
Management of Obesity and Obesity-Related Disorders: From Stem Cells and Epigenetics to Its Treatment
 , , and
, , and 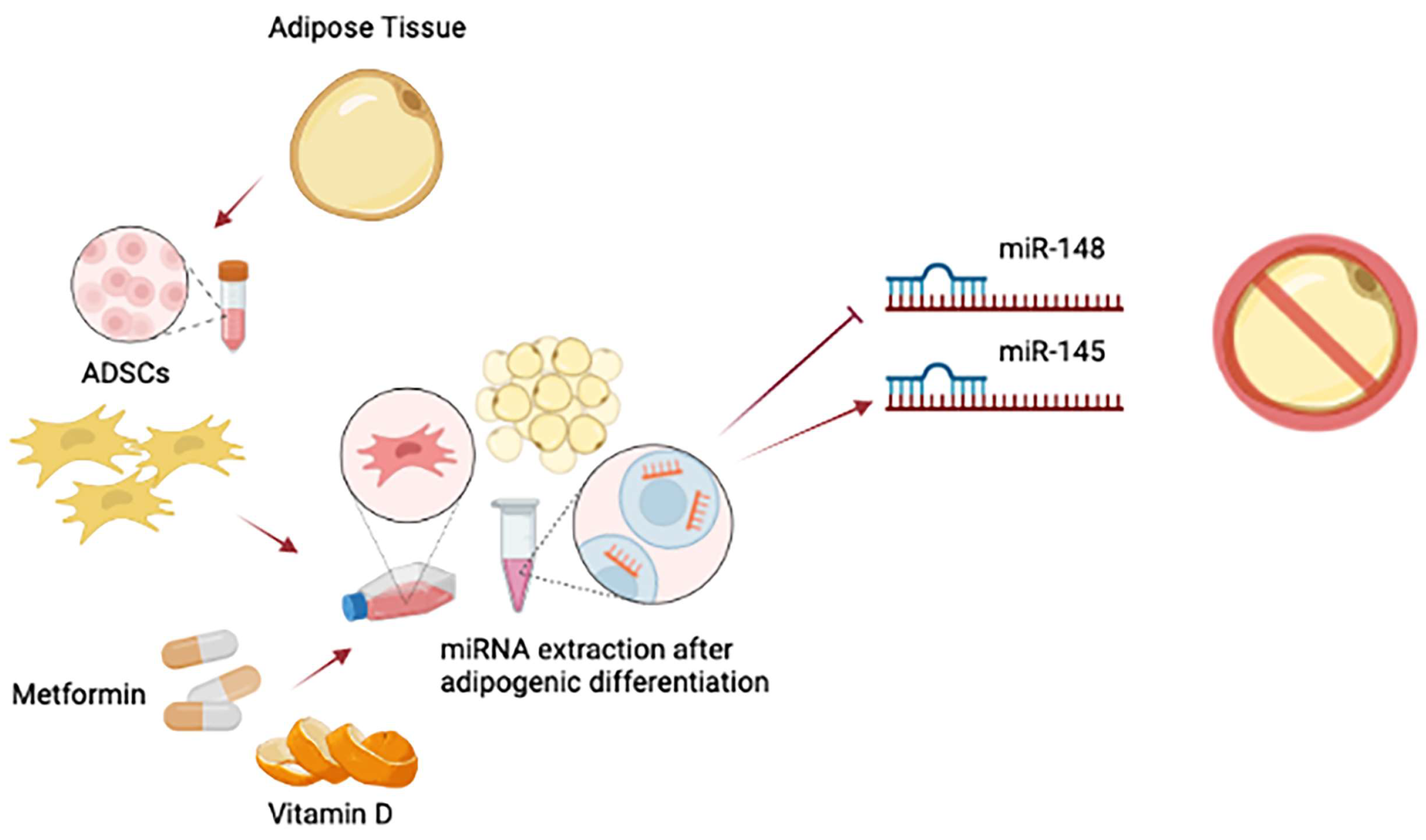{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epigenetic Programming of Adipose-Derived Stem Cells (ADSCs)
3. Biological Features of Adipose Tissue
4. Obesity and Obesity-Related Metabolic Syndrome
5. Surgical Management of Obesity
6. Pharmacological Interventions for Obesity
6.1. Sympathicomimetic
6.2. Sympathicomimetic and Anticonvulsivant
6.3. Pancreatic Lipase Inhibitor
6.4. 5-HT2c Serotonin Agonist
6.5. Glucagon-like Peptide 1 Agonists
6.6. Opioid Receptor Antagonist/Dopamine and Noradrenaline Reuptake Inhibitor
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vicente, A.J.-L.; Beltrán-Carrillo, J.; Megías, Á.; González-Cutre, D. Elements behind sedentary lifestyles and unhealthy eating habits in individuals with severe obesity. Int. J. Qual. Stud. Health Well-Being 2022, 17, 2056967. [Google Scholar]
- Scully, T.; Ettela, A.; LeRoith, D.; Gallagher, E.J. Obesity, Type 2 Diabetes, and Cancer Risk. Front. Oncol. 2021, 10, 615375. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajer, G.R.; Van Haeften, T.W.; Visseren, F.L.J. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Onis, M.; Blössner, M.; Borghi, E. Global prevalence and trends of overweight and obesity among preschool children. Am. J. Clin. Nutr. 2010, 92, 1257–1264. [Google Scholar] [CrossRef] [Green Version]
- Ling, T.-S.A.K.J.; Chen, S.; Zahry, N.R. Economic burden of childhood overweight and obesity: A systematic review and meta-analysis. Obes. Rev. 2022, 24, e13535. [Google Scholar] [CrossRef]
- Lee, Y.H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Welte, M.A.; Gould, A.P. Lipid droplet functions beyond energy storage. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Lazar, M.A. Brown fat in humans: Turning up the heat on obesity. Diabetes 2009, 58, 1482–1484. [Google Scholar] [CrossRef] [Green Version]
- Maliszewska, K.; Kretowski, A. Brown adipose tissue and its role in insulin and glucose homeostasis. Int. J. Mol. Sci. 2021, 22, 1530. [Google Scholar] [CrossRef]
- Pilkington, A.C.; Paz, H.A.; Wankhade, U.D. Beige Adipose Tissue Identification and Marker Specificity—Overview. Front. Endocrinol. 2021, 12, 599134. [Google Scholar] [CrossRef]
- Audano, M.; Pedretti, S.; Caruso, D.; Crestani, M.; De Fabiani, E.; Mitro, N. Regulatory mechanisms of the early phase of white adipocyte differentiation: An overview. Cell. Mol. Life Sci. 2022, 79, 1–14. [Google Scholar] [CrossRef]
- Glenn, J.D. Mesenchymal stem cells: Emerging mechanisms of immunomodulation and therapy. World, J. Stem Cells 2014, 6, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawthorn, W.P.; Scheller, E.L.; MacDougald, O.A. Adipose tissue stem cells meet preadipocyte commitment: Going back to the future. J. Lipid Res. 2012, 53, 227–246. [Google Scholar] [CrossRef] [Green Version]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [Green Version]
- Nic-Can, G.I.; Rodas-Junco, B.A.; Carrillo-Cocom, L.M.; Zepeda-Pedreguera, A.; Peñaloza-Cuevas, R.; Aguilar-Ayala, F.J.; Rojas-Herrera, R.A. Epigenetic regulation of adipogenic differentiation by histone lysine demethylation. Int. J. Mol. Sci. 2019, 20, 3918. [Google Scholar] [CrossRef] [Green Version]
- Ying, H.; Pan, R.; Chen, Y. Epigenetic Control of Mesenchymal Stromal Cell Fate Decision. In Post-Translational Modifications in Cell. Functions and Diseases; InTechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Szymczak-Pajor, I.; Miazek, K.; Selmi, A.; Balcerczyk, A.; Śliwińska, A. The Action of Vitamin D in Adipose Tissue: Is There the Link between Vitamin D Deficiency and Adipose Tissue-Related Metabolic Disorders? Int. J. Mol. Sci. 2022, 23, 956. [Google Scholar] [CrossRef]
- Santaniello, S.; Cruciani, S.; Basoli, V.; Balzano, F.; Bellu, E.; Garroni, G.; Ginesu, G.C.; Cossu, M.L.; Facchin, F.; Delitala, A.P.; et al. Melatonin and Vitamin D Orchestrate Adipose Derived Stem Cell Fate by Modulating Epigenetic Regulatory. Genes 2018, 15, 1631–1639. [Google Scholar] [CrossRef]
- Fathi, E.; Farahzadi, R. Isolation, culturing, characterization and aging of adipose tissue-derived mesenchymal stem cells: A brief overview. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef] [Green Version]
- Schäffler, A.; Büchler, C. Concise Review: Adipose Tissue-Derived Stromal Cells—Basic and Clinical Implications for Novel Cell-Based Therapies. Stem Cells 2007, 25, 818–827. [Google Scholar] [CrossRef]
- Basoli, V.; Santaniello, S.; Cruciani, S.; Ginesu, G.C.; Cossu, M.L.; Delitala, A.P.; Serra, P.A.; Ventura, C.; Maioli, M. Melatonin and vitamin D interfere with the adipogenic fate of adipose-derived stem cells. Int. J. Mol. Sci. 2017, 18, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Maioli, M.; Leonardi, E.; Olivi, E.; Pasquinelli, G.; Valente, S.; Mendez, A.J.; Ricordi, C.; Raffaini, M.; Tremolada, C.; et al. A new nonenzymatic method and device to obtain a fat tissue derivative highly enriched in pericyte-like elements by mild mechanical forces from human lipoaspirates. Cell Transplant. 2013, 22, 2063–2077. [Google Scholar] [CrossRef] [PubMed]
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (ADSC) in plastic and reconstructive surgery. Int. Wound, J. 2017, 14, 112–124. [Google Scholar] [CrossRef]
- Rooj, A.K.; Bronisz, A.; Godlewski, J. The role of octamer binding transcription factors in glioblastoma multiforme. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 18, 1093–1108. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, V.; Rezende, N.C.; Scotland, K.B.; Shaffer, S.M.; Persson, J.L.; Gudas, L.J.; Mongan, A.P. Regulation of Stem cell pluripotency and differentiation involves a mutual regulatory circuit of the Nanog, OCT4, and SOX2 pluripotency transcription factors with polycomb Repressive Complexes and Stem Cell microRNAs. Stem Cells Dev. 2009, 18, 1093–1108. [Google Scholar] [CrossRef]
- Perez-Campo, F.; Riancho, J. Epigenetic Mechanisms Regulating Mesenchymal Stem Cell Differentiation. Curr. Genom. 2015, 16, 368–383. [Google Scholar] [CrossRef] [Green Version]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A complex interplay of multiple molecular determinants and pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Tinahones, F.J. Impaired adipose tissue expandability and lipogenic capacities as ones of the main causes of metabolic disorders. J. Diabetes Res. 2015, 2015, 970375. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.B.; Benkusky, N.A.; Sen, B.; Rubin, J.; Pike, J.W. Epigenetic plasticity drives adipogenic and osteogenic differentiation of marrow-derived mesenchymal stem cells. J. Biol. Chem. 2016, 291, 17829–17847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Yin, C.; Zhao, F.; Ali, A.; Ma, J.; Qian, A. Mesenchymal stem cells: Cell fate decision to osteoblast or adipocyte and application in osteoporosis treatment. Int. J. Mol. Sci. 2018, 19, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Li, N.; Zhang, G. Spontaneous adipogenic differentiation potential of adipose-derived stem cells decreased with increasing cell passages. Mol. Med. Rep. 2018, 17, 6109–6115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Yi, D.; Nguyen, H.P.; Sul, H.S. Epigenetic dynamics of the thermogenic gene program of adipocytes. Biochem. J. 2020, 477, 1137–1148. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Kim, J.; Yuan, X.; Braun, T. Epigenetic modifications of stem cells: A paradigm for the control of cardiac progenitor cells. Circ. Res. 2011, 109, 1067–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, A.I.; Handschin, C. How epigenetic modifications drive the expression and mediate the action of PGC-1α in the regulation of metabolism. Int. J. Mol. Sci. 2019, 20, 5449. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Hwang, J.; Jeong, H.-S. The Role of Histone Acetylation in Mesenchymal Stem Cell Differentiation. Chonnam Med. J. 2022, 58, 6–12. [Google Scholar] [CrossRef]
- Li, H.X.; Xiao, L.; Wang, C.; Gao, J.L.; Zhai, Y.G. Epigenetic regulation of adipocyte differentiation and adipogenesis. J. Zhejiang Univ. Sci. B 2010, 11, 784–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pant, R.; Firmal, P.; Shah, V.K.; Alam, A.; Chattopadhyay, S. Epigenetic Regulation of Adipogenesis in Development of Metabolic Syndrome. Front. Cell Dev. Biol. 2021, 8, 619888. [Google Scholar] [CrossRef] [PubMed]
- Perrini, S.; Porro, S.; Nigro, P.; Cignarelli, A.; Caccioppoli, C.; Genchi, V.A.; Martines, G.; De Fazio, M.; Capuano, P.; Natalicchio, A.; et al. Reduced SIRT1 and SIRT2 expression promotes adipogenesis of human visceral adipose stem cells and associates with accumulation of visceral fat in human obesity. Int. J. Obes. 2020, 44, 307–319. [Google Scholar] [CrossRef]
- Castellano-Castillo, D.; Moreno-Indias, I.; Sanchez-Alcoholado, L.; Ramos-Molina, B.; Alcaide-Torres, J.; Morcillo, S.; Ocaña-Wilhelmi, L.; Tinahones, F.; Queipo-Ortuño, M.I.; Cardonao, F. Altered adipose tissue DNA methylation status in metabolic syndrome: Relationships between global DNA methylation and specific methylation at adipogenic, lipid metabolism and inflammatory candidate genes and metabolic variables. J. Clin. Med. 2019, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Mirzaeicheshmeh, E.; Zerrweck, C.; Centeno-Cruz, F.; Baca-Peynado, P.; Martinez-Hernandez, A.; García-Ortiz, H.; Contreras-Cubas, C.; Salas-Martínez, M.G.; Saldaña-Alvarez, Y.; Mendoza-Caamal, E.C.; et al. Alterations of DNA methylation during adipogenesis differentiation of mesenchymal stem cells isolated from adipose tissue of patients with obesity is associated with type 2 diabetes. Adipocyte 2021, 10, 493–504. [Google Scholar] [CrossRef]
- Mayoral, R.; Osborn, O.; McNelis, J.; Johnson, A.M.; Oh, D.Y.; Izquierdo, C.L.; Chung, H.; Li, P.; Traves, P.G.; Bandyopadhyay, G.; et al. Adipocyte SIRT1 knockout promotes PPARγ activity, adipogenesis and insulin sensitivity in chronic-HFD and obesity. Mol. Metab. 2015, 4, 378–391. [Google Scholar] [CrossRef]
- Eung, J.Y.; Chung, J.J.; Sung, S.C.; Kang, H.K.; Jae, B.K. Down-regulation of histone deacetylases stimulates adipocyte differentiation. J. Biol. Chem. 2006, 281, 6608–6615. [Google Scholar] [CrossRef] [Green Version]
- Cruciani, S.; Garroni, G.; Balzano, F.; Pala, R.; Bellu, E.; Cossu, M.L.; Ginesu, G.C.; Ventura, C.; Maioli, M. Tuning adipogenic differentiation in adscs by metformin and vitamin d: Involvement of mirnas. Int. J. Mol. Sci. 2020, 21, 6181. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Zhang, S.; Mei, G.C.C.; Li, S.; Zhang, W.; Junjvlieke, Z. MiR-145 reduces the activity of PI3K/Akt and MAPK signaling pathways and inhibits adipogenesis in bovine preadipocytes. Genomics 2020, 112, 2688–2694. [Google Scholar] [CrossRef]
- Wang, N.; He, L.; Lin, H.; Tan, L.; Sun, Y.; Zhang, X.; Danser, A.H.J.; Lu, H.S.; He, Y.; Lu, X. MicroRNA-148a regulates low-density lipoprotein metabolism by repressing the (pro)renin receptor. PLoS ONE 2020, 15, e0225356. [Google Scholar] [CrossRef]
- Cruciani, S.; Garroni, G.; Pala, R.; Cossu, M.L.; Ginesu, G.C.; Ventura, C.; Maioli, M. Metformin and vitamin d modulate inflammation and autophagy during adipose-derived stem cell differentiation. Int. J. Mol. Sci. 2021, 22, 6686. [Google Scholar] [CrossRef] [PubMed]
- Cruciani, M.M.S.; Garroni, G.; Pala, R.; Coradduzza, D.; Cossu, M.L.; Ginesu, G.C.; Capobianco, G.; Dessole, S.; Ventura, C. Metformin and vitamin D modulate adipose-derived stem cell differentiation towards the beige phenotype. Adipocyte 2022, 11, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Harvey, I.; Boudreau, A.; Stephens, J.M. Adipose tissue in health and disease: Adipose Tissue in Health and Disease. Open Biol. 2020, 10, 200291. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M.T. Adipose tissue: An endocrine organ playing a role in metabolic regulation. Horm. Mol. Biol. Clin. Investig. 2016, 26, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Bora, P.; Majumdar, A.S. Adipose tissue-derived stromal vascular fraction in regenerative medicine: A brief review on biology and translation. Stem Cell Res. Ther. 2017, 8, 145. [Google Scholar] [CrossRef]
- Ramakrishnan, V.M.; Boyd, N.L. The Adipose Stromal Vascular Fraction as a Complex Cellular Source for Tissue Engineering Applications. Tissue Eng. Part B Rev. 2018, 24, 289–299. [Google Scholar] [CrossRef]
- Song, T.; Kuang, S. Adipocyte dedifferentiation in health and diseases. Clinical Sci. 2019, 133, 2107–2119. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Pontecorvi, P.; Anastasiadou, E.; Napoli, C.; Marchese, C. Immunomodulatory Effect of Adipose-Derived Stem Cells: The Cutting Edge of Clinical Application. Front. Cell Dev. Biol. 2020, 8, 236. [Google Scholar] [CrossRef]
- Abbasi, B.; Shamsasenjan, K.; Ahmadi, M.; Beheshti, S.A.; Saleh, M. Mesenchymal stem cells and natural killer cells interaction mechanisms and potential clinical applications. Stem Cell Res. Ther. 2022, 13, 97. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef]
- Hsiao, W.Y.; Guertin, D.A. De Novo Lipogenesis as a Source of Second Messengers in Adipocytes. Curr. Diabetes Rep. 2019, 19, 138. [Google Scholar] [CrossRef] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giralt, M.; Villarroya, F. White, brown, beige/brite: Different adipose cells for different functions? Endocrinology 2013, 154, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Brown and beige adipose tissues in health and disease. Compr. Physiol. 2017, 7, 1281–1306. [Google Scholar] [CrossRef]
- Al-Mansoori, L.; Al-Jaber, H.; Prince, M.S.; Elrayess, M.A. Role of Inflammatory Cytokines, Growth Factors and Adipokines in Adipogenesis and Insulin Resistance. Inflammation 2022, 45, 31–44. [Google Scholar] [CrossRef]
- Smith, U.; Kahn, B.B. Adipose tissue regulates insulin sensitivity: Role of adipogenesis, de novo lipogenesis and novel lipids. J. Intern. Med. 2016, 280, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, D.E.; Bergman, B.C. Keeping It Local in Metabolic Disease: Adipose Tissue Paracrine Signaling and Insulin Resistance. Diabetes 2022, 71, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, F.; Arroyave, F. Control of adipose cell browning and its therapeutic potential. Metabolites 2020, 10, 471. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, J.; Dai, H.; Duan, Y.; An, Y.; Shi, L.; Lv, Y.; Li, H.; Wang, C.; Ma, Q.; et al. Brown and beige adipose tissue: A novel therapeutic strategy for obesity and type 2 diabetes mellitus. Adipocyte 2021, 10, 48–65. [Google Scholar] [CrossRef]
- Li, Y.; Yun, K.; Mu, R. A review on the biology and properties of adipose tissue macrophages involved in adipose tissue physiological and pathophysiological processes. Lipids Health Dis. 2020, 19, 164. [Google Scholar] [CrossRef]
- Zoico, E.; Rubele, S.; De Caro, A.; Nori, N.; Mazzali, G.; Fantin, F.; Rossi, A.; Zamboni, M. Brown and beige adipose tissue and aging. Front. Endocrinol. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Bank, H.; Kirsh, C.; Simcox, J. Aging adipose: Depot location dictates age-associated expansion and dysfunction. Ageing Res. Rev. 2021, 67, 101259. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F.; Cereijo, R.; Gavaldà-Navarro, A.; Villarroya, J.; Giralt, M. Inflammation of brown/beige adipose tissues in obesity and metabolic disease. J. Intern. Med. 2018, 284, 492–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omran, F.; Christian, M. Inflammatory Signaling and Brown Fat Activity. Front. Endocrinol. 2020, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Purnell, J.Q. Definitions, Classification, and Epidemiology of Obesity; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. [Google Scholar]
- Flegal, K.M.; Graubard, B.I. Estimates of excess deaths associated with body mass index and other anthropometric variables. Am. J. Clin. Nutr. 2009, 89, 1213–1219. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Pathophysiological molecular mechanisms of obesity: A link between mafld and nash with cardiovascular diseases. Int. J. Mol. Sci. 2021, 22, 11629. [Google Scholar] [CrossRef]
- Caussy, C.; Aubin, A.; Loomba, R. The Relationship Between Type 2 Diabetes, NAFLD, and Cardiovascular Risk. Curr. Diabetes Rep. 2021, 21, 15. [Google Scholar] [CrossRef]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. PharmacoEconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Ryan, D.; Barquera, S.; Cavalcanti, O.B.; Ralston, J. The Global Pandemic of Overweight and Obesity. In Handbook of Global Health; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Lagerros, Y.T.; Rössner, S. Obesity management: What brings success? Ther. Adv. Gastroenterol. 2013, 6, 77–88. [Google Scholar] [CrossRef]
- Pigeyre, M.; Yazdi, F.T.; Kaur, Y.; Meyre, D. Recent progress in genetics, epigenetics and metagenomics unveils the pathophysiology of human obesity. Clin. Sci. 2016, 130, 943–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Townshend, T.; Lake, A. Obesogenic environments: Current evidence of the built and food environments. Perspect. Public Health 2017, 137, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-induced changes in adipose tissue microenvironment and their impact on cardiovascular disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [Green Version]
- Baptista, L.S. Obesity and weight loss could alter the properties of adipose stem cells? World, J. Stem Cells 2015, 7, 165–173. [Google Scholar] [CrossRef]
- Payab, M.; Goodarzi, P.; Foroughi Heravani, N.; Hadavandkhani, M.; Zarei, Z.; Falahzadeh, K.; Larijani, B.; Rahim, F.; Arjmand, B. Stem cell and obesity: Current state and future perspective. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Oestreich, A.K.; Collins, K.H.; Harasymowicz, N.S.; Wu, C.L.; Guilak, F. Is Obesity a Disease of Stem Cells? Cell Stem Cell 2020, 27, 15–18. [Google Scholar] [CrossRef]
- Caruso, C.; Balistreri, C.R.; Candore, G. The role of adipose tissue and adipokines in obesity-related inflammatory diseases. Mediat. Inflamm. 2010, 2010, 802078. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-adipogenic Role of AMP-Activated Protein Kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef]
- Benomar, Y.; Taouis, M. Molecular mechanisms underlying obesity-induced hypothalamic inflammation and insulin resistance: Pivotal role of resistin/tlr4 pathways. Front. Endocrinol. 2019, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Campia, U.; Tesauro, M.; Cardillo, C. Human obesity and endothelium-dependent responsiveness. Br. J. Pharmacol. 2012, 165, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Peng, D.Q. New insights into the mechanism of low high-density lipoprotein cholesterol in obesity. Lipids Health Dis. 2011, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Liu, J.L.; Lu, X.; Yang, Q. Epigenetic regulation of energy metabolism in obesity. J. Mol. Cell Biol. 2021, 13, 480–499. [Google Scholar] [CrossRef]
- Park, Y.J.; Han, S.M.; Huh, J.Y.; Kim, J.B. Emerging roles of epigenetic regulation in obesity and metabolic disease. J. Biol. Chem. 2021. [CrossRef] [PubMed]
- Van Dijk, S.J.; Molloy, P.L.; Varinli, H.; Morrison, J.L.; Muhlhausler, B.S. Members of EpiSCOPE. Epigenetics and human obesity. Int. J. Obes. 2015, 39, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Kang, S. Functional implications of DNA methylation in adipose biology. Diabetes 2019, 68, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, E.C.; Garretson, J.T.; Wu, R.; Shi, H.; Xue, B. Neuronal Dnmt1 deficiency attenuates diet-Induced obesity in mice. Endocrinology 2018, 159, 145–162. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wu, R.; Cui, X.; Zha, L.; Yu, L.; Shi, H.; Xue, B. Histone deacetylase 1 (HDAC1) negatively regulates thermogenic program in brown adipocytes via coordinated regulation of histone H3 lysine 27 (H3K27) deacetylation and methylation. J. Biol. Chem. 2016, 291, 4523–4536. [Google Scholar] [CrossRef]
- Paulo, E.; Wu, D.; Hecker, P.; Zhang, Y.; Wang, B. Adipocyte HDAC4 activation leads to beige adipocyte expansion and reduced adiposity. J. Endocrinol. 2018, 239, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Mikłosz, A.; Nikitiuk, B.E.; Chabowski, A. Using adipose-derived mesenchymal stem cells to fight the metabolic complications of obesity: Where do we stand? Obes. Rev. 2022, 23, e13413. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.; Keiran, N.; Núñez-Roa, C.; Maymó-Masip, E.; Ejarque, M.; Sabadell-Basallote, J.; Del Mar Rodríguez-Peña, M.; Ceperuelo-Mallafré, V.; Seco, J.; Benaiges, E.; et al. Effects of stem cells from inducible brown adipose tissue on diet-induced obesity in mice. Sci. Rep. 2021, 11, 13923. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Mazini, L.; Malka, G.; Zeller, M.; Cottin, Y.; Vergely, C. The crosstalk of adipose-derived stem cells (Adsc), oxidative stress, and inflammation in protective and adaptive responses. Int. J. Mol. Sci. 2020, 21, 9262. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, C.; Li, Y.; Tai, R.; Sun, C. Adipose-derived stem cells and obesity: The spear and shield relationship. Genes Dis. 2021; in press. [Google Scholar] [CrossRef]
- Jaber, H.; Issa, K.; Eid, A.; Saleh, F.A. The therapeutic effects of adipose-derived mesenchymal stem cells on obesity and its associated diseases in diet-induced obese mice. Sci. Rep. 2021, 11, 6291. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.M.; Kvach, E.; Eckel, R.H. Treatment of Obesity: Weight Loss and Bariatric Surgery. Circ. Res. 2016, 118, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Hlinnik, A.A.; Aulas, S.D.; Stebounov, S.S.; Rummo, O.O.; Hermanovich, V.I. Bariatric Surgery for Morbid Obesity. Nov. Khirurgii 2021, 29, 662–670. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Varela, J.E. Bariatric surgery for obesity and metabolic disorders: State of the art. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 160–169. [Google Scholar] [CrossRef]
- Eisenberg, D.; Duffy, A.J.; Bell, R.L. Update on obesity surgery. World, J. Gastroenterol. 2006, 12, 3196–3203. [Google Scholar] [CrossRef]
- De Luca, M.; Angrisani, L.; Himpens, J.; Busetto, L.; Scopinaro, N.; Weiner, R.; Sartori, A.; Stier, C.; Lakdawala, M.; Bhasker, A.G.; et al. Indications for Surgery for Obesity and Weight-Related Diseases: Position Statements from the International Federation for the Surgery of Obesity and Metabolic Disorders (IFSO). Obes. Surg. 2016, 26, 1659–1696. [Google Scholar] [CrossRef]
- Cummings, D.E.; Rubino, F. Metabolic surgery for the treatment of type 2 diabetes in obese individuals. Diabetologia 2018. [CrossRef] [PubMed] [Green Version]
- Santarpia, L.; Del Piano, C.; Amato, V.; Marra, M.; De Caprio, C.; De Rosa, E.; Contaldo, F.; Pasanisi, F. Impact of laparoscopic sleeve gastrectomy on metabolism and liver structure. Obes. Facts 2014. [Google Scholar]
- Bužga, M.; Zavadilová, V.; Holéczy, P.; Švagera, Z.; Švorc, P.; Foltys, A.; Zonča, P. Dietary intake and ghrelin and leptin changes after sleeve gastrectomy. Wideochir. Inne Tech. Maloinwazyjne 2014, 9, 554–561. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Zhang, G.; Peng, B.; Cheng, Z.; Du, X. Roux-En-Y Gastric Bypass Versus Sleeve Gastrectomy Plus Procedures for Treatment of Morbid Obesity: Systematic Review and Meta-Analysis. Obes. Surg. 2021, 31, 3303–3311. [Google Scholar] [CrossRef] [PubMed]
- Laurenius, A.M.A.; Wallengren, O.; Alaraj, A.; Forslund, H.B.; Thorell, A.; Wallenius, V. Resolution of diabetes, gastrointestinal symptoms, and self-reported dietary intake after gastric bypass versus sleeve gastrectomy: A randomized study. Surg. Obes. Relat. Dis. 2022; in press. [Google Scholar] [CrossRef]
- Strain, G.W.; Torghabeh, M.H.; Gagner, M.; Ebel, F.; Dakin, G.F.; Abelson, J.S.; Connolly, D.; Pomp, A. The Impact of Biliopancreatic Diversion with Duodenal Switch (BPD/DS) Over 9 Years. Obes. Surg. 2017, 27, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.S.; Guimarães, M.; Almeida, R.; Pereira, A.M.; Lobato, C.B.; Hartmann, B.; Hilsted, L.; Holst, J.J.; Nora, M. Biliopancreatic diversion with duodenal switch (BPD-DS) and single-anastomosis duodeno-ileal bypass with sleeve gastrectomy (SADI-S) result in distinct post-prandial hormone profiles. Int. J. Obes. 2019, 43, 2518–2527. [Google Scholar] [CrossRef] [PubMed]
- Mitzman, B.; Cottam, D.; Goriparthi, R.; Cottam, S.; Zaveri, H.; Surve, A.; Roslin, M.S. Stomach Intestinal Pylorus Sparing (SIPS) Surgery for Morbid Obesity: Retrospective Analyses of Our Preliminary Experience. Obes. Surg. 2016, 26, 2098–2104. [Google Scholar] [CrossRef]
- Neichoy, B.T.; Schniederjan, B.; Cottam, D.R.; Surve, A.K.; Zaveri, H.M.; Cottam, A.; Cottam, S. Stomach intestinal pylorus-sparing surgery for morbid obesity. J. Soc. Laparoendosc. Surg. 2018, 22, e2017.00063. [Google Scholar] [CrossRef] [PubMed]
- Monkhouse, S.J.W.; Morgan, J.D.T.; Norton, S.A. Complication of bariatric surgery: Presentation and emergency management—A review. Ann. R. Coll. Surg. Engl. 2009, 91, 280–286. [Google Scholar] [CrossRef]
- Decker, G.A.; Swain, J.M.; Crowell, M.D.; Scolapio, J.S. Gastrointestinal and nutritional complications after bariatric surgery. Am. J. Gastroenterol. 2007, 102, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Collazo-Clavell, M.L.; Shah, M. Common and Rare Complications of Bariatric Surgery. Endocrinol. Metab. Clin. N. Am. 2020, 49, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Tjeertes, E.E.K.M.; Hoeks, S.S.E.; Beks, S.S.B.J.C.; Valentijn, T.T.M.; Hoofwijk, A.A.G.M.; Stolker, R.J.R.J. Obesity—A risk factor for postoperative complications in general surgery? BMC Anesthesiol. 2015, 15, 112. [Google Scholar] [CrossRef] [Green Version]
- Rashti, F.; Gupta, E.; Ebrahimi, S.; Shope, T.R.; Koch, T.R.; Gostout, C.J. Development of minimally invasive techniques for management of medically-complicated obesity. World J. Gastroenterol. 2014, 20, 13424–13445. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.; Fobi, M.A.L.; Buchwald, J.N. Bariatric Metabolic Surgery Standardization (BMSS) Working Group. Standardization of Bariatric Metabolic Procedures: World Consensus Meeting Statement. Obes. Surg. 2019, 29, 309–345. [Google Scholar] [CrossRef] [Green Version]
- Al-Mulhim, A.S.; Al-Hussaini, H.A.; Al-Jalal, B.A.; Al-Moagal, R.O.; Al-Najjar, S.A. Obesity Disease and Surgery. Int. J. Chronic Dis. 2014, 2014, 652341. [Google Scholar] [CrossRef]
- Král, J.; Machytka, E.; Horká, V.; Selucká, J.; Doleček, F.; Špičák, J.; Kovářová, V.; Haluzík, M.; Bužga, M. Endoscopic treatment of obesity and nutritional aspects of bariatric endoscopy. Nutrients 2021, 13, 4268. [Google Scholar] [CrossRef]
- Picot, J.; Jones, J.; Colquitt, J.L.; Gospodarevskaya, E.; Loveman, E.; Baxter, L.; Clegg, A.J. The clinical effectiveness and cost-effectiveness of bariatric (weight loss) surgery for obesity: A systematic review and economic evaluation. Health Technol. Assess. 2009, 13. [Google Scholar] [CrossRef] [Green Version]
- Zurawel, R.; Gluck, M.; Piecuch, J.; Nowowiejska-Wiewiora, A.; Niedziela, J.; Wiewiora, M. Effectiveness and Safety of Adjustable Gastric Banding in Morbidly Obese Patients After 5 Years of Follow-up. Indian, J. Surg. 2021, 77, 853–862. [Google Scholar] [CrossRef]
- Mandrioli, M.; Inaba, K.; Piccinini, A.; Biscardi, A.; Sartelli, M.; Agresta, F.; Catena, F.; Cirocchi, R.; Jovine, E.; Tugnoli, G.; et al. Advances in laparoscopy for acute care surgery and trauma. World J. Gastroenterol. 2016, 22, 668–680. [Google Scholar] [CrossRef]
- Torgerson, J.S.; Sjöström, L. The Swedish Obese Subjects (SOS) study—Rationale and results. Int. J. Obes. 2001, 25, S2–S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheen, A.J.; Letiexhe, M.; Rorive, M.; De Flines, J.; Luyckx, F.H.; Desaive, C. Bariatric surgery: 10-Year results of Swedish Obese Subjects Study. Rev. Med. 2005, 60, 121–125. [Google Scholar]
- Wiggins, T.; Guidozzi, N.; Welbourn, R.; Ahmed, A.R.; Markar, S.R. Association of bariatric surgery with all-cause mortality and incidence of obesity-related disease at a population level: A systematic review and meta-analysis. PLoS Med. 2020, 17, e1003206. [Google Scholar] [CrossRef] [PubMed]
- Mechanick, J.I.; Apovian, C.; Brethauer, S.; Garvey, W.T.; Joffe, A.M.; Kim, J.; Kushner, R.F.; Lindquist, R.; Pessah-Pollack, R.; Seger, J.; et al. Clinical Practice Guidelines for the Perioperative Nutrition, Metabolic, and Nonsurgical Support of Patients Undergoing Bariatric Procedures—2019 Update: Cosponsored by American Association of Clinical Endocrinologists/American College of Endocrinology. Endocr. Pract. 2019, 25, 1–75. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef]
- Apovian, C.M.; Aronne, L.J.; Bessesen, D.H.; McDonnell, M.E.; Murad, M.H.; Pagotto, U.; Ryan, D.H.; Still, C.D.; Endocrine Society. Pharmacological management of obesity: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2015, 100, 342–362. [Google Scholar] [CrossRef] [Green Version]
- Bray, G.A. Drug treatment of obesity. Psychiatr. Clin. N. Am. 2005, 28, 193–217. [Google Scholar] [CrossRef]
- Van Dieren, S.; Czernichow, S.; Chalmers, J.; Kengne, A.P.; de Galan, B.E.; Poulter, N.; Woodward, M.; Beulens, J.W.; Grobbee, D.E.; van der Schouw, Y.T.; et al. Weight changes and their predictors amongst 11 140 patients with type 2 diabetes in the ADVANCE trial. Diabetes Obes. Metab. 2012, 14, 464–469. [Google Scholar] [CrossRef]
- Turenius, C.I.; Htut, M.M.; Prodon, D.A.; Ebersole, P.L.; Ngo, P.T.; Lara, R.N.; Wilczynski, J.L.; Stanley, B.G. GABAA receptors in the lateral hypothalamus as mediators of satiety and body weight regulation. Brain Res. 2009, 1262, 16–24. [Google Scholar] [CrossRef]
- Garvey, W.T.; Ryan, D.H.; Look, M.; Gadde, K.M.; Allison, D.B.; Peterson, C.A.; Schwiers, M.; Day, W.W.; Bowden, C.H. Two-year sustained weight loss and metabolic benefits with controlled-release phentermine/topiramate in obese and overweight adults (SEQUEL): A randomized, placebo-controlled, phase 3 extension study. Am. J. Clin. Nutr. 2012, 95, 297–308. [Google Scholar] [CrossRef]
- Drent, M.L.; Popp-Snijders, C.; Adér, H.J.; Jansen, J.B.M.J.; van der Veen, E.A. Lipase Inhibition and Hormonal Status, Body Composition and Gastrointestinal Processing of a Liquid High-Fat Mixed Meal in Moderately Obese Subjects. Obes. Res. 1995, 3, 573–581. [Google Scholar] [CrossRef] [PubMed]
- McDuffie, J.R.; Calis, K.A.; Booth, S.L.; Uwaifo, G.I.; Yanovski, J.A. Effects of orlistat on fat-soluble vitamins in obese adolescents. Pharmacotherapy 2002, 22, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Ioannides-Demos, L.L.; Piccenna, L.; McNeil, J.J. Pharmacotherapies for obesity: Past, current, and future therapies. J. Obes. 2011, 2011, 179674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, W.J.; Grottick, A.J.; Menzaghi, F.; Reyes-Saldana, H.; Espitia, S.; Yuskin, D.; Whelan, K.; Martin, M.; Morgan, M.; Chen, W.; et al. Lorcaserin, a novel selective human 5-hydroxytryptamine2C agonist: In vitro and in vivo pharmacological characterization. J. Pharmacol. Exp. Ther. 2008, 325, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.R.; Weissman, N.J.; Anderson, C.M.; Sanchez, M.; Chuang, E.; Stubbe, S.; Bays, H.; Shanahan, W.R. Behavioral Modification and Lorcaserin for Overweight and Obesity Management (BLOOM) Study Group. Multicenter, Placebo-Controlled Trial of Lorcaserin for Weight Management. N. Engl. J. Med. 2010, 363, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Toft-Nielson, M.; Madsbad, S.; Holst, J.J. The Effect of Glucagon-Like Peptide I (GLP-I) on Glucose Elimination in Healthy Subjects Depends on the Pancreatic Glucoregulatory Hormones. Diabetes 1996, 45, 552–556. [Google Scholar] [CrossRef]
- Flint, A.; Raben, A.; Astrup, A.; Holst, J.J. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J. Clin. Investig. 1998, 101, 515–520. [Google Scholar] [CrossRef]
- Schirra, J.; Nicolaus, M.; Roggel, R.; Katschinski, M.; Storr, M.; Woerle, H.J.; Göke, B. Endogenous glucagon-like peptide 1 controls endocrine pancreatic secretion and antro-pyloro-duodenal motility in humans. Gut 2006, 55, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Pi-Sunyer, X.; Astrup, A.; Fujioka, K.; Greenway, F.; Halpern, A.; Krempf, M.; Lau, D.C.; le Roux, C.W.; Violante Ortiz, R.; Jensen, C.B.; et al. A Randomized, Controlled Trial of 3.0 mg of Liraglutide in Weight Management. N. Engl. J. Med. 2015, 373, 11–22. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Pan, A.; Sun, Q.; Czernichow, S.; Kivimaki, M.; Okereke, O.I.; Lucas, M.; Manson, J.E.; Ascherio, A.; Hu, F.B. Bidirectional association between depression and obesity in middle-aged and older women. Int. J. Obes. 2012, 36, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenway, F.L.; Fujioka, K.; Plodkowski, R.A.; Mudaliar, S.; Guttadauria, M.; Erickson, J.; Kim, D.D.; Dunayevich, E.; COR-I Study Group. Effect of naltrexone plus bupropion on weight loss in overweight and obese adults (COR-I): A multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2010, 376, 595–605. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruciani, S.; Delitala, A.P.; Cossu, M.L.; Ventura, C.; Maioli, M. Management of Obesity and Obesity-Related Disorders: From Stem Cells and Epigenetics to Its Treatment. Int. J. Mol. Sci. 2023, 24, 2310. https://doi.org/10.3390/ijms24032310
Cruciani S, Delitala AP, Cossu ML, Ventura C, Maioli M. Management of Obesity and Obesity-Related Disorders: From Stem Cells and Epigenetics to Its Treatment. International Journal of Molecular Sciences. 2023; 24(3):2310. https://doi.org/10.3390/ijms24032310
Chicago/Turabian StyleCruciani, Sara, Alessandro Palmerio Delitala, Maria Laura Cossu, Carlo Ventura, and Margherita Maioli. 2023. "Management of Obesity and Obesity-Related Disorders: From Stem Cells and Epigenetics to Its Treatment" International Journal of Molecular Sciences 24, no. 3: 2310. https://doi.org/10.3390/ijms24032310