patients with DDD 15 patients with idiopathic scoliosis (controls) | Patients with DDD: 30.2 years (27–28) Patients with idiopathic scoliosis: 18.7 years (16–24) | NP | NP tissue explants stimulated in BioPress compression culture plates using the Flexercell Compression System (FX-4000C; Flexcell International) | ADAMTSs, aggrecan and TIMP-3 | CASES: Compressive load of 0.35–0.95 MPa at 1 Hz for 2 h twice a day for 7 days. 1 h prior to load, samples were treated with or without TIMP-3 (Physiological mechanical loading) CONTROLS: uncompressed | Compressive loading without TIMP-3 ↑ ADAMTS-1, 4, and 5 ↓ Aggrecan | ||
| Compressive loading + TIMP-3 ↑↑ Aggrecan | ||||||||||
| Gilbert et al. 2010 [41] | To investigate the effect of CTS on AF cells derived from both degenerative and non-degenerative IVDs | 6 human AF samples 2 surgical samples classified as degenerative 4 postmortem samples classified as non-degenerative | Degenerative: 2 patients aged 29 and 66 Non-degenerative: 49.3 years (37–57) | AF | 2D cultures of AF cells on Silicone membranes stretched by the FX4000 Flexercell Tension System (Flexcell International) | MMP-3, ADAMTS-4, aggrecan and collagen I | CASES: application of 20% CTS for 20 min at a frequency of 1.0 Hz or 0.33 Hz (Physiological and less than physiological mechanical loading frequency respectively) CONTROLS: unstrained | CTS at 1 Hz in non-degenerative AF cells ↓ ADAMTS-4 and MMP-3 | CTS at 0.33 Hz in non-degenerative AF cells ↓ Collagen I and II | |
| CTS at 1 Hz in degenerative AF cells ↓ Aggrecan and Collagen I | CTS at 0.33 Hz in degenerative AF cells ↓ Collagen I, II and aggrecan | |||||||||
| Neidlinger-Wilke et al. 2009 [42] | To assess the effects of mechanical load on expression of pleiotropin (PTN) and aggrecan in IVD cells and to determine the factors influencing endothelial cells migration. | Human IVD cells isolated from discs of 11 patients 8 disc degenerations 3 underwent surgery for trauma 1 scoliosis | 44.1 ± 18.3 years -6 males -5 females | Both NP and AF | Cells embedded in collagen gels exposed to strain (AF) or hydrostatic pressure (NP) | PTN, aggrecan | CASES: 4% CTS at 1 Hz for AF cells (n 6). Hydrostatic pressure (0.25 or 2.5 MPa; 0.1 Hz) for NP cells (n 11). (Physiological mechanical loading) CONTROLS: unstrained/unloaded | CTS on AF cells for 1 h ↑↑ PTN | Hydrostatic pressure on NP cells for 1 h ↑ PTN | |
| CTS on AF cells for 24 h ↑↑ Aggrecan No difference in PTN expression between loaded and unloaded cells | Hydrostatic pressure on NP cells for 24 h ↓ Aggrecan ↑ Endothelial cell migration/ adhesion No difference in PTN expression between loaded and unloaded cells | |||||||||
| Le Maitre et al. 2009 [43] | To assess the role of integrin signaling in IVD cells exposed to mechanical stimulation. | 24 human IVD samples 15 surgical samples classified as degenerative 9 postmortem samples classified as non-degenerative | Degenerative patients: 56.2 ± 15.8 years -10 males -4 females -1 unknown Non-degenerative patients: 43.8 ± 8.5 years -6 males | NP | NP cells seeded in alginate constructs loaded in BioFlex compression plates and stimulated with a Flexercell FX-4000 C compression loading system (Flexcell International) | Aggrecan | CASES: compression loading at 0.35–0.95 MPa at 1 Hz applied for 2 h. 1 h prior to mechanical loading, constructs were either untreated, treated with RGD integrins inhibitory peptide or with a control peptide. (Physiological mechanical loading) CONTROLS: uncompressed | Compressive loading of degenerative and non-degenerative NP cells + control peptide ↓ Aggrecan | Compressive loading of non-degenerative NP cells + RGD-integrin ligand site inhibitor ⊗ load-induced decrease in expression of aggrecan gene. | |
| Compressive loading of degenerative NP cells + RGD-integrin ligand site inhibitor ↓ Aggrecan | ||||||||||
| Le Maitre et al. 2008 [44] | To investigate the effect of physiological hydrostatic pressure on NP and AF cells. | Human IVD cells isolated from discs of 15 patients 7 surgical 8 postmortem | Surgical samples: 43.8 ± 15.6 -6 males -1 female Postmortem samples 74.8 ± 0.5 -8 males | NP | Alginate NP cell cultures in Sterile Whirl-Pak bags placed within a pressure vessel attached to a piston | Catabolic and anabolic genes (multiple genes, mainly aggrecan and MMP-3) | CASES: dynamic hydrostatic pressure at 0.8–1.7 MPa at 0,5 Hz for 2 h. (Physiological mechanical loading) CONTROLS: uncompressed | HP on degenerative and non-degenerative AF cells No effect on gene expression | ||
| HP on non-degenerative NP cells ↑↑ Aggrecan, c-fos ↑ Sox-9, collagen II No effect on MMP-3 | HP on degenerative NP cells No effect on gene expression | |||||||||
| Aladin et al. 2007 [45] | To compare the mechanical properties of human NP tissue in those that carry the Trp2 allele to those without, before any sign of disc degeneration. | 12 non-degenerated IVD samples from patients who underwent scoliosis surgery (6 Trp2+ samples and 6 Trp2- samples). 6 degenerated IVD samples from patients who underwent anterior discectomy (Trp2-). | Non-degenerated samples (Trp2+): mean age 15.2 years Non degenerated samples (Trp2-): mean age 18.5 years Degenerated samples mean age 37.67 years | NP | Frozen tissue explants stimulated in a compression apparatus composed of non-porous chamber made of Teflon, placed coaxially to an upper rigid and porous platen having 50% porosity with an average pore size of 50 mm | COL9A2 Trp2 allele | NP samples were tested in confined compression. Swelling pressure and compressive moduli were measured and compared between groups (harboring or not the COL9 Trp2 allele) | / | ||
| Neidlinger-Wilke et al. 2005 [46] | To evaluate the response of IVD cells, in AP and NF cells as well, to different mechanical stimuli in terms of mRNA expression of anabolic and catabolic matrix proteins | Human IVD cells isolated from discs of 18 patients surgically treated for DDD | 41.1 ± 12.0 years -6 males -5 females | Both NP and AF | 2D cultures in Silicone dishes (cyclic strain experiments) or into standard culture dishes (hydrostatic pressure experiments) | Catabolic and anabolic genes (multiple genes, mainly aggrecan and MMP-3) | STRAIN CASES: CTS of 1, 2, 4, and 8% at a frequency of 1 Hz for 24 h HYDROSTATIC PRESSURE CASES: 0.25 MPa at a frequency of 0.1 Hz for 30 min (Physiological mechanical loading) CONTROLS: unstrained and uncompressed | HP on NP cells ↑ Proliferation ↑ Aggrecan, collagen I ↓ MMP-2, MMP-3 | NP cells without HP or CTS ↑ Proliferation | |
| HP on AF cells ↑ Collagen I ↓ MMP-3, aggrecan | ||||||||||
| CTS on NP cells ↑ Collagen I ↓ MMP-2, MMP-3 | ||||||||||
| CTS on AF cells ↑ Aggrecan, collagen II ↓ MMP-3 | ||||||||||
| Author | Key Findings |
|---|---|
| Song et al. 2022 [32] | The RhoA/MRTF-A signaling pathway is activated by mechanical stress and promotes ECM degeneration in the NP. The RhoA/MRTF-A axis regulates MMP-12 and CTGF expression. NP degeneration caused by mechanical stress overload is mitigated by the RhoA/MRTF-A inhibitor CCG-1423, which has the potential therapeutic effect of NPCs’ functional recovery facilitation. |
| Sun et al. 2020 [33] | HUVECs’ migration is strongly inhibited by 0.5 Mpa/NC-exos, showing a dose-dependent effect. Thus, NC-exos induced by compressive load have the potential to inhibit endothelial cell angiogenesis. The anti-angiogenic role of 0.5 Mpa/NC-exos appears to be mediated by miRNAs transferred from NCs to endothelial cells via NC-exos. Among these miRNAs, the upregulation of miR-140-5p shows a significant inhibitive effect on angiogenesis in endothelial cells via the downregulation of Wnt11 expression and inhibition of b-catenin nuclear accumulation. |
| Cambria et al. 2020 [34] | A TRPV4 antagonist has shown the capacity to lessen the mechanoflammation caused by hyperphysiological CTS, thus revealing the novel mechanoinflammatory role of TRPV4 in human primary AF cells. TRPV4 mediates stretch-induced inflammation, possibly via the activation of the p38 MAPK pathway. TRPV4 KO via CRISPR-Cas9 prevents the stretch-induced upregulation of IL8 mRNA and tends to reduce the stretch-induced IL6 mRNA. |
| Franco-Obregon et al. 2018 [35] | IVD cells exposed to simulated microgravity or to TRPC channel inhibition showed reduced proliferation and increased senescence. The TRPC6 gene expression was reduced in cells subjected to simulated microgravity. Mechanotransduction, cell proliferation and senescence regulation in IVDs may be partially controlled by TRPC6 ion channels. |
| Hodson et al. 2018 [36] | NP cells cultured both at pH 7.1 and 6.5 showed similar gene expression patterns. NP cells’ response to compression shifted from matrix homeostasis at a pH of 7.1, representative of non-degenerated tissue, to matrix catabolism at a pH of 6.5, representative of degenerated tissue. Thus, a catabolic shift in human NP cell phenotype occurred under acidic pH conditions. The aberrant mechanotransduction induced by an acidic environment may have a potential role in the progression of IVD degeneration. |
| Likhitpanichkul et al. 2016 [37] | TNF-α-treated AF cells showed increased IL-1β and IL-8 production and cytoskeletal network modification with enhanced stress fiber formation from actin polymerization and microtubule network disruption; these cytoskeletal changes reflect increased mechanosensitivity. Pro-inflammatory cytokines produced by IVD cells may be a key factor in the inflamed environment associated with IVD degeneration, potentially altering physiological IVD mechanobiology. |
| Gilbert et al. 2013 [38] | RGD-recognizing integrins may not be involved in the altered mechanoresponse of AF cells derived from degenerate IVDs. The aberrant response in degenerate cells may be recognized by different mechanoreceptors via an alternative mechanotransduction pathway. The involvement of RGD integrins in mechanoresponses appeared to be lost with degeneration. The findings suggested FAK activation is caused by CTS via an RGD-integrin-dependent pathway. The ADAMTS-4 decreased gene expression seen in CTS-stimulated non-degenerated AF cells may occur through an integrin-, and potentially FAK-, dependent mechanism. Non-degenerated AF cells’ mechanoresponse to CTS appears to be mediated by RGD-recognizing integrins. The aforementioned cells showed the integrin-dependent phosphorylation of FAK in response to strain stimulation. RGD integrins and FAK may not be involved in the mechanoresponse of degenerated AF cells. |
| Gilbert et al. 2011 [39] | For the first time, immunopositivity for the IL-4r subunits IL-4Ra and IL2Rg and immunonegativity for the receptor subunit IL-13Ra1 was described in human IVD cells. Stimulated with CTS, AF cells derived from non-degenerative IVDs showed a shift to a less catabolic phenotype. The reduced catabolic response appeared to be IL-1- and IL-4-dependent. The increment in MMP-3 gene expression may be correlated with its decreased activity (negative feedback regulation). The inhibition of MMPs’ activity due to IL-1Ra treatment seems to have determined increased MMP gene expression in IVD cells, outlining the potential of targeting IL-1R in DDD treatment. The findings show the potential role of IL-1, not only in a catabolic shift typical of DDD, but also in transducing physiological mechanical stimuli, leading to tissue remodeling. Neither IL-1 nor IL-4 appeared to be necessary for the matrix remodeling mechanoresponse of AF cells derived from degenerative IVDs. AF cells derived from degenerative and non-degenerative tissue regulated matrix protein and matrix-degrading enzyme gene expression differently; this difference may be determined by altered cytokine-dependent transcription factor activation. |
| Huang et al. 2011 [40] | Compressive load, with the increase in ADAMTS-1, 4, 5 expression, appears to be a key factor in aggrecan depletion in human NP cells, which might have an etiologic implication in the development of IDD. The physiological expression of ADAMTS-1, 4, 9, 15 and TIMP-3 in non-degenerative discs indicates a possible role for these proteins in the normal turn-over of aggrecan and other matrix molecules in the healthy disc matrix. The imbalance between ADAMTSs and their inhibitor (TIMP-3) could play a role in the pathogenesis of IDD. TIMP-3 might be a potent therapeutic target preventing aggrecan loss during IDD. |
| Gilbert et al. 2010 [41] | The response of AF cells derived from both degenerative and non-degenerative IVDs to CTS appeared to be frequency-dependent. At 1 Hz of CTS (similar to normal physiologic motion), AF cells derived from non-degenerative tissue showed a shift to a less catabolic phenotype. The 0.33 Hz of CTS resulted in a catabolic response. The response of AF cells to CTS was both frequency-dependent and dependent on whether the cells are derived from degenerated or non -degenerated tissue. AF cells derived from degenerated tissue not only lost their ability to downregulate matrix enzyme gene expression in response to a 1.0 Hz frequency of CTS but also responded by reducing their matrix protein gene expression. Matrix homeostasis and matrix anabolism are promoted by physiological mechanical loads in healthy disc cells. Frequencies below physiological levels could lead to the degradation of the IVD matrix. On the contrary, physiologic mechanical loads could be detrimental to disc cell matrix homeostasis in degenerated IVDs, potentially leading to the progression of DDD. |
| Neidlinger-Wilke et al. 2009 [42] | Relative expression levels of PTN in human IVD cells are influenced by mechanical load. Increased PTN expression may have pro-angiogenic effects, which could be either enforced or impaired by the down- or upregulation of the antiangiogenic protein aggrecan. PTN expression showed high variability between different patients; this variation may be attributable to different degrees of disc degeneration. Mechanical load influences the presence of angiostatic and angiogenic factors within the IVD. |
| Le Maitre et al. 2009 [43] | In non-degenerated IVDs, RGD-binding integrins contribute to the mechanotransduction response. Mechanotransduction pathways appeared to be altered in NP cells derived from degenerative IVDs. Mechanosensing in NP cells from non-degenerative discs occurred via RGD integrins, possibly via the a5b1-integrin, while cells from degenerated discs might use a different receptor to sense and respond to mechanical signaling. Since the a5b1 integrin is physiologically expressed in degenerated IVDs, with no change in expression observed during disc degeneration, the cited mechanosensing could occur through a non-RGD integrin. |
| Le Maitre et al. 2008 [44] | IVD cells modify matrix homeostasis in response to physiological hydrostatic pressure. Cells obtained from healthy IVD tissues respond to dynamic HP by upregulating the gene expression of anabolic genes, indicative of healthy matrix homeostasis. Altered mechanotransduction pathways may be operational in degenerative IVD tissues. |
| Aladin et al. 2007 [45] | A non-degenerated (COL9 A2 Trp2+) control group appeared to have an increasing swelling stress pattern. Degenerated (COL9 A2Trp2-) groups exhibited decreasing swelling stress patterns. The non-degenerated (Trp2+) and degenerated (Trp2-) groups’ swelling pressure and compressive modulus were considerably lower than those of the non-degenerated (Trp2-) controls. A significant difference occurred in the swelling pressure (p = 0.006) and compressive modulus (p = 0.025) between the non-degenerated (Trp2-) controls and (Trp2+) cases. The swelling pressure difference between degenerated (Trp2+) controls and (Trp2-) cases was not significant (p = 0.34), suggesting that the Trp2 allele is related with a decrease in the swelling pressure to an extent approximately similar to but slightly better than that of the degenerated discs. The Trp2 allele seems to be linked to IVD mechanical properties and consequently to disc degeneration pathomechanism. Alterations in collagen IX result in cross-linking abnormalities between collagen IX and collagen II, thus reducing the ability of the collagen network to hold the ECM together, manifested as changes in the compression modulus and swelling pressure. The Trp2 allele is associated with deteriorating mechanical properties of the nucleus pulposus as early as adolescence, with no other signs of disc degeneration. |
| Neidlinger-Wilke et al. 2005 [46] | CTS appeared to have an anabolic effect both in AF and NP cells, apparently without differences among different strain magnitudes. The application of mechanical stimuli had no negative effect on IVD cells. Hydrostatic pressure also appeared to determine an anabolic effect in both AP and NP cells. Anulus cells were more responsive to cyclic strain, whereas nucleus cells were more responsive to hydrostatic pressure. Mechanically stimulated disc cells, either subjected to hydrostatic pressure or cyclic strain, tended to decrease the mRNA expression of MMP-2 and -3. These findings seem to suggest that moderate mechanical loading may prevent disc matrix degradation via the suppression of cellular production of MMPs. Cells obtained from different donor patients showed high inter-individual variations, which could be influenced by the degree of degeneration of the disc samples. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruffilli, A.; Viroli, G.; Neri, S.; Traversari, M.; Barile, F.; Manzetti, M.; Assirelli, E.; Ialuna, M.; Vita, F.; Faldini, C. Mechanobiology of the Human Intervertebral Disc: Systematic Review of the Literature and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 2728. https://doi.org/10.3390/ijms24032728
Ruffilli A, Viroli G, Neri S, Traversari M, Barile F, Manzetti M, Assirelli E, Ialuna M, Vita F, Faldini C. Mechanobiology of the Human Intervertebral Disc: Systematic Review of the Literature and Future Perspectives. International Journal of Molecular Sciences. 2023; 24(3):2728. https://doi.org/10.3390/ijms24032728
Chicago/Turabian StyleRuffilli, Alberto, Giovanni Viroli, Simona Neri, Matteo Traversari, Francesca Barile, Marco Manzetti, Elisa Assirelli, Marco Ialuna, Fabio Vita, and Cesare Faldini. 2023. "Mechanobiology of the Human Intervertebral Disc: Systematic Review of the Literature and Future Perspectives" International Journal of Molecular Sciences 24, no. 3: 2728. https://doi.org/10.3390/ijms24032728