

Metaproteomic Analysis of an Oral Squamous Cell Carcinoma Dataset Suggests Diagnostic Potential of the Mycobiome


Abstract
:1. Introduction
2. Results
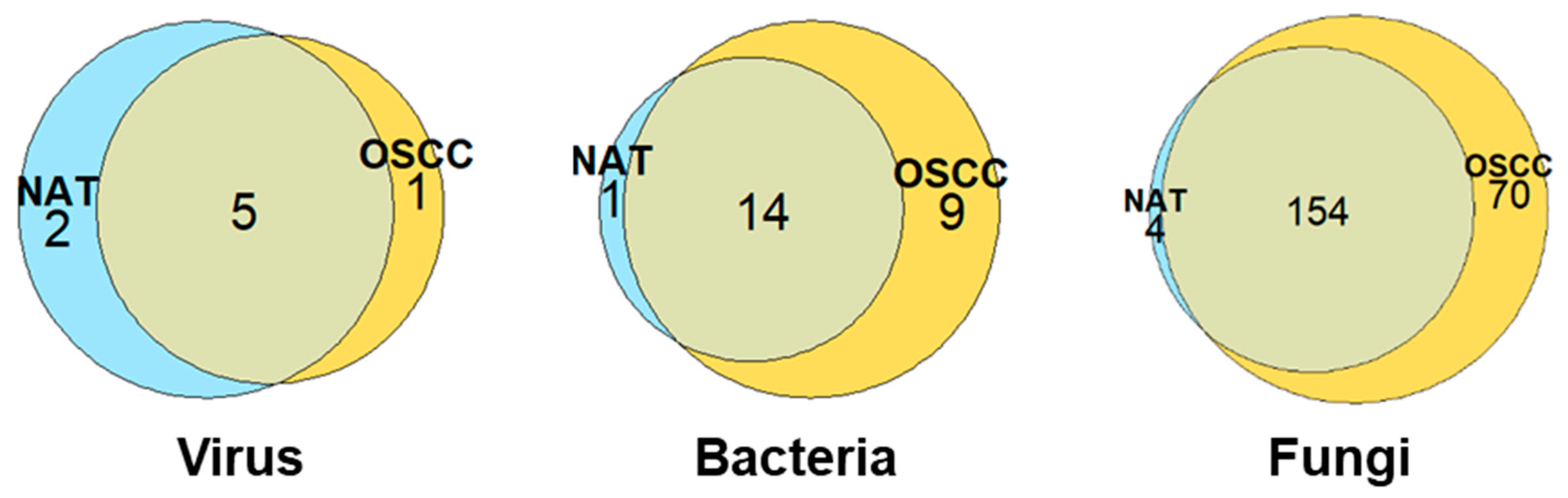
2.1. An Increased Overall Microbial Species Diversity Is Observed in OSCC Conditions
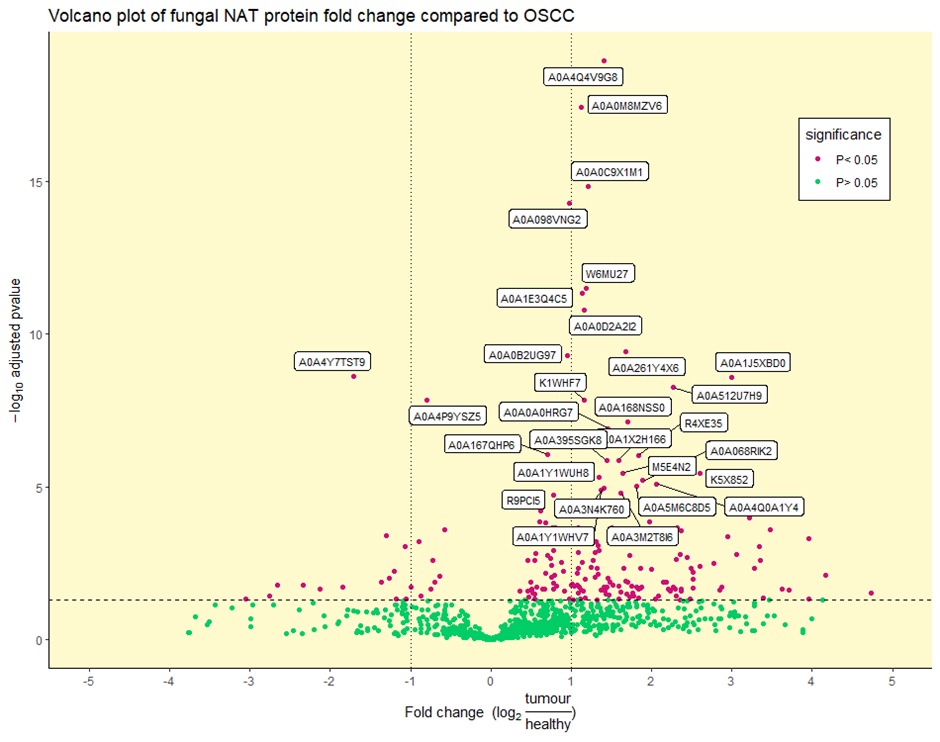
2.2. Identification of Differentially Abundant Fungal Proteins
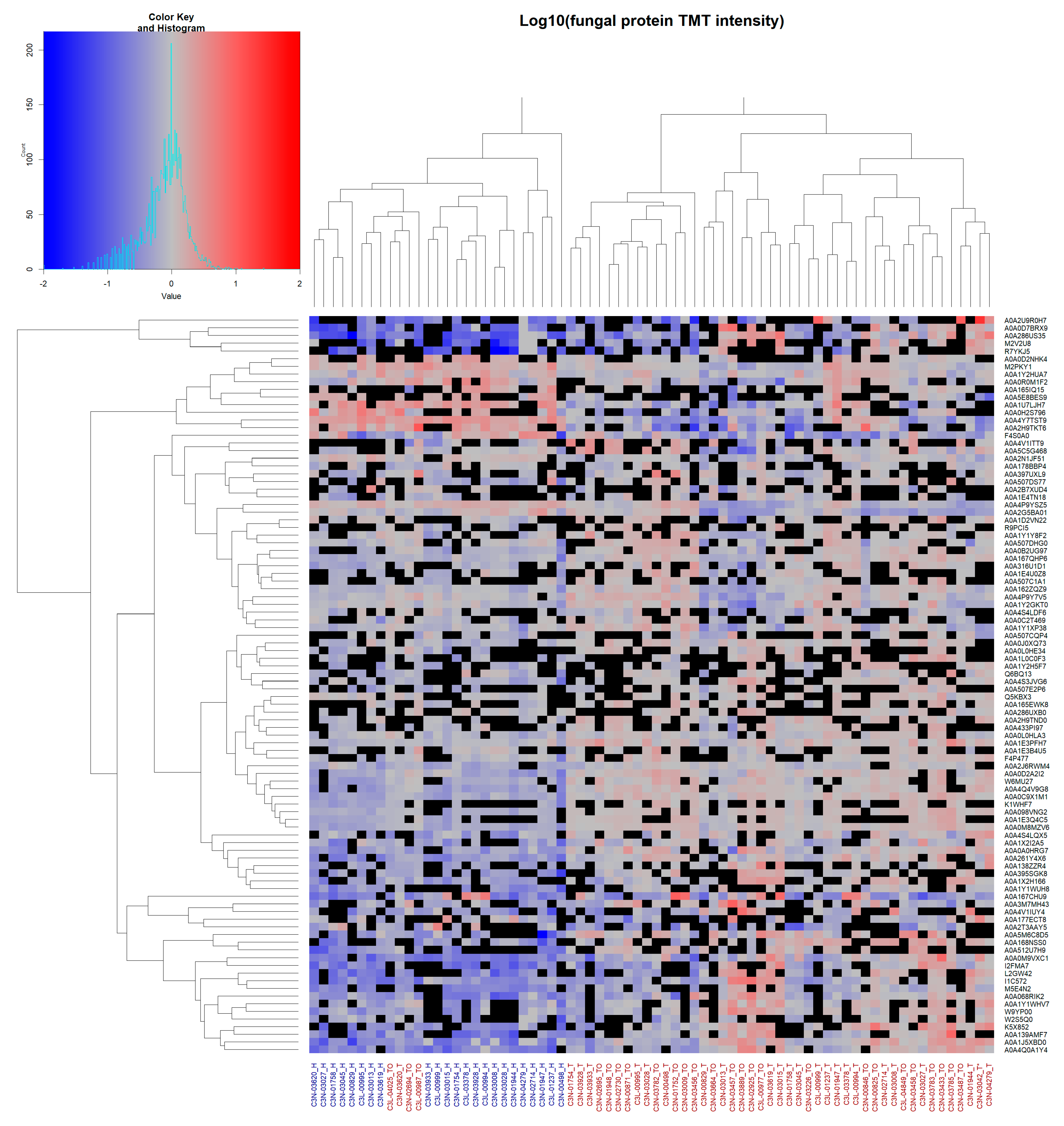
2.3. Hierarchical Clustering Reveals Diagnostic Potential of Fungal Microbiome
3. Discussion
3.1. Enrichment of Nucleocytoplasmic Large DNA Viruses in the Oral Virome
3.2. Periodontal Pathogens and Opportunistic Bacteria Are Enriched in OSCC
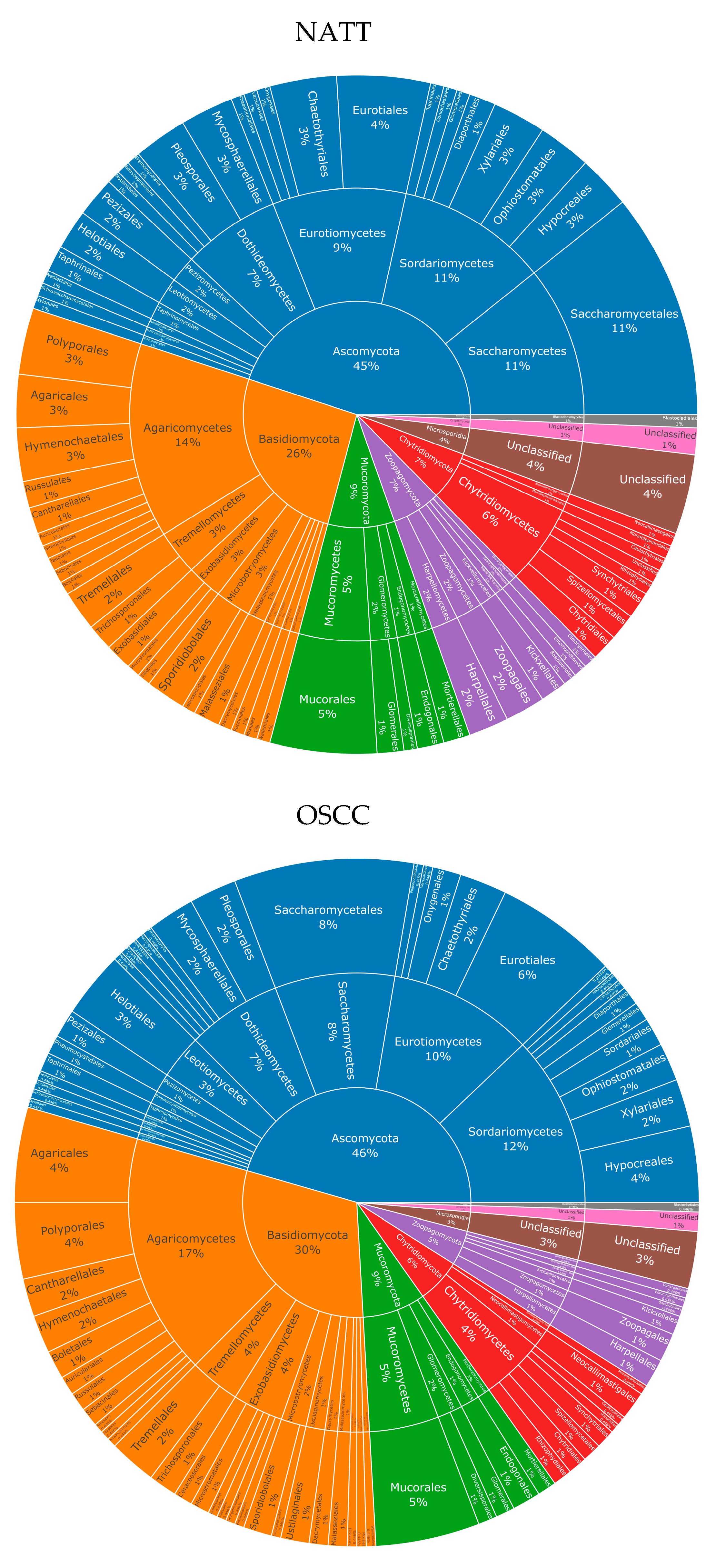
3.3. An Unexpected Fungal Diversity Is Observed in the OSCC Patient Samples
3.4. Fungal Proteins Implicate Pathogens Capable of Soft Tissue Damage
3.5. Clustering of the Mycobiome Protein Profile Shows Diagnostic Potential
4. Methods
4.1. Data Collection
4.2. Trans-Proteomic Pipeline Analysis
4.3. Secondary Analysis and Visualisation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-Z.; Xie, L.; Shang, Z.-J. Burden of Oral Cancer on the 10 Most Populous Countries from 1990 to 2019: Estimates from the Global Burden of Disease Study 2019. Int. J. Environ. Res. Public Health 2022, 19, 875. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinkugbe, A.A.; Garcia, D.T.; Brickhouse, T.H.; Mosavel, M. Lifestyle Risk Factor Related Disparities in Oral Cancer Examination in the U.S: A Population-Based Cross-Sectional Study. BMC Public Health 2020, 20, 153. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.-C.; Kung, P.-T.; Lung, C.-H.; Tsai, M.-H.; Liu, S.-A.; Chiu, L.-T.; Huang, K.-H.; Tsai, W.-C. Assessment of the Risk of Oral Cancer Incidence in A High-Risk Population and Establishment of a Predictive Model for Oral Cancer Incidence Using a Population-Based Cohort in Taiwan. Int. J. Environ. Res. Public Health 2020, 17, 665. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.H.; Hu, C.Y.; He, H.R.; Li, Y.J.; Lyu, J. Global and Regional Burdens of Oral Cancer from 1990 to 2017: Results from the Global Burden of Disease Study. Cancer Commun. 2020, 40, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheraghlou, S.; Schettino, A.; Zogg, C.K.; Judson, B.L. Changing Prognosis of Oral Cancer: An Analysis of Survival and Treatment between 1973 and 2014. Laryngoscope 2018, 128, 2762–2769. [Google Scholar] [CrossRef]
- He, S.; Chakraborty, R.; Ranganathan, S. Proliferation and Apoptosis Pathways and Factors in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 1562. [Google Scholar] [CrossRef]
- Chakraborty, R.; Hu, H.; Mangani, A.S.; Vickery, K.; Ranganathan, S. Combined Bacterial Antigen Lipopolysaccharide and Lipoteichoic Acid Increase Cal 27 Oral Cancer Cell Proliferation. Dent. Oral Maxillofac. Res. 2021, 4, 1–6. [Google Scholar]
- Zhang, L.; Liu, Y.; Zheng, H.J.; Zhang, C.P. The Oral Microbiota May Have Influence on Oral Cancer. Front. Cell. Infect. Microbiol. 2020, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Marwick, C. Helicobacter: New Name, New Hypothesis Involving Type of Gastric Cancer. JAMA 1990, 264, 2724–2727. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Sipponen, P.; Naumann, M.; Moayyedi, P.; Mégraud, F.; Xiao, S.-D.; Sugano, K.; Nyrén, O.; Lejondal, H. Pylori-Gastric Cancer Task Force. Helicobacter Pylori Eradication Has the Potential to Prevent Gastric Cancer: A State-of-the-Art Critique. Off. J. Am. Coll. Gastroenterol. 2005, 100, 2100–2115. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Jobin, C. The Microbiome and Cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Z.; Qi, Y.; Wen, X.; Zhang, L. Metagenomic Analysis Reveals a Changing Microbiome Associated with the Depth of Invasion of Oral Squamous Cell Carcinoma. Front. Microbiol. 2022, 13, 795777. [Google Scholar] [CrossRef]
- Torralba, M.G.; Aleti, G.; Li, W.; Moncera, K.J.; Lin, Y.-H.; Yu, Y.; Masternak, M.M.; Golusinski, W.; Golusinski, P.; Lamperska, K.; et al. Oral Microbial Species and Virulence Factors Associated with Oral Squamous Cell Carcinoma. Microb. Ecol. 2021, 82, 1030–1046. [Google Scholar] [CrossRef]
- Yang, C.Y.; Yeh, Y.M.; Yu, H.Y.; Chin, C.Y.; Hsu, C.W.; Liu, H.; Huang, P.J.; Hu, S.N.; Liao, C.T.; Chang, K.P.; et al. Oral Microbiota Community Dynamics Associated with Oral Squamous Cell Carcinoma Staging. Front. Microbiol. 2018, 9, 862. [Google Scholar] [CrossRef] [Green Version]
- Gallimidi, A.B.; Fischman, S.; Revach, B.; Bulvik, R.; Maliutina, A.; Rubinstein, A.M.; Nussbaum, G.; Elkin, M. Periodontal Pathogens Porphyromonas gingivalis and Fusobacterium nucleatum Promote Tumor Progression in an Oral-Specific Chemical Carcinogenesis Model. Oncotarget 2015, 6, 22613–22623. [Google Scholar] [CrossRef] [Green Version]
- Das, B.R.; Nagpal, J.K. Understanding the Biology of Oral Cancer. Med. Sci. Monit. 2002, 8, 258–268. [Google Scholar]
- Thompson, I.O.C.; van der Bijl, P.; van Wyk, C.W.; van Eyk, A.D. A Comparative Light-Microscopic, Electron-Microscopic and Chemical Study of Human Vaginal and Buccal Epithelium. Arch. Oral Biol. 2001, 46, 1091–1098. [Google Scholar] [CrossRef]
- Campisi, G.; Panzarella, V.; Giuliani, M.; Lajolo, C.; di Fede, O.; Falaschini, S.; di Liberto, C.; Scully, C.; Lo Muzio, L. Human Papillomavirus: Its Identikit and Controversial Role in Oral Oncogenesis, Premalignant and Malignant Lesions (Review). Int. J. Oncol. 2007, 30, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Park, N.-H.; Min, B.-M.; Li, S.; Huang, M.Z.; Doniger, J. Immortalization of Normal Human Oral Keratinocytes with Type 16 Human Papillomavirus. Carcinogenesis 1991, 12, 1627–1631. [Google Scholar] [CrossRef]
- Lechner, M.; Liu, J.; Masterson, L.; Fenton, T.R. HPV-Associated Oropharyngeal Cancer: Epidemiology, Molecular Biology and Clinical Management. Nat. Rev. Clin. Oncol. 2022, 19, 306–327. [Google Scholar] [CrossRef]
- Farah, C.S. Molecular Landscape of Head and Neck Cancer and Implications for Therapy. Ann. Transl. Med. 2021, 9, 915. [Google Scholar] [CrossRef]
- Wu, W.; Wang, Z.; Zhou, Z. Role of the Human Papillomavirus in Malignant Transformation of Oral Leukoplakia Distinct from Oropharyngeal Squamous Cell Carcinoma: A Study of 76 Patients with Internal-Control Specimens. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 128, 273–279. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Clifford, G.M.; Boyle, P.; Franceschi, S. Human Papillomavirus Types in Head and Neck Squamous Cell Carcinomas Worldwide: A Systematic Review. Cancer Epidemiol. Biomark. Prev. 2005, 14, 467–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidry, J.; Birdwell, C.; Scott, R. Epstein-Barr Virus in the Pathogenesis of Oral Cancers. Oral Dis. 2018, 24, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.I.; Fauci, A.S.; Varmus, H.; Nabel, G.J. Epstein-Barr Virus: An Important Vaccine Target for Cancer Prevention. Sci. Transl. Med. 2011, 3, 107fs7. [Google Scholar] [CrossRef] [Green Version]
- Mäkinen, A.; Nawaz, A.; Mäkitie, A.; Meurman, J.H. Role of Non-Albicans Candida and Candida albicans in Oral Squamous Cell Cancer Patients. J. Oral Maxillofac. Surg. 2018, 76, 2564–2571. [Google Scholar] [CrossRef] [Green Version]
- Gainza-Cirauqui, M.L.; Nieminen, M.T.; Novak Frazer, L.; Aguirre-Urizar, J.M.; Moragues, M.D.; Rautemaa, R. Production of Carcinogenic Acetaldehyde by Candida albicans from Patients with Potentially Malignant Oral Mucosal Disorders. J. Oral Pathol. Med. 2013, 42, 243–249. [Google Scholar] [CrossRef]
- Perera, M.; Al-Hebshi, N.N.; Perera, I.; Ipe, D.; Ulett, G.C.; Speicher, D.J.; Chen, T.; Johnson, N.W. A Dysbiotic Mycobiome Dominated by Candida albicans Is Identified within Oral Squamous-Cell Carcinomas. J. Oral Microbiol. 2017, 9, 1385369. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Chen, L.; Savage, S.R.; Eguez, R.V.; Dou, Y.; Li, Y.; da Veiga Leprevost, F.; Jaehnig, E.J.; Lei, J.T.; Wen, B.; et al. Proteogenomic Insights into the Biology and Treatment of HPV-Negative Head and Neck Squamous Cell Carcinoma. Cancer Cell 2021, 39, 361–379.e16. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Farrah, T.; Lam, H.; Sun, Z.; Nilsson, E.; Pratt, B.; Prazen, B.; Eng, J.K.; et al. A Guided Tour of the TPP. Proteomics 2011, 10, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.C.; Neves, L.X.; Trino, L.D.; Carnielli, C.M.; Lopes, A.F.B.; Yokoo, S.; Pauletti, B.A.; Domingues, R.R.; Sá, J.O.; Persinoti, G.; et al. Meta-Omics Analysis Indicates the Saliva Microbiome and Its Proteins Associated with the Prognosis of Oral Cancer Patients. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2021, 1869, 140659. [Google Scholar] [CrossRef] [PubMed]
- Shay, E.; Sangwan, N.; Padmanabhan, R.; Lundy, S.; Burkey, B.; Eng, C. Bacteriome and Mycobiome and Bacteriome-Mycobiome Interactions in Head and Neck Squamous Cell Carcinoma. Oncotarget 2020, 11, 2375–2386. [Google Scholar] [CrossRef]
- Colson, P.; de Lamballerie, X.; Yutin, N.; Asgari, S.; Bigot, Y.; Bideshi, D.K.; Cheng, X.-W.; Federici, B.A.; van Etten, J.L.; Koonin, E.V.; et al. “Megavirales”, a Proposed New Order for Eukaryotic Nucleocytoplasmic Large DNA Viruses. Arch. Virol. 2013, 158, 2517–2521. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in Microalgae Incorporation into Innovative Food Products with Potential Health Benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Van der Spiegel, M.; Noordam, M.Y.; van der Fels-Klerx, H.J. Safety of Novel Protein Sources (Insects, Microalgae, Seaweed, Duckweed, and Rapeseed) and Legislative Aspects for Their Application in Food and Feed Production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Lasek, R.; Szuplewska, M.; Mitura, M.; Decewicz, P.; Chmielowska, C.; Pawłot, A.; Sentkowska, D.; Czarnecki, J.; Bartosik, D. Genome Structure of the Opportunistic Pathogen Paracoccus yeei (Alphaproteobacteria) and Identification of Putative Virulence Factors. Front. Microbiol. 2018, 9, 2553. [Google Scholar] [CrossRef] [Green Version]
- Çiçek, M.; Hasçelik, G.; Müştak, H.K.; Diker, K.S.; Şener, B. Accurate Diagnosis of Pseudomonas Luteola in Routine Microbiology Laboratory: On the Occasion of Two Isolates. Mikrobiyoloji Bul. 2016, 50, 621–624. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus Epidermidis—The “accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Malani, A.N.; Aronoff, D.M.; Bradley, S.F.; Kauffman, C.A. Cardiobacterium hominis Endocarditis: Two Cases and a Review of the Literature. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heldt Manica, L.A.; Cohen, P.R. Staphylococcus lugdunensis Infections of the Skin and Soft Tissue: A Case Series and Review. Dermatol. Ther. 2017, 7, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Al-Hebshi, N.N.; Borgnakke, W.S.; Johnson, N.W. The Microbiome of Oral Squamous Cell Carcinomas: A Functional Perspective. Curr. Oral Health Rep. 2019, 6, 145–160. [Google Scholar] [CrossRef] [Green Version]
- You, Y.-O.; Kim, K.-J.; Min, B.-M.; Chung, C.-P. Staphylococcus lugdunensis—A Potential Pathogen in Oral Infection. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontology 1999, 88, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Feller, L.; Altini, M.; Lemmer, J. Inflammation in the Context of Oral Cancer. Oral Oncol. 2013, 49, 887–892. [Google Scholar] [CrossRef]
- Niklander, S.E. Inflammatory Mediators in Oral Cancer: Pathogenic Mechanisms and Diagnostic Potential. Front. Oral Health 2021, 2, 642238. [Google Scholar] [CrossRef] [PubMed]
- Abusleme, L.; Dupuy, A.K.; Dutzan, N.; Silva, N.; Burleson, J.A.; Strausbaugh, L.D.; Gamonal, J.; Diaz, P.I. The Subgingival Microbiome in Health and Periodontitis and Its Relationship with Community Biomass and Inflammation. ISME J. 2013, 7, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Anesti, V.; McDonald, I.R.; Ramaswamy, M.; Wade, W.G.; Kelly, D.P.; Wood, A.P. Isolation and Molecular Detection of Methylotrophic Bacteria Occurring in the Human Mouth. Environ. Microbiol. 2005, 7, 1227–1238. [Google Scholar] [CrossRef]
- Genco, R.J.; LaMonte, M.J.; McSkimming, D.I.; Buck, M.J.; Li, L.; Hovey, K.M.; Andrews, C.A.; Sun, Y.; Tsompana, M.; Zheng, W.; et al. The Subgingival Microbiome Relationship to Periodontal Disease in Older Women. J. Dent. Res. 2019, 98, 975–984. [Google Scholar] [CrossRef]
- Poco, S.E.; Nakazawa, F.; Sato, M.; Hoshino, E. Eubacterium minutum sp. nov., Isolated from Human Periodontal Pockets. Int. J. Syst. Bacteriol. 1996, 46, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Pehl, M.J.; Jamieson, W.D.; Kong, K.; Forbester, J.L.; Fredendall, R.J.; Gregory, G.A.; McFarland, J.E.; Healy, J.M.; Orwin, P.M. Genes That Influence Swarming Motility and Biofilm Formation in Variovorax Paradoxus EPS. PLoS ONE 2012, 7, e31832. [Google Scholar] [CrossRef] [PubMed]
- Irani, S.; Barati, I.; Badiei, M. Periodontitis and Oral Cancer—Current Concepts of the Etiopathogenesis. Oncol. Rev. 2020, 14, 465. [Google Scholar] [CrossRef] [PubMed]
- Javed, F.; Warnakulasuriya, S. Is There a Relationship between Periodontal Disease and Oral Cancer? A Systematic Review of Currently Available Evidence. Crit. Rev. Oncol. Hematol. 2016, 97, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I.; Hong, B.Y.; Dupuy, A.K.; Strausbaugh, L.D. Mining the Oral Mycobiome: Methods, Components, and Meaning. Virulence 2017, 8, 313–323. [Google Scholar] [CrossRef]
- Baker, J.L.; Bor, B.; Agnello, M.; Shi, W.; He, X. Ecology of the Oral Microbiome: Beyond Bacteria. Trends Microbiol. 2017, 25, 362–374. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the Oral Fungal Microbiome (Mycobiome) in Healthy Individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [Green Version]
- Huffnagle, G.B.; Noverr, M.C. The Emerging World of the Fungal Microbiome. Trends Microbiol. 2013, 21, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H.; Morris, A. The Emerging Importance and Challenges of the Human Mycobiome. Virulence 2017, 8, 310–312. [Google Scholar] [CrossRef]
- Kannan, R. Oral Biopsy: Principles and Practice. In Textbook of Oral Cancer: Prevention, Diagnosis and Management; Warnakulasuriya, S., Greenspan, J.S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-32316-5. [Google Scholar]
- Malik, N.A. Textbook of Oral and Maxillofacial Surgery, 2nd ed.; Jaypee Brothers Medical Publishers Ltd.: New Delhi, India, 2008. [Google Scholar]
- Miloro, M.; Peterson, L.J. Peterson’s Principles of Oral and Maxillofacial Surgery, 3rd ed.; Shelton, C.T., Ed.; Springer: New York, NY, USA, 2012; ISBN 978-1-60795-111-7. [Google Scholar]
- Rogawansamy, S.; Gaskin, S.; Taylor, M.; Pisaniello, D. An Evaluation of Antifungal Agents for the Treatment of Fungal Contamination in Indoor Air Environments. Int. J. Environ. Res. Public Health 2015, 12, 6319–6332. [Google Scholar] [CrossRef] [Green Version]
- Leikin, J.B.; Paloucek, F.P. (Eds.) Poisoning and Toxicology Handbook, 4th ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-4479-9. [Google Scholar]
- Lachapelle, J.-M.; Castel, O.; Casado, A.F.; Leroy, B.; Micali, G.; Tennstedt, D.; Lambert, J. Antiseptics in the Era of Bacterial Resistance: A Focus on Povidone Iodine. Clin. Pract. 2013, 10, 579–592. [Google Scholar] [CrossRef]
- Fathilah, A.R.; Himratul-Aznita, W.H.; Fatheen, A.R.N.; Suriani, K.R. The Antifungal Properties of Chlorhexidine Digluconate and Cetylpyrinidinium Chloride on Oral Candida. J. Dent. 2012, 40, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Tabe, Y.; Yamada, T.; Misawa, S.; Oguri, T.; Ohsaka, A.; Miida, T. Comparison of Antifungal Activities of Gentian Violet and Povidone-Iodine Against Clinical Isolates of Candida Species and Other Yeasts: A Framework to Establish Topical Disinfectant Activities. Mycopathologia 2012, 173, 21–25. [Google Scholar] [CrossRef]
- Kanagalingam, J.; Feliciano, R.; Hah, J.H.; Labib, H.; Le, T.A.; Lin, J.-C. Practical Use of Povidone-iodine Antiseptic in the Maintenance of Oral Health and in the Prevention and Treatment of Common Oropharyngeal Infections. Int. J. Clin. Pract. 2015, 69, 1247–1256. [Google Scholar] [CrossRef]
- Theofilou, V.I.; Alfaifi, A.; Montelongo-Jauregui, D.; Pettas, E.; Georgaki, M.; Nikitakis, N.G.; Jabra-Rizk, M.; Sultan, A.S. The Oral Mycobiome: Oral Epithelial Dysplasia and Oral Squamous Cell Carcinoma. J. Oral Pathol. Med. 2022, 51, 413–420. [Google Scholar] [CrossRef]
- Jennings, Z.; Kable, K.; Halliday, C.L.; Nankivell, B.J.; Kok, J.; Wong, G.; Chen, S.C.-A. Verruconis gallopava Cardiac and Endovascular Infection with Dissemination after Renal Transplantation: Case Report and Lessons Learned. Med. Mycol. Case Rep. 2017, 15, 5–8. [Google Scholar] [CrossRef]
- Irshad, M.; Nasir, N.; Hashmi, U.H.; Farooqi, J.; Mahmood, S.F. Invasive Pulmonary Infection by Syncephalastrum Species: Two Case Reports and Review of Literature. IDCases 2020, 21, e00913. [Google Scholar] [CrossRef]
- Jeffries, P.; Young, T.W.K. Ultrastructure of the Haustorial Apparatus of Dimargaris cristalligena. Ann. Bot. 1981, 47, 107–119. [Google Scholar] [CrossRef]
- Kemna, M.E.; Neri, R.C.; Ali, R.; Salkin, I.F. Cokeromyces recurvatus, a Mucoraceous Zygomycete Rarely Isolated in Clinical Laboratories. J. Clin. Microbiol. 1994, 32, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Kalaskar, R.R.; Kalaskar, A.R.; Ganvir, S. Oral Mucormycosis in an 18-Month-Old Child: A Rare Case Report with a Literature Review. J. Korean Assoc. Oral Maxillofac. Surg. 2016, 42, 105–110. [Google Scholar] [CrossRef]
- Rajendra Santosh, A.B.; Muddana, K.; Bakki, S.R. Fungal Infections of Oral Cavity: Diagnosis, Management, and Association with COVID-19. SN Compr. Clin. Med. 2021, 3, 1373–1384. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Wang, H.; Retuerto, M.; Zhang, H.; Burkey, B.; Ghannoum, M.A.; Eng, C. Bacteriome and Mycobiome Associations in Oral Tongue Cancer. Oncotarget 2017, 8, 97273–97289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, I.; Ghose, A.; Gupta, D.; Manasvi, M.; Bhandari, S.; Das, A.; Sanchez, E.; Boussios, S. COVID-19, Mucormycosis and Cancer: The Triple Threat—Hypothesis or Reality? J. Pers. Med. 2022, 12, 1119. [Google Scholar] [CrossRef] [PubMed]
- Pagano, L.; Offidani, M.; Fianchi, L.; Nosari, A.; Candoni, A.; Picardi, M.; Corvatta, L.; D’Antonio, D.; Girmenia, C.; Martino, P.; et al. Mucormycosis in Hematologic Patients. Haematologica 2004, 89, 207–214. [Google Scholar] [PubMed]
- Meyer, R.D. Phycomycosis Complicating Leukemia and Lymphoma. Ann. Intern. Med. 1972, 77, 871. [Google Scholar] [CrossRef]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the Human Oral Mycobiome with Improved Practices in Amplicon-Based Taxonomy: Discovery of Malassezia as a Prominent Commensal. PLoS ONE 2014, 9, e90899. [Google Scholar] [CrossRef] [Green Version]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The Fungal Mycobiome Promotes Pancreatic Oncogenesis via Activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Almeida, O.P.; Jorge, J., Jr.; Scully, C. Paracoccidioidomycosis of the Mouth: An Emerging Deep Mycosis. Crit. Rev. Oral Biol. Med. 2003, 14, 268–274. [Google Scholar] [CrossRef]
- De Abreu e Silva, M.À.; Salum, F.G.; Figueiredo, M.A.; Cherubini, K. Important Aspects of Oral Paracoccidioidomycosis—A Literature Review: Oral Paracoccidioidomycosis. Mycoses 2013, 56, 189–199. [Google Scholar] [CrossRef]
- Azevedo, R.S.; Gouvêa, A.F.; Lopes, M.A.; Corrêa, M.B.; Jorge, J. Synchronous Oral Paracoccidioidomycosis and Oral Squamous Cell Carcinomas with Submandibular Enlargement. Med. Mycol. 2011, 49, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A.; Conceição, Y.M.T.; Kono, A.; Rivitti, E.; Campos, A.F.; Campos, S.V. Neoplasia and Paracoccidioidomycosis. Mycopathologia 2008, 165, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Rodrigues, G.; Severo, C.B.; de Mattos Oliveira, F.; da Silva Moreira, J.; Prolla, J.C.; Severo, L.C. Association between Paracoccidioidomycosis and Cancer. J. Bras. Pneumol. 2010, 36, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Rutala, W.A.; Kanamori, H.; Gergen, M.F.; Sickbert-Bennett, E.E.; Weber, D.J. Susceptibility of Candida auris and Candida albicans to 21 Germicides Used in Healthcare Facilities. Infect. Control Hosp. Epidemiol. 2019, 40, 380–382. [Google Scholar] [CrossRef]
- Escapa, I.F.; Chen, T.; Huang, Y.; Gajare, P.; Dewhirst, F.E.; Lemon, K.P. New Insights into Human Nostril Microbiome from the Expanded Human Oral Microbiome Database (EHOMD): A Resource for the Microbiome of the Human Aerodigestive Tract. mSystems 2018, 3, e00187-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Yu, W.H.; Izard, J.; Baranova, O.V.; Lakshmanan, A.; Dewhirst, F.E. The Human Oral Microbiome Database: A Web Accessible Resource for Investigating Oral Microbe Taxonomic and Genomic Information. Database J. Biol. Databases Curation 2010, 2010, baq013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Ranganathan, S. Bioinformatic Analysis to Investigate Metaproteome Composition Using Trans-Proteomic Pipeline. Curr. Protoc. 2022, 2, e506. [Google Scholar] [CrossRef]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Slagel, J.; Sun, Z.; Moritz, R.L. Trans-Proteomic Pipeline, a Standardized Data Processing Pipeline for Large-Scale Reproducible Proteomics Informatics. Proteom. Clin. Appl. 2015, 9, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Sievert, C. Interactive Web-Based Data Visualization with R, Plotly, and Shiny; Chapman and Hall/CRC: Boca Raton, FL, USA, 2020; ISBN 978-1-138-33145-7. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. Available online: https://cran.r-project.org/package=gplots (accessed on 1 March 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein Description | Fungal Species | Log2 Fold Change | Adjusted p-Value |
|---|---|---|---|---|
| A0A4Q4V9G8 | HET domain-containing protein | Monosporascus sp. MG133 | 1.41 | 1.17 × 10−19 |
| A0A0M8MZV6 | Tubulin beta chain | Escovopsis weberi | 1.13 | 3.94 × 10−18 |
| A0A0C9X1M1 | Unplaced genomic scaffold K443scaffold_229, whole genome shotgun sequence | Laccaria amethystina | 1.22 | 1.49 × 10−15 |
| A0A098VNG2 | Tubulin beta chain | Mitosporidium daphniae | 0.98 | 5.31 × 10−15 |
| W6MU27 | ERF-3 | Kuraishia capsulata | 1.19 | 3.16 × 10−12 |
| A0A1E3Q4C5 | Tubulin beta chain | Lipomyces starkeyi | 1.14 | 4.80 × 10−12 |
| A0A0D2A2I2 | Endoplasmic reticulum chaperone BiP | Verruconis gallopava | 1.16 | 1.66 × 10−11 |
| A0A261Y4X6 | Adenine phosphoribosyltransferase | Bifiguratus adelaidae | 1.68 | 3.66 × 10−10 |
| A0A0B2UG97 | Tubulin beta chain | Ordospora colligata | 0.95 | 5.10 × 10−10 |
| A0A4Y7TST9 | Actin-1 | Coprinellus micaceus | −1.70 | 2.45 × 10−09 |
| A0A1J5XBD0 | Actin | Amphiamblys sp. WSBS2006 | 3.00 | 2.57 × 10−09 |
| A0A512U7H9 | Uncharacterized protein | Metschnikowia sp. JCM 33374 | 2.27 | 5.62 × 10−09 |
| K1WHF7 | DNA-directed RNA polymerase subunit | Marssonina brunnea f. sp. multigermtubi | 1.17 | 1.42 × 10−08 |
| A0A4P9YSZ5 | Glyceraldehyde-3-phosphate dehydrogenase | Syncephalis pseudoplumigaleata | −0.80 | 1.42 × 10−08 |
| A0A168NSS0 | SAM_MT_RSMB_NOP domain-containing protein | Absidia glauca | 1.71 | 7.38 × 10−08 |
| A0A0A0HRG7 | Actin | Paracoccidioides brasiliensis | 1.46 | 1.25 × 10−07 |
| A0A167QHP6 | Uncharacterized protein | Phycomyces blakesleeanus | 0.70 | 8.77 × 10−07 |
| R4XE35 | DNA-directed RNA polymerase subunit | Taphrina deformans | 1.84 | 9.27 × 10−07 |
| A0A1X2H166 | Eukaryotic peptide chain release factor subunit 1 | Syncephalastrum racemosum | 1.60 | 1.32 × 10−06 |
| A0A395SGK8 | Non-ribosomal peptide synthetase | Fusarium longipes | 1.44 | 1.32 × 10−06 |
| K5X852 | WD_REPEATS_REGION domain-containing protein | Phanerochaete carnosa | 2.60 | 3.48 × 10−06 |
| M5E4N2 | Tryptophanyl-tRNA synthetase | Malassezia sympodialis | 1.64 | 3.48 × 10−06 |
| A0A1Y1WUH8 | Beta-glucosidase | Anaeromyces robustus | 1.34 | 4.96 × 10−06 |
| A0A068RIK2 | Phosphoglycerate kinase | Lichtheimia corymbifera | 1.89 | 6.27 × 10−06 |
| A0A4Q0A1Y4 | 14-3-3 domain-containing protein | Dimargaris cristalligena | 2.06 | 7.94 × 10−06 |
| A0A5M6C8D5 | ADP-ribosylation factor | Kwoniella shandongensis | 1.82 | 9.64 × 10−06 |
| A0A3N4K760 | Tubulin alpha chain | Morchella conica | 1.41 | 1.10 × 10−05 |
| A0A1Y1WHV7 | Dynein heavy chain, cytoplasmic | Linderina pennispora | 1.37 | 1.27 × 10−05 |
| A0A3M2T8I6 | Uncharacterized protein | Aspergillus sp. HF37 | 1.62 | 1.63 × 10−05 |
| R9PCI5 | Tubulin beta chain | Pseudozyma hubeiensis | 0.78 | 1.90 × 10−05 |
| Organism Type | No. of Reference Proteomes | No. of Protein Entries | Size (GB) | URL/Directory |
|---|---|---|---|---|
| Virus | 10,062 | 517,610 | 0.18 | https://ftp.uniprot.org/pub/databases/uniprot/current_release/knowledgebase/reference_proteomes/Viruses/ (release no. 2021_3; accessed on 2 June 2021) |
| Fungi | 784 | 8,370,376 | 4.61 | https://www.uniprot.org/uniprot/?query=proteome%3a(taxonomy%3a%22Fungi+%5b4751%5d%22+AND+reference%3ayes) (release no. 2021_3; accessed on 2 June 2021) |
| Bacteria | 2087 | 5,044,213 | 1.89 | http://homd.org/ftp/genomes/PROKKA/current/faa/ (ver 9.1.4: accessed on 9 September 2020) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, S.; Chakraborty, R.; Ranganathan, S. Metaproteomic Analysis of an Oral Squamous Cell Carcinoma Dataset Suggests Diagnostic Potential of the Mycobiome. Int. J. Mol. Sci. 2023, 24, 1050. https://doi.org/10.3390/ijms24021050
He S, Chakraborty R, Ranganathan S. Metaproteomic Analysis of an Oral Squamous Cell Carcinoma Dataset Suggests Diagnostic Potential of the Mycobiome. International Journal of Molecular Sciences. 2023; 24(2):1050. https://doi.org/10.3390/ijms24021050
Chicago/Turabian StyleHe, Steven, Rajdeep Chakraborty, and Shoba Ranganathan. 2023. "Metaproteomic Analysis of an Oral Squamous Cell Carcinoma Dataset Suggests Diagnostic Potential of the Mycobiome" International Journal of Molecular Sciences 24, no. 2: 1050. https://doi.org/10.3390/ijms24021050