
Morphological Methods to Evaluate Peripheral Nerve Fiber Regeneration: A Comprehensive Review



Abstract
:1. Introduction
2. General Concepts
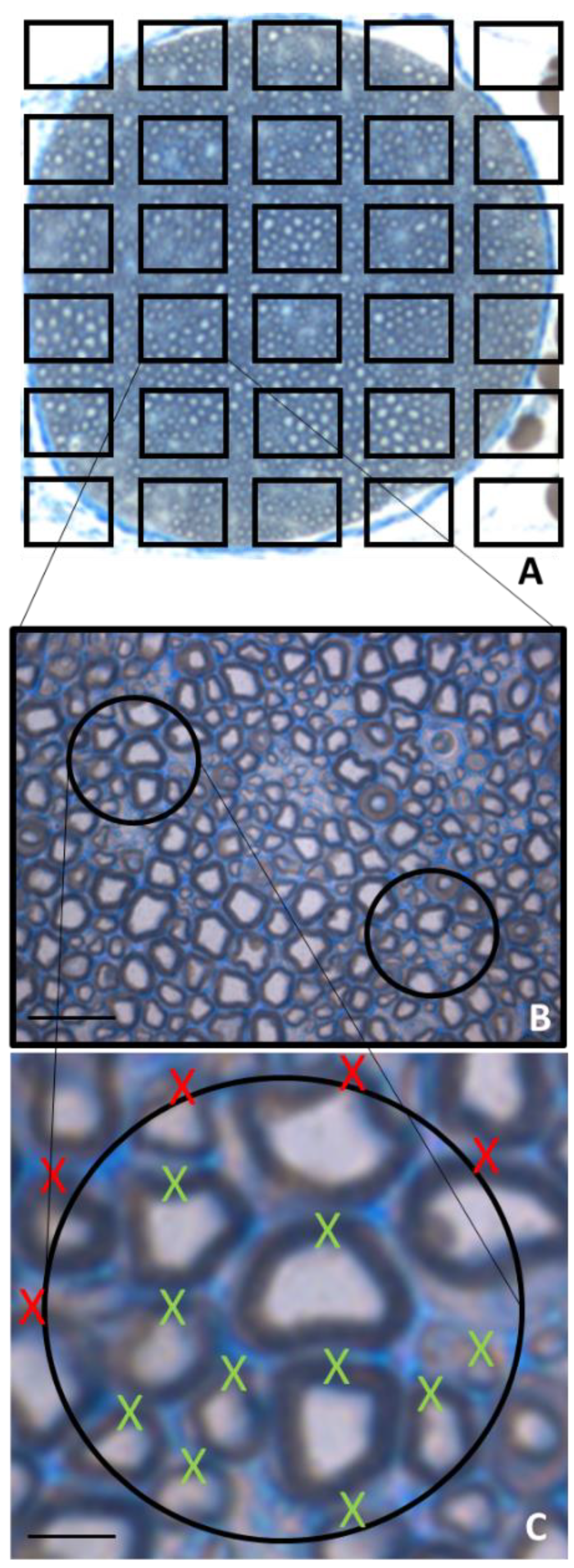
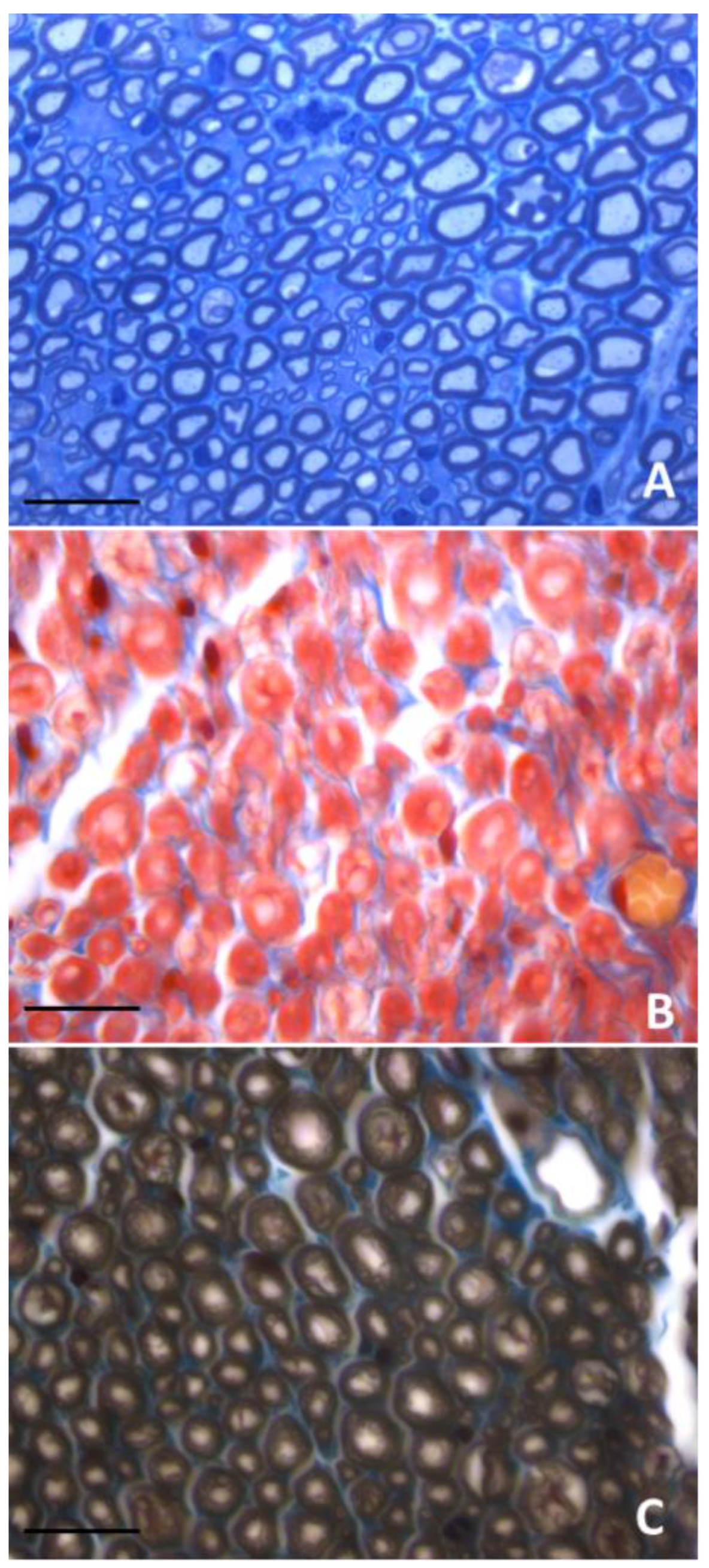
3. Morphological Methods and Myelinated Fiber Quantification
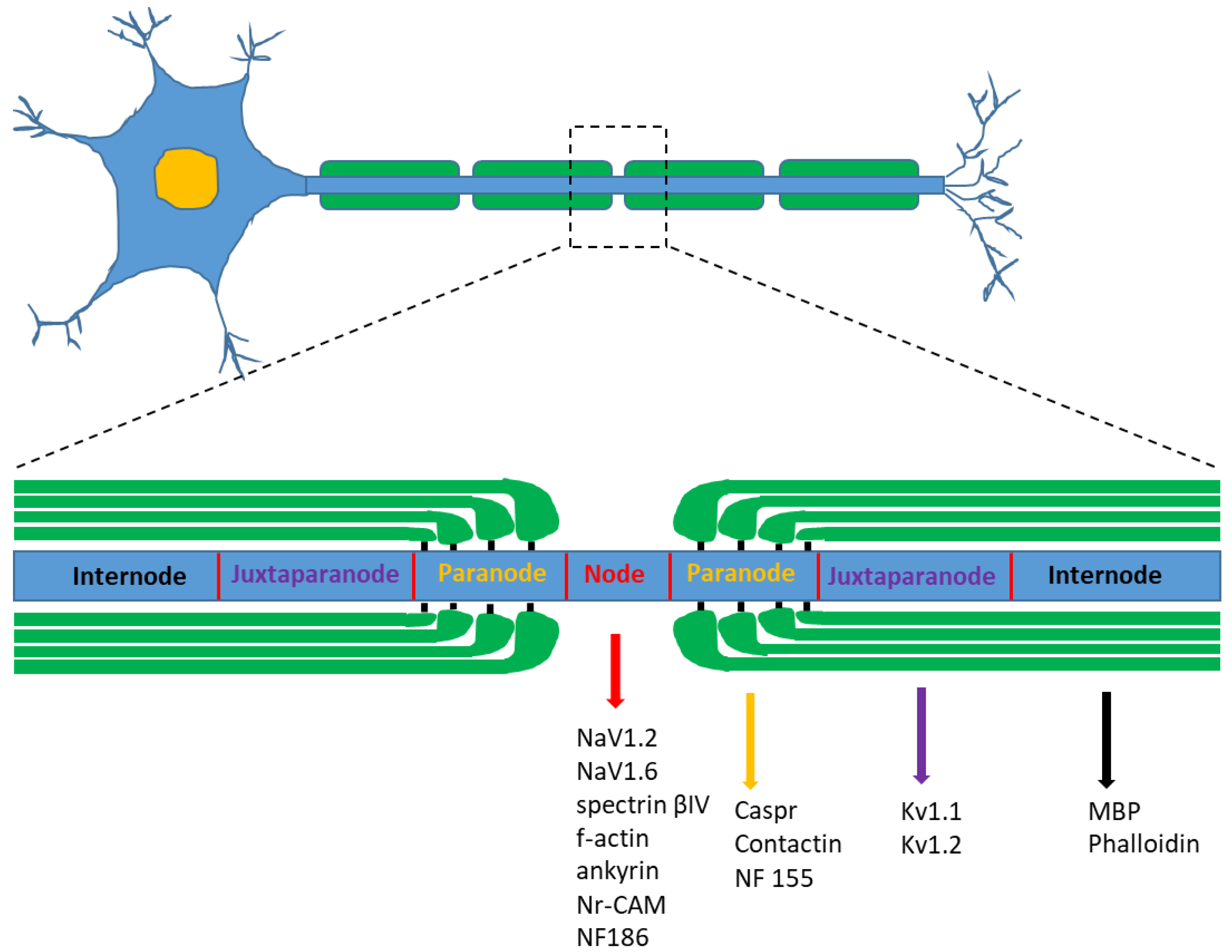
4. Nodal and Internodal Length Analysis
5. Retrograde Labelling Technique for Regenerating Fiber and Neuron Staining
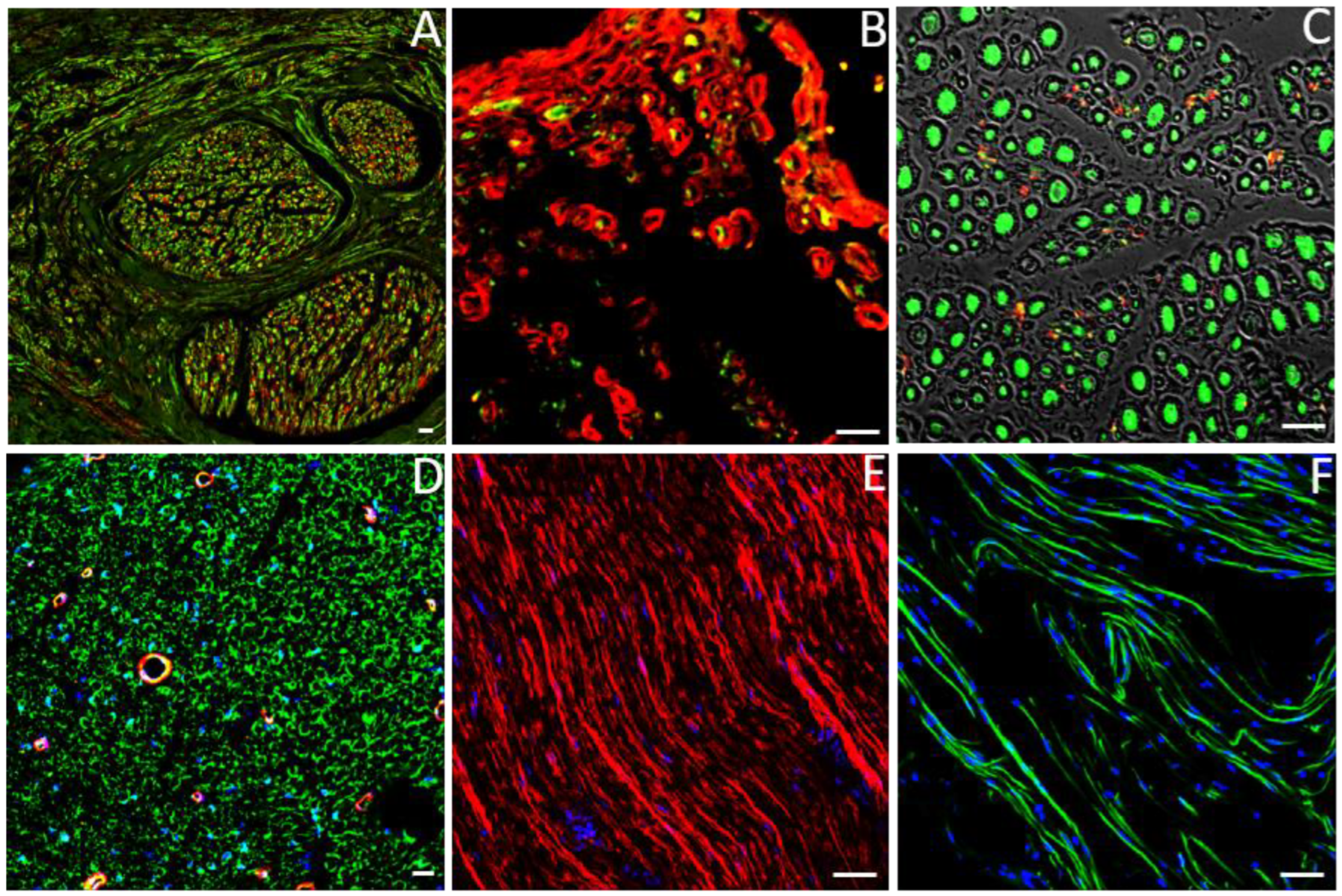
6. Immunohistochemistry for Identifying Proteins Specific for the Different Cellular Components of the Peripheral Nerve
7. Fluorescent Transgenic Animal Models Specific for Nerve Fiber Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transgenic Animal Models | Fluorescent Structures | Aim of the Study | References |
|---|---|---|---|
| B6.Cg-Tg (Thy1-YFP) 16Jrs/J mice | YFP is expressed under the control of Thy1 promoter |
| Mohan and colleagues [77] Tu and colleagues [78] |
| Thy1-YFP-H mice | YFP is expressed under the control of neurospecific regulatory elements of Thy1 gene |
| Nguyen and colleagues [79] |
| Thy1-CFP/S100β-GFP/mice | Expression of cyan fluorescent protein (CFP) in axons under the control of Thy1 promoter and of GFP in Schwann cells under the control of S100β promoter |
| Magill and colleagues [80] |
| Thy1-GFP/rats | GFP is expressed under the control of Thy1 promoter |
| Moore and colleagues [81] Kemp et al. [82] |
| “Kosmos” mice | EGFP is expressed under the control of human S100β regulatory sequences |
| Zuo, Y., et al. [83] |
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daly, W.; Yao, L.; Zeugolis, D.; Windebank, A.; Pandit, A. A biomaterials approach to peripheral nerve regeneration: Bridging the peripheral nerve gap and enhancing functional recovery. J. R. Soc. Interface 2012, 9, 202–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinzel, J.C.; Dadun, L.F.; Prahm, C.; Winter, N.; Bressler, M.; Lauer, H.; Ritter, J.; Daigeler, A.; Kolbenschlag, J. Beyond the Knife-Reviewing the Interplay of Psychosocial Factors and Peripheral Nerve Lesions. J. Pers. Med. 2021, 11, 1200. [Google Scholar] [CrossRef]
- Griffin, J.W.; Hogan, M.V.; Chhabra, A.B.; Deal, D.N. Peripheral nerve repair and reconstruction. J. Bone Joint Surg. Am. 2013, 95, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Hoke, A. A (heat) shock to the system promotes peripheral nerve regeneration. J. Clin. Investig. 2011, 121, 4231–4234. [Google Scholar] [CrossRef]
- Grinsell, D.; Keating, C.P. Peripheral nerve reconstruction after injury: A review of clinical and experimental therapies. Biomed. Res. Int. 2014, 2014, 698256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, X. Functional evaluation of peripheral nerve regeneration and target reinnervation in animal models: A critical overview. Eur. J. Neurosci. 2016, 43, 271–286. [Google Scholar] [CrossRef]
- Varejão, A.S.; Meek, M.F.; Ferreira, A.J.; Patrício, J.A.; Cabrita, A.M. Functional evaluation of peripheral nerve regeneration in the rat: Walking track analysis. J. Neurosci. Methods 2001, 108, 1–9. [Google Scholar] [CrossRef]
- Navarro, X.; Udina, E. Chapter 6: Methods and protocols in peripheral nerve regeneration experimental research: Part III-electrophysiological evaluation. Int. Rev. Neurobiol. 2009, 87, 105–126. [Google Scholar]
- Ronchi, G.; Morano, M.; Fregnan, F.; Pugliese, P.; Crosio, A.; Tos, P.; Geuna, S.; Haastert-Talini, K.; Gambarotta, G. The Median Nerve Injury Model in Pre-clinical Research—A Critical Review on Benefits and Limitations. Front. Cell Neurosci. 2019, 13, 288. [Google Scholar] [CrossRef] [Green Version]
- Geuna, S. The sciatic nerve injury model in pre-clinical research. J. Neurosci. Methods 2015, 243, 39–46. [Google Scholar] [CrossRef]
- Vela, F.J.; Martínez-Chacón, G.; Ballestín, A.; Campos, J.L.; Sánchez-Margallo, F.M.; Abellán, E. Animal models used to study direct peripheral nerve repair: A systematic review. Neural. Regen. Res. 2020, 15, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Fronaro, M.; Di Scipio, F.; Ronchi, G.; Giacobini-Robecchi, M.G.; Geuna, S. Chapter 5: Methods and protocols in peripheral nerve regeneration experimental research: Part II-morphological techniques. Int. Rev. Neurobiol. 2009, 87, 81–103. [Google Scholar]
- Carriel, V.; Garzón, I.; Alaminos, M.; Cornelissen, M. Histological assessment in peripheral nerve tissue engineering. Neural. Regen. Res. 2014, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Carriel, V.; Campos, A.; Alaminos, M.; Raimondo, S.; Geuna, S. Staining Methods for Normal and Regenerative Myelin in the Nervous System. Methods Mol. Biol. 2017, 1560, 207–218. [Google Scholar] [PubMed]
- Muratori, L.; Ronchi, G.; Raimondo, S.; Giacobini-Robecchi, M.G.; Fornaro, M.; Geuna, S. Can regenerated nerve fibers return to normal size? A long-term post-traumatic study of the rat median nerve crush injury model. Microsurgery 2012, 32, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Cercignani, M.; Giulietti, G.; Dowell, N.G.; Gabel, M.; Broad, R.; Leigh, P.N.; Harrison, N.A.; Bozzali, M. Characterizing axonal myelination within the healthy population: A tract-by-tract mapping of effects of age and gender on the fiber g-ratio. Neurobiol. Aging 2017, 49, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Orfahli, L.M.; Rezaei, M.; Figueroa, B.A.; Crawford, A.V.; Annunziata, M.J.; Rao, M.; Fahradyan, V.; Ordenana, C.X.; Silver, J.; Rampazzo, A.; et al. Histomorphometry in Peripheral Nerve Regeneration: Comparison of Different Axon Counting Methods. J. Surg. Res. 2021, 268, 354–362. [Google Scholar] [CrossRef]
- Kemp, S.W.; Cederna, P.; Midha, R. Comparative outcome measures in peripheral regeneration studies. Exp. Neurol. 2017, 287, 348–357. [Google Scholar] [CrossRef]
- Geuna, S.; Gigo-Benato, D.; Ade, C.R. On sampling and sampling errors in histomorphometry of peripheral nerve fibers. Microsurgery 2004, 24, 72–76. [Google Scholar] [CrossRef]
- Wang, W.; Itoh, S.; Konno, K.; Kikkawa, T.; Ichinose, S.; Sakai, K.; Ohkuma, T.; Watabe, K. Effects of Schwann cell alignment along the oriented electrospun chitosan nanofibers on nerve regeneration. J. Biomed. Mater. Res. A 2009, 91, 994–1005. [Google Scholar] [CrossRef]
- Zarei, K.; Scheetz, T.; Christopher, M.; Miller, K.; Hedberg-Buenz, A.; Tandon, A.; Anderson, M.; Fingert, J.; Abramoff, M. Automated Axon Counting in Rodent Optic Nerve Sections with AxonJ. Sci. Rep. 2016, 6, 26559. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.L.; Hricz, N.; Malapati, H.; von Guionneau, N.; Wong, M.; Harris, T.; Boudreau, M.; Cohen-Adad, J.; Tuffaha, S. A simple and robust method for automating analysis of naive and regenerating peripheral nerves. PLoS ONE 2021, 16, e0248323. [Google Scholar] [CrossRef] [PubMed]
- Daeschler, S.C.; Bourget, M.-H.; Derakhshan, D.; Sharma, V.; Asenov, S.I.; Gordon, T.; Cohen-Adad, J.; Borschel, G.H. Rapid, automated nerve histomorphometry through open-source artificial intelligence. Sci. Rep. 2022, 12, 5975. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Z.; Wang, X.; Feng, Z.; Xu, H.; Feng, S.; Wang, K.; Ma, A.; Chen, J.; Wo, Y. Qualitative analysis of axon regeneration by semi-automatic programs on axon photomicrographs proven through concepts of calculus. Int. J. Clin. Exp. Med. 2018, 11, 6560–6570. [Google Scholar]
- Di Scipio, F.; Raimondo, S.; Tos, P.; Geuna, S. A simple protocol for paraffin-embedded myelin sheath staining with osmium tetroxide for light microscope observation. Microsc. Res. Tech. 2008, 71, 497–502. [Google Scholar] [CrossRef]
- Soltanpour, N.; Vostacolaee, Y.A.; Pourghasem, M. Comparison of Morphometric Aspects of Light and Electron Microscopy of the Hypoglossal Nerve between Young and Aged Male Wistar Rats. Cell J. 2012, 13, 229–236. [Google Scholar] [PubMed]
- Ronchi, G.; Jager, S.B.; Vaegter, C.B.; Raimondo, S.; Giacobini-Robecchi, M.G.; Geuna, S. Discrepancies in quantitative assessment of normal and regenerated peripheral nerve fibers between light and electron microscopy. J. Peripher. Nerv. Syst. 2014, 19, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.L. Schwann cell myelination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Xin, W.; Tan, J.-R.; Zhu, R.-P.; Li, T.; Wang, D.; Kan, S.-S.; Xiong, D.-K.; Li, H.-H.; Zhang, M.-M.; et al. Myelin sheath structure and regeneration in peripheral nerve injury repair. Proc. Natl. Acad. Sci. USA 2019, 116, 22347–22352. [Google Scholar] [CrossRef] [Green Version]
- Ziv-Polat, O.; Shahar, A.; Levy, I.; Skaat, H.; Neuman, S.; Fregnan, F.; Geuna, S.; Grothe, C.; Haastert-Talini, K.; Margel, S. The role of neurotrophic factors conjugated to iron oxide nanoparticles in peripheral nerve regeneration: In vitro studies. Biomed. Res. Int. 2014, 2014, 267808. [Google Scholar] [CrossRef] [Green Version]
- Zoupi, L.; Savvaki, M.; Karagogeos, D. Axons and myelinating glia: An intimate contact. IUBMB Life 2011, 63, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.L.; Brophy, P.; Peles, E. Molecular domains of myelinated axons in the peripheral nervous system. Glia 2008, 56, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, A.D.; Young, J.Z. Internode length and fiber diameter in developing and regenerating nerves. J. Anat. 1948, 82, 110–134. [Google Scholar] [PubMed]
- Hinman, J.D.; Peters, A.; Cabral, H.; Rosene, D.; Hollander, W.; Rasband, M.; Abraham, C.R. Age-related molecular reorganization at the node of Ranvier. J. Comp. Neurol. 2006, 495, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Arancibia-Cárcamo, I.L.; Ford, M.C.; Cossell, L.; Ishida, K.; Tohyama, K.; Attwell, D. Node of Ranvier length as a potential regulator of myelinated axon conduction speed. Elife 2017, 6, e23329. [Google Scholar] [CrossRef]
- Ford, M.; Alexandrova, O.; Cossell, L.; Stange-Marten, A.; Sinclair, J.; Kopp-Scheinpflug, C.; Pecka, M.; Attwell, D.; Grothe, B. Tuning of Ranvier node and internode properties in myelinated axons to adjust action potential timing. Nat. Commun. 2015, 6, 8073. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, E.J.; Scherer, S.S. On the molecular architecture of myelinated fibers. Histochem. Cell Biol. 2000, 113, 1–18. [Google Scholar] [CrossRef]
- Kordeli, E.; Davis, J.; Trapp, B.; Bennett, V. An isoform of ankyrin is localized at nodes of Ranvier in myelinated axons of central and peripheral nerves. J. Cell Biol. 1990, 110, 1341–1352. [Google Scholar] [CrossRef] [Green Version]
- Corfas, G.; Velardez, M.O.; Ko, C.-P.; Ratner, N.; Peles, E. Mechanisms and roles of axon-Schwann cell interactions. J. Neurosci. 2004, 24, 9250–9260. [Google Scholar] [CrossRef] [Green Version]
- Rasband, M.; Trimmer, J.S.; Schwarz, T.L.; Levinson, S.R.; Ellisman, M.H.; Schachner, M.; Shrager, P. Potassium channel distribution, clustering, and function in remyelinating rat axons. J. Neurosci. 1998, 18, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kim, M.J.; Li, L.; Velumian, A.A.; Aui, P.M.; Fehlings, M.G.; Petratos, S. Nogo receptor 1 regulates Caspr distribution at axo–glial units in the central nervous system. Sci. Rep. 2017, 7, 8958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreshuk, A.; Walecki, R.; Koethe, U.; Gierthmuehlen, M.; Plachta, D.; Genoud, C.; Haastert-Talini, K.; Hamprecht, F. Automated tracing of myelinated axons and detection of the nodes of Ranvier in serial images of peripheral nerves. J. Microsc. 2015, 259, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Este, E.; Kamin, D.; Balzarotti, F.; Hell, S.W. Ultrastructural anatomy of nodes of Ranvier in the peripheral nervous system as revealed by STED microscopy. Proc. Natl. Acad. Sci. USA 2017, 114, E191–E199. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, G.; Raimondo, S. Chronically denervated distal nerve stump inhibits peripheral nerve regeneration. Neural. Regen. Res. 2017, 12, 739–740. [Google Scholar] [CrossRef]
- Otani, Y.; Yermakov, L.M.; Dupree, J.L.; Susuki, K. Chronic peripheral nerve compression disrupts paranodal axoglial junctions. Muscle Nerve 2017, 55, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Waxman, S.G.; Kocsis, J.D.; Stys, P.K. (Eds.) 553Regeneration of peripheral nervous system axons. In The Axon: Structure, Function and Pathophysiology; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Puigdellívol-Sánchez, A.; Valero-Cabré, A.; Prats-Galino, A.; Navarro, X.; Molander, C. On the use of fast blue, fluoro-gold and diamidino yellow for retrograde tracing after peripheral nerve injury: Uptake, fading, dye interactions, and toxicity. J. Neurosci. Methods 2002, 115, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Mi, D.; Yuan, Y.; Zhang, Y.; Niu, J.; Wang, Y.; Yan, J.; Yang, Y.; Hu, W. Injection of Fluoro-Gold into the tibial nerve leads to prolonged but reversible functional deficits in rats. Sci. Rep. 2019, 9, 9906. [Google Scholar] [CrossRef] [Green Version]
- Puigdellıvol-Sánchez, A.; Prats-Galino, A.; Ruano-Gil, D.; Molander, C. Persistence of tracer in the application site—A potential confounding factor in nerve regeneration studies. J. Neurosci. Methods 2003, 127, 105–110. [Google Scholar] [CrossRef] [Green Version]
- de Ruiter, G.C.; Spinner, R.J.; Verhaagen, J.; Malessy, M.J.A. Misdirection and guidance of regenerating axons after experimental nerve injury and repair. J. Neurosurg. 2014, 120, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.; Moradzadeh, A.; Hunter, D.A.; Kawamura, D.H.; Puppala, V.K.; Tung, T.H.H.; Mackinnon, S.E.; Myckatyn, T.M. Retrograde labeling in peripheral nerve research: It is not all black and white. J. Reconstr. Microsurg. 2007, 23, 381–389. [Google Scholar] [CrossRef]
- Novikova, L.; Novikov, L.; Kellerth, J. Persistent neuronal labeling by retrograde fluorescent tracers: A comparison between Fast Blue, Fluoro-Gold and various dextran conjugates. J. Neurosci. Methods 1997, 74, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Schmued, L.C.; Kyriakidis, K.; Fallon, J.H.; Ribak, C.E. Neurons containing retrogradely transported Fluoro-Gold exhibit a variety of lysosomal profiles: A combined brightfield, fluorescence, and electron microscopic study. J. Neurocytol. 1989, 18, 333–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radtke, C.; Kocsis, J.D.; Baumgärtner, W.; Vogt, P.M. Fluoro-Ruby as a reliable marker for regenerating fiber tracts. Innov. Surg. Sci. 2017, 2, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Žygelytė, E.; Bernard, M.E.; Tomlinson, J.; Martin, M.J.; Terhorst, A.; Bradford, H.E.; Lundquist, S.A.; Sledziona, M.; Cheetham, J. RetroDISCO: Clearing technique to improve quantification of retrograde labeled motor neurons of intact mouse spinal cords. J. Neurosci. Methods 2016, 271, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Holmes, T.C.; Luo, M.-H.; Beier, K.T.; Horwitz, G.D.; Zhao, F.; Zeng, W.; Hui, M.; Semler, B.L.; Sandri-Goldin, R.M. Viral Vectors for Neural Circuit Mapping and Recent Advances in Trans-synaptic Anterograde Tracers. Neuron 2020, 107, 1029–1047. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hegarty, S.V.; Winter, C.; Wang, F.; He, Z. Viral vectors for neuronal cell type-specific visualization and manipulations. Curr. Opin. Neurobiol. 2020, 63, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, B.; Gao, Z. Lighting Up Neural Circuits by Viral Tracing. Neurosci. Bull. 2022, 38, 1383–1396. [Google Scholar] [CrossRef]
- Im, K.; Mareninov, S.; Diaz, M.F.P.; Yong, W.H. An Introduction to Performing Immunofluorescence Staining. Methods Mol. Biol. 2019, 1897, 299–311. [Google Scholar]
- Kemp, S.W.P.; Syed, S.; Walsh, S.K.; Zochodne, D.W.; Midha, R. Collagen nerve conduits promote enhanced axonal regeneration, schwann cell association, and neovascularization compared to silicone conduits. Tissue Eng. Part A 2009, 15, 1975–1988. [Google Scholar] [CrossRef]
- Verdu, E.; Navarro, X. Comparison of immunohistochemical and functional reinnervation of skin and muscle after peripheral nerve injury. Exp. Neurol. 1997, 146, 187–198. [Google Scholar] [CrossRef]
- Fornasari, B.E.; Zen, F.; Nato, G.; Fogli, M.; Luzzati, F.; Ronchi, G.; Raimondo, S.; Gambarotta, G. Blood Vessels: The Pathway Used by Schwann Cells to Colonize Nerve Conduits. Int. J. Mol. Sci. 2022, 23, 2254. [Google Scholar] [CrossRef] [PubMed]
- Križ, J.; Zhu, Q.; Julien, J.-P.; Padjen, A.L. Electrophysiological properties of axons in mice lacking neurofilament subunit genes: Disparity between conduction velocity and axon diameter in absence of NF-H. Brain Res. 2000, 885, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Hausrat, T.J.; Radwitz, J.; Lombino, F.L.; Breiden, P.; Kneussel, M. Alpha- and beta-tubulin isotypes are differentially expressed during brain development. Dev. Neurobiol. 2021, 81, 333–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornaro, M.; Lee, J.; Raimondo, S.; Nicolino, S.; Geuna, S.; Giacobini-Robecchi, M. Neuronal intermediate filament expression in rat dorsal root ganglia sensory neurons: An in vivo and in vitro study. Neuroscience 2008, 153, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, S.M.; Vartanian, T.; Fishman, M. GAP-43 as a plasticity protein in neuronal form and repair. J. Neurobiol. 1992, 23, 507–520. [Google Scholar] [CrossRef]
- Romeo, H.E.; Weihe, E.; Müller, S.; Vollrath, L. Protein gene product (PGP) 9.5 immunoreactivity in nerve fibers and pinealocytes of guinea-pig pineal gland: Interrelationship with tyrosine- hydroxylase- and neuropeptide-Y-immunoreactive nerve fibers. Cell Tissue Res. 1993, 271, 477–484. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, K.K. Glial fibrillary acidic protein: From intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015, 38, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Smith-Slatas, C.; Barbarese, E. Myelin basic protein gene dosage effects in the PNS. Mol. Cell Neurosci. 2000, 15, 343–354. [Google Scholar] [CrossRef]
- Morton, P.D.; Dellarole, A.; Theus, M.H.; Walters, W.M.; Berge, S.S.; Bethea, J.R. Activation of NF-kappaB in Schwann cells is dispensable for myelination in vivo. J. Neurosci. 2013, 33, 9932–9936. [Google Scholar] [CrossRef] [Green Version]
- Muratori, L.; Gnavi, S.; Fregnan, F.; Mancardi, A.; Raimondo, S.; Perroteau, I.; Geuna, S. Evaluation of Vascular Endothelial Growth Factor (VEGF) and Its Family Member Expression After Peripheral Nerve Regeneration and Denervation. Anat. Rec. 2018, 301, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Du, J.; Qing, L.; Mee, T.; Xu, X.; Wang, Z.; Xu, C.; Jia, X. Identification of sensory and motor nerve fascicles by immunofluorescence staining after peripheral nerve injury. J. Transl. Med. 2021, 19, 207. [Google Scholar] [CrossRef] [PubMed]
- Renier, N.; Wu, Z.; Simon, D.J.; Yang, J.; Ariel, P.; Tessier-Lavigne, M. iDISCO: A simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 2014, 159, 896–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huesing, C.; Qualls-Creekmore, E.; Lee, N.; François, M.; Torres, H.; Zhang, R.; Burk, D.H.; Yu, S.; Morrison, C.D.; Berthoud, H.; et al. Sympathetic innervation of inguinal white adipose tissue in the mouse. J. Comp. Neurol. 2021, 529, 1465–1485. [Google Scholar] [CrossRef] [PubMed]
- Sathyanesan, A.; Ogura, T.; Lin, W. Automated measurement of nerve fiber density using line intensity scan analysis. J. Neurosci. Methods 2012, 206, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Richard, L.; Védrenne, N.; Vallat, J.-M.; Funalot, B. Characterization of Endoneurial Fibroblast-like Cells from Human and Rat Peripheral Nerves. J. Histochem. Cytochem. 2014, 62, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.; Hernández, I.C.; Wang, W.; Yin, K.; Sundback, C.A.; Wegst, U.G.; Jowett, N. Fluorescent Reporter Mice for Nerve Guidance Conduit Assessment: A High-Throughput in vivo Model. Laryngoscope 2018, 128, E386–E392. [Google Scholar] [CrossRef]
- Tu, N.H.; Katano, T.; Matsumura, S.; Pham, V.M.; Muratani, T.; Minami, T.; Ito, S. Role of c-Jun N-terminal kinase in late nerve regeneration monitored by in vivo imaging of thy1-yellow fluorescent protein transgenic mice. Eur. J. Neurosci. 2016, 43, 548–560. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Sanes, J.; Lichtman, J. Pre-existing pathways promote precise projection patterns. Nat. Neurosci. 2002, 5, 861–867. [Google Scholar] [CrossRef]
- Magill, C.K.; Tong, A.; Kawamura, D.; Hayashi, A.; Hunter, D.A.; Parsadanian, A.; MacKinnon, S.E.; Myckatyn, T.M. Reinnervation of the tibialis anterior following sciatic nerve crush injury: A confocal microscopic study in transgenic mice. Exp. Neurol. 2007, 207, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.M.; Borschel, G.H.; Santosa, K.B.; Flagg, E.R.; Tong, A.Y.; Kasukurthi, R.; Newton, P.; Yan, Y.; Hunter, D.A.; Johnson, P.J.; et al. A transgenic rat expressing green fluorescent protein (GFP) in peripheral nerves provides a new hindlimb model for the study of nerve injury and regeneration. J. Neurosci. Methods 2012, 204, 19–27. [Google Scholar] [CrossRef]
- Kemp, S.W.P.; Phua, P.D.; Stanoulis, K.N.; Wood, M.D.; Liu, E.H.; Gordon, T.; Borschel, G.H. Functional recovery following peripheral nerve injury in the transgenic Thy1-GFP rat. J. Peripher. Nerv. Syst. 2013, 18, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Lubischer, J.; Kang, H.; Tian, L.; Mikesh, M.; Marks, A.; Scofield, V.L.; Maika, S.; Newman, C.; Krieg, P.; et al. Fluorescent proteins expressed in mouse transgenic lines mark subsets of glia, neurons, macrophages, and dendritic cells for vital examination. J. Neurosci. 2004, 24, 10999–11009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vleggeert-Lankamp, C.L. The role of evaluation methods in the assessment of peripheral nerve regeneration through synthetic conduits: A systematic review. Laboratory investigation. J. Neurosurg. 2007, 107, 1168–1189. [Google Scholar] [CrossRef]
- Frykman, G.K.; McMillan, P.; Yegge, S. A review of experimental methods measuring peripheral nerve regeneration in animals. Orthop. Clin. North Am. 1988, 19, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.D.; Kemp, S.W.; Weber, C.; Borschel, G.H.; Gordon, T. Outcome measures of peripheral nerve regeneration. Ann. Anat. 2011, 193, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, M.; Razavi, S.; Bakhtiari, A. The advances in nerve tissue engineering: From fabrication of nerve conduit to in vivo nerve regeneration assays. J. Tissue Eng. Regen. Med. 2019, 13, 2077–2100. [Google Scholar] [CrossRef]
- Janjic, J.M.; Gorantla, V.S. Peripheral Nerve Nanoimaging: Monitoring Treatment and Regeneration. AAPS J. 2017, 19, 1304–1316. [Google Scholar] [CrossRef]




| Name | Excitation | Emission |
|---|---|---|
| Fast Blue (FB) | 365 | 420 |
| Diamidino Yellow (DY) | 365 | >500 |
| True Blue (TB) | 365 | 405 |
| Granular Blue (GB) | 365 | 410 |
| Antibody | Role | Target | References |
|---|---|---|---|
| NF-200 | Intermediate filament protein of neuronal cytoskeleton | Axons | Kriz and colleagues [63] |
| βIII tubulin | Cytoskeletal component expressed almost exclusively by neurons and axons | Axons and neurons | Hausrat and colleagues [64] |
| Peripherin | Type III intermediate filament protein | Small-sized axons | Fornaro and colleagues [65] |
| GAP43 | Major component of “growth cones” of elongating axons | Developing and regenerating neurons | Strittmatter and colleagues [66] |
| PGP 9.5/UCHL-1 | Neuronal specific protein member of ubiquitin carboxyl-terminal hydrolase domain | Neurons and nerve fibers | Romeo and colleagues [67] |
| GFAP | Type III intermediate filament protein | Non-myelinating Schwann cells | Yang and colleagues [68] |
| MBP | Major constituents of myelin sheath of Schwann cells | Myelinating Schwann cells | Smith-Slatas and colleagues [69] |
| S100 β | Calcium-binding protein | Adult Schwann cells | Morton and colleagues [70] |
| Morphological Methods | Type of Analysis | Limits of the Technique | References |
|---|---|---|---|
| Histological Staining |
|
| Raimondo and colleagues [12] |
| Transmission Electron Microscopy |
|
| |
| Retrograde Labeling |
|
|
|
| Immunohistochemistry Analysis |
|
| |
| Transgenic Models |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronchi, G.; Fregnan, F.; Muratori, L.; Gambarotta, G.; Raimondo, S. Morphological Methods to Evaluate Peripheral Nerve Fiber Regeneration: A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 1818. https://doi.org/10.3390/ijms24031818
Ronchi G, Fregnan F, Muratori L, Gambarotta G, Raimondo S. Morphological Methods to Evaluate Peripheral Nerve Fiber Regeneration: A Comprehensive Review. International Journal of Molecular Sciences. 2023; 24(3):1818. https://doi.org/10.3390/ijms24031818
Chicago/Turabian StyleRonchi, Giulia, Federica Fregnan, Luisa Muratori, Giovanna Gambarotta, and Stefania Raimondo. 2023. "Morphological Methods to Evaluate Peripheral Nerve Fiber Regeneration: A Comprehensive Review" International Journal of Molecular Sciences 24, no. 3: 1818. https://doi.org/10.3390/ijms24031818