
Recent Advances and Future Potential of Long Non-Coding RNAs in Insects

Key Laboratory of Bio-Pesticide Innovation and Application of Guangdong Province, College of Plant Protection, South China Agricultural University, Guangzhou 510642, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(3), 2605; https://doi.org/10.3390/ijms24032605
Submission received: 8 December 2022
/
Revised: 28 December 2022
/
Accepted: 4 January 2023
/
Published: 30 January 2023
(This article belongs to the Special Issue Hormonal/Noncoding RNA Regulation in Invertebrate Models)
Abstract
:Over the last decade, long non-coding RNAs (lncRNAs) have witnessed a steep rise in interest amongst the scientific community. Because of their functional significance in several biological processes, i.e., alternative splicing, epigenetics, cell cycle, dosage compensation, and gene expression regulation, lncRNAs have transformed our understanding of RNA’s regulatory potential. However, most knowledge concerning lncRNAs comes from mammals, and our understanding of the potential role of lncRNAs amongst insects remains unclear. Technological advances such as RNA-seq have enabled entomologists to profile several hundred lncRNAs in insect species, although few are functionally studied. This article will review experimentally validated lncRNAs from different insects and the lncRNAs identified via bioinformatic tools. Lastly, we will discuss the existing research challenges and the future of lncRNAs in insects.
1. Introduction
With the inception of high-throughput genomic technologies, including microarray and next-generation sequencing (NGS), researchers have detected several RNA transcripts in particular tissues or cells at a precise time. Apart from the conventional messenger RNAs (mRNAs) that translate into functional proteins, RNA-sequencing (RNA-seq) has put the spotlight on various classes of non-coding RNAs (ncRNAs) [1]. Once believed to be transcriptional noise, junk RNAs, or the by-products of genetic operations, ncRNAs have emerged as promising biomarkers [2]. The ncRNAs comprise the largest class of RNAs and are classified into small ncRNAs (sncRNAs) and long ncRNAs (lncRNAs) [3]. In the past decade, lncRNAs have achieved global interest due to their potential role in numerous biological processes, including alternative splicing, cell cycle, epigenetics, dosage compensation, and gene expression regulation [4,5].
2. Historical Overview
In the 1980s, the intricate mechanism of genomic imprinting was discovered, an epigenetic process that determines gene expression based on whether they are inherited from the father or the mother. Two imprinted genes: Igf2r (encoding the receptor for insulin-like growth factor type-2), the paternally expressed protein-coding gene, and the maternally expressed H19, were identified in mice. These genes located on chromosome 7 in close proximity, formed the H19/IGF2 cluster [8,9]. However, the surprising lack of translation in H19 continued to puzzle the researchers until a functional description of an additional lncRNA, Xist (X-inactive-specific transcript), involved in dosage compensation among mammals [10,11] was performed. Based on this information, H19 can be considered the first eukaryotic non-coding gene ever discovered; however, it was initially described as mRNA [12]. The ground-breaking discoveries of H19 and Xist transformed our perspective of ncRNAs and their functional relevance.
However, as the traditional biological methods were not powerful enough, the research concerning lncRNAs remained steady until the dawn of the 21st century, when the FANTOM project revealed several putative lncRNA transcripts by employing full-length cDNA cloning [13]. This evidence rejuvenated the scientific community’s interest, which led to the development of several methods describing the lncRNAs, including the Tiling array and ChIP-Seq [14,15,16]. Figure 1 illustrates some of the significant discoveries made in the field of lncRNAs. Despite these technological advances, rapid progress in insects was made after the development of RNA-seq [17], and several lncRNAs were discovered and cataloged from the Drosophila melanogaster genome [18].
3. Biogenesis and Classification
Although lncRNAs are classified as ncRNAs, they are distinctly different from other types of ncRNAs; hence, it is essential to understand their biogenesis for functional significance. LncRNAs are described as RNA molecules with a length of ≥200 nucleotides (nt) that lack protein-coding ability [19,20]. Similar to mRNAs, lncRNAs are transcribed by RNA polymerase (RNAP) II/III, spliced and capped at the 5` [21], polyadenylated, and contain exons (which are longer but fewer in contrast to mRNAs) [22].
Researchers have established that different types of lncRNAs are transcribed from numerous DNA elements such as promoters, enhancers, and intergenic regions [23]. Unlike other classes of ncRNAs, lncRNAs benefit from secondary and 3D structures, enabling them to have RNA and protein-like functions [24]. The lncRNAs are more highly enriched in the nucleus than in the cytoplasm. For instance, in humans, 17% of lncRNAs are enriched in the nucleus compared to 4% in the cytoplasm [25]. However, multiple lncRNAs have been detected that are shuttled into the cytoplasm after synthesis in the nucleus and mediate numerous modes of gene regulation [26,27]. The cytoplasmic lncRNAs play crucial roles, including mRNA translation regulation, mRNA stability, microRNA (miRNA) precursors, and miRNA sponges/decoys by sequestering miRNAs and reducing their regulatory effects on target mRNAs [28,29,30,31]. In addition, lncRNAs are released into extracellular space as circulating lncRNAs, which could potentially be biomarkers [32]. The biogenesis of lncRNAs is very complicated, and additional in-depth investigations are required for a better understanding [33,34]. Even though lncRNAs and mRNAs share various characteristics, several types of lncRNAs are unique in biogenesis, form, and function [35]. Unlike mRNAs, lncRNAs are expressed in a highly spatiotemporal pattern and at substantially lower levels. They commonly display poor conservation across species [36], making them challenging to study using conventional biological techniques. Relative to their genomic location, lncRNAs are divided into five subclasses: sense (overlap exon on the same strand); antisense (located complementary to the sense strand); long intergenic (lincRNA) (located between coding genes, but their transcription occurs individually); bidirectional (located between coding genes and transcribed simultaneously); and intronic (produced by an intronic region) [37]. The schematic diagram of lncRNA biogenesis, types, and mechanism of gene regulation is presented in Figure 2.
4. LncRNA Databases
Several databases have been established to collect information regarding ncRNAs [41]. Databases such as LNCipedia [42], lncRNome [43], deepBase [44], and LncBook [45] are specially curated to categorize lncRNAs. However, they are limited to non-insect species, mainly humans and mice. NonCode, a more generalist database, comprises lncRNAs reported (experimental or computational approaches) from several model organisms, including D. melanogaster [46]. BmncRNAdb offers extensive information on the lncRNAs identified in the silkworms by utilizing RNA-seq and unigenes [47]. Similarly, InsectBase 2.0, a recently created database dedicated to insects, has curated several genomes and reported 1,293,430 lncRNAs from 376 insect species using computational tools such as FEELnc (v.0.2) [48,49]. Some of the available databases for lncRNA studies are listed in Table 1.
5. Functional Evidence of lncRNAs in Insects
In the last decade, researchers have identified some lncRNAs from insect species; however, nearly all developments in defining the functions of lncRNAs are focused on plants and mammals. D. melanogaster was the first insect species to have lncRNA identified from RNA-seq data [18]. Subsequently, researchers focused on various other insect species, such as Apis cerana and Apis mellifera [55], Nilaparvata lugens [56], Plutella xylostella [57], Pectinophora gossypiella [58] Tribolium castaneum [59] Anopheles gambiae [60], Bombyx mori [61], Aedes aegypti [62], Sogatella furcifera [63], and Bactrocera dorsalis [64].
LncRNAs are poorly characterized in insects, and most of our understanding comes from RNA-seq analysis. However, recent studies on model insect species, i.e., Drosophila, have identified lncRNAs involved in the development, defense mechanism, and resistance. In the subsequent segments, we will review the progress and mechanisms of lncRNAs in different insect species. The lncRNAs identified and functionally characterized in insect arthropods are presented in Table 2.
5.1. Drosophila melanogaster
D. melanogaster is an established model insect species for studying disciplines from fundamental genetics to deciphering complex genetic mechanisms. LncRNAs have been attributed to a variety of biological roles in D. melanogaster, including immunity [79], nervous system [93], spermatogenesis [94,95], and dosage compensation [66]. We will individually discuss the lncRNAs that have been studied in Drosophila.
5.1.1. LncRNA-RNA-on-X (roX)
Dosage compensation is an epigenetic mechanism by which an organism equalizes the gene expressions in both sexes of a species. Organisms have evolved specialized molecular solutions to perform this task. In Drosophila, a dosage compensation complex (DCC) is formed by the two lncRNAs, roX1 and roX2, with five male-specific lethal (MSL) proteins (MLE, MSL1/2/3, and MFO) and up-regulate genes on the single male X chromosome by facilitating the acetylation of histone at lysine 16 (H4K16ac), subsequently increasing gene transcription to balance the expression of two X chromosomes in females [65,66]. MSL1 binds to other proteins, including MSL3, which comprises of chromobarrel and binds to RNA [96]. MSL1/2 recognizes the binding sites, known as high-affinity sites, over the X chromosome, and MOF triggers the H4K16ac, which spreads this chromatin modification [65]. A schematic diagram of this mechanism is presented in Figure 3. It is worth mentioning that both of these lncRNAs function independently. The absence of roX1 diminishes the expression of X-chromosome genes and the loss of roX2 results in MSL-independent gene expression [97].
Contrary to Xist, which predominantly spreads and functions in cis of X chromosomes in mammalian females, roX1 and roX2 in Drosophila possess a unique feature and act in trans, indicating the functional versatility of lncRNAs in the modulation of a single chromosome in the two different systems [98]. Since sex determination and dosage compensation have co-evolved, scientists have recognized sex-lethal (Sxl) as the gene responsible for determining gender in Drosophila [99]. SxlPe, a dose-sensitive promotor, can distinguish between X chromosomes (1 vs. 2), which are only transcribed in females. Because sex determination and dosage compensation are connected, the lncRNAs may have a potential role in promotor activation and, eventually, gender decision [100].
Figure 3.
Regulatory role of roX1/2 lncRNAs on the X chromosome of male Drosophila. Two lncRNAs and five male-specific lethal (MSL) proteins form a dosage compensation complex (DCC) [66]. The DCC is established and extends to the target site via diffusion. MLE is responsible for RNA/DNA helicase and ATPase activity [101]. MSL1/MSL2 identify the target site and MOF initiates the H4K16ac, spreading this chromatin modification over numerous kilobases [65].
Figure 3.
Regulatory role of roX1/2 lncRNAs on the X chromosome of male Drosophila. Two lncRNAs and five male-specific lethal (MSL) proteins form a dosage compensation complex (DCC) [66]. The DCC is established and extends to the target site via diffusion. MLE is responsible for RNA/DNA helicase and ATPase activity [101]. MSL1/MSL2 identify the target site and MOF initiates the H4K16ac, spreading this chromatin modification over numerous kilobases [65].

5.1.2. LncRNA-Heat Shock RNA Omega (hsrω)
The lncRNA heat shock RNA omega (hsrω) is a stress-induced and developmentally expressed lncRNA. Identified from the single-cell transcriptome atlas of the Drosophila brain, the lncRNA hsrω is involved in cellular aging [68]. The lncRNA hsrω (previously termed 93D) is one of the earliest documented lncRNA genes [69], and its multiple transcripts (nuclear and cytoplasmic) are expressed in approximately all cells during development. Under stressful conditions, including heat shock stress, hsrω is critical for normal development [70]. Various species of Drosophila, i.e., D. melanogaster, D. hydei, and D. pseudoobscura, were compared for the structural analysis of lncRNA hsrω, which showed common primary structural characteristics with one intron and two exons. The gene produces three splice variants, with one cytoplasmic (hsrω-c) and the remaining two confined to the nucleus (hsrω-pre-c and hsrω-n) with non-significant ORF [102,103]. The nucleoplasmic transcript (hsrω-n) accumulates and restores vital regulatory factors (i.e., hnRNPs, RNAP II) to an optimum level (pre-stress) in cells that are recovering from heat stress, failure of which can be fatal [104,105]. Under thermal pressure, the silencing/overexpression of hsrω can be lethal to the embryonic and larval stages [106]. These findings highlight the crucial regulatory function of lncRNA hsrω in Drosophila cellular reprogramming, which is essential for survival during thermal stress.
5.1.3. LncRNA-yar
The sleeping pattern in Drosophila is regulated by a conserved lincRNA, termed yellow-achaete intergenic RNA (yar), and expressed through mid-embryogenesis. Conserved across Drosophila species, yar represents an evolution of over 50 million years. [67,107]. Located in the neural gene cluster, the upstream of yar is the yellow gene (y) which codes a protein essential for cuticular pigmentation and sexual performance in males [108,109,110,111], whereas the downstream is the achaete gene (ac), encoding one of four bHLH (basic helix–loop–helix) transcription factors of theachaete–scute complex (AS-C), crucial for the development of the central and peripheral nervous system [112,113,114,115]. The functional studies revealed that nullisomy of yar did not affect flies morphologically; however, it caused deprived and fragmented nighttime sleep, with yar mutants showing reduced rebound sleep followed by sleep deficiency, indicating a critical role of yar in sleep homeostasis. Since yar is a cytoplasmic lincRNA and no evidence advocates its role in the transcription of neighboring genes, a potential relationship between yar and miRNA was studied. The subsequent investigation identified a total of 33 miRNAs. Seed matching confirmed 19 miRNAs in the exon of yar, suggesting that yar may function as a miRNA sponge [67]. Notably, among the matched miRNA seeds, one corresponds to the miR-310 cluster, and studies revealed that the lack of miRNAs 310–313 disrupts synaptic transmission at the neuromuscular junction [116]. These findings demonstrated the potential function of yar in the transcriptional circuit that affects miRNAs in nervous and, ultimately, sleep homeostasis.
5.1.4. LncRNA-iab-8
In Drosophila, a bithorax complex (BX-C) is in charge of the determination of abdominal sections and posterior thorax via regulation of three homeotic genes, i.e., Ultrabithorax (Ubx), abdominal-A (abd-A), and Abdominal-B (Abd-B) [117]. Several ncRNAs are concurrently expressed from the BX-C to cis-modulate specific proteins in abdominal segments [118,119]. The lncRNA iab-8 (∼92kb) is spliced, polyadenylated, and encoded from the intergenic region flanked by the abd-A and Abd-B genes. It is expressed in nerve cells from the eighth abdominal segment and regulates the expression of abd-A via two mechanisms: (1) by acting as a precursor for miRNA, which results in the production of miR-iab-8 embedded within the intronic region of lncRNA iab-8, and (2) by regulating transcriptional interference (TI) of the abd-A promotor in a cis action [71]. The target sites of miR-iab-8 reside in the homeotic genes and their cofactors (hth and exd) [120,121]. A schematic diagram of this mechanism is presented in Figure 4. The knockdown of lncRNA iab-8 can lead to sterility, mainly due to behavioral phenotype changes. In male flies, the inability to bend the abdomen prevents them from mating, whereas in females, it potentially causes peristaltic wave disorder, preventing the oviduct from facilitating the eggs’ movement [71].
5.1.5. LncRNA-oskar
Scientists have discovered that some protein-coding genes can potentially function as lncRNAs [122]. In humans, several mRNAs, such as insulin receptor substrate 1 (IRS1) and p53, can function as regulatory RNA [123,124]. Similarly, a maternal gene oskar in Drosophila is essential for posterior abdominal segmentation and germline determination [72]. During the initial stages of oogenesis, oskar RNA plays a translation-independent role and acts as a lncRNA. Reduction in oskar levels results in a sterile phenotype chiefly due to oogenesis arrest. However, the expression of oskar 3’UTR is adequate for the recovery of egg-less defects of the RNA null mutant independent of protein [73]. Staufen, an RNA-binding protein, is localized inside oocytes in association with the oskar RNA [125]. In oskar null mutants, the Staufen protein fails to transport from the nurse cells into the oocyte, revealing the interdependency of Staufen and oskar RNA regulated via the interaction of Staufen with oskar 3’UTR [73]. However, this non-coding function may probably be mediated via the sequestration of Bruno, a translation regulator which binds to the Bruno responsive element in 3’UTR [122].
5.1.6. LncRNA-CR46018
The Toll pathway is an integral part of insect immunity that recognizes invading pathogens and induces antimicrobial peptides (AMPs) to neutralize the microorganisms [126]. In mammals, numerous lncRNAs, e.g., Lethe [127], lincRNA-Cox2 [128], THRIL [129], and PACER [130], represent the growing list of lncRNAs involved in regulating gene expression in the innate immune system. However, the implication of lncRNAs in regulating insect immune responses is yet to be studied. In Drosophila, the infection of Micrococcus luteus induced the expression of lncRNA-CR46018, suggesting its potential role in the immune response. RNA-seq analysis of the lncRNA-CR46018 mutant flies showed that the overexpression of CR46018 results in the up-regulation of several immune-related genes, mainly in the Toll and IMD pathways. Additional experiments revealed that lncRNA-CR46018 interacted with transcription factors Dif and Dorsal in the Toll pathway. When infected with M. luteus, the CR46018 overexpressing flies showed promising survival abilities compared to the control [74]. The same group studied the function of another novel lncRNA called lncRNA-CR11538. The results showed that lncRNA-CR11538 acts as a Dif/Dorsal decoy resulting in the down-regulation of AMPs and restoration of Drosophila Toll immunity homeostasis [75]. The above research highlights the regulatory function of lncRNAs in Drosophila Toll immunity.
5.2. Apis mellifera
A. mellifera is among the most widely studied insects owing to its environmental and economic potential. However, the studies concerning the role of lncRNAs are insufficient, and only a few have been investigated functionally. Most of the lncRNAs studied are involved in neuronal functions [83]. Other lncRNAs, such as Lncov1/2 (intronic), have been reported from queen ovaries and are responsible for juvenile hormone (JH) dependent maintenance of ovarioles [131]. Similarly, lncRNAs TCONS_00356023, TCONS_00357367, and TCONS_00159909 have also been identified for their potential roles in behavioral transition [132].
5.2.1. LncRNA-Nb-1
Honey bees are eusocial insect species that represent a highly synchronized working of a society. The adult females are divided into two castes, queen (reproduction) and workers (labor). The honey bees have an age-dependent work division, with young workers responsible for brood care. In contrast, elderly workers are responsible for foraging pollen and nectar [133]. Studies have shown that the nurse-forager transition is mediated by a plethora of carefully coordinated interactions of hormonal levels [134], colony demography [135], gene expressional changes in the brain [136], exocrine gland activity, brain chemistry and structure [137,138]. Studies have been conducted to detect the possible involvement of lncRNAs in the nursing-foraging transition in A. mellifera [132]. LncRNA-Nb-1 (Nurse bee brain-selective gene-1), a 700 nt transcript, appears to function in age-dependent transition by modulating the production and secretion of octopamine and JH. In comparison, the expression of lncRNA-Nb-1 was more significant in the brains of nursing bees relative to foraging and queen bees [81], implying its critical role in the behavioral transition.
5.2.2. LncRNA-Ks-1
In the western honey bee A. mellifera, a 17kb nuclear lncRNA-Ks-1 (Kenyon cell/small-type preferential gene-1) is specifically expressed in the mushroom body (MB) of Kenyon cells in the brain. The Kenyon cells play critical roles in the regulatory sections of the insect brain. The lncRNA-Ks-1 is chiefly expressed in the brain of drones compared to queen bees and is present between the brain’s optic lobes and lateral calyx, signifying its crucial role in drone-related functions [82]. This study identified another transcript termed LncRNA-AncR-1, mainly expressed in the sexual tissues, secretory organs, and the brain [83]. Both these transcripts are primarily present in the neuronal nuclei, suggesting their potential neural functions.
5.2.3. LncRNA-kakusei
In A. mellifera, a nuclear lncRNA, termed kakusei (∼7 kb long), was identified and presented the neuronal activity pattern in foragers [139]. The lncRNA-kakusei comprises various constitutive and inducible variants, and its expression is briefly up-regulated during neuronic activities and is essential to multiple neural functions and RNA metabolism.
Additionally, a homolog of kakusei was also detected and isolated from active neurons of worker bees in A. cerana and called Acks (A. cerana kakusei). It is predominantly expressed in worker bees while forming a hot defensive ball to counter a giant hornet attack [85]. These discoveries highlighted the neuronal function of lncRNAs in honey bees; however, a comprehensive investigation is required to elucidate the underlying mechanisms.
5.3. Bombyx mori
B. mori is among the widely known insects, owing to its economic (silk production) and scientific (model for lepidopteran studies) significance [140]. Over the years, it has continuously generated interest from the scientific community to investigate several known and novel scientific phenomena, including lncRNAs [61,141,142]. We will discuss the progress of lncRNAs in silkworms in the following section.
5.3.1. LncRNA-Bmdsx-AS1
Insects exhibit a series of sex-determination mechanisms, including the composition of sex chromosomes, environmental factors, and pathogen manipulation [143,144]. An antisense lncRNA-Bmdsx-AS1 is abundantly expressed in the B. mori testicle. It functions in the sex-specific alternative splicing of its doublesex (Bmdsx) gene [86]. In insects, the dsx is alternatively spliced into several transcripts, eventually producing sex-specific proteins responsible for sexual dimorphism [145]. The lncRNA-Bmdsx-AS1 binds specifically to the splicing factor hnRNPH and interacts with B. mori P element somatic inhibitor (BmPSI) [86], a male-specific protein responsible for splicing of Bmdsx pre-mRNA [146]. Subsequent studies were performed in transgenic Bmdsx-AS1 to investigate changes in male genitalia. The overexpression of lncRNA-Bmdsx-AS1 altered the morphological structure of male genitalia by increasing the number of claspers compared to the wild-type. Additionally, the overexpression of Bmdsx-AS1 reduced the expression of genes in the EGFR (Epidermal Growth Factor Receptor) signaling pathway and vice versa [87]. EGFR is crucial for developing the eighth abdominal segment in B. mori [147]. Promotor analysis demonstrated that BmAbd-B (gene related to exterior genital development) can negatively modulate the expression of Bmdsx-AS1 [87]. These studies presented a multilayered regulatory network involving BmAbd-B, Bmdsx, and Bmdsx-AS1.
5.3.2. LncRNA-Fben-1
The silkworm B. mori displays sexually dimorphic behavior [148]. The female releases pheromones by extending her pheromones glands to attract mates, whereas the males exhibit typical sexual behavior in response to sex pheromones [149]. Molecular studies of this sexually dimorphic behavior suggested the potential role of a nuclear lncRNA, the lncRNA-Fben-1 (female-brain expressed non-coding RNA-1). The lncRNA-Fben-1 is one of the earliest known lncRNA identified in silkworms. In the B. mori genome, the lncRNA-Fben-1 is located ~6 kb upstream of the fruitless (fru) gene. The expression of lncRNA-Fben-1 is preferential to the cells around the mushroom bodies of the adult female brain, signifying its crucial role in sexually dimorphic neuronal functions in females [88].
5.3.3. LncRNA-209997
The B. mori nucleopolyhedrovirus virus (BmNPV) is an important pathogen. The pathogen severely infects silkworms and seriously damages the sericulture industry [150]. Transcriptomic investigations revealed the role of lncRNAs in B. mori immune responses during baculovirus infection [151,152]. Viral infections can manipulate the expression of lncRNAs, and these lncRNA transcripts can either promote or inhibit viral replication [153,154]. Cellular lncRNA, termed Lnc_209997, was found in silkworm’s virus-infected fat body tissues. The expression of Lnc_209997 was significantly downregulated during infection, thus promoting virus replication. However, the overexpression of Lnc_209997 inhibited viral replication, suggesting its critical role during infection [89]. Researchers have documented that lncRNAs can bind to miRNAs as competitive endogenous RNAs (ceRNAs) and control their target genes, ultimately affecting viral replication [155]. A similar interaction was observed between Lnc_209997 and miR-275-5p during BmNPV infection. Overexpression of Lnc_209997 resulted in the inhibition of miR-275-5p and vice versa. These results suggest that BmNPV can potentially promote its proliferation by suppressing Lnc_209997, which diminished the relationship between Lnc_209997 and miR-275-5p and increased the production of miR-275-5p, therefore promoting its replication [89]. Notably, this hypothesis was not experimentally validated and needs further evidence to decipher the molecular mechanism involved.
5.4. Plutella xylostella
P. xylostella is one of the brassica crop’s most destructive lepidopteran pests [156]. The insect is notorious for resisting multiple insecticides, such as Bacillus thuringiensis (Bt), Fipronil, and Chlorpyrifos. Researchers have identified several lncRNAs for their potential role in developing resistance. Many of these lncRNAs may act as precursors for the production of miRNAs [157]. The glutathione S-transferase (GST) is a crucial detoxification enzyme that contributes to resistance by aiding metabolism or chemical sequestration, resulting in cell excretion [158]. An antisense lncRNA, lnc-GSTu1-AS was identified from chlorantraniliprole-resistant populations of P. xylostella. The lncRNA is transcribed from the opposing strand of GSTu1, a gene associated with chlorantraniliprole resistance. Studies have shown that the knockdown of GSTu1 significantly reduced the P. xylostella resistance to chlorantraniliprole. Furthermore, miR-8525-5p was identified to be regulating the activity of GSTu1. It is worth mentioning that in vivo suppression/overexpression did not change the expression of GSTu1, signifying that an additional layer of gene regulation may be present via lncRNAs. Subsequent experimentations demonstrated that the lnc-GSTu1-AS steadied the GSTu1 expression by blocking the binding site of miR-8525-5p at the 3’UTR of GSTu1, thus preventing its degradation and ultimately increasing resistance to chlorantraniliprole [91]. LncRNAs are expressed differentially under various biotic and abiotic stresses [159]. In response to Metarhizum anisopliae infection, multiple lncRNAs were identified, regulating mRNA expression and acting as miRNA precursors. Bioinformatic analysis predicted the function of these lncRNAs in regulating many immune-related genes, including βGRP, toll, serpin and transferrin [160]. Several differentially expressed transcripts were identified from dsRNA-induced P. xylostella libraries. Dicer-2 and the lncRNA targeting it were significantly expressed post-dsRNA treatment, indicating the crucial roles lncRNAs in the modulation of RNAi pathways [161].
5.5. Aedes aegypti and Aedes albopictus
Ae. albopictus and Ae. aegypti are the primary vectors of chikungunya, dengue fever, yellow fever, and Zika virus [162]. Researchers have investigated the critical modulatory role of host lncRNAs during virus interactions [163]. Recent studies identified and functionally annotated lncRNAs in Ae. aegypti [62,164]. The significance of lncRNAs was investigated in dengue virus serotype 2 (DENV-2) and Wolbachia-infected groups. The results showed that the DENV-2 infection amplified the profusion of lncRNAs in mosquito cells, with some able to suppress virus replication. The silencing of lincRNA_1317 promoted viral replication, suggesting its role in mosquito defense mechanisms against viruses.
Similarly, Wolbachia infection led to several differently expressed lncRNAs [62,165]. Subsequent studies revealed that Wolbachia employs the host lncRNA to maintain intracellular oxidative stress and initiate immune responses. The lncRNA, aae-lnc-7598, was significantly induced during Wolbachia infection and modulated the reactive oxygen species (ROS) via the trans-regulation of an antioxidant gene, CAT1B. Furthermore, Wolbachia reduced the expression levels of another lncRNA, aae-lnc-0165. The suppression of aae-lnc-0165 induced the expression of REL1, a gene crucial for downstream activation of the Toll pathway, by sequence-specific binding of aae-miR-980-5p. Notably, suppressing aae-lnc-0165 also reduced cell ROS levels [92]. Transcriptional analysis performed in chikungunya infected midgut of Ae. albopictus identified several differentially expressed lncRNA transcripts, likely in response to infection [166]. Besides their role in vector–virus interaction, lncRNAs are crucial players during development. RNA-seq data from different development stages in Ae. albopictus revealed that most of the lncRNAs are up-regulated at the start of metamorphosis, whereas the egg and the early larval stage had a greater number of differentially expressed transcripts, signifying their highly spatiotemporal expression patterns [167].
5.6. Other Insects
LncRNAs have become a global research hotspot, generating increased interest from various entomological research groups. Even though most of the work is conducted on the species mentioned above, RNA-seq has enabled us to identify lncRNAs from several other insect species. In the malarial vector A. gambiae, 2949 lncRNAs were identified from different life stages, most being intergenic [168]. S. furcifera, a destructive pest of rice, was profiled for lncRNAs during different developmental stages and identified 1861 lncRNAs. Most of the lncRNAs were up-regulated in embryo, fourth and fifth instars, indicating the potential role of lncRNAs during development [63]. In order to address the problem related to Bt-resistance in bollworm Helicoverpa zea, a comparative study was conducted between Cry1Ac-resistant and Cry1Ac-susceptible strains. Results showed that multiple lncRNAs were differentially expressed in the resistant strains. Further analysis showed a pseudogenic lncRNA similar to cadherin. Additionally, these lncRNAs showed their proximity to protein-coding genes, including ABC transporter and serine protease, possibly crucial to the mechanism of resistance development [169].
Similarly, in pink bollworm, P. gossypiella, the lncRNA PgCad1 was identified. It regulates P. gossypiella cadherin (PgCad1), a gene involved in resistance development. Knockdown of lncRNA PgCad1 suppressed the transcription of PgCad1 and reduced the susceptibility to Cry1Ac, suggesting that this lncRNA positively regulates cadherin’s expression and increases the vulnerability of pink bollworm larvae [58]. Whole-transcriptome RNA-seq, performed in two strains of Aphis gossypii (spirotetramat-resistant and susceptible), a notorious sap-sucking polyphagous pest of crops. Differential expression analysis revealed 874 lncRNAs, with five lncRNAs predicted as acetyl-CoA carboxylase (ACC) targets. RT-qPCR combined with RNAi confirmed the interaction of lncRNAs with the expression of ACC genes. Additionally, two transcription factors, C/EBPzeta and C/EBP, were found to regulate the transcription level of lncRNAs [170]. The study presented valuable information for understanding the function of A. gossypii lncRNAs in the development of resistance. In B. dorsalis, malathion-resistant and susceptible strains were subjected to RNA-seq, resulting in the identification of 6171 lncRNAs with multiple differentially expressed transcripts. RT-qPCR analysis identified lncRNAs associated with malathion resistance, with lnc3774.2 and lnc15010.10 significantly expressed in the cuticle of the malathion-resistant population, signifying their roles in resistance. Suppression of lnc3774.2 indicated that cuticular changes could be critical in malathion resistance [64]. The testis-specific lncRNA, lnc94638, was identified in the melon fly, Zeugodacus cucurbitae, an extremely destructive insect pest of cucurbit crops. Localized in the apical testicle region containing mature spermatozoa, the knockdown of lnc94638 drastically lowered the number of spermatozoa and affected male fertility [171]. Genome-wide studies of different developmental stages in T. castaneum demonstrated that lncRNAs are crucial for metabolism and cell differentiation [59].
6. Research Challenges and Future
Research on lncRNAs has gained momentum in many plant and animal groups, but insects are still poorly understood. Studies conducted among insects are limited to a few model species, mainly in D. melanogaster, and our knowledge of lncRNAs from other insect species is still in its primitive stage. Since lncRNAs interact with several genetic molecules, i.e., protein, DNA, and RNA, and subsequently modulate gene expression at the epigenetic, transcriptional, and post-transcriptional levels, studying gene expression could help us understand their biological phenomena. An in-depth elucidation of the sequence and structural elements related to lncRNA function will permit us to predict and classify lncRNAs into families, as has happened for proteins with similar structural domains.
Technological advances such as RNA-seq and bioinformatic tools enabled us to detect and characterize lncRNAs to a certain extent. However, due to a lack of available functional information, filtering the lncRNAs from protein-coding RNAs is still challenging. Recently we have seen the reclassification of former ncRNAs (e.g., tarsal-less/polished rice and pgc) as coding for small peptides in Drosophila and Tribolium spp. [172]. The development of devoted bioinformatic tools is required to identify multifunctional RNAs. High-resolution in vivo RNA imaging, comprehensive identification of RNA-binding proteins (ChIRP), and capture hybridization analysis of RNA targets (CHART) are some of the modern experimental methods that can be employed to identify the protein, RNA, and DNA binding partners of lncRNAs. However, these experiments must be performed across various species to determine the success ratios. Computational approaches, including the construction of designated databases and the compilation of available annotation resources, are required to collect and integrate information about lncRNAs across various species.
Another direction is understanding how lncRNAs cross-talk with each other and how these peculiar cross-talks are modulated. The spatiotemporal transcriptome can provide a comprehensive resource for understanding the function and interaction of lncRNAs in different tissues and cells at a particular time. Similar studies in Drosophila identified a set of conserved lncRNAs, regulated in a tissue-specific pattern, and revealed a complex spatiotemporal expression of lncRNAs during neurogenesis [173]. Novel sequencing technologies that directly sequence RNA and RNA modifications without the need for reverse transcription could lay the ground for further studies. However, developing advanced genetic model systems is essential to understand lncRNA functions in vivo and in vitro.
7. Conclusions
Compiling evidence suggests the critical function of lncRNAs in correctly executing gene expression. The examples reviewed and discussed here demonstrate the functional versatility of lncRNAs in various species of insects. Despite the number of proposed models, only a fraction lncRNAs have been ascribed to a specific target and named. For most insect lncRNAs, information comes only from RNA-seq experiments. As mentioned above, technological advances have enabled researchers to obtain results for thousands of lncRNA transcripts in a single investigation. The focus of biological research has shifted towards a holistic approach with the integration of systematic biology, which gives us a comprehensive interpretation of the lncRNA data via computational modeling. However, there is a clear need to develop experimental techniques that would allow us to differentiate lncRNAs from mRNAs and to understand the interaction of lncRNAs with other genetic molecules. Hopefully, the current flurry of attention on lncRNAs will be critical for resolving these complications from widespread experimental discoveries concerning the structural and functional mechanism of lncRNAs in insects.
Author Contributions
J.Z. and F.J. conceived the project. J.Z. and J.H. wrote the manuscript and drew the pictures. X.X. and F.J. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the National Natural Science Foundation of China. Grant numbers 31972345 and 32172498.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge the support of the Laboratory of Bio-Pesticide Innovation and Application of Guangdong Province, Guangzhou, China. We also thank anonymous referees for their valuable comments and suggestion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kukurba, K.R.; Montgomery, S.B. RNA Sequencing and Analysis. Cold Spring Harb. Protoc. 2015, 2015, 951–969. [Google Scholar] [CrossRef] [Green Version]
- Brosius, J. Waste not, want not–transcript excess in multicellular eukaryotes. Trends Genet. 2005, 21, 287–288. [Google Scholar] [CrossRef]
- Gomes, A.Q.; Nolasco, S.; Soares, H. Non-coding RNAs: Multi-tasking molecules in the cell. Int. J. Mol. Sci. 2013, 14, 16010–16039. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Koch, L. Screening for lncRNA function. Nat. Rev. Genet. 2017, 18, 70. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulos, I.; Marin, R.; Cardoso-Moreira, M.; Kaessmann, H. Developmental dynamics of lncRNAs across mammalian organs and species. Nature 2019, 571, 510–514. [Google Scholar] [CrossRef]
- Wang, H.-L.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. Adv. Exp. Med. Biol. 2017, 1008, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Bartolomei, M.S.; Zemel, S.; Tilghman, S.M. Parental imprinting of the mouse H19 gene. Nature 1991, 351, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Barlow, D.P.; Stöger, R.; Herrmann, B.G.; Saito, K.; Schweifer, N. The mouse insulin-like growth factor type-2 receptor is imprinted and closely linked to the Tme locus. Nature 1991, 349, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Brockdorff, N.; Ashworth, A.; Kay, G.F.; McCabe, V.M.; Norris, D.P.; Cooper, P.J.; Swift, S.; Rastan, S. The product of the mouse Xist gene is a 15 kb inactive X-specific transcript containing no conserved ORF and located in the nucleus. Cell 1992, 71, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J.; Hendrich, B.D.; Rupert, J.L.; Lafreniere, R.G.; Xing, Y.; Lawrence, J.; Willard, H.F. The human XIST gene: Analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell 1992, 71, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. In Long Non Coding RNA Biology; Rao, M.R.S., Ed.; Springer: Singapore, 2017; pp. 1–46. [Google Scholar]
- Okazaki, Y.; Furuno, M.; Kasukawa, T.; Adachi, J.; Bono, H.; Kondo, S.; Nikaido, I.; Osato, N.; Saito, R.; Suzuki, H.; et al. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature 2002, 420, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertone, P.; Stolc, V.; Royce, T.E.; Rozowsky, J.S.; Urban, A.E.; Zhu, X.; Rinn, J.L.; Tongprasit, W.; Samanta, M.; Weissman, S. Global identification of human transcribed sequences with genome tiling arrays. Science 2004, 306, 2242–2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-H.; Li, J.-H.; Jiang, S.; Zhou, H.; Qu, L.-H. ChIPBase: A database for decoding the transcriptional regulation of long non-coding RNA and microRNA genes from ChIP-Seq data. Nucleic Acids Res. 2013, 41, D177–D187. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Young, R.S.; Marques, A.C.; Tibbit, C.; Haerty, W.; Bassett, A.R.; Liu, J.-L.; Ponting, C.P. Identification and properties of 1,119 candidate lincRNA loci in the Drosophila melanogaster genome. Genome Biol. Evol. 2012, 4, 427–442. [Google Scholar] [CrossRef] [Green Version]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.r.; Hofacker, I.L. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [Green Version]
- Perkel, J.M. Visiting “Noncodarnia”. BioTechniques 2013, 54, 301–304. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Derakhshan, M.; Rashidi Noshabad, F.Z.; Razaviyan, J.; Hadavi, R.; Jafarpour, H.; Jafari, A.; Rajabi, A.; Hamblin, M.R.; Mahabady, M.K.; et al. Non-Coding RNAs and Brain Tumors: Insights Into Their Roles in Apoptosis. Front. Cell Dev. Biol. 2022, 9, 792185. [Google Scholar] [CrossRef]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The Transcriptional Landscape of the Mammalian Genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Fullwood, M.J. Roles, functions, and mechanisms of long non-coding RNAs in cancer. Genom. Proteom. Bioinform. 2016, 14, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Ingolia, N.T.; Lareau, L.F.; Weissman, J.S. Ribosome profiling of mouse embryonic stem cells reveals the complexity and dynamics of mammalian proteomes. Cell 2011, 147, 789–802. [Google Scholar] [CrossRef] [Green Version]
- Montes, M.; Nielsen, M.M.; Maglieri, G.; Jacobsen, A.; Højfeldt, J.; Agrawal-Singh, S.; Hansen, K.; Helin, K.; Van De Werken, H.J.; Pedersen, J.S. The lncRNA MIR31HG regulates p16INK4A expression to modulate senescence. Nat. Commun. 2015, 6, 6967. [Google Scholar] [CrossRef] [Green Version]
- Rashid, F.; Shah, A.; Shan, G. Long Non-coding RNAs in the Cytoplasm. Genom. Proteom. Bioinform. 2016, 14, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA-LncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; McClusky, W.G.; Abdelmohsen, K.; Gorospe, M. Cytoplasmic functions of long noncoding RNAs. Wiley Interdiscip Rev. RNA 2018, 9, e1471. [Google Scholar] [CrossRef]
- Bosson, A.D.; Zamudio, J.R.; Sharp, P.A. Endogenous miRNA and target concentrations determine susceptibility to potential ceRNA competition. Mol. Cell 2014, 56, 347–359. [Google Scholar] [CrossRef]
- Qi, P.; Zhou, X.-y.; Du, X. Circulating long non-coding RNAs in cancer: Current status and future perspectives. Mol. Cancer 2016, 15, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Jandura, A.; Krause, H.M. The New RNA World: Growing Evidence for Long Noncoding RNA Functionality. Trends Genet. 2017, 33, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, K.; Leonard, V.E.; Kv, S.; Lalwani, M.K.; Jalali, S.; Patowary, A.; Joshi, A.; Scaria, V.; Sivasubbu, S. Dynamic expression of long non-coding RNAs (lncRNAs) in adult zebrafish. PloS ONE 2013, 8, e83616. [Google Scholar] [CrossRef] [Green Version]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wei, Z.; Dong, H.; Zhou, J.; Yuan, J.; Ni, B.; Wu, Y.; Han, C.; Tian, Y. Regulation of mRNA stability by RBPs and noncoding RNAs contributing to the pathogenicity of Th17 cells. RNA Biol. 2021, 18, 647–656. [Google Scholar] [CrossRef]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef] [Green Version]
- Dey, B.K.; Pfeifer, K.; Dutta, A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014, 28, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Hoeppner, M.P.; Barquist, L.E.; Gardner, P.P. An introduction to RNA databases. In RNA Sequence, Structure, and Function: Computational and Bioinformatic Methods; Humana Press: Totowa, NJ, USA, 2014; Volume 1097, pp. 107–123. [Google Scholar]
- Volders, P.-J.; Verheggen, K.; Menschaert, G.; Vandepoele, K.; Martens, L.; Vandesompele, J.; Mestdagh, P. An update on LNCipedia: A database for annotated human lncRNA sequences. Nucleic Acids Res. 2014, 43, D174–D180. [Google Scholar] [CrossRef]
- Bhartiya, D.; Pal, K.; Ghosh, S.; Kapoor, S.; Jalali, S.; Panwar, B.; Jain, S.; Sati, S.; Sengupta, S.; Sachidanandan, C. lncRNome: A comprehensive knowledgebase of human long noncoding RNAs. Database 2013, 2013, bat034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.-L.; Li, J.-H.; Wu, J.; Sun, W.-J.; Liu, S.; Wang, Z.-L.; Zhou, H.; Yang, J.-H.; Qu, L.-H. deepBase v2.0: Identification, expression, evolution and function of small RNAs, LncRNAs and circular RNAs from deep-sequencing data. Nucleic Acids Res. 2015, 44, D196–D202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Cao, J.; Liu, L.; Du, Q.; Li, Z.; Zou, D.; Bajic, V.B.; Zhang, Z. LncBook: A curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D128–D134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D.; et al. NONCODEV6: An updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef]
- Zhou, Q.-Z.; Zhang, B.; Yu, Q.-Y.; Zhang, Z. BmncRNAdb: A comprehensive database of non-coding RNAs in the silkworm, Bombyx mori. BMC Bioinform. 2016, 17, 370. [Google Scholar] [CrossRef] [Green Version]
- Wucher, V.; Legeai, F.; Hedan, B.; Rizk, G.; Lagoutte, L.; Leeb, T.; Jagannathan, V.; Cadieu, E.; David, A.; Lohi, H. FEELnc: A tool for long non-coding RNA annotation and its application to the dog transcriptome. Nucleic Acids Res. 2017, 45, e57. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Jing, D.; Tang, S.; Chen, X.; Chen, H.; Duanmu, H.; Cong, Y.; Chen, M.; Ye, X.; Zhou, H.; et al. InsectBase 2.0: A comprehensive gene resource for insects. Nucleic Acids Res. 2021, 50, D1040–D1045. [Google Scholar] [CrossRef]
- Gramates, L.S.; Agapite, J.; Attrill, H.; Calvi, B.R.; Crosby, M.A.; dos Santos, G.; Goodman, J.L.; Goutte-Gattat, D.; Jenkins, V.K.; Kaufman, T.; et al. FlyBase: A guided tour of highlighted features. Genetics 2022, 220, bat034. [Google Scholar] [CrossRef]
- Das, T.; Deb, A.; Parida, S.; Mondal, S.; Khatua, S.; Ghosh, Z. LncRBase V. 2: An updated resource for multispecies lncRNAs and ClinicLSNP hosting genetic variants in lncRNAs for cancer patients. RNA Biol. 2021, 18, 1136–1151. [Google Scholar] [CrossRef]
- The RNAcentral Consortium. RNAcentral: A hub of information for non-coding RNA sequences. Nucleic Acids Res. 2018, 47, D221–D229. [Google Scholar] [CrossRef]
- Gong, J.; Liu, C.; Liu, W.; Xiang, Y.; Diao, L.; Guo, A.-Y.; Han, L. LNCediting: A database for functional effects of RNA editing in lncRNAs. Nucleic Acids Res. 2016, 45, D79–D84. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, G.; Li, J.; Zhang, X.; Huang, S.; Xiang, S.; Hu, X.; Liu, C. CRISPRlnc: A manually curated database of validated sgRNAs for lncRNAs. Nucleic Acids Res. 2018, 47, D63–D68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakodi, M.; Jung, J.W.; Park, D.; Ahn, Y.-J.; Lee, S.-C.; Shin, S.-Y.; Shin, C.; Yang, T.-J.; Kwon, H.W. Genome-wide characterization of long intergenic non-coding RNAs (lincRNAs) provides new insight into viral diseases in honey bees Apis cerana and Apis mellifera. BMC Genom. 2015, 16, 680. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Yuan, Z.; Guo, D.; Hou, B.; Yin, C.; Zhang, W.; Li, F. Genome-wide identification of long noncoding RNA genes and their potential association with fecundity and virulence in rice brown planthopper, Nilaparvata lugens. BMC Genom. 2015, 16, 749. [Google Scholar] [CrossRef] [Green Version]
- Etebari, K.; Furlong, M.J.; Asgari, S. Genome wide discovery of long intergenic non-coding RNAs in Diamondback moth (Plutella xylostella) and their expression in insecticide resistant strains. Sci. Rep. 2015, 5, 14642. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hussain, F.; Unnithan, G.C.; Dong, S.; UlAbdin, Z.; Gu, S.; Mathew, L.G.; Fabrick, J.A.; Ni, X.; Carrière, Y. A long non-coding RNA regulates cadherin transcription and susceptibility to Bt toxin Cry1Ac in pink bollworm, Pectinophora gossypiella. Pest. Biochem. Physiol. 2019, 158, 54–60. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.W.; Lu, Y.Y.; Li, B.; Chen, K.P.; Li, C.J. Genome-wide identification and characterization of long non-coding RNAs in Tribolium castaneum. Insect. Sci. 2021, 28, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, A.M.; Waterhouse, R.M.; Muskavitch, M.A. Long non-coding RNA discovery across the genus anopheles reveals conserved secondary structures within and beyond the Gambiae complex. BMC Genom. 2015, 16, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Cheng, T.; Liu, C.; Liu, D.; Zhang, Q.; Long, R.; Zhao, P.; Xia, Q. Systematic identification and characterization of long non-coding RNAs in the silkworm, Bombyx mori. PloS ONE 2016, 11, e0147147. [Google Scholar] [CrossRef] [Green Version]
- Etebari, K.; Asad, S.; Zhang, G.; Asgari, S. Identification of Aedes aegypti long intergenic non-coding RNAs and their association with Wolbachia and dengue virus infection. PLoS Negl. Trop. Dis. 2016, 10, e0005069. [Google Scholar] [CrossRef]
- Chang, Z.X.; Ajayi, O.E.; Guo, D.Y.; Wu, Q.F. Genome-wide characterization and developmental expression profiling of long non-coding RNAs in Sogatella furcifera. Insect. Sci. 2020, 27, 987–997. [Google Scholar] [CrossRef]
- Meng, L.W.; Yuan, G.R.; Chen, M.L.; Dou, W.; Jing, T.X.; Zheng, L.S.; Peng, M.L.; Bai, W.J.; Wang, J.J. Genome-wide identification of long non-coding RNAs (lncRNAs) associated with malathion resistance in Bactrocera dorsalis. Pest Manag. Sci. 2021, 77, 2292–2301. [Google Scholar] [CrossRef]
- Conrad, T.; Akhtar, A. Dosage compensation in Drosophila melanogaster: Epigenetic fine-tuning of chromosome-wide transcription. Nat. Rev. Genet. 2012, 13, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.; Baker, B.S. The rox1 and rox2 RNAs are essential components of the compensasome, which mediates dosage compensation in Drosophila. Mol. Cell 1999, 4, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Soshnev, A.A.; Ishimoto, H.; McAllister, B.F.; Li, X.; Wehling, M.D.; Kitamoto, T.; Geyer, P.K. A conserved long noncoding RNA affects sleep behavior in Drosophila. Genetics 2011, 189, 455–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davie, K.; Janssens, J.; Koldere, D.; De Waegeneer, M.; Pech, U.; Kreft, Ł.; Aibar, S.; Makhzami, S.; Christiaens, V.; González-Blas, C.B. A single-cell transcriptome atlas of the aging Drosophila brain. Cell 2018, 174, 982–998.e920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhotia, S.C.; Mukherjee, T. Absence of novel translation products in relation to induced activity of the 93D puff in Drosophila melanogaster. Chromosoma 1982, 85, 369–374. [Google Scholar] [CrossRef]
- Lakhotia, S.C. Long non-coding RNAs coordinate cellular responses to stress. Wiley Interdiscip. Rev. RNA 2012, 3, 779–796. [Google Scholar] [CrossRef]
- Gummalla, M.; Maeda, R.K.; Castro Alvarez, J.J.; Gyurkovics, H.; Singari, S.; Edwards, K.A.; Karch, F.; Bender, W. abd-A Regulation by the iab-8 Noncoding RNA. PLoS Genet. 2012, 8, e1002720. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, R.; Ephrussi, A. Germ plasm formation and germ cell determination in Drosophila. Germline Dev. 1994, 182, 282–300. [Google Scholar]
- Jenny, A.; Hachet, O.; Závorszky, P.; Cyrklaff, A.; Weston, M.D.; Johnston, D.S.; Erdélyi, M.; Ephrussi, A. A translation-independent role of oskar RNA in early Drosophila oogenesis. Development 2006, 133, 2827–2833. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ni, J.; Wu, S.; Ma, F.; Jin, P.; Li, S. lncRNA-CR46018 positively regulates the Drosophila Toll immune response by interacting with Dif/Dorsal. Dev. Comp. Immunol. 2021, 124, 104183. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, S.; Wu, S.; Jin, P.; Ma, F. LncRNA-CR11538 Decoys Dif/Dorsal to Reduce Antimicrobial Peptide Products for Restoring Drosophila Toll Immunity Homeostasis. Int. J. Mol. Sci. 2021, 22, 10117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wu, S.; Liu, L.; Li, R.; Jin, P.; Li, S. Drosophila Relish Activating lncRNA-CR33942 Transcription Facilitates Antimicrobial Peptide Expression in Imd Innate Immune Response. Front. Immunol. 2022, 13, 905899. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Y.; Qi, G. LncRNA-IRAR-mediated regulation of insulin receptor transcripts in Drosophila melanogaster during nutritional stress. Insect Mol. Biol. 2022, 31, 261–272. [Google Scholar] [CrossRef]
- Li, M.; Wen, S.; Guo, X.; Bai, B.; Gong, Z.; Liu, X.; Wang, Y.; Zhou, Y.; Chen, X.; Liu, L.; et al. The novel long non-coding RNA CRG regulates Drosophila locomotor behavior. Nucleic Acids Res. 2012, 40, 11714–11727. [Google Scholar] [CrossRef] [Green Version]
- Valanne, S.; Salminen, T.S.; Järvelä-Stölting, M.; Vesala, L.; Rämet, M. Immune-inducible non-coding RNA molecule lincRNA-IBIN connects immunity and metabolism in Drosophila melanogaster. PLoS Pathog. 2019, 15, e1007504. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Dai, H.; Chen, S.; Zhang, L.; Long, M. Highly tissue specific expression of Sphinx supports its male courtship related role in Drosophila melanogaster. PloS ONE 2011, 6, e18853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadano, H.; Yamazaki, Y.; Takeuchi, H.; Kubo, T. Age-and division-of-labour-dependent differential expression of a novel non-coding RNA, Nb-1, in the brain of worker honeybees, Apis mellifera L. Insect Mol. Biol. 2009, 18, 715–726. [Google Scholar] [CrossRef]
- Sawata, M.; Yoshino, D.; Takeuchi, H.; Kamikouchi, A.; Ohashi, K.; Kubo, T. Identification and punctate nuclear localization of a novel noncoding RNA, Ks-1, from the honeybee brain. RNA 2002, 8, 772–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawata, M.; Takeuchi, H.; Kubo, T. Identification and analysis of the minimal promoter activity of a novel noncoding nuclear RNA gene, AncR-1, from the honeybee (Apis mellifera L.). RNA 2004, 10, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Humann, F.C.; Hartfelder, K. Representational Difference Analysis (RDA) reveals differential expression of conserved as well as novel genes during caste-specific development of the honey bee (Apis mellifera L.) ovary. Insect. Biochem. Mol. Biol. 2011, 41, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Kiya, T.; Kunieda, T.; Kubo, T. Inducible-and constitutive-type transcript variants of kakusei, a novel non-coding immediate early gene, in the honeybee brain. Insect Mol. Biol. 2008, 17, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, K.; Zha, X. An antisense lncRNA functions in alternative splicing of Bmdsx in the silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 2019, 516, 639–644. [Google Scholar] [CrossRef]
- Wang, K.-X.; Chen, C.-B.; Wan, Q.-X.; Zha, X.-F. Long Non-Coding RNA Bmdsx-AS1 Effects on Male External Genital Development in Silkworm. Insects 2022, 13, 188. [Google Scholar] [CrossRef]
- Taguchi, S.; Iwami, M.; Kiya, T. Identification and characterization of a novel nuclear noncoding RNA, Fben-1, which is preferentially expressed in the higher brain center of the female silkworm moth, Bombyx mori. Neurosci. Lett. 2011, 496, 176–180. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, S.L.; Yin, H.T.; Zhao, Z.M.; Chen, Z.K.; Shen, M.M.; Zhang, Z.D.; Guo, X.J.; Wu, P. Cellular Lnc_209997 suppresses Bombyx mori nucleopolyhedrovirus replication by targeting miR-275-5p in B. mori. Insect Mol. Biol. 2022, 31, 308–316. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y.; Huang, Q.; Zhao, C.; Li, X.; Kan, Y.; Li, D. Long Noncoding RNA lncR17454 Regulates Metamorphosis of Silkworm Through let-7 miRNA Cluster. J. Insect Sci. 2022, 22, 13. [Google Scholar] [CrossRef]
- Zhu, B.; Li, L.; Wei, R.; Liang, P.; Gao, X. Regulation of GSTu1-mediated insecticide resistance in Plutella xylostella by miRNA and lncRNA. PLoS Genet. 2021, 17, e1009888. [Google Scholar] [CrossRef]
- Mao, W.; Zeng, Q.; She, L.; Yuan, H.; Luo, Y.; Wang, R.; She, Y.; Wang, W.; Wang, C.; Pan, X. Wolbachia Utilizes lncRNAs to Activate the Anti-Dengue Toll Pathway and Balance Reactive Oxygen Species Stress in Aedes aegypti Through a Competitive Endogenous RNA Network. Front. Cell Infect. Microbiol. 2022, 11, 823403. [Google Scholar] [CrossRef]
- Li, K.; Tian, Y.; Yuan, Y.; Fan, X.; Yang, M.; He, Z.; Yang, D. Insights into the Functions of LncRNAs in Drosophila. Int. J. Mol. Sci. 2019, 20, 4646. [Google Scholar] [CrossRef]
- Maeda, R.K.; Sitnik, J.L.; Frei, Y.; Prince, E.; Gligorov, D.; Wolfner, M.F.; Karch, F. The lncRNA male-specific abdominal plays a critical role in Drosophila accessory gland development and male fertility. PLoS Genet. 2018, 14, e1007519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedelek, V.; Bodai, L.; Grézal, G.; Kovács, B.; Boros, I.M.; Laurinyecz, B.; Sinka, R. Analysis of Drosophila melanogaster testis transcriptome. BMC Genom. 2018, 19, 697. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.A.; Vermaak, D.; Bayes, J.J.; Malik, H.S. Species-specific positive selection of the male-specific lethal complex that participates in dosage compensation in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 15412–15417. [Google Scholar] [CrossRef] [Green Version]
- Ilik, I.; Akhtar, A. roX RNAs: Non-coding regulators of the male X chromosome in flies. RNA Biol. 2009, 6, 113–121. [Google Scholar] [CrossRef]
- Ramirez, F.; Lingg, T.; Toscano, S.; Lam, K.C.; Georgiev, P.; Chung, H.-R.; Lajoie, B.R.; de Wit, E.; Zhan, Y.; de Laat, W. High-affinity sites form an interaction network to facilitate spreading of the MSL complex across the X chromosome in Drosophila. Mol. Cell 2015, 60, 146–162. [Google Scholar] [CrossRef] [Green Version]
- Mulvey, B.B.; Olcese, U.; Cabrera, J.R.; Horabin, J.I. An interactive network of long non-coding RNAs facilitates the Drosophila sex determination decision. Biochim. Biophys. Acta 2014, 1839, 773–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morra, R.; Yokoyama, R.; Ling, H.; Lucchesi, J.C. Role of the ATPase/helicase maleless (MLE) in the assembly, targeting, spreading and function of the male-specific lethal (MSL) complex of Drosophila. Epigenetics Chromatin 2011, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Lakhotia, S.C. Expression of hsrω-RNAi transgene prior to heat shock specifically compromises accumulation of heat shock-induced Hsp70 in Drosophila melanogaster. Cell Stress Chaperones 2016, 21, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Garbe, J.; Bendena, W.; Alfano, M.; Pardue, M. A Drosophila heat shock locus with a rapidly diverging sequence but a conserved structure. J. Biol. Chem. 1986, 261, 16889–16894. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Lakhotia, S.C. Dynamics of hnRNPs and omega speckles in normal and heat shocked live cell nuclei of Drosophila melanogaster. Chromosoma 2015, 124, 367–383. [Google Scholar] [CrossRef]
- Prasanth, K.; Rajendra, T.; Lal, A.; Lakhotia, S. Omega speckles-a novel class of nuclear speckles containing hnRNPs associated with noncoding hsr-omega RNA in Drosophila. J. Cell Sci. 2000, 113, 3485–3497. [Google Scholar] [CrossRef] [PubMed]
- Lakhotia, S.C. The non-coding, developmentally active and stress inducible hsrω gene of Drosophila melanogaster integrates post-transcriptional processing of other nuclear transcripts. Nc RNAs Mol. Biol. Mol. Med. 2003, 203–221, 203–221. [Google Scholar]
- Li, M.; Liu, L. Neural functions of long noncoding RNAs in Drosophila. J. Comp. Physiol. 2015, 201, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, M.D.; Radovic, A.; Wittkopp, P.J.; Long, A.D. A gene necessary for normal male courtship, yellow, acts downstream of fruitless in the Drosophila melanogaster larval brain. J. Neurobiol. 2003, 55, 53–72. [Google Scholar] [CrossRef]
- Massey, J.H.; Chung, D.; Siwanowicz, I.; Stern, D.L.; Wittkopp, P.J. The yellow gene influences Drosophila male mating success through sex comb melanization. Elife 2019, 8, e49388. [Google Scholar] [CrossRef]
- Nash, W.G.; Yarkin, R.J. Genetic regulation and pattern formation: A study of the yellow locus in Drosophila melanogaster. Genet. Res. 1974, 24, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.K.; Corces, V. Separate regulatory elements are responsible for the complex pattern of tissue-specific and developmental transcription of the yellow locus in Drosophila melanogaster. Genes Dev. 1987, 1, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Gibert, J.-M.; Simpson, P. Evolution of cis-regulation of the proneural genes. Int. J. Dev. Biol. 2003, 47, 643–651. [Google Scholar]
- Negre, B.; Simpson, P. Evolution of the achaete-scute complex in insects: Convergent duplication of proneural genes. Trends Genet. 2009, 25, 147–152. [Google Scholar] [CrossRef]
- García-Bellido, A.; de Celis, J.F. The complex tale of the achaete–scute complex: A paradigmatic case in the analysis of gene organization and function during development. Genetics 2009, 182, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcellini, S.; Gibert, J.-M.; Simpson, P. achaete, but not scute, is dispensable for the peripheral nervous system of Drosophila. Dev. Biol. 2005, 285, 545–553. [Google Scholar] [CrossRef]
- Tsurudome, K.; Tsang, K.; Liao, E.H.; Ball, R.; Penney, J.; Yang, J.-S.; Elazzouzi, F.; He, T.; Chishti, A.; Lnenicka, G. The Drosophila miR-310 cluster negatively regulates synaptic strength at the neuromuscular junction. Neuron 2010, 68, 879–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, R.K.; Karch, F. The ABC of the BX-C: The bithorax complex explained. Development 2006, 133, 1413–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipshitz, H.; Peattie, D.; Hogness, D. Novel transcripts from the Ultrabithorax domain of the bithorax complex. Genes Dev. 1987, 1, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Pease, B.; Borges, A.C.; Bender, W. Noncoding RNAs of the Ultrabithorax domain of the Drosophila bithorax complex. Genetics 2013, 195, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Bender, W. MicroRNAs in the Drosophila bithorax complex. Genes Dev. 2008, 22, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garaulet, D.L.; Castellanos, M.C.; Bejarano, F.; Sanfilippo, P.; Tyler, D.M.; Allan, D.W.; Sánchez-Herrero, E.; Lai, E.C. Homeotic function of Drosophila Bithorax-complex miRNAs mediates fertility by restricting multiple Hox genes and TALE cofactors in the CNS. Dev. Cell 2014, 29, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Sampath, K.; Ephrussi, A. CncRNAs: RNAs with both coding and non-coding roles in development. Development 2016, 143, 1234–1241. [Google Scholar] [CrossRef] [Green Version]
- Gajjar, M.; Candeias, M.M.; Malbert-Colas, L.; Mazars, A.; Fujita, J.; Olivares-Illana, V.; Fåhraeus, R. The p53 mRNA-Mdm2 interaction controls Mdm2 nuclear trafficking and is required for p53 activation following DNA damage. Cancer cell 2012, 21, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, H.; Yamagishi, N.; Tomida, C.; Yano, C.; Aibara, K.; Kohno, S.; Abe, T.; Ohno, A.; Hirasaka, K.; Okumura, Y. A novel myogenic function residing in the 5′ non-coding region of Insulin receptor substrate-1 (Irs-1) transcript. BMC Cell Biol. 2015, 16, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Johnston, D.; Beuchle, D.; Nüsslein-Volhard, C. staufen, a gene required to localize maternal RNAs in the Drosophila egg. Cell 1991, 66, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Rapicavoli, N.A.; Qu, K.; Zhang, J.; Mikhail, M.; Laberge, R.-M.; Chang, H.Y. A mammalian pseudogene lncRNA at the interface of inflammation and anti-inflammatory therapeutics. Elife 2013, 2, e00762. [Google Scholar] [CrossRef]
- Carpenter, S.; Aiello, D.; Atianand, M.K.; Ricci, E.P.; Gandhi, P.; Hall, L.L.; Byron, M.; Monks, B.; Henry-Bezy, M.; Lawrence, J.B. A long noncoding RNA mediates both activation and repression of immune response genes. Science 2013, 341, 789–792. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Chao, T.-C.; Chang, K.-Y.; Lin, N.; Patil, V.S.; Shimizu, C.; Head, S.R.; Burns, J.C.; Rana, T.M. The long noncoding RNA THRIL regulates TNFα expression through its interaction with hnRNPL. Proc. Natl. Acad. Sci. USA 2014, 111, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, M.; Emerson, B.M. p50-associated COX-2 extragenic RNA (PACER) activates COX-2 gene expression by occluding repressive NF-κB complexes. Elife 2014, 3, e01776. [Google Scholar] [CrossRef]
- Humann, F.C.; Tiberio, G.J.; Hartfelder, K. Sequence and expression characteristics of long noncoding RNAs in honey bee caste development–potential novel regulators for transgressive ovary size. PloS ONE 2013, 8, e78915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Shi, T.; Qi, L.; Su, X.; Wang, D.; Dong, J.; Huang, Z.Y. lncRNA profile of Apis mellifera and its possible role in behavioural transition from nurses to foragers. BMC Genom. 2019, 20, 393. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Elekonich, M.M.; Schulz, D.J.; Bloch, G.; Robinson, G.E. Juvenile hormone levels in honey bee (Apis mellifera L.) foragers: Foraging experience and diurnal variation. J. Insect Physiol. 2001, 47, 1119–1125. [Google Scholar] [CrossRef]
- Huang, Z.-Y.; Robinson, G.E. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996, 39, 147–158. [Google Scholar] [CrossRef]
- Weinstock, G.M.; Robinson, G.E.; Gibbs, R.A.; Worley, K.C.; Evans, J.D.; Maleszka, R.; Robertson, H.M.; Weaver, D.B.; Beye, M.; Bork, P. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar]
- Robinson, G.E. Genomics and integrative analyses of division of labor in honeybee colonies. Am. Nat. 2002, 160, S160–S172. [Google Scholar] [CrossRef]
- Heylen, K.; Gobin, B.; Billen, J.; Hu, T.-T.; Arckens, L.; Huybrechts, R. Amfor expression in the honeybee brain: A trigger mechanism for nurse–forager transition. J. Insect Physiol. 2008, 54, 1400–1403. [Google Scholar] [CrossRef]
- Kiya, T.; Kunieda, T.; Kubo, T. Increased Neural Activity of a Mushroom Body Neuron Subtype in the Brains of Forager Honeybees. PLoS ONE 2007, 2, e371. [Google Scholar] [CrossRef] [Green Version]
- The International Silkworm Genome, C. The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect. Biochem. Mol Biol. 2008, 38, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Wu, M.; Ye, X.; Zhao, S.; Liang, J.; Ye, L.; You, Z.; Zhong, B. Comparative mRNA and LncRNA Analysis of the Molecular Mechanisms Associated With Low Silk Production in Bombyx mori. Front. Genet. 2021, 11, 592128. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Z.; Lin, S.; Wu, W.; Tang, W.; Dong, Y.; Shen, M.; Wu, P.; Guo, X. Identification of long noncoding RNAs in silkworm larvae infected with Bombyx mori cypovirus. Arch. Insect Biochem. Physiol. 2021, 106, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, E.C.; van de Zande, L. Insect Sex Determination: A Cascade of Mechanisms. Sex. Dev. 2014, 8, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Blackmon, H.; Ross, L.; Bachtrog, D. Sex Determination, Sex Chromosomes, and Karyotype Evolution in Insects. J. Hered. 2016, 108, 78–93. [Google Scholar] [CrossRef] [Green Version]
- Black, D.L. Mechanisms of Alternative Pre-Messenger RNA Splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.G.; Imanishi, S.; Dohmae, N.; Asanuma, M.; Matsumoto, S. Identification of a male-specific RNA binding protein that regulates sex-specific splicing of Bmdsx by increasing RNA binding activity of BmPSI. Mol. Cell. Biol. 2010, 30, 5776–5786. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Xu, H.; Ma, S.; Guo, H.; Wang, F.; Zhang, L.; Zha, X.; Zhao, P.; Xia, Q. Ectopic expression of the male BmDSX affects formation of the chitin plate in female Bombyx mori. Mol. Reprod. Dev. 2014, 81, 240–247. [Google Scholar] [CrossRef]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T.; Nakagawa, T.; Mitsuno, H.; Mori, H.; Endo, Y.; Tanoue, S.; Yasukochi, Y.; Touhara, K.; Nishioka, T. Identification and functional characterization of a sex pheromone receptor in the silkmoth Bombyx mori. Proc. Natl. Acad. Sci. USA 2004, 101, 16653–16658. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, M. Bombyx mori. In Encyclopedia of Genetics; Brenner, S., Miller, J.H., Eds.; Academic Press: New York, NY, USA, 2001; pp. 231–233. [Google Scholar]
- Satyavathi, V.; Ghosh, R.; Subramanian, S. Long Non-Coding RNAs Regulating Immunity in Insects. Noncoding RNA 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yin, H.; Shen, M.; Huang, H.; Hou, Q.; Zhang, Z.; Zhao, W.; Guo, X.; Wu, P. Analysis of lncRNA-mediated gene regulatory network of Bombyx mori in response to BmNPV infection. J. Invertebr. Pathol. 2020, 170, 107323. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Zhou, R.; Zhao, J.; Zhang, Y.; Yi, D.; Li, Q.; Zhou, J.; Guo, F.; Liang, C. Host long noncoding RNA lncRNA-PAAN regulates the replication of influenza A virus. Viruses 2018, 10, 330. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Liu, X.; Li, J.; Zhou, H.; Carr, M.J.; Zhang, Z.; Shi, W. Long noncoding RNAs: Novel regulators of virus-host interactions. Rev. Med. Virol. 2019, 29, e2046. [Google Scholar] [CrossRef]
- Zhao, X.; Bai, Z.; Li, C.; Sheng, C.; Li, H. Identification of a Novel Eight-lncRNA Prognostic Signature for HBV-HCC and Analysis of Their Functions Based on Coexpression and ceRNA Networks. Biomed Res. Int. 2020, 2020, 8765461. [Google Scholar] [CrossRef] [Green Version]
- Zafar, J.; Zhang, Y.; Huang, J.; Freed, S.; Shoukat, R.F.; Xu, X.; Jin, F. Spatio-Temporal Profiling of Metarhizium anisopliae—Responsive microRNAs Involved in Modulation of Plutella xylostella Immunity and Development. J. Fungi 2021, 7, 942. [Google Scholar] [CrossRef]
- Liu, F.; Guo, D.; Yuan, Z.; Chen, C.; Xiao, H. Genome-wide identification of long non-coding RNA genes and their association with insecticide resistance and metamorphosis in diamondback moth, Plutella xylostella. Sci. Rep. 2017, 7, 15870. [Google Scholar] [CrossRef] [Green Version]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef]
- Valadkhan, S.; Valencia-Hipólito, A. lncRNAs in Stress Response. In Long Non-coding RNAs in Human Disease; Morris, K.V., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 394, pp. 203–236. [Google Scholar]
- Zafar, J.; Huang, J.; Xu, X.; Jin, F. Analysis of Long Non-Coding RNA-Mediated Regulatory Networks of Plutella xylostella in Response to Metarhizium anisopliae Infection. Insects 2022, 13, 916. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Li, H.; Zhang, H.; An, S. Comparative analysis of dsRNA-induced lncRNAs in three kinds of insect species. Arch. Insect Biochem. Physiol. 2020, 103, e21640. [Google Scholar] [CrossRef]
- Shoukat, R.F.; Zafar, J.; Shakeel, M.; Zhang, Y.; Freed, S.; Xu, X.; Jin, F. Assessment of lethal, sublethal, and transgenerational effects of Beauveria bassiana on the demography of Aedes albopictus (Culicidae: Diptera). Insects 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterling, C.; Koch, M.; Koeppel, M.; Garcia-Alcalde, F.; Karlas, A.; Meyer, T.F. Evidence for a crucial role of a host non-coding RNA in influenza A virus replication. RNA Biol. 2014, 11, 66–75. [Google Scholar] [CrossRef]
- Azlan, A.; Obeidat, S.M.; Yunus, M.A.; Azzam, G. Systematic identification and characterization of Aedes aegypti long noncoding RNAs (lncRNAs). Sci. Rep. 2019, 9, 12147. [Google Scholar] [CrossRef] [Green Version]
- Leitner, M.; Etebari, K.; Asgari, S. Transcriptional response of Wolbachia-transinfected Aedes aegypti mosquito cells to dengue virus at early stages of infection. J. Gen. Virol. 2022, 103, 001694. [Google Scholar] [CrossRef] [PubMed]
- Vedururu, R.k.; Neave, M.J.; Tachedjian, M.; Klein, M.J.; Gorry, P.R.; Duchemin, J.-B.; Paradkar, P.N. RNASeq Analysis of Aedes albopictus Mosquito Midguts after Chikungunya Virus Infection. Viruses 2019, 11, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Cheng, P.; Zhang, K.; Gong, M.; Zhang, Z.; Zhang, R. Systematic identification and characterization of long noncoding RNAs (lncRNAs) during Aedes albopictus development. PLOS Negl. Trop. Dis. 2022, 16, e0010245. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, A.M.; Waterhouse, R.M.; Kopin, A.S.; Muskavitch, M.A. Long non-coding RNA discovery in Anopheles gambiae using deep RNA sequencing. BioRxiv 2014. [Google Scholar] [CrossRef] [Green Version]
- Lawrie, R.D.; Mitchell, R.D.; Deguenon, J.M.; Ponnusamy, L.; Reisig, D.; Pozo-Valdivia, A.D.; Kurtz, R.W.; Roe, R.M. Characterization of Long Non-Coding RNAs in the Bollworm, Helicoverpa zea, and Their Possible Role in Cry1Ac-Resistance. Insects 2022, 13, 12. [Google Scholar] [CrossRef]
- Peng, T.; Pan, Y.; Tian, F.; Xu, H.; Yang, F.; Chen, X.; Gao, X.; Li, J.; Wang, H.; Shang, Q. Identification and the potential roles of long non-coding RNAs in regulating acetyl-CoA carboxylase ACC transcription in spirotetramat-resistant Aphis gossypii. Pest. Biochem. Physiol. 2021, 179, 104972. [Google Scholar] [CrossRef]
- Li, W.-J.; Wei, D.; Han, H.-L.; Song, Y.-J.; Wang, Y.; Xu, H.-Q.; Smagghe, G.; Wang, J.-J. lnc94638 is a testis-specific long non-coding RNA involved in spermatozoa formation in Zeugodacus cucurbitae (Coquillett). Insect Mol. Biol. 2021, 30, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M. Everything old is new again: (linc)RNAs make proteins! Embo J. 2014, 33, 937–938. [Google Scholar] [CrossRef] [Green Version]
- McCorkindale, A.L.; Wahle, P.; Werner, S.; Jungreis, I.; Menzel, P.; Shukla, C.J.; Abreu, R.L.P.; Irizarry, R.A.; Meyer, I.M.; Kellis, M. A gene expression atlas of embryonic neurogenesis in Drosophila reveals complex spatiotemporal regulation of lncRNAs. Development 2019, 146, dev175265. [Google Scholar] [CrossRef]
Figure 1.
Some of the significant developments in long non-coding RNA research. Note: lncRNA denotes long non-coding RNA; roX denotes RNA-on-X; XIST denotes X-inactive-specific transcript; Nb-1 denotes Nurse bee brain-selective gene-1; yar denotes yellow-achaete intergenic RNA; and lincRNA denotes long intergenic non-coding RNAs.
Figure 1.
Some of the significant developments in long non-coding RNA research. Note: lncRNA denotes long non-coding RNA; roX denotes RNA-on-X; XIST denotes X-inactive-specific transcript; Nb-1 denotes Nurse bee brain-selective gene-1; yar denotes yellow-achaete intergenic RNA; and lincRNA denotes long intergenic non-coding RNAs.

Figure 2.
Biogenesis, classification, and mechanism of gene regulation by lncRNAs. LncRNA is transcribed by RNA polymerase, which can be categorized into several types: (a) sense; (b) antisense; (c) bidirectional; (d) intronic, and (e) intergenic [37]. The mechanism of gene regulation of lncRNAs is presented: (1) mRNA stability (via recruitment or impairment of RNA degrading enzymes such as exonucleases) [38], (2) mRNA translation (via recruitment or impairment of ribosomes) [39], (3) miRNA sponge (lncRNAs can prevent miRNA by targeting mRNAs by acting as competing endogenous RNAs (ceRNAs) [31], and (4) miRNA precursors (lncRNAs harbor miRNAs by acting as precursors) [40].
Figure 2.
Biogenesis, classification, and mechanism of gene regulation by lncRNAs. LncRNA is transcribed by RNA polymerase, which can be categorized into several types: (a) sense; (b) antisense; (c) bidirectional; (d) intronic, and (e) intergenic [37]. The mechanism of gene regulation of lncRNAs is presented: (1) mRNA stability (via recruitment or impairment of RNA degrading enzymes such as exonucleases) [38], (2) mRNA translation (via recruitment or impairment of ribosomes) [39], (3) miRNA sponge (lncRNAs can prevent miRNA by targeting mRNAs by acting as competing endogenous RNAs (ceRNAs) [31], and (4) miRNA precursors (lncRNAs harbor miRNAs by acting as precursors) [40].

Figure 4.
Schematic diagram of the bithorax complex (BX-C) and the lncRNA-iab-8. The segment-specific domains are presented in rectangles. The lncRNA-iab-8 encodes from the intergenic region between the abd-A and Abd-B genes. It is expressed in neural cells from the eighth abdominal segment and regulates the expression of abd-A. The lncRNA-iab-8 regulates via acting as a precursor for miR-iab-8 production or by controlling transcriptional interference of the abd-A promotor in a cis manner [71] Note: Figure adapted with permission from Daniel L. Garaulet, Eric C. Lai, Mechanisms of Development; published by Elsevier, 2015. Copyright license no. 5462280266230.
Figure 4.
Schematic diagram of the bithorax complex (BX-C) and the lncRNA-iab-8. The segment-specific domains are presented in rectangles. The lncRNA-iab-8 encodes from the intergenic region between the abd-A and Abd-B genes. It is expressed in neural cells from the eighth abdominal segment and regulates the expression of abd-A. The lncRNA-iab-8 regulates via acting as a precursor for miR-iab-8 production or by controlling transcriptional interference of the abd-A promotor in a cis manner [71] Note: Figure adapted with permission from Daniel L. Garaulet, Eric C. Lai, Mechanisms of Development; published by Elsevier, 2015. Copyright license no. 5462280266230.


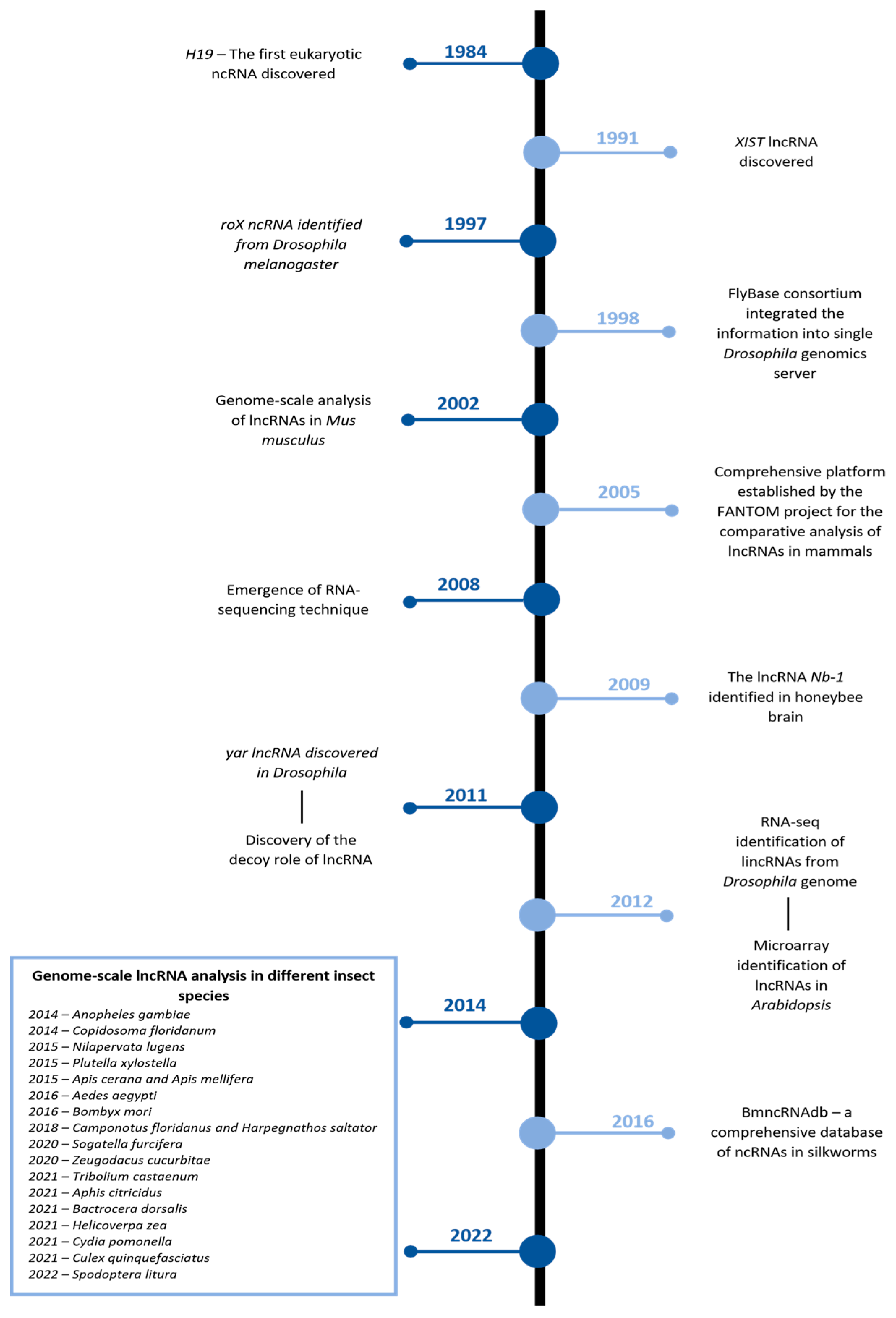{kind=link}
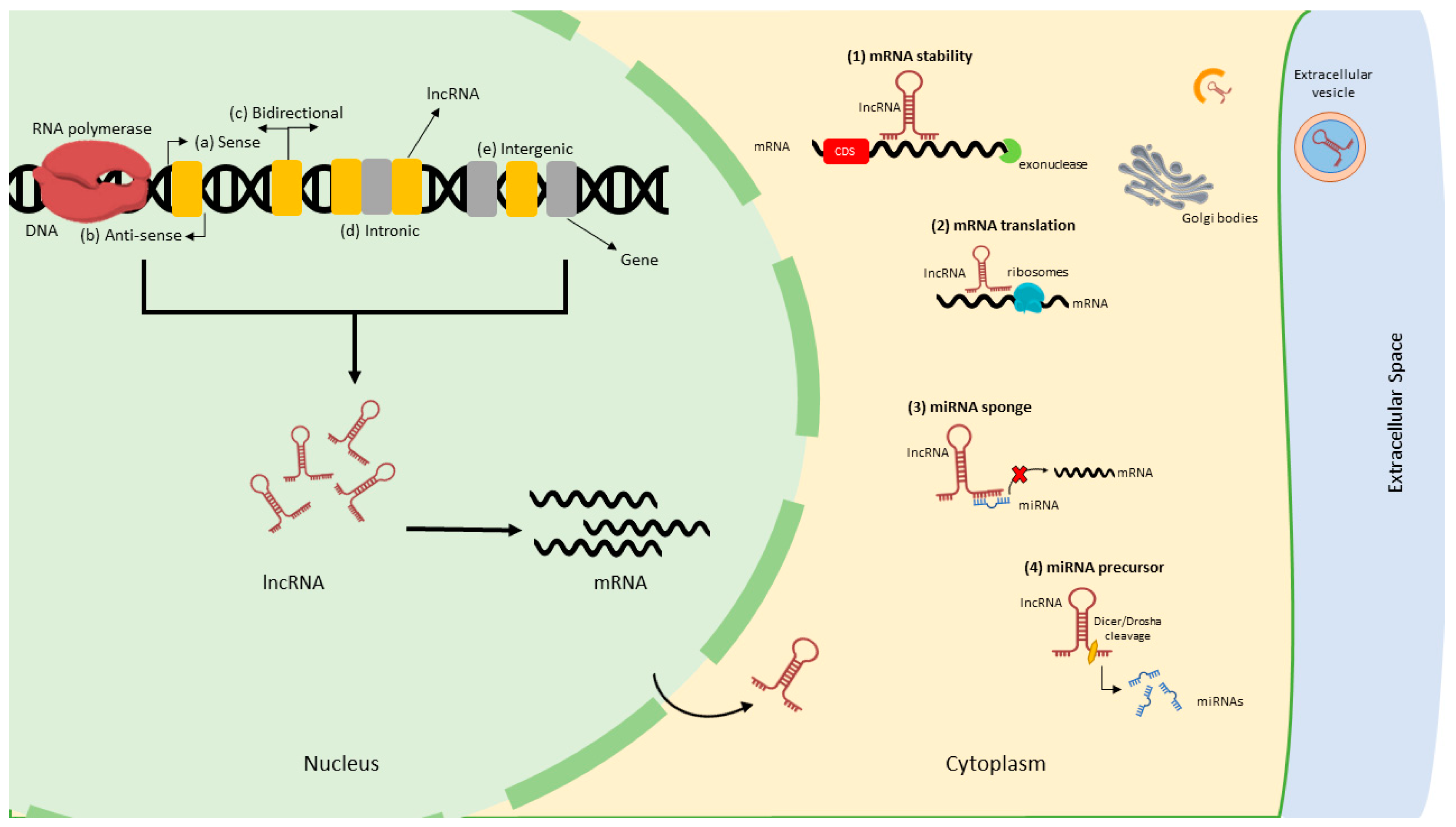{kind=link}
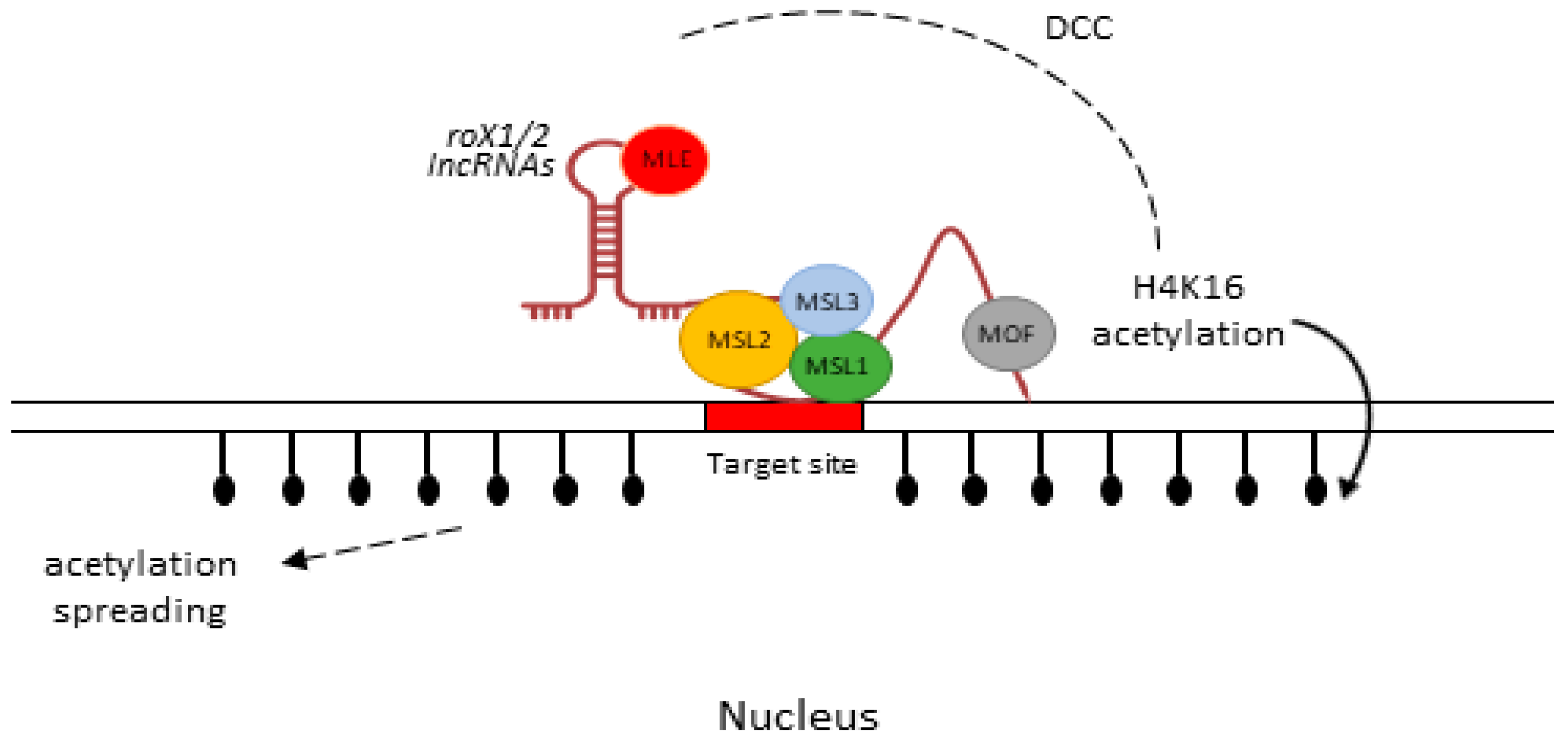{kind=link}
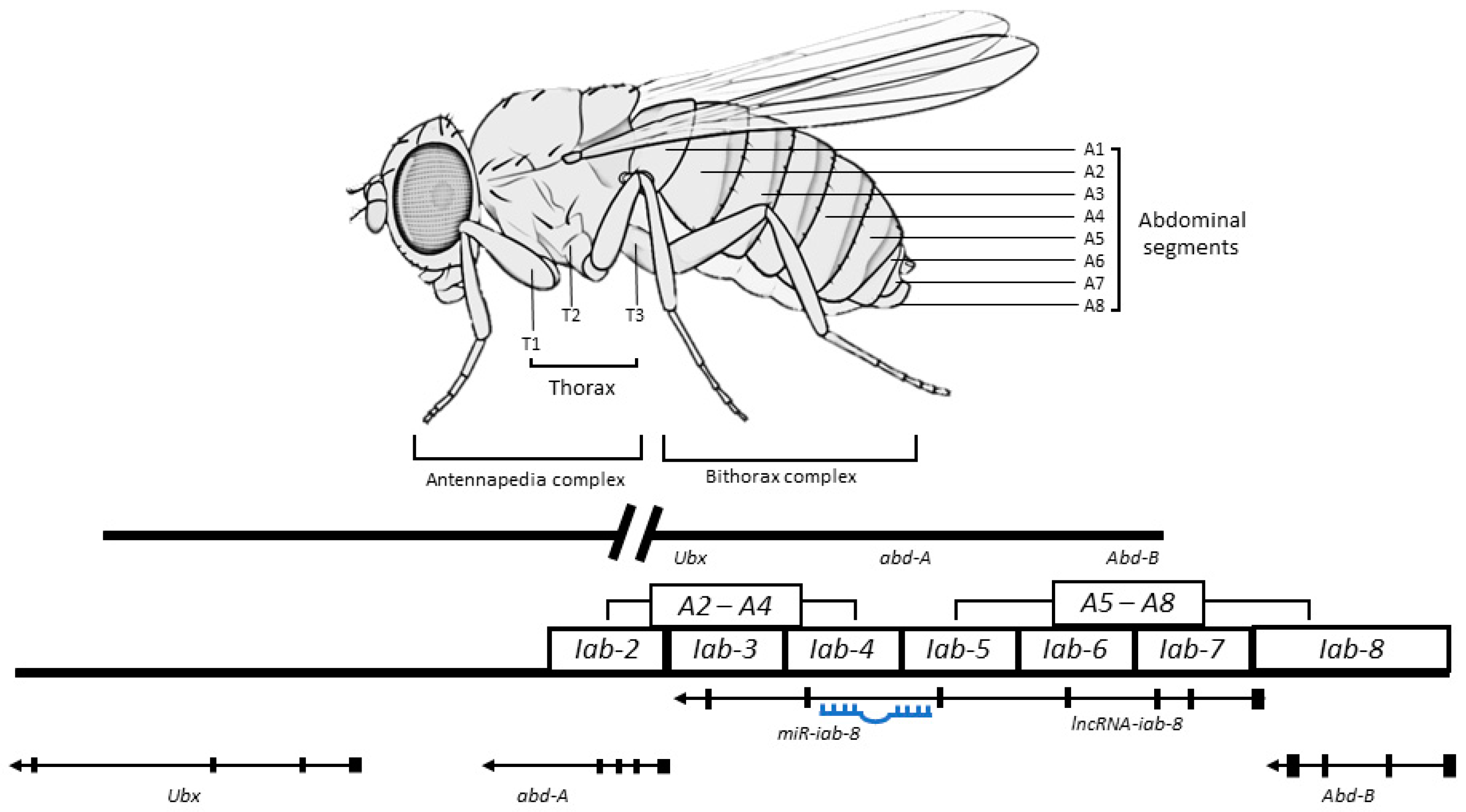{kind=link}
Table 1.
List of some crucial databases available for lncRNA studies.
| Databases | Features | Links | References |
|---|---|---|---|
| InsectBase 2.0 | The database contains more than 16 million sequences, including 817 insect genomes in 20 orders, 158 families, and 457 genera. It includes 1,293,430 lncRNAs from 376 species. | http://v2.insect-genome.com (accessed on 15 November 2022) | [49] |
| BmncRNAdb | The database offers extensive information on the lncRNAs identified in the silkworms. It compiled a list of 6281 lncRNAs with the help of RNA-seq data and unigenes. | http://gene.cqu.edu.cn/BmncRNAdb (accessed on 30 November 2022) | [47] |
| FlyBase | A dedicated repository of Drosophila genes and genomes. | http://flybase.org (accessed on 15 November 2022) | [50] |
| LncRBase V.2 | The database contains information about lncRNAs from eight species, including Drosophila. It provides information on transcript features, genomic location, promoter information, and sORF information within lncRNAs. | http://dibresources.jcbose.ac (accessed on 10 November 2022) | [51] |
| RNAcentral v20 | Multispecies database. RNAcentral imports ncRNA sequences from multiple databases. It has imported data from 47 databases, including Ensemble, LNCipedia, lncRNAdb, and NonCode. | https://rnacentral.org (accessed on 15 November 2022) | [52] |
| NonCode v6.0 | Dedicated database for ncRNAs. Collection of 39 species, including D. melanogaster. It provides basic and advanced information on lncRNAs, such as expression profile, conservation info, and functional prediction. | http://www.noncode.org (accessed on 15 November 2022) | [46] |
| LNCediting | It offers a comprehensive resource for the functional prediction of novel RNA editing sites in lncRNAs amongst multiple species, including Drosophila. | http://bioinfo.life.hust.edu.cn/LNCediting (accessed on 10 November 2022) | [53] |
| CRISPRlnc | Manual of confirmed CRISPR/Cas9 sgRNAs for lncRNA. | http://www.crisprlnc.org/browse (accessed on 8 November 2022) | [54] |
Table 2.
List of lncRNAs identified from different insects.
| LncRNAs | Functions | References |
|---|---|---|
| Drosophila melanogaster | ||
| roX1 and roX2 | Dosage compensation | [65,66] |
| yellow-achaete intergenic RNA (yar) | Sleeping mechanism | [67] |
| heat shock RNA omega (hsrω) | Cellular aging | [68,69,70] |
| iab 8 | Behavioral phenotype | [71] |
| oskar | Abdominal segmentation | [72,73] |
| CR46018 and CR11538 | Toll immunity | [74,75] |
| CR33942 | Imd immune response | [76] |
| IRAR | Insulin signaling | [77] |
| CRG | Locomotive behavior | [78] |
| IBIN | Immunity and metabolism | [79] |
| sphinx | Male courtship behavior | [80] |
| Apis mellifera | ||
| Nb-1 | The behavioral transition from nursing to foraging | [81] |
| Ks-1 and AncR-1 | Neuronal functions | [82,83] |
| Lncov1 and Lncov2 | Cell death of ovarioles in worker embryogenesis. | [84] |
| kakusei | Neural functions in forager bees | [85] |
| Bombyx mori | ||
| Bmdsx-AS1 | Sexual behavior | [86,87] |
| Fben-1 | Role in sexually dimorphic neural functions in females | [88] |
| Lnc_209997 | BmNPV replication | [89] |
| lncR17454 | Metamorphosis | [90] |
| Plutella xylostella | ||
| lnc-GSTu1-AS | Chlorantraniliprole resistance | [91] |
| Aedes aegypti | ||
| lincRNA_1317 | Dengue virus replication | [62] |
| aae-lnc-7598 | Antioxidant functions | [92] |
| aae-lnc-0165 | Toll pathway | [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zafar, J.; Huang, J.; Xu, X.; Jin, F. Recent Advances and Future Potential of Long Non-Coding RNAs in Insects. Int. J. Mol. Sci. 2023, 24, 2605. https://doi.org/10.3390/ijms24032605
AMA Style
Zafar J, Huang J, Xu X, Jin F. Recent Advances and Future Potential of Long Non-Coding RNAs in Insects. International Journal of Molecular Sciences. 2023; 24(3):2605. https://doi.org/10.3390/ijms24032605
Chicago/Turabian StyleZafar, Junaid, Junlin Huang, Xiaoxia Xu, and Fengliang Jin. 2023. "Recent Advances and Future Potential of Long Non-Coding RNAs in Insects" International Journal of Molecular Sciences 24, no. 3: 2605. https://doi.org/10.3390/ijms24032605
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.