
Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity

 , and
, and
Abstract
:1. Introduction
2. Anthracyclines—Toxicity and Mechanism of Action
3. Flavonoids—Structure, Bioavailability and Molecular Activity
3.1. General Structure of Flavonoids
3.2. Absorption and Metabolism of Flavonoids Determine In Vivo Activity
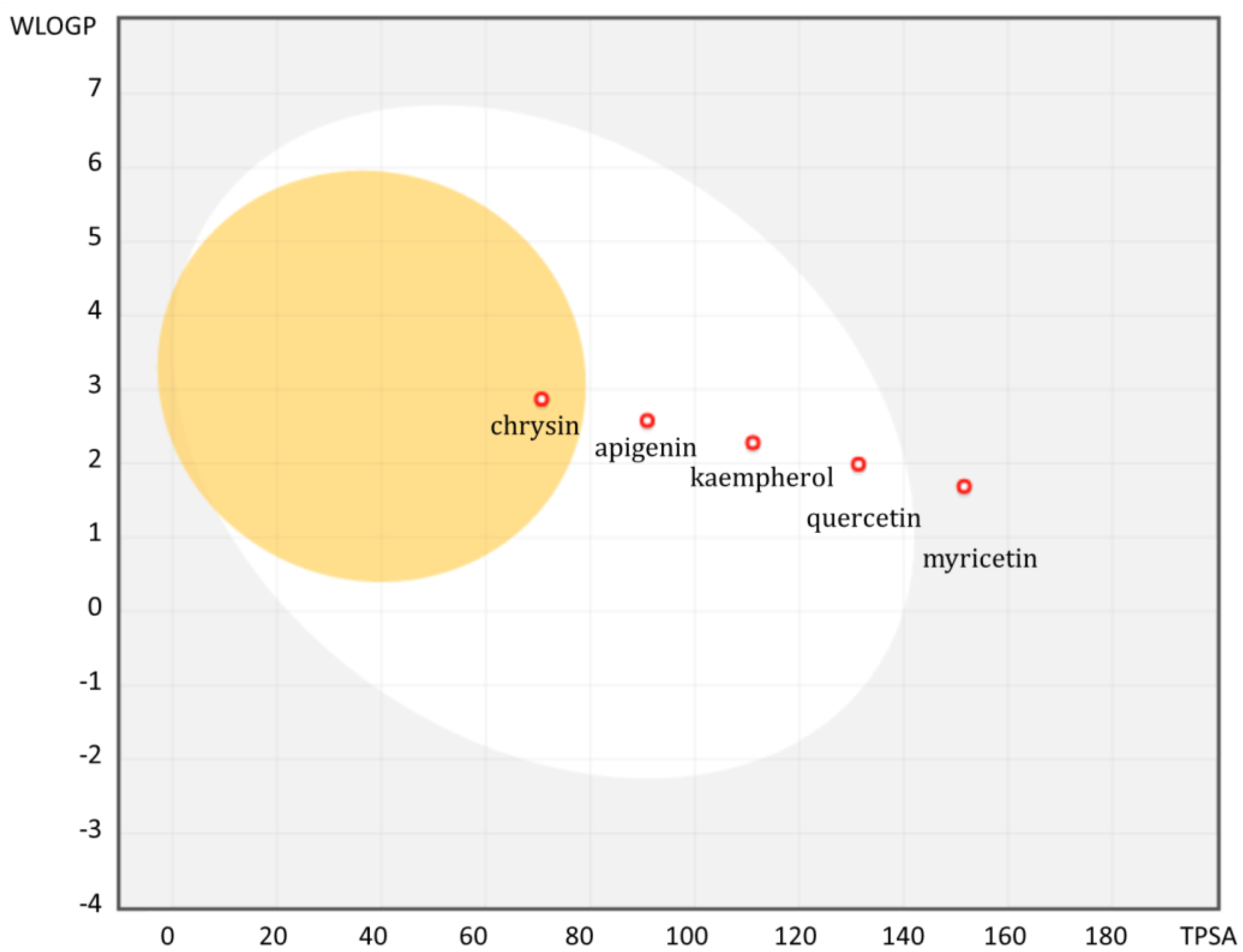
3.3. Bioavailability of Flavonoids Conditioned by Physicochemical Parameters

4. Anthracycline and Flavonoids Crosstalk
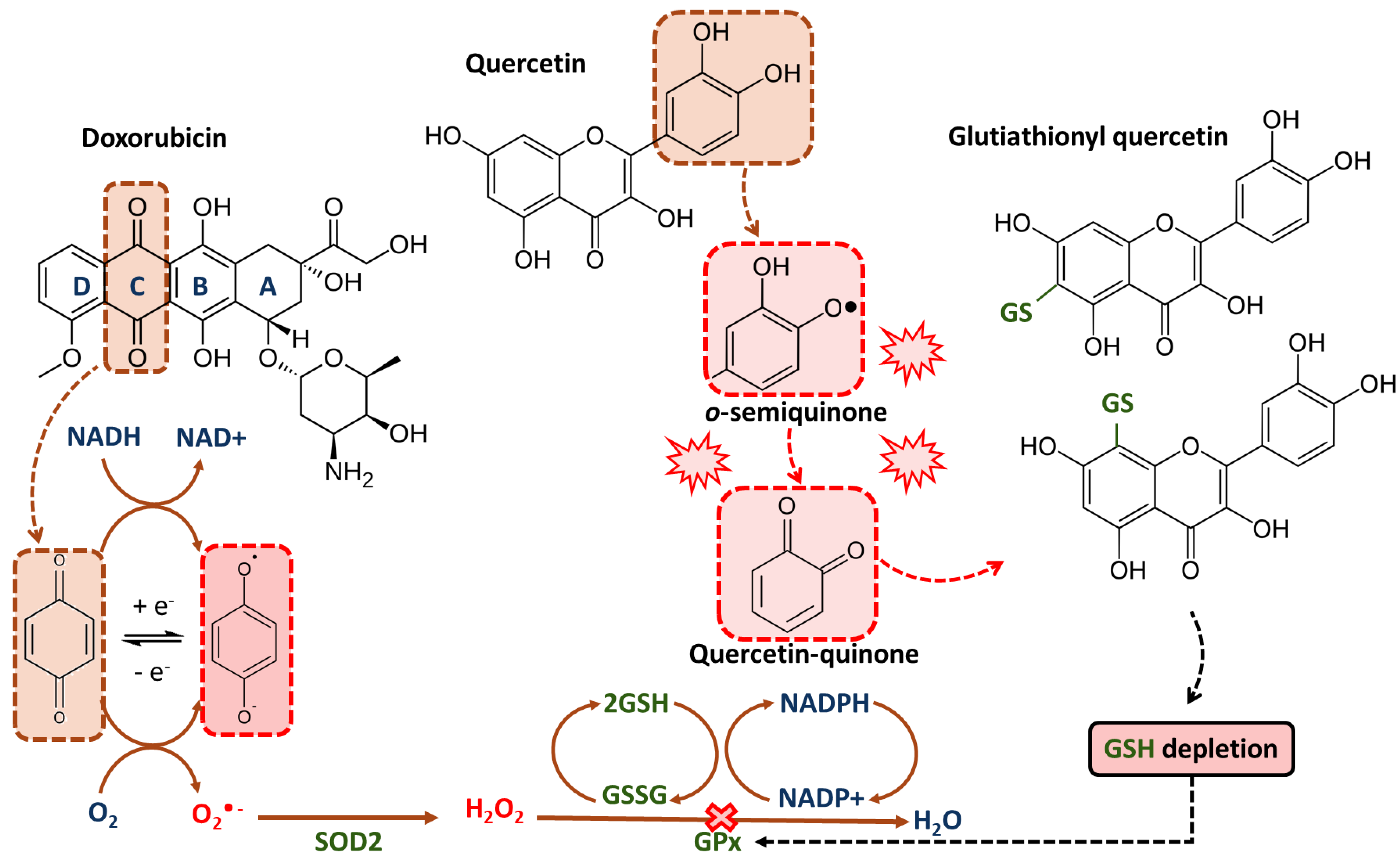
4.1. Glutathione Depletion and Oxidative Stress
4.2. Flavonoids As Inhibitors on Nrf-2/ARE Pathway
4.3. Flavonoids As Agonist and Antagonist of Nuclear Receptors
5. Modulatory Effect of Flavonoids on the Activity of Multidrug Response Proteins
5.1. Modulation Effect of Flavonoids on Cytochrome P450 3A4 (CYP3A4) Activity
5.2. Flavonoids Interactions with Organic Anion-Transporting Polypeptide (OATP)
5.3. Role of Flavonoids in ABC Transporters Activity in Cancer Cells
6. Genotoxicity vs. Genoprotection in Anthracycline-Treated Cells
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Cell Line | CHRY | API | KAE | QUE | MIR | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50 | Assay | Ref | IC50 | Assay | Ref | IC50 | Assay | Ref | IC50 | Assay | Ref | IC50 | Assay | Ref | |
| MDA-MB-468 | 5.84 ± 0.67 μM | MTT, 72 h | [235] | 8.9 μg/mL | 72 h | [237] | 23 μg/mL | 72 h MTT | [238] | 18.1 + 1.6 μM | TIA | [239] | |||
| MDA-MB-231 | 47.0 + 2.9 μM | TIA | [239] | >100 μM | MTT, 24 h | [240] | 114.75 µM | MTT, 72 h | [241] | ||||||
| 40 (35–45) μM | TBE + CTG 72 h | [242] | 75.12 ± 1.85 μM | MTT. 24 h | [243] | 34 μg/mL | MTT, T72 h | [238] | >200 μM | MTT, 24 h | [243] | 30.29 ± 1.87 μM | MTT, 24 h | [243] | |
| >200 | MTT, 24 h | [243] | 101 (82–124) μM | TBE + CTG 72 h | [242] | >200 μM | TBE + CTG 72 h | [242] | |||||||
| 38 μM | TBE + CTG 72 h | [242] | |||||||||||||
| MCF-7 | 4.20 ± 1.70 μM | MTT, 72 h | [235] | 37.2 ± 3.7 μM | WST-1, 48 h | [244] | 25 (16–40) μM | TBE + CTG 72 h | [242] | 37 μM | MTT, 24 h | [240] | >200 μM | TBE + CTG 72 h | [242] |
| 34 (28–40) μM | TBE + CTG 72 h | [242] | 7.8 μg/mL | 72 h | [237] | 102 (62–161) μM | TBE + CTG 72 h | [242] | |||||||
| 25.6 ± 1.2 µM | MTT, 72 h | [245] | >80 µM | CQ, 72 h | [246] | ||||||||||
| 13.7 ± 0.6 µM | MTT, 72 h | [245] | |||||||||||||
| HeLa | 86.51 ± 2.9 | MTT, 48 h | [247] | 10 µM | MTT, 48 h | [248] | 50 µM | MTT, 48 h | [249] | 35.5 ± 1.1 µM | MTT, 72 h | [250] | 9.06 µM | MTT, 24 h | [251] |
| 41.0 ± 4.5 | WST-1.48 h | [244] | |||||||||||||
| 74.71 ± 3.6 µg/mL | MTT, 48 h. | [252] | |||||||||||||
| HepG2 | 10.69 ± 0.94 μM | MTT, 72 h | [235] | 57.86 ± 2.9 µg/mL | MTT, 48 h | [252] | 30.92 μM | RFR, 24 h | [253] | 87 μM | TBE, 18 h | [254] | 87.84 μM | CCK-8, 72 h | [255] |
| 37.4 ± 3.8 | WST-1, 48 h | [244] | |||||||||||||
| HT-29 | 126.5 ± 6.3 | SG, 72 h | [256] | 130.6 ± 6.8 | SG, 72 h | [256] | 136.9 ± 7.4 | SG, 72 h | [256] | 85.59 ± 8.6 μM | SG, 72 h | [256] | 47.6 ± 2.3 µM | SG, 72 h | [256] |
| Caco-2 | 4.84 0.19 µM | MTT, 72 h | [235] | 115.4 ± 10.3 μM | SG 72 h | [256] | 163.2 ± 1.6 μM | SG, 72 h | [256] | 96.3 ± 6.7 μM | SG 72 h | [256] | 88.4 ± 3.4 µM. | SG, 72 h | [256] |
| 115.6 ± 2.1 μM | SG, 72 h | [256] | |||||||||||||
| HCT-116 | 4.07 ± 0.29 µM | MTT, 72 h | [235] | 34.4 ± 3.6 µM | WST-1, 48 h | [244] | 7.2 μM | WST-8, 72 h | [257] | 40 µM | MTT, 48 h | [258] | No effect | MTT, 72 h | [241] |
| LNCaP | 75 µM | MTT, 48 h | [258] | 25 µM | DC, 72 h | [259] | 53.8 μM | xC, 24 h | [260] | ||||||
| 56.81 µM | CQ, 72 h | [246] | 28:8 ± 1:5 μM | MTT, 48 h | [261] | 19.44 µM | CQ, 72 h | [246] | |||||||
| 20 µM | DC, 72 h | [259] | 33.3 μM | xC, 24 h | [260] | ||||||||||
| PC-3 | 50 µM | DC, 72 h | [259] | 52.24 µM | CQ 72 h | [246] | 14.5 μM | xC 24 h | [260] | 47.6 μM | CCK-8, 48 h | [262] | |||
| >20 μM | MTT, 48 h | [263] | 58:3 ± 3:5 μM | MTT, 48 h | [261] | 50 µM | DC, 72 h | [259] | |||||||
| >100 µM | CQ, 72 h | [246] | 38.8 ± 3.9 | WST-1, 48 h | [244] | 41.98 µM | CQ, 72 h | [246] | 33.41 µM | CQ, 72 h | [246] | ||||
| A549 | 4.64 ± 1.36 µM | MTT, 72 h | [235] | 93.7 ± 3.7 μM | MTT, 4 h | [264] | 72 μM | MTT, 24 h | [265] | 8.65 µg/mL | 24 h, MTT | [266] | 154.3 ± 4.6 | CCK-8, 48 h | [267] |
| 49.2 ± 0.6 µM | WST-8, 48 h | [268] | 35.4 ± 3.7 | WST-1, 48 h | [244] | ||||||||||
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination Therapy in Combating Cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Model Lists of Essential Medicines. Available online: https://www.who.int/groups/expert-committee-on-selection-and-use-of-essential-medicines/essential-medicines-lists (accessed on 15 November 2022).
- Sobczuk, P.; Czerwińska, M.; Kleibert, M.; Cudnoch-Jędrzejewska, A. Anthracycline-Induced Cardiotoxicity and Renin-Angiotensin-Aldosterone System—From Molecular Mechanisms to Therapeutic Applications. Heart. Fail. Rev. 2022, 27, 295–319. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, R.; Margulets, V.; Kirshenbaum, L.A. Chapter 2—Molecular Mechanisms Underlying Anthracycline Cardiotoxicity: Challenges in Cardio-Oncology. In Cardio-Oncology; Gottlieb, R.A., Mehta, P.K., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 25–34. ISBN 978-0-12-803547-4. [Google Scholar]
- El-Readi, M.Z.; Al-Abd, A.M.; Althubiti, M.A.; Almaimani, R.A.; Al-Amoodi, H.S.; Ashour, M.L.; Wink, M.; Eid, S.Y. Multiple Molecular Mechanisms to Overcome Multidrug Resistance in Cancer by Natural Secondary Metabolites. Front. Pharmacol. 2021, 12, 658513. [Google Scholar] [CrossRef]
- Liu, F.; Peng, Y.; Qiao, Y.; Huang, Y.; Song, F.; Zhang, M.; Song, F. Consumption of Flavonoids and Risk of Hormone-Related Cancers: A Systematic Review and Meta-Analysis of Observational Studies. Nutr. J. 2022, 21, 27. [Google Scholar] [CrossRef]
- Sak, K. Role of Flavonoids in the Prevention of Gynecological Cancers: Epidemiological Clues. Curr. Women’s Health Rev. 2017, 13, 103–113. [Google Scholar] [CrossRef]
- Woo, H.D.; Kim, J. Dietary Flavonoid Intake and Risk of Stomach and Colorectal Cancer. World. J. Gastroenterol. 2013, 19, 1011–1019. [Google Scholar] [CrossRef]
- Dai, R.; Liu, M.; Xiang, X.; Li, Y.; Xi, Z.; Xu, H. OMICS Applications for Medicinal Plants in Gastrointestinal Cancers: Current Advancements and Future Perspectives. Front. Pharmacol. 2022, 13, 842203. [Google Scholar] [CrossRef]
- Singh, S.; Singh, D.B.; Singh, S.; Shukla, R.; Ramteke, P.W.; Misra, K. Exploring Medicinal Plant Legacy for Drug Discovery in Post-Genomic Era. Proc. Natl. Acad. Sci. India. Sect. B Biol. Sci. 2019, 89, 1141–1151. [Google Scholar] [CrossRef]
- Lu, W.; Zhang, R.; Jiang, H.; Zhang, H.; Luo, C. Computer-Aided Drug Design in Epigenetics. Front. Chem. 2018, 6, 57. [Google Scholar] [CrossRef]
- Muroi, M.; Osada, H. Proteomics-Based Target Identification of Natural Products Affecting Cancer Metabolism. J. Antibiot. 2021, 74, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Stuart, K.A.; Welsh, K.; Walker, M.C.; Edrada-Ebel, R. Metabolomic Tools Used in Marine Natural Product Drug Discovery. Expert Opin. Drug Discov. 2020, 15, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Biharee, A.; Yadav, A.; Jangid, K.; Singh, Y.; Kulkarni, S.; Sawant, D.M.; Kumar, P.; Thareja, S.; Jain, A.K. Flavonoids as Promising Anticancer Agents: An in Silico Investigation of ADMET, Binding Affinity by Molecular Docking and Molecular Dynamics Simulations. J. Biomol. Struct. Dyn. 2022, 27, 1–12. [Google Scholar] [CrossRef]
- Boniface, P.K.; Elizabeth, F.I. Flavones as a Privileged Scaffold in Drug Discovery: Current Developments. Curr. Org. Synth 2019, 16, 968–1001. [Google Scholar] [CrossRef]
- Doytchinova, I. Drug Design-Past, Present, Future. Molecules 2022, 27, 1496. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Evolving Drug Design Methodology: From QSAR to AIDD. ChemRxiv 2022. [Google Scholar] [CrossRef]
- Selevsek, N.; Caiment, F.; Nudischer, R.; Gmuender, H.; Agarkova, I.; Atkinson, F.L.; Bachmann, I.; Baier, V.; Barel, G.; Bauer, C.; et al. Network Integration and Modelling of Dynamic Drug Responses at Multi-Omics Levels. Commun. Biol. 2020, 3, 1–15. [Google Scholar] [CrossRef]
- Grigalius, I.; Petrikaite, V. Relationship between Antioxidant and Anticancer Activity of Trihydroxyflavones. Molecules 2017, 22, 2169. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Qi, Y.; Zhao, Y.; Jin, N. Effects of Hydroxyl Group on the Interaction of Carboxylated Flavonoid Derivatives with S. cerevisiae α-Glucosidase. Curr. Comput. Aided Drug Des. 2020, 16, 31–44. [Google Scholar] [CrossRef]
- Lee, V.J.; Heffern, M.C. Structure-Activity Assessment of Flavonoids as Modulators of Copper Transport. Front. Chem. 2022, 10, 930. [Google Scholar] [CrossRef]
- Matesun, D.A.; Mensah, K.B.; Yamoah, P.; Bangalee, V.; Padayachee, N. Adverse Drug Reactions Associated with Doxorubicin and Epirubicin: A Descriptive Analysis from VigiBase. J. Oncol. Pharm. Pract. 2022, 13, 107815522211135. [Google Scholar] [CrossRef] [PubMed]
- Toroz, D.; Gould, I.R. A Computational Study of Anthracyclines Interacting with Lipid Bilayers: Correlation of Membrane Insertion Rates, Orientation Effects and Localisation with Cytotoxicity. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Kemp, C.J.; Henikoff, S. Anthracyclines Induce Double-Strand DNA Breaks at Active Gene Promoters. Mutat. Res. 2015, 773, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Marinello, J.; Delcuratolo, M.; Capranico, G. Anthracyclines as Topoisomerase II Poisons: From Early Studies to New Perspectives. Int. J. Mol. Sci. 2018, 19, 3480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulbright, J.M.; Egas-Bejar, D.E.; Huh, W.W.; Chandra, J. Analysis of Redox and Apoptotic Effects of Anthracyclines to Delineate a Cardioprotective Strategy. Cancer Chemother. Pharmacol. 2015, 76, 1297–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shandilya, M.; Sharma, S.; Das, P.P.; Charak, S.; Shandilya, M.; Sharma, S.; Das, P.P.; Charak, S. Molecular-Level Understanding of the Anticancer Action Mechanism of Anthracyclines; IntechOpen: London, UK, 2020; ISBN 978-1-83968-868-3. [Google Scholar]
- Kelly, R.A.; Leedale, J.; Calleja, D.; Enoch, S.J.; Harrell, A.; Chadwick, A.E.; Webb, S. Modelling Changes in Glutathione Homeostasis as a Function of Quinone Redox Metabolism. Sci. Rep. 2019, 9, 6333. [Google Scholar] [CrossRef] [Green Version]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin Pathways: Pharmacodynamics and Adverse Effects. Pharmacogenet. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Geisberg, C.; Sawyer, D.B. Mechanisms of Anthracycline Cardiotoxicity and Strategies to Decrease Cardiac Damage. Curr. Hypertens. Rep. 2010, 12, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Kadioglu, O.; Elbadawi, M.; Fleischer, E.; Efferth, T. Identification of Novel Anthracycline Resistance Genes and Their Inhibitors. Pharmaceuticals 2021, 14, 1051. [Google Scholar] [CrossRef]
- Cardinale, D.; Iacopo, F.; Cipolla, C.M. Cardiotoxicity of Anthracyclines. Front. Cardiovasc. Med. 2020, 7, 26. [Google Scholar] [CrossRef]
- Mechanisms of Doxorubicin-Mediated Reproductive Toxicity—A Review. Reprod. Toxicol. 2021, 102, 80–89. [CrossRef]
- van der Zanden, S.Y.; Qiao, X.; Neefjes, J. New Insights into the Activities and Toxicities of the Old Anticancer Drug Doxorubicin. FEBS J. 2021, 288, 6095–6111. [Google Scholar] [CrossRef]
- Nguyen, P.H.; Sigdel, K.P.; Kings, G.M.; Roberts, A.G. The Basis of Anthracycline Drugs Transport by P-Glycoprotein. FASEB J. 2019, 33, 507.3. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, P.; Ali, V.; Verma, M. Role of Membrane-Embedded Drug Efflux ABC Transporters in the Cancer Chemotherapy. Oncol. Rev. 2020, 14, 448. [Google Scholar] [CrossRef]
- Elmeliegy, M.; Vourvahis, M.; Guo, C.; Wang, D.D. Effect of P-Glycoprotein (P-Gp) Inducers on Exposure of P-Gp Substrates: Review of Clinical Drug–Drug Interaction Studies. Clin. Pharm. 2020, 59, 699–714. [Google Scholar] [CrossRef] [Green Version]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A Knowledge-Based Approach in Designing Combinatorial or Medicinal Chemistry Libraries for Drug Discovery. 1. A Qualitative and Quantitative Characterization of Known Drug Databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef]
- Waghray, D.; Zhang, Q. Inhibit or Evade Multidrug Resistance P-Glycoprotein in Cancer Treatment. J. Med. Chem. 2018, 61, 5108–5121. [Google Scholar] [CrossRef]
- Guttman, Y.; Kerem, Z. Computer-Aided (In Silico) Modeling of Cytochrome P450-Mediated Food–Drug Interactions (FDI). Int. J. Mol. Sci. 2022, 23, 8498. [Google Scholar] [CrossRef]
- Jabłońska-Trypuć, A.; Świderski, G.; Krętowski, R.; Lewandowski, W. Newly Synthesized Doxorubicin Complexes with Selected Metals-Synthesis, Structure and Anti-Breast Cancer Activity. Molecules 2017, 22, 1106. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, H.; Sacco, P.; Djaker, N.; de la Chapelle, M.L.; Marsich, E.; Li, X.; Spadavecchia, J. CTL–Doxorubicin (DOX)–Gold Complex Nanoparticles (DOX–AuGCs): From Synthesis to Enhancement of Therapeutic Effect on Liver Cancer Model. Nanoscale Adv. 2020, 2, 5231–5241. [Google Scholar] [CrossRef]
- Paukovcekova, S.; Krchniakova, M.; Chlapek, P.; Neradil, J.; Skoda, J.; Veselska, R. Thiosemicarbazones Can Act Synergistically with Anthracyclines to Downregulate CHEK1 Expression and Induce DNA Damage in Cell Lines Derived from Pediatric Solid Tumors. Int. J. Mol. Sci. 2022, 23, 8549. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.N.; Gradishar, W.J. Adjuvant Anthracyclines in Breast Cancer: What Is Their Role? Oncologist 2018, 23, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wu, R.; Chen, L.; Yang, Z.; Yan, D.; Li, M. Understanding Anthracycline Cardiotoxicity From Mitochondrial Aspect. Front. Pharmacol. 2022, 13, 811406. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.; Alven, S.; Maseko, R.B.; Aderibigbe, B.A. Doxorubicin-Based Hybrid Compounds as Potential Anticancer Agents: A Review. Molecules 2022, 27, 4478. [Google Scholar] [CrossRef]
- Ejiofor, I.I.; Igbokwe, M.-G.C.; Ejiofor, I.I.; Igbokwe, M.-G.C. Flavonoids: Understanding Their Biosynthetic Pathways in Plants and Health Benefits; IntechOpen: London, UK, 2021; ISBN 978-1-83969-270-3. [Google Scholar]
- Escobar-Cévoli, R.; Castro-Espín, C.; Béraud, V.; Buckland, G.; Zamora-Ros, R.; Escobar-Cévoli, R.; Castro-Espín, C.; Béraud, V.; Buckland, G.; Zamora-Ros, R. An Overview of Global Flavonoid Intake and Its Food Sources; IntechOpen: London, UK, 2017; ISBN 978-953-51-3424-4. [Google Scholar]
- Xie, L.; Deng, Z.; Zhang, J.; Dong, H.; Wang, W.; Xing, B.; Liu, X. Comparison of Flavonoid O-Glycoside, C-Glycoside and Their Aglycones on Antioxidant Capacity and Metabolism during In Vitro Digestion and In Vivo. Foods 2022, 11, 882. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in Flavonoid Research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Yue, Y.; Petimar, J.; Willett, W.C.; Smith-Warner, S.A.; Yuan, C.; Rosato, S.; Sampson, L.; Rosner, B.; Cassidy, A.; Rimm, E.B.; et al. Dietary Flavonoids and Flavonoid-Rich Foods: Validity and Reproducibility of Food Frequency Questionnaire-Derived Intake Estimates. Public Health Nutr. 2020, 23, 3295. [Google Scholar] [CrossRef]
- Egert, S.; Rimbach, G. Which Sources of Flavonoids: Complex Diets or Dietary Supplements? Adv. Nutr. 2011, 2, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Cheatham, C.L.; Nieman, D.C.; Neilson, A.P.; Lila, M.A. Enhancing the Cognitive Effects of Flavonoids With Physical Activity: Is There a Case for the Gut Microbiome? Front. Neurosci. 2022, 16, 833202. [Google Scholar] [CrossRef]
- Gouveia, H.J.C.B.; Urquiza-Martínez, M.V.; Manhães-de-Castro, R.; Costa-de-Santana, B.J.R.; Villarreal, J.P.; Mercado-Camargo, R.; Torner, L.; de Souza Aquino, J.; Toscano, A.E.; Guzmán-Quevedo, O. Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 8344. [Google Scholar] [CrossRef]
- Amawi, H.; Ashby, C.R.; Tiwari, A.K. Cancer Chemoprevention through Dietary Flavonoids: What’s Limiting? Chin. J. Cancer 2017, 36, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janabi, A.H.W.; Kamboh, A.A.; Saeed, M.; Xiaoyu, L.; BiBi, J.; Majeed, F.; Naveed, M.; Mughal, M.J.; Korejo, N.A.; Kamboh, R.; et al. Flavonoid-Rich Foods (FRF): A Promising Nutraceutical Approach against Lifespan-Shortening Diseases. Iran. J. Basic Med. Sci. 2020, 23, 140. [Google Scholar] [CrossRef]
- Šudomová, M.; Berchová-Bímová, K.; Mazurakova, A.; Šamec, D.; Kubatka, P.; Hassan, S.T.S. Flavonoids Target Human Herpesviruses That Infect the Nervous System: Mechanisms of Action and Therapeutic Insights. Viruses 2022, 14, 592. [Google Scholar] [CrossRef] [PubMed]
- Husain, I.; Bala, K.; Khan, I.A.; Khan, S.I. A Review on Phytochemicals, Pharmacological Activities, Drug Interactions, and Associated Toxicities of Licorice (Glycyrrhiza sp.). Food Front. 2021, 2, 449–485. [Google Scholar] [CrossRef]
- Singh, P.; Sharma, S.; Rath, S.K. A Versatile Flavonoid Quercetin: Study of Its Toxicity and Differential Gene Expression in the Liver of Mice. Phytomed. Plus 2022, 2, 100148. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Azimi-Nezhad, M. Protective Effects of Chrysin Against Drugs and Toxic Agents. Dose Response 2017, 15, 1559325817711782. [Google Scholar] [CrossRef]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin Bioactive Effects: Moving from Preclinical Evidence to Potential Clinical Applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef]
- Lewandowski, W.; Lewandowska, H.; Golonko, A.; Świderski, G.; Świsłocka, R.; Kalinowska, M. Correlations between Molecular Structure and Biological Activity in “Logical Series” of Dietary Chromone Derivatives. PLoS ONE 2020, 15, e0229477. [Google Scholar] [CrossRef]
- Aparicio, S. A Systematic Computational Study on Flavonoids. Int. J. Mol. Sci. 2010, 11, 2017–2038. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, M. Current Trends in Computational Quantum Chemistry Studies on Antioxidant Radical Scavenging Activity. J. Chem. Inf. Model. 2022, 62, 2639–2658. [Google Scholar] [CrossRef]
- Zheng, Y.-Z.; Deng, G.; Guo, R.; Fu, Z.-M.; Chen, D.-F. Theoretical Insight into the Antioxidative Activity of Isoflavonoid: The Effect of the C2=C3 Double Bond. Phytochemistry 2019, 166, 112075. [Google Scholar] [CrossRef]
- Nam, G.; Hong, M.; Lee, J.; Lee, H.J.; Ji, Y.; Kang, J.; Baik, M.-H.; Lim, M.H. Multiple Reactivities of Flavonoids towards Pathological Elements in Alzheimer’s Disease: Structure–Activity Relationship. Chem. Sci. 2020, 11, 10243–10254. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of Angiotensin-Converting Enzyme Activity by Flavonoids: Structure-Activity Relationship Studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veiko, A.G.; Lapshina, E.A.; Zavodnik, I.B. Comparative Analysis of Molecular Properties and Reactions with Oxidants for Quercetin, Catechin, and Naringenin. Mol. Cell. Biochem. 2021, 476, 4287–4299. [Google Scholar] [CrossRef] [PubMed]
- Shafreen, R.M.B.; Lakshmi, S.A.; Pandian, S.K.; Park, Y.S.; Kim, Y.M.; Paśko, P.; Deutsch, J.; Katrich, E.; Gorinstein, S. Unraveling the Antioxidant, Binding and Health-Protecting Properties of Phenolic Compounds of Beers with Main Human Serum Proteins: In Vitro and In Silico Approaches. Molecules 2020, 25, 4962. [Google Scholar] [CrossRef] [PubMed]
- Moazzen, A.; Öztinen, N.; Ak-Sakalli, E.; Koşar, M. Structure-Antiradical Activity Relationships of 25 Natural Antioxidant Phenolic Compounds from Different Classes. Heliyon 2022, 8, e10467. [Google Scholar] [CrossRef]
- Furger, C. Live Cell Assays for the Assessment of Antioxidant Activities of Plant Extracts. Antioxidants 2021, 10, 944. [Google Scholar] [CrossRef]
- Isika, D.K.; Özkömeç, F.N.; Çeşme, M.; Sadik, O.A. Synthesis, Biological and Computational Studies of Flavonoid Acetamide Derivatives. RSC Adv. 2022, 12, 10037–10050. [Google Scholar] [CrossRef]
- Grootaert, C.; Gonzales, G.B.; Vissenaekens, H.; Van De Wiele, T.; Raes, K.; Smagghe, G.; Van Camp, J. Flow Cytometric Method for the Detection of Flavonoids in Cell Lines. J. Biomol. Screen. 2016, 21, 858–865. [Google Scholar] [CrossRef] [Green Version]
- Vissenaekens, H.; Smagghe, G.; Criel, H.; Grootaert, C.; Raes, K.; Rajkovic, A.; Goeminne, G.; Boon, N.; De Schutter, K.; Van Camp, J. Intracellular Quercetin Accumulation and Its Impact on Mitochondrial Dysfunction in Intestinal Caco-2 Cells. Food Res. Int. 2021, 145, 110430. [Google Scholar] [CrossRef]
- Forbes, A.M.; Lin, H.; Meadows, G.G.; Meier, G.P. Synthesis and Anticancer Activity of New Flavonoid Analogs and Inconsistencies in Assays Related to Proliferation and Viability Measurements. Int. J. Oncol. 2014, 45, 831–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, L.A.; Da Silva, H.C.; De Almeida, W.B. Structural Determination of Antioxidant and Anticancer Flavonoid Rutin in Solution through DFT Calculations of 1H NMR Chemical Shifts. ChemistryOpen 2018, 7, 902–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, B.T.; Thompson, E.P. A Method for Visualizing Fluorescence of Flavonoid Therapeutics in Vivo in the Model Eukaryote Dictyostelium Discoideum. BioTechniques 2019, 66, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheidt, H.A.; Pampel, A.; Nissler, L.; Gebhardt, R.; Huster, D. Investigation of the Membrane Localization and Distribution of Flavonoids by High-Resolution Magic Angle Spinning NMR Spectroscopy. Biochim. Biophys. Acta (BBA) Biomembr. 2004, 1663, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Ulrih, N.P.; Maričić, M.; Ota, A.; Šentjurc, M.; Abram, V. Kaempferol and Quercetin Interactions with Model Lipid Membranes. Food Res. Int. 2015, 71, 146–154. [Google Scholar] [CrossRef]
- Chodari, L.; Aytemir, M.D.; Vahedi, P.; Alipour, M.; Vahed, S.Z.; Khatibi, S.M.H.; Ahmadian, E.; Ardalan, M.; Eftekhari, A. Targeting Mitochondrial Biogenesis with Polyphenol Compounds. Oxidative Med. Cell. Longev. 2021, 2021, 4946711. [Google Scholar] [CrossRef]
- Zheng, Y.; Ma, L.; Sun, Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front. Pharmacol. 2021, 12, 648407. [Google Scholar]
- Walle, T.; Otake, Y.; Brubaker, J.A.; Walle, U.K.; Halushka, P.V. Disposition and Metabolism of the Flavonoid Chrysin in Normal Volunteers. Br. J. Clin. Pharmacol. 2001, 51, 143–146. [Google Scholar] [CrossRef]
- Gugler, R.; Leschik, M.; Dengler, H.J. Disposition of Quercetin in Man after Single Oral and Intravenous Doses. Eur. J. Clin. Pharmacol. 1975, 9, 229–234. [Google Scholar] [CrossRef]
- Riva, A.; Ronchi, M.; Petrangolini, G.; Bosisio, S.; Allegrini, P. Improved Oral Absorption of Quercetin from Quercetin Phytosome®, a New Delivery System Based on Food Grade Lecithin. Eur. J. Drug Metab. Pharmacokinet. 2019, 44, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Chavez, E.; Garcia-Quiroz, A.; Santiago-Jiménez, J.C.; Díaz-Góngora, J.A.I.; Díaz-López, R.; de Landa Castillo-Alvarado, F. Quantum–Mechanical Characterization of the Doxorubicin Molecule to Improve Its Anticancer Functions. MRS Adv. 2021, 6, 897–902. [Google Scholar] [CrossRef]
- Tosca, E.M.; Bartolucci, R.; Magni, P. Application of Artificial Neural Networks to Predict the Intrinsic Solubility of Drug-Like Molecules. Pharmaceutics 2021, 13, 1101. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Khan, F. 3D-QSAR, Docking, ADME/Tox Studies on Flavone Analogs Reveal Anticancer Activity through Tankyrase Inhibition. Sci. Rep. 2019, 9, 5414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žuvela, P.; David, J.; Wong, M.W. Interpretation of ANN-Based QSAR Models for Prediction of Antioxidant Activity of Flavonoids. J. Comput. Chem. 2018, 39, 953–963. [Google Scholar] [CrossRef]
- Zhang, X.; Lionberger, R.A.; Davit, B.M.; Yu, L.X. Utility of Physiologically Based Absorption Modeling in Implementing Quality by Design in Drug Development. AAPS J. 2011, 13, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast Calculation of Molecular Polar Surface Area as a Sum of Fragment-Based Contributions and Its Application to the Prediction of Drug Transport Properties. J. Med. Chem. 2000, 43, 3714–3717. [Google Scholar] [CrossRef]
- Bitew, M.; Desalegn, T.; Demissie, T.B.; Belayneh, A.; Endale, M.; Eswaramoorthy, R. Pharmacokinetics and Drug-Likeness of Antidiabetic Flavonoids: Molecular Docking and DFT Study. PLoS ONE 2021, 16, e0260853. [Google Scholar] [CrossRef]
- Schnider, P. CHAPTER 1: Overview of Strategies for Solving ADMET Challenges. In The Medicinal Chemist’s Guide to Solving ADMET Challenges; Royal Society of Chemistry: London, UK, 2021; pp. 1–15. [Google Scholar]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. Prediction of Hydrophobic (Lipophilic) Properties of Small Organic Molecules Using Fragmental Methods: An Analysis of ALOGP and CLOGP Methods. J. Phys. Chem. A 1998, 102, 3762–3772. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg to Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Prachayasittikul, V.; Worachartcheewan, A.; Shoombuatong, W.; Prachayasittikul, V.; Nantasenamat, C. Classification of P-Glycoprotein-Interacting Compounds Using Machine Learning Methods. EXCLI J. 2015, 14, 958. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.B.; de Oliveira, P.V.; Fagan, S.B.; Oliveira, B.R.; da Silva Nornberg, B.F.; Almeida, D.V.; Marins, L.F.; González-Durruthy, M. Modeling Drug-Drug Interactions of AZD1208 with Vincristine and Daunorubicin on Ligand-Extrusion Binding TMD-Domains of Multidrug Resistance P-Glycoprotein (ABCB1). Toxicology 2019, 411, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Fang, Y.; Cao, W.; Liang, F.; Pan, S.; Xu, X. Quantitative Structure–Activity Relationships for the Flavonoid-Mediated Inhibition of P-Glycoprotein in KB/MDR1 Cells. Molecules 2019, 24, 1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, F.; Bahlake, A.; Chamani, Z.; Bagheri, S. Theoretical Study on the Conjugation of PLGA and PLGA-PEG Carriers to Doxorubicin and Daunorubicin. Eur. J. Exp. Biol. 2012, 2, 2055–2060. [Google Scholar]
- Beck, T.C.; Beck, K.R.; Morningstar, J.; Benjamin, M.M.; Norris, R.A. Descriptors of Cytochrome Inhibitors and Useful Machine Learning Based Methods for the Design of Safer Drugs. Pharmaceuticals 2021, 14, 472. [Google Scholar] [CrossRef]
- Miar, M.; Shiroudi, A.; Pourshamsian, K.; Oliaey, A.R.; Hatamjafari, F. Theoretical Investigations on the HOMO–LUMO Gap and Global Reactivity Descriptor Studies, Natural Bond Orbital, and Nucleus-Independent Chemical Shifts Analyses of 3-Phenylbenzo[d]Thiazole-2(3H)-Imine and Its Para-Substituted Derivatives: Solvent and Substituent Effects. J. Chem. Res. 2021, 45, 147–158. [Google Scholar] [CrossRef]
- Lagoa, R.; Graziani, I.; Lopez-Sanchez, C.; Garcia-Martinez, V.; Gutierrez-Merino, C. Complex I and Cytochrome c Are Molecular Targets of Flavonoids That Inhibit Hydrogen Peroxide Production by Mitochondria. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1807, 1562–1572. [Google Scholar] [CrossRef] [Green Version]
- Jomova, K.; Hudecova, L.; Lauro, P.; Simunková, M.; Barbierikova, Z.; Malcek, M.; Alwasel, S.H.; Alhazza, I.M.; Rhodes, C.J.; Valko, M. The Effect of Luteolin on DNA Damage Mediated by a Copper Catalyzed Fenton Reaction. J. Inorg. Biochem. 2022, 226, 111635. [Google Scholar] [CrossRef]
- Vásquez-Espinal, A.; Yañez, O.; Osorio, E.; Areche, C.; García-Beltrán, O.; Ruiz, L.M.; Cassels, B.K.; Tiznado, W. Theoretical Study of the Antioxidant Activity of Quercetin Oxidation Products. Front. Chem. 2019, 7, 818. [Google Scholar] [CrossRef]
- Spiegel, M.; Andruniów, T.; Sroka, Z. Flavones’ and Flavonols’ Antiradical Structure–Activity Relationship—A Quantum Chemical Study. Antioxidants 2020, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-Y.; Hsieh, Y.-A.; Tsai, C.-I.; Kang, Y.-F.; Chang, F.-R.; Wu, Y.-C.; Wu, C.-C. Protoapigenone, a Natural Derivative of Apigenin, Induces Mitogen-Activated Protein Kinase-Dependent Apoptosis in Human Breast Cancer Cells Associated with Induction of Oxidative Stress and Inhibition of Glutathione S-Transferase π. Invest. New Drugs 2011, 29, 1347–1359. [Google Scholar] [CrossRef]
- Santini, C.; Pellei, M.; Gandin, V.; Porchia, M.; Tisato, F.; Marzano, C. Advances in Copper Complexes as Anticancer Agents. Chem. Rev. 2014, 114, 815–862. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Rizvi, A. Understanding the Prooxidant Action of Plant Polyphenols in the Cellular Microenvironment of Malignant Cells: Role of Copper and Therapeutic Implications. Front. Pharmacol. 2022, 13, 929853. [Google Scholar] [CrossRef]
- He, H.; Wang, L.; Qiao, Y.; Zhou, Q.; Li, H.; Chen, S.; Yin, D.; Huang, Q.; He, M. Doxorubicin Induces Endotheliotoxicity and Mitochondrial Dysfunction via ROS/ENOS/NO Pathway. Front. Pharmacol. 2020, 10, 1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. CPB 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative Stress and Cellular Response to Doxorubicin: A Common Factor in the Complex Milieu of Anthracycline Cardiotoxicity. Oxid. Med. Cell. Longev. 2017, 2017, 1521020. [Google Scholar] [CrossRef]
- Carrasco-Torres, G.; Baltiérrez-Hoyos, R.; Andrade-Jorge, E.; Villa-Treviño, S.; Trujillo-Ferrara, J.G.; Vásquez-Garzón, V.R. Cytotoxicity, Oxidative Stress, Cell Cycle Arrest, and Mitochondrial Apoptosis after Combined Treatment of Hepatocarcinoma Cells with Maleic Anhydride Derivatives and Quercetin. Oxidative Med. Cell. Longev. 2017, 2017, e2734976. [Google Scholar] [CrossRef]
- Iqbal, M. Flavonoid-Mediated Modulation of CYP3A Enzyme and P-Glycoprotein Transporter: Potential Effects on Bioavailability and Disposition of Tyrosine Kinase Inhibitors; IntechOpen: London, UK, 2020; ISBN 978-1-83880-888-4. [Google Scholar]
- Ju, J.; Picinich, S.C.; Yang, Z.; Zhao, Y.; Suh, N.; Kong, A.-N.; Yang, C.S. Cancer-Preventive Activities of Tocopherols and Tocotrienols. Carcinogenesis 2010, 31, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.U.; Dagur, H.S.; Khan, M.; Malik, N.; Alam, M.; Mushtaque, M. Therapeutic Role of Flavonoids and Flavones in Cancer Prevention: Current Trends and Future Perspectives. Eur. J. Med. Chem. Rep. 2021, 3, 100010. [Google Scholar] [CrossRef]
- Bidoli, E.; Talamini, R.; Zucchetto, A.; Bosetti, C.; Negri, E.; Lenardon, O.; Maso, L.D.; Polesel, J.; Montella, M.; Franceschi, S.; et al. Dietary Vitamins E and C and Prostate Cancer Risk. Acta Oncol. 2009, 48, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Zhang, V.X.; Sze, K.M.-F.; Chan, L.-K.; Ho, D.W.-H.; Tsui, Y.-M.; Chiu, Y.-T.; Lee, E.; Husain, A.; Huang, H.; Tian, L.; et al. Antioxidant Supplements Promote Tumor Formation and Growth and Confer Drug Resistance in Hepatocellular Carcinoma by Reducing Intracellular ROS and Induction of TMBIM1. Cell. Biosci. 2021, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Jardim, B.V.; Moschetta, M.G.; Leonel, C.; Gelaleti, G.B.; Regiani, V.R.; Ferreira, L.C.; Lopes, J.R.; de Campos Zuccari, D.A.P. Glutathione and Glutathione Peroxidase Expression in Breast Cancer: An Immunohistochemical and Molecular Study. Oncol. Rep. 2013, 30, 1119–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criscuolo, D.; Avolio, R.; Parri, M.; Romano, S.; Chiarugi, P.; Matassa, D.S.; Esposito, F. Decreased Levels of GSH Are Associated with Platinum Resistance in High-Grade Serous Ovarian Cancer. Antioxidants 2022, 11, 1544. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.L.; Heerdt, P.; Sachar, S.; Abolhoda, A.; Hochwald, S.; Cheng, H.; Burt, M. Glutathione Metabolism in Patients with Non-Small Cell Lung Cancers. Cancer Res. 1997, 57, 152–155. [Google Scholar] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Kachadourian, R.; Day, B.J. Flavonoid-Induced Glutathione Depletion: Potential Implications for Cancer Treatment. Free Radic. Biol. Med. 2006, 41, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Mahbub, A.A.; Le Maitre, C.L.; Haywood-Small, S.L.; Cross, N.A.; Jordan-Mahy, N. Polyphenols Act Synergistically with Doxorubicin and Etoposide in Leukaemia Cell Lines. Cell Death Discov. 2015, 1, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Chen, C.; Xu, Y.; Shen, J.; Guo, H.; Li, H.; Li, X.; Kang, D.; Shao, Y.; Zhu, Z.; et al. Is the Combinational Administration of Doxorubicin and Glutathione a Reasonable Proposal? Acta Pharmacol. Sin. 2019, 40, 699–709. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, K.; Zong, Q.; Tu, Y.; Dong, Y.; Yuan, Y. Polyprodrug with Glutathione Depletion and Cascade Drug Activation for Multi-Drug Resistance Reversal. Biomaterials 2021, 270, 120649. [Google Scholar] [CrossRef]
- Ferraresi, R.; Troiano, L.; Roat, E.; Lugli, E.; Nemes, E.; Nasi, M.; Pinti, M.; Garcia Fernandez, M.I.; Copper, E.L.; Cossarizza, A. Essential Requirement of Reduced Glutathione (GSH) for the Anti-Oxidant Effect of the Flavonoid Quercetin. Free Radic. Res. 2005, 39, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Robaszkiewicz, A.; Balcerczyk, A.; Bartosz, G. Antioxidative and Prooxidative Effects of Quercetin on A549 Cells. Cell Biol. Int. 2007, 31, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L. Quercetin and Cancer Chemoprevention. Evid.-Based Complement. Altern. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef] [Green Version]
- Brechbuhl, H.M.; Kachadourian, R.; Min, E.; Chan, D.; Day, B.J. Chrysin Enhances Doxorubicin-Induced Cytotoxicity in Human Lung Epithelial Cancer Cell Lines: The Role of Glutathione. Toxicol. Appl. Pharmacol. 2012, 258, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Poku, V.O.; Iram, S.H. A Critical Review on Modulators of Multidrug Resistance Protein 1 in Cancer Cells. PeerJ 2022, 10, e12594. [Google Scholar] [CrossRef]
- Li, Y.; Revalde, J.L.; Reid, G.; Paxton, J.W. Interactions of Dietary Phytochemicals with ABC Transporters: Possible Implications for Drug Disposition and Multidrug Resistance in Cancer. Drug Metab. Rev. 2010, 42, 590–611. [Google Scholar] [CrossRef]
- Cebula, M.; Schmidt, E.E.; Arnér, E.S.J. TrxR1 as a Potent Regulator of the Nrf2-Keap1 Response System. Antioxid. Redox. Signal 2015, 23, 823–853. [Google Scholar] [CrossRef] [Green Version]
- Shan, S.; Shi, J.; Yang, P.; Jia, B.; Wu, H.; Zhang, X.; Li, Z. Apigenin Restrains Colon Cancer Cell Proliferation via Targeted Blocking of Pyruvate Kinase M2-Dependent Glycolysis. J. Agric. Food Chem. 2017, 65, 8136–8144. [Google Scholar] [CrossRef]
- Durmus, S.; Naik, J.; Buil, L.; Wagenaar, E.; Van Tellingen, O.; Schinkel, A.H. In Vivo Disposition of Doxorubicin Is Affected by Mouse Oatp1a/1b and Human OATP1A/1B Transporters. Int. J. Cancer 2014, 135, 1700–1710. [Google Scholar] [CrossRef]
- Gao, A.M.; Ke, Z.P.; Shi, F.; Sun, G.C.; Chen, H. Chrysin Enhances Sensitivity of BEL-7402/ADM Cells to Doxorubicin by Suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 Pathway. Chem.-Biol. Interact. 2013, 206, 100–108. [Google Scholar] [CrossRef]
- Gao, A.-M.; Ke, Z.-P.; Wang, J.-N.; Yang, J.-Y.; Chen, S.-Y.; Chen, H. Apigenin Sensitizes Doxorubicin-Resistant Hepatocellular Carcinoma BEL-7402/ADM Cells to Doxorubicin via Inhibiting PI3K/Akt/Nrf2 Pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi-Pour, Z.; Ramezani, F.; Keshavarzi, F.; Samadi, N. The Role of Quercetin and Vitamin C in Nrf2-dependent Oxidative Stress Production in Breast Cancer Cells. Oncol. Lett. 2017, 13, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Majumder, D.; Saha, C. Correlation of Binding Efficacies of DNA to Flavonoids and Their Induced Cellular Damage. J. Photochem. Photobiol. B Biol. 2017, 170, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Sabzichi, M.; Mohammadian, J.; Bazzaz, R.; Pirouzpanah, M.B.; Shaaker, M.; Hamishehkar, H.; Chavoshi, H.; Salehi, R.; Samadi, N. Chrysin Loaded Nanostructured Lipid Carriers (NLCs) Triggers Apoptosis in MCF-7 Cancer Cells by Inhibiting the Nrf2 Pathway. Process Biochem. 2017, 60, 84–91. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Sun, K.; Wang, X.; Pan, H.; Zhu, J.; Ji, X.; Li, X. Chrysin Suppresses Proliferation, Migration, and Invasion in Glioblastoma Cell Lines via Mediating the ERK/Nrf2 Signaling Pathway. Drug Des. Dev. Ther. 2018, 12, 721. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.M.; Zhang, X.Y.; Ke, Z.P. Apigenin Sensitizes BEL-7402/ADM Cells to Doxorubicin through Inhibiting MiR-101/Nrf2 Pathway. Oncotarget 2017, 8, 82085. [Google Scholar] [CrossRef] [Green Version]
- Fouzder, C.; Mukhuty, A.; Kundu, R. Kaempferol Inhibits Nrf2 Signalling Pathway via Downregulation of Nrf2 MRNA and Induces Apoptosis in NSCLC Cells. Arch. Biochem. Biophys. 2021, 697, 108700. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Qu, C.; Chen, L.; Geng, Y.; Cheng, C.; Yu, S.; Wang, D.; Yang, L.; Meng, Z.; et al. Kaempferol Induces ROS-Dependent Apoptosis in Pancreatic Cancer Cells via TGM2-Mediated Akt/MTOR Signaling. BMC Cancer 2021, 21, 1–11. [Google Scholar] [CrossRef]
- De Prax, M.C.A.; Ferro, K.P.V.; Santos, I.; Torello, C.O.; Salazar-Terreros, M.; Olalla Saad, S.T. NRF2 Is Targeted By the Polyphenol Quercetin and Induces Apoptosis, in Part, through up Regulation of Pro Apoptotic Mirs. Blood 2019, 134, 2529. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Guo, Y.; Yang, A.Y.; Paredes-Gonzalez, X.; Ramirez, C.; Pung, D.; Kong, A.N.T. The Berry Constituents Quercetin, Kaempferol, and Pterostilbene Synergistically Attenuate Reactive Oxygen Species: Involvement of the Nrf2-ARE Signaling Pathway. Food Chem. Toxicol. 2014, 72, 303–311. [Google Scholar] [CrossRef]
- Sharma, A.; Parikh, M.; Shah, H.; Gandhi, T. Modulation of Nrf2 by Quercetin in Doxorubicin-Treated Rats. Heliyon 2020, 6, e03803. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Chen, T.; Zhang, Z.; Chen, X.; Chen, C.; Chen, L.; Wang, X.; Ying, X. Activation of Nrf2/HO-1 Signal with Myricetin for Attenuating ECM Degradation in Human Chondrocytes and Ameliorating the Murine Osteoarthritis. Int. Immunopharmacol. 2019, 75, 105742. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, A.Y.; Ye, X.; Li, B.; Rojanasakul, Y.; Rankin, G.O.; Chen, Y.C. Myricetin Inhibits Proliferation of Cisplatin-Resistant Cancer Cells through a P53-Dependent Apoptotic Pathway. Int. J. Oncol. 2015, 47, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.-C.; Hsiao, J.-R.; Jiang, S.-S.; Chang, J.-Y.; Chu, P.-Y.; Liu, K.-J.; Fang, H.-L.; Lin, L.-M.; Chen, H.-H.; Huang, Y.-W.; et al. C-MYC-Directed NRF2 Drives Malignant Progression of Head and Neck Cancer via Glucose-6-Phosphate Dehydrogenase and Transketolase Activation. Theranostics 2021, 11, 5232–5247. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Chen, J.; Tanigawa, S.; Hou, D.-X. Microarray and Pathway Analysis Highlight Nrf2/ARE-Mediated Expression Profiling by Polyphenolic Myricetin. Mol. Nutr. Food Res. 2013, 57, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Avior, Y.; Bomze, D.; Ramon, O.; Nahmias, Y. Flavonoids as Dietary Regulators of Nuclear Receptor Activity. Food Funct. 2013, 4, 831–844. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.F.A.; Van Winkle, L.S.; Esser, C.; Haarmann-Stemmann, T. The Aryl Hydrocarbon Receptor as a Target of Environmental Stressors—Implications for Pollution Mediated Stress and Inflammatory Responses. Redox. Biol. 2020, 34, 101530. [Google Scholar] [CrossRef]
- Husain, I.; Dale, O.R.; Martin, K.; Gurley, B.J.; Adams, S.J.; Avula, B.; Chittiboyina, A.G.; Khan, I.A.; Khan, S.I. Screening of Medicinal Plants for Possible Herb-Drug Interactions through Modulating Nuclear Receptors, Drug-Metabolizing Enzymes and Transporters. J. Ethnopharmacol. 2023, 301, 115822. [Google Scholar] [CrossRef]
- Goya-Jorge, E.; Jorge Rodríguez, M.E.; Veitía, M.S.-I.; Giner, R.M. Plant Occurring Flavonoids as Modulators of the Aryl Hydrocarbon Receptor. Molecules 2021, 26, 2315. [Google Scholar] [CrossRef]
- Sandanaraj, E.; Lal, S.; Selvarajan, V.; Ooi, L.L.; Wong, Z.W.; Wong, N.S.; Ang, P.C.S.; Lee, E.J.D.; Chowbay, B. PXR Pharmacogenetics: Association of Haplotypes with Hepatic CYP3A4 and ABCB1 Messenger RNA Expression and Doxorubicin Clearance in Asian Breast Cancer Patients. Clin. Cancer Res. 2008, 14, 7116–7126. [Google Scholar] [CrossRef] [Green Version]
- Jin, U.-H.; Park, H.; Li, X.; Davidson, L.A.; Allred, C.; Patil, B.; Jayaprakasha, G.; Orr, A.A.; Mao, L.; Chapkin, R.S.; et al. Structure-Dependent Modulation of Aryl Hydrocarbon Receptor-Mediated Activities by Flavonoids. Toxicol. Sci. 2018, 164, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carazo Fernández, A.; Smutny, T.; Hyrsová, L.; Berka, K.; Pavek, P. Chrysin, Baicalein and Galangin Are Indirect Activators of the Human Constitutive Androstane Receptor (CAR). Toxicol. Lett. 2015, 233, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Lin, W.; Wu, J.; Chen, T. Flavonoids Activate Pregnane x Receptor-Mediated CYP3A4 Gene Expression by Inhibiting Cyclin-Dependent Kinases in HepG2 Liver Carcinoma Cells. BMC Biochem. 2010, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Li, D.; Yu, W.; Zhang, Q.; Hou, X.; He, Y.; Kou, X. Mechanisms and Therapeutic Prospects of Polyphenols as Modulators of the Aryl Hydrocarbon Receptor. Food Funct. 2017, 8, 1414–1437. [Google Scholar] [CrossRef]
- Rajaraman, G.; Yang, G.; Chen, J.; Chang, T.K. Modulation of CYP1B1 and CYP1A1 Gene Expression and Activation of Aryl Hydrocarbon Receptor by Ginkgo Biloba Extract in MCF-10A Human Mammary Epithelial Cells. Can. J. Physiol. Pharmacol. 2009, 87, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, B.; Wang, H. Mechanisms of Xenobiotic Receptor Activation: Direct vs. Indirect. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2016, 1859, 1130–1140. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.; Sakamuru, S.; Huang, R.; Niebler, J.; Ferguson, S.S.; Xia, M. Characterization of Human Pregnane X Receptor Activators Identified from a Screening of the Tox21 Compound Library. Biochem. Pharmacol. 2021, 184, 114368. [Google Scholar] [CrossRef]
- Hakkola, J.; Hukkanen, J.; Turpeinen, M.; Pelkonen, O. Inhibition and Induction of CYP Enzymes in Humans: An Update. Arch. Toxicol. 2020, 94, 3671–3722. [Google Scholar] [CrossRef]
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28, 1359–1390. [Google Scholar] [CrossRef]
- Basheer, L.; Kerem, Z. Interactions between CYP3A4 and Dietary Polyphenols. Oxid. Med. Cell Longev. 2015, 2015, 854015. [Google Scholar] [CrossRef] [Green Version]
- Harahap, Y.; Ardiningsih, P.; Winarti, A.C.; Purwanto, D.J. Analysis of the Doxorubicin and Doxorubicinol in the Plasma of Breast Cancer Patients for Monitoring the Toxicity of Doxorubicin. Drug Des. Dev. Ther. 2020, 14, 3469. [Google Scholar] [CrossRef] [PubMed]
- Schulte, R.R.; Ho, R.H. Organic Anion Transporting Polypeptides: Emerging Roles in Cancer Pharmacology. Mol. Pharmacol. 2019, 95, 490. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Leake, B.F.; Kim, R.B.; Ho, R.H. Contribution of Organic Anion-Transporting Polypeptides 1A/1B to Doxorubicin Uptake and Clearance. Mol. Pharmacol. 2017, 91, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, T.; Tamai, I. Genetic polymorphisms of OATP transporters and their impact on intestinal absorption and hepatic disposition of drugs. Drug Metab. Pharmacokinet. 2012, 27, 106–121. [Google Scholar] [CrossRef]
- Damodar, G.; Smitha, T.; Gopinath, S.; Vijayakumar, S.; Rao, Y. An Evaluation of Hepatotoxicity in Breast Cancer Patients Receiving Injection Doxorubicin. Ann. Med. Health. Sci. Res. 2014, 4, 74. [Google Scholar] [CrossRef]
- Passamonti, S.; Terdoslavich, M.; Franca, R.; Vanzo, A.; Tramer, F.; Braidot, E.; Petrussa, E.; Vianello, A. Bioavailability of Flavonoids: A Review of Their Membrane Transport and the Function of Bilitranslocase in Animal and Plant Organisms. Curr. Drug Metab. 2009, 10, 369–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otter, M.; Csader, S.; Keiser, M.; Oswald, S. Expression and Functional Contribution of Different Organic Cation Transporters to the Cellular Uptake of Doxorubicin into Human Breast Cancer and Cardiac Tissue. Int. J. Mol. Sci. 2022, 23, 255. [Google Scholar] [CrossRef]
- Miladiyah, I.; Yuanita, E.; Nuryadi, S.; Jumina, J.; Haryana, S.M.; Mustofa, M. Synergistic Effect of 1,3,6-Trihydroxy-4,5,7-Trichloroxanthone in Combination with Doxorubicin on B-Cell Lymphoma Cells and Its Mechanism of Action Through Molecular Docking. Curr. Ther. Res. Clin. Exp. 2020, 92, 100576. [Google Scholar] [CrossRef]
- Özkan, A.; Stolley, D.L.; Cressman, E.N.K.; McMillin, M.; DeMorrow, S.; Yankeelov, T.E.; Rylander, M.N. Tumor Microenvironment Alters Chemoresistance of Hepatocellular Carcinoma Through CYP3A4 Metabolic Activity. Front. Oncol. 2021, 11, 662135. [Google Scholar] [CrossRef]
- van Eijk, M.; Boosman, R.J.; Schinkel, A.H.; Huitema, A.D.R.; Beijnen, J.H. Cytochrome P450 3A4, 3A5, and 2C8 Expression in Breast, Prostate, Lung, Endometrial, and Ovarian Tumors: Relevance for Resistance to Taxanes. Cancer Chemother. Pharmacol. 2019, 84, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaini, H.R.; Thomas, D.G.; Giordano, T.J.; Johnson, T.D.; Biermann, J.S.; Leu, K.; Hollenberg, P.F.; Baker, L.H. Cytochrome P450 CYP3A4/5 Expression as a Biomarker of Outcome in Osteosarcoma. J. Clin. Oncol. 2003, 21, 2481–2485. [Google Scholar] [CrossRef] [PubMed]
- Floriano-Sanchez, E.; Rodriguez, N.C.; Bandala, C.; Coballase-Urrutia, E.; Lopez-Cruz, J. CYP3A4 Expression in Breast Cancer and Its Association with Risk Factors in Mexican Women. Asian. Pac. J. Cancer Prev. 2014, 15, 3805–3809. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Buddha, B.; Dey, S.; Pal, D.; Mitra, A.K. In Vitro Interaction of the HIV Protease Inhibitor Ritonavir with Herbal Constituents: Changes in P-gp and CYP3A4 Activity. Am. J. Ther. 2004, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Raucy, J.L. Regulation of CYP3A4 Expression in Human Hepatocytes by Pharmaceuticals and Natural Products. Drug Metab. Dispos. 2003, 31, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-S.; Piao, Y.-J.; Kang, K.W. Effects of Quercetin on the Bioavailability of Doxorubicin in Rats: Role of CYP3A4 and P-Gp Inhibition by Quercetin. Arch. Pharm. Res. 2011, 34, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Alrushaid, S.; Zhao, Y.; Sayre, C.L.; Maayah, Z.H.; Forrest, M.L.; Senadheera, S.N.; Chaboyer, K.; Anderson, H.D.; El-Kadi, A.O.S.; Davies, N.M. Mechanistically Elucidating the in Vitro Safety and Efficacy of a Novel Doxorubicin Derivative. Drug Deliv. Transl. Res. 2017, 7, 582–597. [Google Scholar] [CrossRef]
- Yu, C.P.; Wu, P.P.; Hou, Y.C.; Lin, S.P.; Tsai, S.Y.; Chen, C.T.; Chao, P.D.L. Quercetin and Rutin Reduced the Bioavailability of Cyclosporine from Neoral, an Immunosuppressant, through Activating P-Glycoprotein and CYP 3A4. J. Agric. Food Chem. 2011, 59, 4644–4648. [Google Scholar] [CrossRef]
- Choi, S.-J.; Shin, S.-C.; Choi, J.-S. Effects of Myricetin on the Bioavailability of Doxorubicin for Oral Drug Delivery in Rats: Possible Role of CYP3A4 and P-Glycoprotein Inhibition by Myricetin. Arch. Pharm. Res. 2011, 34, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Ito, H.; Ohnishi, R.; Hatano, T. Inhibitory Effects of Polyphenols on Human Cytochrome P450 3A4 and 2C9 Activity. Food Chem. Toxicol. 2010, 48, 429–435. [Google Scholar] [CrossRef]
- Šarić Mustapić, D.; Debeljak, Ž.; Maleš, Ž.; Bojić, M. The Inhibitory Effect of Flavonoid Aglycones on the Metabolic Activity of CYP3A4 Enzyme. Molecules 2018, 23, 2553. [Google Scholar] [CrossRef] [Green Version]
- The Promotive Effects of Antioxidative Apigenin on the Bioavailability of Paclitaxel for Oral Delivery in Rats-Biomolecules & Therapeutics|Korea Science. Available online: http://koreascience.or.kr/article/JAKO201032755114848.page (accessed on 14 September 2022).
- Lee, C.A.; Kadwell, S.H.; Kost, T.A.; Serabjit-Singh, C.J. CYP3A4 Expressed by Insect Cells Infected with a Recombinant Baculovirus Containing Both CYP3A4 and Human NADPH-Cytochrome P450 Reductase Is Catalytically Similar to Human Liver Microsomal CYP3A4. Arch. Biochem. Biophys. 1995, 319, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Kiani, Y.S.; Jabeen, I. Lipophilic Metabolic Efficiency (LipMetE) and Drug Efficiency Indices to Explore the Metabolic Properties of the Substrates of Selected Cytochrome P450 Isoforms. ACS Omega 2020, 5, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Liu, S.; Yang, J.; Wang, Z.; Zhang, H.; Gui, C. Investigation of the Interactions between Flavonoids and Human Organic Anion Transporting Polypeptide 1B1 Using Fluorescent Substrate and 3D-QSAR Analysis. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183210. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Bai, J.; Hu, M.; Xu, Y.; Zhao, S.; Sun, Y.; Wang, B.; Hu, J.; Li, Y. Drug Interaction Study of Flavonoids toward OATP1B1 and Their 3D Structure Activity Relationship Analysis for Predicting Hepatoprotective Effects. Toxicology 2020, 437, 152445. [Google Scholar] [CrossRef] [PubMed]
- Mandery, K.; Bujok, K.; Schmidt, I.; Keiser, M.; Siegmund, W.; Balk, B.; König, J.; Fromm, M.F.; Glaeser, H. Influence of the Flavonoids Apigenin, Kaempferol, and Quercetin on the Function of Organic Anion Transporting Polypeptides 1A2 and 2B1. Biochem. Pharmacol. 2010, 80, 1746–1753. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Yu, A.-M. ABC Transporters in Multidrug Resistance and Pharmacokinetics, and Strategies for Drug Development. Curr. Pharm. Des. 2014, 20, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Q.; Wu, Z.-X.; Yang, Y.; Teng, Q.-X.; Li, Y.-D.; Lei, Z.-N.; Jani, K.A.; Kaushal, N.; Chen, Z.-S. ATP-Binding Cassette (ABC) Transporters in Cancer: A Review of Recent Updates. J. Evid. Based. Med. 2021, 14, 232–256. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The Human ATP-Binding Cassette (ABC) Transporter Superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Stefan, S.M. Multi-Target ABC Transporter Modulators: What next and Where to Go? Future Med. Chem. 2019, 11, 2353–2358. [Google Scholar] [CrossRef]
- Jaramillo, A.C.; Saig, F.A.; Cloos, J.; Jansen, G.; Peters, G.J. How to Overcome ATP-Binding Cassette Drug Efflux Transporter-Mediated Drug Resistance? Cancer Drug Resist. 2018, 1, 6–29. [Google Scholar] [CrossRef] [Green Version]
- Molnár, J.; Engi, H.; Hohmann, J.; Molnár, P.; Deli, J.; Wesolowska, O.; Michalak, K.; Wang, Q. Reversal of Multidrug Resitance by Natural Substances from Plants. Curr. Top. Med. Chem. 2010, 10, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Kadioglu, O.; Khalid, H.; Sugimoto, Y.; Efferth, T. Activity of the Dietary Flavonoid, Apigenin, against Multidrug-Resistant Tumor Cells as Determined by Pharmacogenomics and Molecular Docking. J. Nutr. Biochem. 2015, 26, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Khantamat, O.; Pintha, K. Inhibition of P-Glycoprotein Function and Expression by Kaempferol and Quercetin. J. Chemother. 2005, 17, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Huang, C.; Ma, T.; Jiang, L.; Tang, L.; Shi, T.; Zhang, S.; Zhang, L.; Zhu, P.; Li, J.; et al. Reversal Effect of Quercetin on Multidrug Resistance via FZD7/β-Catenin Pathway in Hepatocellular Carcinoma Cells. Phytomedicine 2018, 43, 37–45. [Google Scholar] [CrossRef] [PubMed]
- van Zanden, J.J.; Wortelboer, H.M.; Bijlsma, S.; Punt, A.; Usta, M.; van Bladeren, P.J.; Rietjens, I.M.C.M.; Cnubben, N.H.P. Quantitative Structure Activity Relationship Studies on the Flavonoid Mediated Inhibition of Multidrug Resistance Proteins 1 and 2. Biochem. Pharmacol. 2005, 69, 699–708. [Google Scholar] [CrossRef]
- Wong, I.L.K.; Chan, K.-F.; Tsang, K.H.; Lam, C.Y.; Zhao, Y.; Chan, T.H.; Chow, L.M.C. Modulation of Multidrug Resistance Protein 1 (MRP1/ABCC1)-Mediated Multidrug Resistance by Bivalent Apigenin Homodimers and Their Derivatives. J. Med. Chem. 2009, 52, 5311–5322. [Google Scholar] [CrossRef]
- Borska, S.; Sopel, M.; Chmielewska, M.; Zabel, M.; Dziegiel, P. Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin. Molecules 2010, 15, 857. [Google Scholar] [CrossRef]
- Wang, S.Y.; Duan, K.M.; Li, Y.; Mei, Y.; Sheng, H.; Liu, H.; Mei, X.; Ouyang, W.; Zhou, H.H.; Liu, Z.Q. Effect of Quercetin on P-Glycoprotein Transport Ability in Chinese Healthy Subjects. Eur. J. Clin. Nutr. 2013, 67, 390–394. [Google Scholar] [CrossRef]
- Singh, A.; Patel, S.K.; Kumar, P.; Das, K.C.; Verma, D.; Sharma, R.; Tripathi, T.; Giri, R.; Martins, N.; Garg, N. Quercetin Acts as a P-Gp Modulator via Impeding Signal Transduction from Nucleotide-Binding Domain to Transmembrane Domain. J. Biomol. Struct. Dyn. 2022, 40, 4507–4515. [Google Scholar] [CrossRef]
- Dudas, B.; Decleves, X.; Cisternino, S.; Perahia, D.; Miteva, M.A. ABCG2/BCRP Transport Mechanism Revealed through Kinetically Excited Targeted Molecular Dynamics Simulations. Comput. Struct. Biotechnol. J. 2022, 20, 4195–4205. [Google Scholar] [CrossRef]
- Nakanishi, T.; Ross, D.D. Breast Cancer Resistance Protein (BCRP/ABCG2): Its Role in Multidrug Resistance and Regulation of Its Gene Expression. Chin. J. Cancer 2012, 31, 73–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Q.; Unadkat, J.D. Role of the Breast Cancer Resistance Protein (BCRP/ABCG2) in Drug Transport—An Update. AAPS J. 2015, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Bai, J.; Zhao, S.; Hu, M.; Sun, Y.; Wang, B.; Ji, M.; Jin, J.; Wang, X.; Hu, J.; et al. Evaluation of Inhibitory Effects of Flavonoids on Breast Cancer Resistance Protein (BCRP): From Library Screening to Biological Evaluation to Structure-Activity Relationship. Toxicol. Vitr. 2019, 61, 104642. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Ross, D.D.; Nakanishi, T.; Bailey-Dell, K.; Zhou, S.; Mercer, K.E.; Sarkadi, B.; Sorrentino, B.P.; Schuetz, J.D. The Stem Cell Marker Bcrp/ABCG2 Enhances Hypoxic Cell Survival through Interactions with Heme. J. Biol. Chem. 2004, 279, 24218–24225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.C.; Chia, W.T.; Chung, M.F.; Lin, K.J.; Hsiao, C.W.; Jin, C.; Lim, W.H.; Chen, C.C.; Sung, H.W. An Implantable Depot That Can Generate Oxygen in Situ for Overcoming Hypoxia-Induced Resistance to Anticancer Drugs in Chemotherapy. J. Am. Chem. Soc. 2016, 138, 5222–5225. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Xue, J.; Li, Z.; Shi, X.; Jiang, B.H.; Fang, J. Chrysin Inhibits Expression of Hypoxia-Inducible Factor-1alpha through Reducing Hypoxia-Inducible Factor-1alpha Stability and Inhibiting Its Protein Synthesis. Mol. Cancer Ther. 2007, 6, 220–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.-C.; Hsieh, Y.-L.; Hung, C.-M.; Chien, P.-H.; Chien, Y.-F.; Chen, L.-C.; Tu, C.-Y.; Chen, C.-H.; Hsu, S.-C.; Lin, Y.-M.; et al. BCRP/ABCG2 Inhibition Sensitizes Hepatocellular Carcinoma Cells to Sorafenib. PLoS One 2013, 8, e83627. [Google Scholar] [CrossRef]
- Ebert, B.; Seidel, A.; Lampen, A. Phytochemicals Induce Breast Cancer Resistance Protein in Caco-2 Cells and Enhance the Transport of Benzo[a]Pyrene-3-Sulfate. Toxicological. Sci. 2007, 96, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Syahputra, R.A.; Harahap, U.; Dalimunthe, A.; Nasution, M.P.; Satria, D. The Role of Flavonoids as a Cardioprotective Strategy against Doxorubicin-Induced Cardiotoxicity: A Review. Molecules 2022, 27, 1320. [Google Scholar] [CrossRef]
- van Acker, F.A.; Hulshof, J.W.; Haenen, G.R.; Menge, W.M.; van der Vijgh, W.J.; Bast, A. New Synthetic Flavonoids as Potent Protectors against Doxorubicin-Induced Cardiotoxicity. Free Radic. Biol. Med. 2001, 31, 31–37. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R.M.M.; Bruynzeel, A.M.E.; Van der Vijgh, W.J.F. Protection by Flavonoids against Anthracycline Cardiotoxicity: From Chemistry to Clinical Trials. Cardiovasc. Toxicol. 2007, 7, 154–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, B. Doxorubicin Induces Cardiotoxicity through Upregulation of Death Receptors Mediated Apoptosis in Cardiomyocytes. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.V.; Kim, J. Iron Promotes Cardiac Doxorubicin Retention and Toxicity Through Downregulation of the Mitochondrial Exporter ABCB8. Front. Pharmacol. 2022, 13, 322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Shi, J.; Nice, E.C.; Huang, C.; Shi, Z. The Multifaceted Role of Flavonoids in Cancer Therapy: Leveraging Autophagy with a Double-Edged Sword. Antioxidants 2021, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Belhan, S.; Özkaraca, M.; Özdek, U.; Kömüroğlu, A.U. Protective Role of Chrysin on Doxorubicin-Induced Oxidative Stress and DNA Damage in Rat Testes. Andrologia 2020, 52, e13747. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Teves, S.S.; Kemp, C.J.; Henikoff, S. Doxorubicin, DNA Torsion, and Chromatin Dynamics. Biochim. Biophys. Acta 2014, 1845, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nafisi, S.; Hashemi, M.; Rajabi, M.; Tajmir-Riahi, H.A. DNA Adducts with Antioxidant Flavonoids: Morin, Apigenin, and Naringin. DNA Cell Biol. 2008, 27, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Waihenya, S.; Şenel, P.; Osonga, F.J.; Erdoğan, T.; Altay, F.; Gölcü, A.; Sadik, O.A. Mechanism of Interactions of DsDNA Binding with Apigenin and Its Sulfamate Derivatives Using Multispectroscopic, Voltammetric, and Molecular Docking Studies. ACS Omega 2021, 6, 5124–5137. [Google Scholar] [CrossRef]
- Kumar, S.; Nair, M.S. Deciphering the Interaction of Flavones with Calf Thymus DNA and Octamer DNA Sequence (CCAATTGG)2. RSC Adv. 2021, 11, 29354–29371. [Google Scholar] [CrossRef]
- Wang, Z.; Cui, M.; Song, F.; Lu, L.; Liu, Z.; Liu, S. Evaluation of Flavonoids Binding to DNA Duplexes by Electrospray Ionization Mass Spectrometry. J. Am. Soc. Mass. Spectrom. 2008, 19, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Sha, Y.; Chen, X.; Niu, B.; Chen, Q. The Interaction Mode of Groove Binding Between Quercetin and Calf Thymus DNA Based on Spectrometry and Simulation. Chem. Biodivers. 2017, 14, e1700133. [Google Scholar] [CrossRef] [PubMed]
- Korga-Plewko, A.; Michalczyk, M.; Adamczuk, G.; Humeniuk, E.; Ostrowska-Lesko, M.; Jozefczyk, A.; Iwan, M.; Wojcik, M.; Dudka, J. Apigenin and Hesperidin Downregulate DNA Repair Genes in MCF-7 Breast Cancer Cells and Augment Doxorubicin Toxicity. Molecules 2020, 25, 4421. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, H.; Shiga, C.; Imai, M.; Ikemura, K.; Kitamura, Y.; Ohta, K.; Miyazawa, D.; Sakanashi, M.; Tahira, T.; Maeda, T.; et al. Idarubicin, an Anthracycline, Induces Oxidative DNA Damage in the Presence of Copper (II). Anticancer Res. 2020, 40, 5399–5404. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, T.; Long, M.; Li, P. Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine. Oxid. Med. Cell Longev. 2020, 2020, 8825387. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Krishnaswamy, S.; Devashya, V.; Sethuraman, S.; Krishnan, U.M. Flavonoid-Metal Ion Complexes: A Novel Class of Therapeutic Agents. Med. Res. Rev. 2014, 34, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Pan, J.; Zhu, X.; Zhang, S.; Liu, C.; Sun, J.; Li, Y.; Chen, S.; Huang, J.; Cao, C.; et al. Pharmacokinetic Herb-Drug Interactions between Aidi Injection and Doxorubicin in Rats with Diethylnitrosamine-Induced Hepatocellular Carcinoma. BMC Pharmacol. Toxicol. 2021, 22, 48. [Google Scholar] [CrossRef]
- Omonga, N.; Zia, Z.; Ghanbour, H.; Ragazzon-Smith, A.; Foster, H.; Hadfield, J.; Ragazzon, P. Facile Synthesis and Biological Evaluation of Chrysin Derivatives. J. Chem. Res. 2021, 45, 1083–1092. [Google Scholar] [CrossRef]
- Xiao, Z.; Jiang, Y.; Chen, X.-F.; Wang, C.-Q.; Xu, W.; Liu, Y.; Hu, S.-S.; Huang, X.-R.; Shan, L.-J.; Tang, Y.-H.; et al. The Hepatorenal Toxicity and Tumor Response of Chemotherapy With or Without Aidi Injection in Advanced Lung Cancer: A Meta-Analysis of 80 Randomized Controlled Trials. Clin. Ther. 2020, 42, 515–543.e31. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Giuliano, A.; Law, R.; Van Herle, A.V. Apigenin Inhibits Growth and Induces G2/M Arrest by Modulating Cyclin-CDK Regulators and ERK MAP Kinase Activation in Breast Carcinoma Cells. Anticancer. Res. 2001, 21, 413–420. [Google Scholar] [PubMed]
- Ayob, Z.; Mohd Bohari, S.P.; Abd Samad, A.; Jamil, S. Cytotoxic Activities against Breast Cancer Cells of Local Justicia Gendarussa Crude Extracts. Evid.-Based Complement. Altern. Med. 2014, 2014, 732980. [Google Scholar] [CrossRef] [Green Version]
- Balabhadrapathruni, S.; Thomas, T.J.; Yurkow, E.J.; Amenta, P.S.; Thomas, T. Effects of Genistein and Structurally Related Phytoestrogens on Cell Cycle Kinetics and Apoptosis in MDA-MB-468 Human Breast Cancer Cells. Oncol. Rep. 2000, 7, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Halagowder, D.; Sivasithambaram, N.D. Quercetin Suppresses Twist to Induce Apoptosis in MCF-7 Breast Cancer Cells. PloS one 2015, 10, e0141370. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, N.L.; Obradovic, A.D.; Vukic, M.D.; Jovanovic, D.; Djurdjevic, P.M. Cytotoxic, Proapoptotic and Antioxidative Potential of Flavonoids Isolated from Propolis against Colon (HCT-116) and Breast (MDA-MB-231) Cancer Cell Lines. Food Res. Int. 2018, 106, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Yadegarynia, S. Profiling Flavonoid Cytotoxicity in Human Breast Cancer Cell Lines. Master’s Thesis, San Jose State University, San Jose, CA, USA, 2012. [Google Scholar] [CrossRef]
- Koh, S.Y.; Moon, J.Y.; Unno, T.; Cho, S.K. Baicalein Suppresses Stem Cell-Like Characteristics in Radio- and Chemoresistant MDA-MB-231 Human Breast Cancer Cells through Up-Regulation of IFIT2. Nutrients 2019, 11, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushina, Y.; Shiomi, K.; Onodera, T.; Kuriyama, I. Inhibitory effects of apigenin on mammalian dna polymerase and proliferation of human cancer cells. In Apigenin and Naringenin: Natural Sources, Pharmacology and Role in Cancer…—Google Książki; Stacks, N.M., Ed.; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2015; pp. 53–75. [Google Scholar]
- Ravishankar, D.; Watson, K.A.; Greco, F.; Osborn, H.M.I. Novel Synthesised Flavone Derivatives Provide Significant Insight into the Structural Features Required for Enhanced Anti-Proliferative Activity. RSC Adv. 2013, 6, 64544–64556. [Google Scholar] [CrossRef] [Green Version]
- Haddad, A.Q.; Venkateswaran, V.; Viswanathan, L.; Teahan, S.J.; Fleshner, N.E.; Klotz, L.H. Novel Antiproliferative Flavonoids Induce Cell Cycle Arrest in Human Prostate Cancer Cell Lines. Prostate Cancer Prostatic Dis. 2006, 9, 68–76. [Google Scholar] [CrossRef]
- Kamat, S.; Kumari, M.; Jayabaskaran, C. Infrared Spectroscopy and Flow Cytometry Studies on the Apoptotic Effect of Nano-Chrysin in HeLa Cells. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2022, 282, 121666. [Google Scholar] [CrossRef]
- Souza, R.P.; Bonfim-Mendonça, P.D.S.; Gimenes, F.; Ratti, B.A.; Kaplum, V.; Bruschi, M.L.; Nakamura, C.V.; Silva, S.O.; Maria-Engler, S.S.; Consolaro, M.E.L. Oxidative Stress Triggered by Apigenin Induces Apoptosis in a Comprehensive Panel of Human Cervical Cancer-Derived Cell Lines. Oxidative Med. Cell. Longev. 2017, 2017, 1512745. [Google Scholar] [CrossRef] [Green Version]
- Afroze, N.; Pramodh, S.; Almutary, A.G.; Rizvi, T.A.; Rais, N.; Raina, R.; Faiyazuddin, M.; Alnuqaydan, A.M.; Hussain, A. Kaempferol Regresses Carcinogenesis through a Molecular Cross Talk Involved in Proliferation, Apoptosis and Inflammation on Human Cervical Cancer Cells, HeLa. Appl. Sci. 2022, 12, 3155. [Google Scholar] [CrossRef]
- Danihelová, M.; Veverka, M.; Šturdík, E.; Jantová, S. Antioxidant Action and Cytotoxicity on HeLa and NIH-3T3 Cells of New Quercetin Derivatives. Interdiscip. Toxicol. 2013, 6, 209. [Google Scholar] [CrossRef] [Green Version]
- Boonmasawai, S.; Sariya, L.; Leesombun, A.; Chaichoun, K.; Taowan, J.; Thongjuy, O. Anti-Proliferative and Total ERK1/2 Inhibitory Effects of Plant Flavonols on Human Cervical Cancer (HeLa) Cells. Thai J. Vet. Med. 2018, 48, 541–549. [Google Scholar]
- Zayed, M.M.M.; Sahyon, H.A.; Hanafy, N.A.N.; El-Kemary, M.A. The Effect of Encapsulated Apigenin Nanoparticles on HePG-2 Cells through Regulation of P53. Pharmaceutics 2022, 14, 1160. [Google Scholar] [CrossRef] [PubMed]
- Kluska, M.; Juszczak, M.; Żuchowski, J.; Stochmal, A.; Woźniak, K. Kaempferol and Its Glycoside Derivatives as Modulators of Etoposide Activity in HL-60 Cells. Int. J. Mol. Sci. 2021, 22, 3520. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Quercetin Induces Apoptosis via Caspase Activation, Regulation of Bcl-2, and Inhibition of PI-3-Kinase/Akt and ERK Pathways in a Human Hepatoma Cell Line (HepG2). J. Nutr. 2006, 136, 2715–2721. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, J.; Yu, X.; Xu, S.; Li, D.; Zheng, Q.; Yin, Y. Myricetin Suppresses the Propagation of Hepatocellular Carcinoma via Down-Regulating Expression of YAP. Cells 2019, 8, 358. [Google Scholar] [CrossRef] [Green Version]
- Kuntz, S.; Wenzel, U.; Daniel, H. Comparative Analysis of the Effects of Flavonoids on Proliferation, Cytotoxicity, and Apoptosis in Human Colon Cancer Cell Lines. Eur. J. Nutr. 1999, 38, 133–142. [Google Scholar] [CrossRef]
- Sezer, E.D.; Oktay, L.M.; Karadadaş, E.; Memmedov, H.; Selvi Gunel, N.; Sözmen, E. Assessing Anticancer Potential of Blueberry Flavonoids, Quercetin, Kaempferol, and Gentisic Acid, Through Oxidative Stress and Apoptosis Parameters on HCT-116 Cells. J. Med. Food 2019, 22, 1118–1126. [Google Scholar] [CrossRef]
- Combination of Quercetin and Kaempferol Enhances in Vitro Cytotoxicity on Human Colon Cancer (HCT-116) Cells. Available online: https://digital.csic.es/bitstream/10261/115037/1/RNP2014_262.pdf (accessed on 10 September 2022).
- Shenouda, N.S.; Zhou, C.; Browning, J.D.; Ansell, P.J.; Sakla, M.S.; Lubahn, D.B.; MacDonald, R.S. Phytoestrogens in Common Herbs Regulate Prostate Cancer Cell Growth in Vitro. Nutr. Cancer 2004, 49, 200–208. [Google Scholar] [CrossRef]
- Özgönül, A.M.; Aşık, A.; Durmaz, B.; Aslaminabad, R.; Gündüz, C.; Sozmen, E.Y. Antiproliferative Effect of Rosehip Tea Phenolics in Prostate Cancer Cell Lines. Turk. J. Biochem. 2020, 45, 423–428. [Google Scholar] [CrossRef]
- Da, J.; Xu, M.; Wang, Y.; Li, W.; Lu, M.; Wang, Z. Kaempferol Promotes Apoptosis While Inhibiting Cell Proliferation via Androgen-Dependent Pathway and Suppressing Vasculogenic Mimicry and Invasion in Prostate Cancer. Anal. Cell. Pathol. 2019, 2019, 1907698. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zhang, C.; Huang, H.; Yang, B.; Xiao, G.; Kong, D.; Tian, Q.; Song, Q.; Song, Y.; Tan, H.; et al. The Natural Compound Myricetin Effectively Represses the Malignant Progression of Prostate Cancer by Inhibiting PIM1 and Disrupting the PIM1/CXCR4 Interaction. Cell. Physiol. Biochem. 2018, 48, 1230–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Yang, J.; Li, K.; Luo, J.; Yang, S.; Song, J.R.; Chen, C.; Pan, W.D. Synthesis of Flavone Derivatives via N-Amination and Evaluation of Their Anticancer Activities. Molecules 2019, 24, 2723. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Ganguli, A.; Dastidar, D.G.; Acharya, B.R.; Das, A.; Chakrabarti, G. Apigenin Shows Synergistic Anticancer Activity with Curcumin by Binding at Different Sites of Tubulin. Biochimie 2013, 95, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Hang, M.; Zhao, F.; Chen, S.B.; Sun, Q.; Zhang, C.X. Kaempferol Modulates the Metastasis of Human Non-Small Cell Lung Cancer Cells by Inhibiting Epithelial-Mesenchymal Transition. Bangladesh J. Pharmacol. 2015, 10, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Zhaorigetu; Farrag, I.M.; Belal, A.; Al Badawi, M.H.; Abdelhady, A.A.; AbouGalala, F.M.A.; El-Sharkawy, A.; EL-Dahshan, A.A.; Mehany, A.B.M. Antiproliferative, Apoptotic Effects and Suppression of Oxidative Stress of Quercetin against Induced Toxicity in Lung Cancer Cells of Rats: In Vitro and In Vivo Study. J. Cancer 2021, 12, 5249. [Google Scholar] [CrossRef]
- Han, J.; Cheng, C.; Zhang, J.; Fang, J.; Yao, W.; Zhu, Y.; Xiu, Z.; Jin, N.; Lu, H.; Li, X.; et al. Myricetin Activates the Caspase-3/GSDME Pathway via ER Stress Induction of Pyroptosis in Lung Cancer Cells. Front. Pharmacol. 2022, 13, 3286. [Google Scholar] [CrossRef]
- Samarghandian, S.; Nezhad, M.; Mohammadi, G. Role of Caspases, Bax and Bcl-2 in Chrysin-Induced Apoptosis in the A549 Human Lung Adenocarcinoma Epithelial Cells. Anti-Cancer Agents Med. Chem. 2014, 14, 901–909. [Google Scholar] [CrossRef]


| CHRY | API | KAE | QUE | MIR | DOX | ||
|---|---|---|---|---|---|---|---|
| Physicochemical Properties 1 | Molecular Weight | 254.24 | 270.24 | 286.24 | 302.24 | 318.24 | 543.52 |
| Volume | 206.92 | 216.03 | 224.05 | 232.07 | 240.08 | ||
| Aromatic heavy atoms | 12 | 16 | 16 | 16 | 16 | 12 | |
| H-bond acceptors | 4 | 5 | 6 | 7 | 8 | 7 | |
| H-bond donors | 2 | 3 | 4 | 5 | 6 | 12 | |
| TPSA | 70.67 | 90.9 | 111.13 | 131.36 | 151.59 | 206.08 | |
| SILICOS-IT Log P 2 | 3.02 | 2.52 | 2.03 | 1.54 | 1.06 | 1.17 | |
| ESOL Log S | −4.19 | −3.94 | −3.31 | −3.16 | −3.01 | 132.66 | |
| Molar Refractivity | 71.97 | 73.99 | 76.01 | 78.03 | 80.06 | 131.52 | |
| Pharmacokinetics 1 | CYP1A2 | + | + | + | + | + | − |
| CYP2C19 | − | − | − | − | − | − | |
| CYP2C9 | + | − | − | − | − | − | |
| CYP2D6 | − | + | + | + | + | − | |
| CYP3A4 | + | + | + | + | + | − | |
| DFT electrochemical parameters 3,4 | HOMO [eV] | −6.43 | −6.24 | −5.78 | −5.65 | −5.71 | −5.32 |
| LUMO [eV] | −2.04 | −1.88 | −1.97 | −1.97 | −2.06 | −4.09 | |
| HOMO-LUMO energy gap [eV] | 4.39 | 4.36 | 3.81 | 3.68 | 3.65 | 1.23 | |
| Dipole moment | 3.84 | 2.47 | 5.47 | 4.23 | 6.78 | 7.77 | |
| Electron affinity | 4.24 | 4.06 | 3.87 | 3.81 | 3.88 | 6.80 |
| Compound | Concentration | Cell Line | Cell Type | Effect | Ref |
|---|---|---|---|---|---|
| Chrysin | 10–20 µM | BEL-7402/ADM cells | Hepatocellular carcinoma | ↓ mRNA and protein expression of Nrf2, HO-1, MRP5, and aldo-keto reductase family 1 member B10 (AKR1B10) | [137] |
| Chrysin [nanostructural lipid carriers] | 5–50 µM | MCF-7 | Breast cancer | ↓mRNA and protein expression of Nrf2; NQO1, HO1 i MRP1 | [141] |
| Chrysin | 10–60 µM | T98, U251, U87 | human glioblastomas | ↓ Nrf2, NQO-1, HO-1 (Keap1-independent); ↓ ERK signaling | [142] |
| Apigenin | 10 μM | BEL-7402/ADM | Hepatocellular carcinoma | inhibiting miR-101/Nrf2 pathway | [143] |
| Kaempferol | 25 μM | A549, NCIH460 | NSCLC | ↓ mRNA and protein expression of Nrf2; ↓ AKR1C1, NQO1, HO1 i GS; ↑ ROS | [144] |
| 25–50 μM | PANC-1, PaCa-2 | pancreatic cancer | ROS-dependent suppression Akt/mTOR signaling; ↓ Keap; Nrf2 (ambiguous impact) | [145] | |
| Quercetin | 50 μM | human xenograft acute myeloid leukemia (AML) models, and in vitro using leukemia cell lines | ↓ Nnf2 nuclear localization; ↓ pNrf2 ↓ HDAC4; | [146] |
| Cell Internalization of Rhodamine-123 in MCF-7/ADR 1 Cells Overexpressing P-gp | Ref | Human CYP3A4 Microsomes Expressed in Baculovirus 2-Insect Cell, Lineweaver–Burk Plot Analysis | Ref | Residual Activity Compared to Control (6β-Hydroxylation of Testosterone-Marker Reaction of CYP3A4 Activity) | Ref | ||
|---|---|---|---|---|---|---|---|
| Flavonols | Myricetin | 7.8 | [185] | 44.5 | [186] | 133 ± 35 | [187] |
| Quercetin | 1.97 | [182] | 22.1 | [186] | 126 ± 10 | [187] | |
| Kaempferol | 8.6 | [182] | 8.8 | [186] | 101 ± 14 | [187] | |
| Flavones | Apigenin | 1.8 | [182] | 0.4 | [186] | 24 ± 3 | [187] |
| Chrysin | nd | 0.9 | [186] | 17 ± 3 | [187] |
| Binging Constants (K[M−]]) | Methods | DNA Model | Measurement Conditions | References | |
|---|---|---|---|---|---|
| CHRYSIN | 1.21 × 105 | Abs | d(CCAATTGG)2 | room temperature; absorption: 200 to 800 nm; fluorescence: Ex 326 nm, Em spectra 320 to 650 nm, | [227] |
| 9.07 × 103 | Fluo | ||||
| 9.03 × 105 | Abs | Ct-DNA | |||
| 3.03 × 105 | Fluo | ||||
| APIGENIN | 7.10 × 104 | FT-IR & Abs | Ct-DNA | FT-IR: sharp DNA band at 968 cm−1 as internal reference, obtained difference spectra [(DNA solution + ligand)-(DNA solution)]; Abs: of DNA at 260 nm, wavelength range of 245–445 upon titration of dsDNA | [225] |
| 9.12 × 104 | Abs | dsDNA | [226] | ||
| 0.73 × 104 | Abs | Ct-DNA | [140] | ||
| QUERCETIN | 17.54 × 105 | Fluo, 300 K | Ct-DNA | fluorescence: Ex 220–660 nm 57; Em 240–680 nm | [229] |
| 8.97 × 105 | Fluo, 310 K | ||||
| 4.80 × 105 | Fluo, 320 K | ||||
| 18.0 × 10 4 | Abs | Ct-DNA | [140] | ||
| KAEMPFEROL | 18.0 × 104 | Abs | Ct-DNA | measurement of absorption between 200 nm and 500 nm | [140] |
| MYRICETIN | 8.63 × 104 | Abs | Ct-DNA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golonko, A.; Olichwier, A.J.; Swislocka, R.; Szczerbinski, L.; Lewandowski, W. Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity. Int. J. Mol. Sci. 2023, 24, 391. https://doi.org/10.3390/ijms24010391
Golonko A, Olichwier AJ, Swislocka R, Szczerbinski L, Lewandowski W. Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity. International Journal of Molecular Sciences. 2023; 24(1):391. https://doi.org/10.3390/ijms24010391
Chicago/Turabian StyleGolonko, Aleksandra, Adam Jan Olichwier, Renata Swislocka, Lukasz Szczerbinski, and Włodzimierz Lewandowski. 2023. "Why Do Dietary Flavonoids Have a Promising Effect as Enhancers of Anthracyclines? Hydroxyl Substituents, Bioavailability and Biological Activity" International Journal of Molecular Sciences 24, no. 1: 391. https://doi.org/10.3390/ijms24010391