
Simulated Microgravity Influences Immunity-Related Biomarkers in Lung Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Results
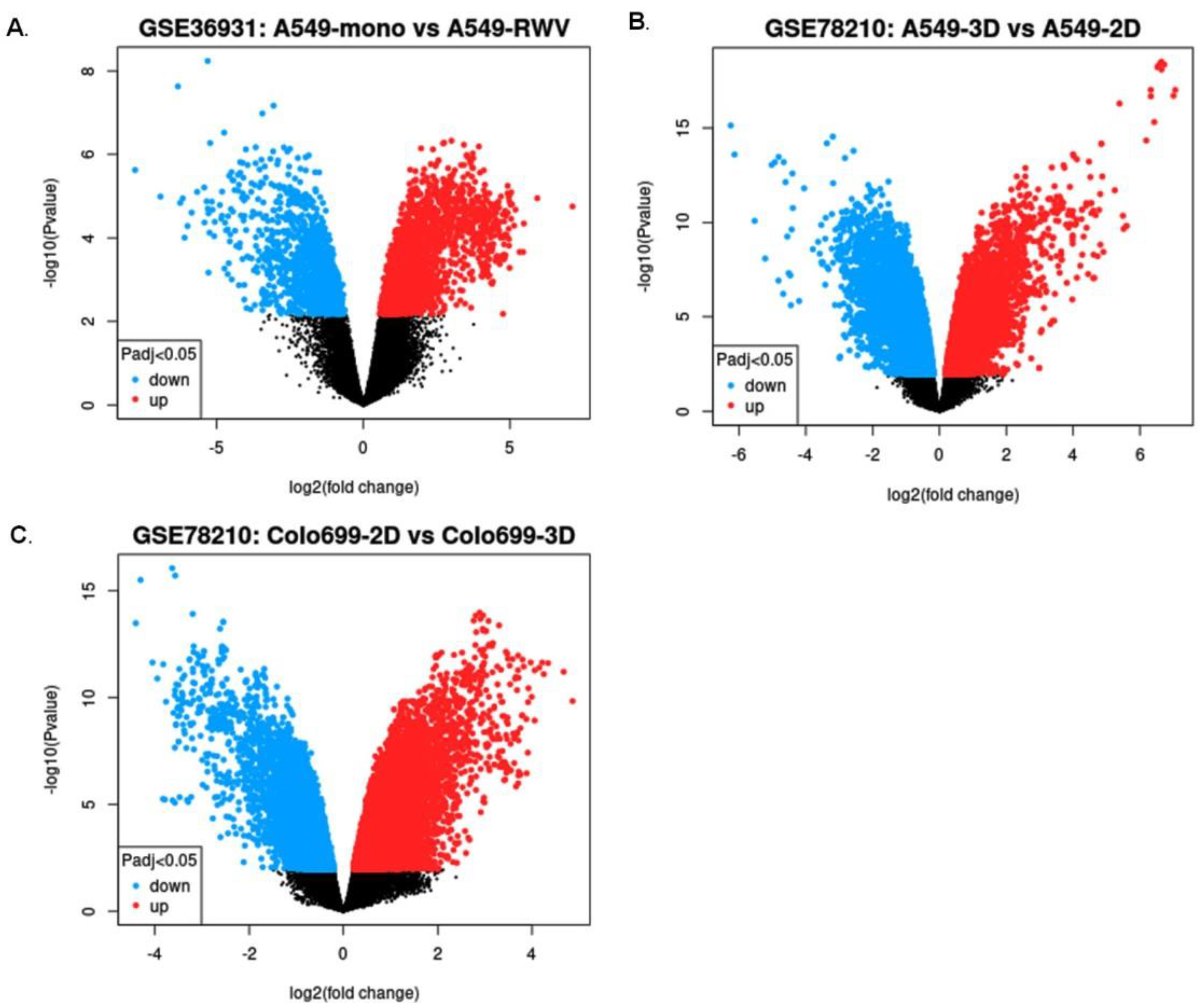
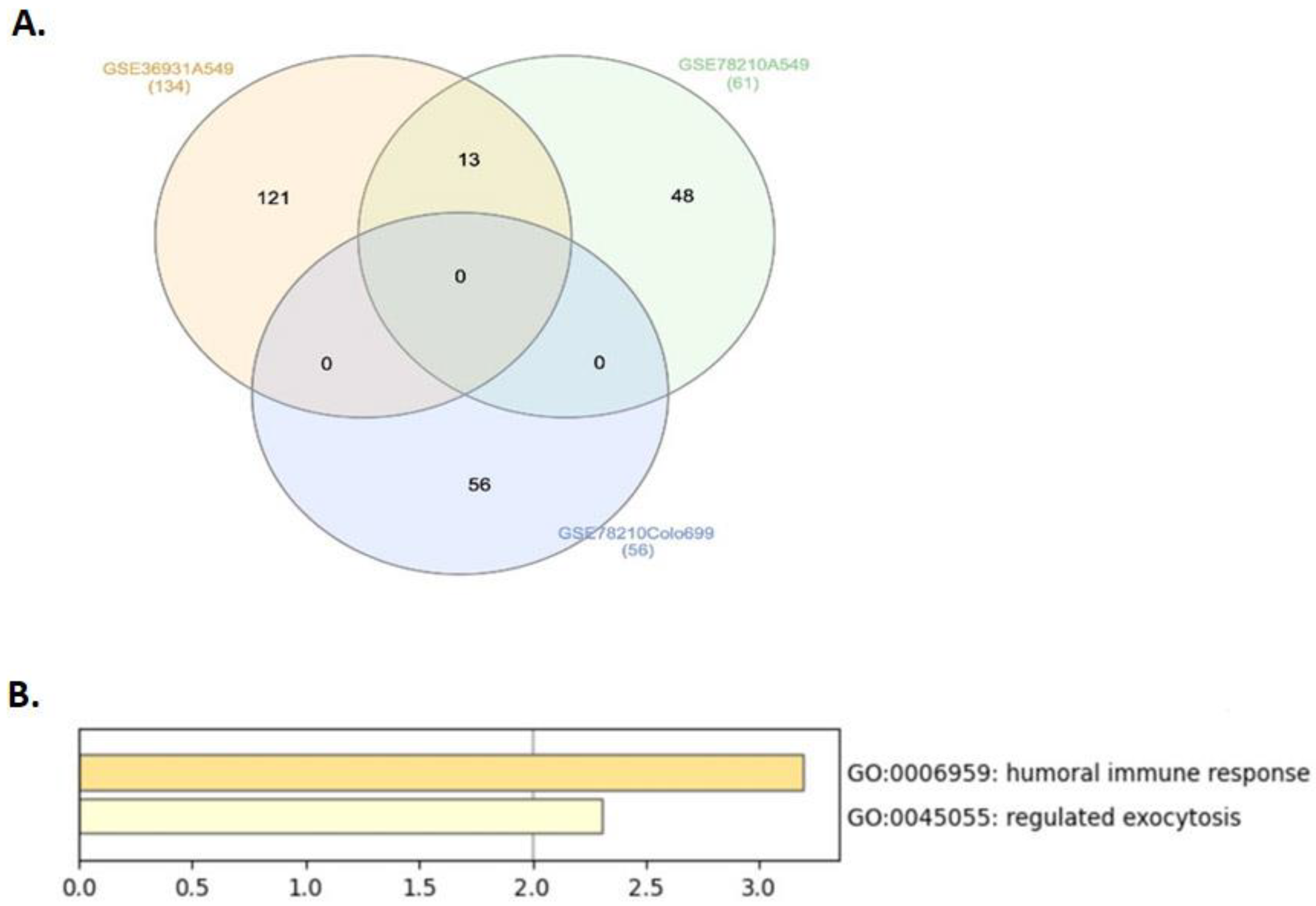
2.1. Top Common Differentially Expressed Genes Identified in Lung Cancer Cells under Simulated Microgravity (SMG) Compared with Ground Gravity (GG)
2.2. The Differentially Expressed Genes Correlated with Lung Adenocarcinoma Patients’ Clinical Prognosis
2.3. Expression of FCGBP, BPIFB1, F5, CFB, and CST1 in A549 Cells Post-Simulated Microgravity
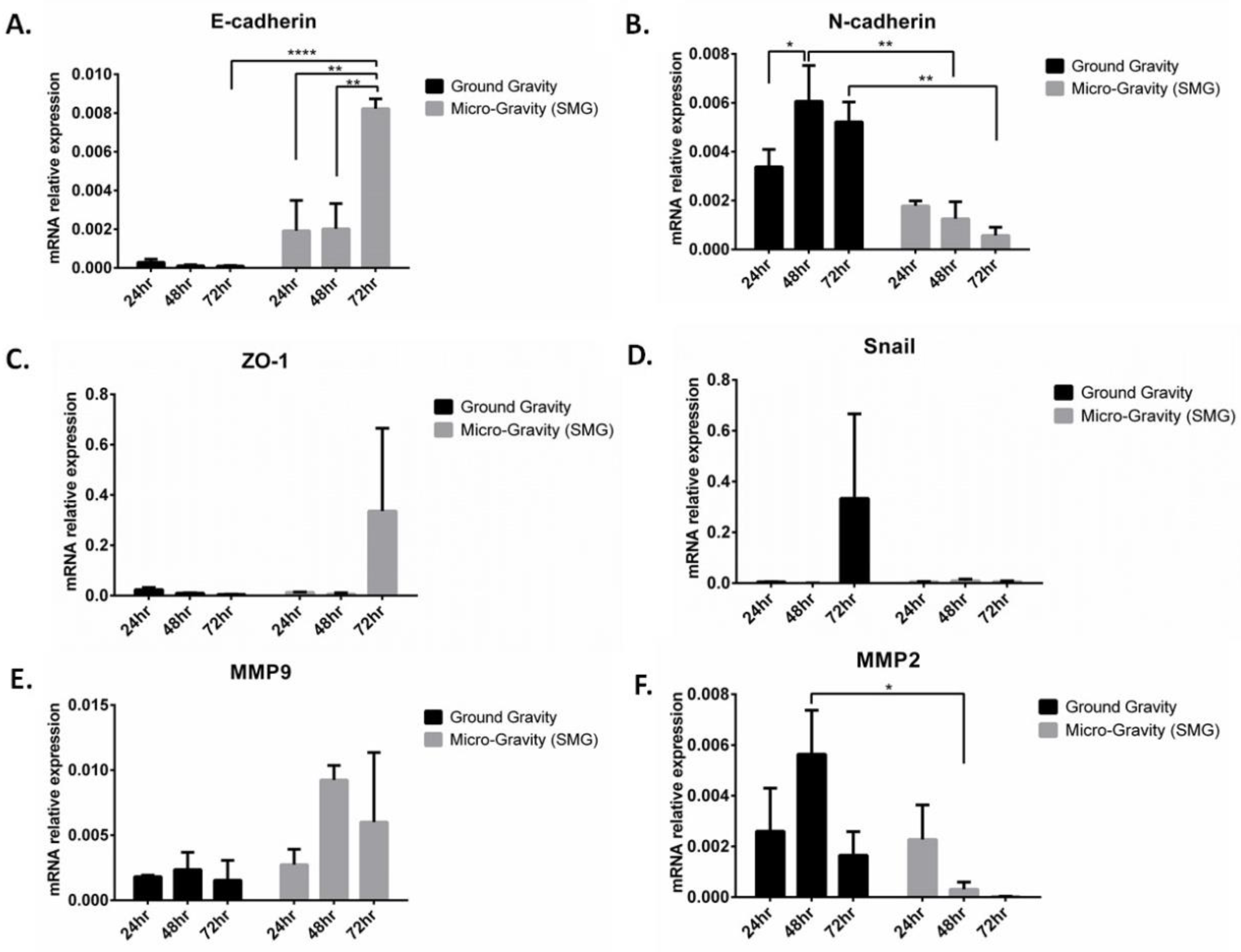
2.4. Simulated Microgravity Reduces Cell Viability and Reverses Epithelial-to-Mesenchymal Transition in A549 Lung Cancer Cell Line
2.5. Epithelial-to-Mesenchymal Transition Pathway Correlates with the Expression of FCGBP, BPIFB1, F5, CFB, and CST1
3. Discussion
4. Materials and Methods
4.1. Datasets to Identify Common Differentially Expressed Genes (DEGs) in Lung Cancer Cell Lines Exposed to Simulated Microgravity
4.2. GEO2R Gene Set Enrichment Analysis to Generate DEGs in Each Dataset
4.3. Quantification of Clinicopathological Involvement of the Identified Genes
4.4. Enriched Ontology Clustering for the Identified Genes
4.5. Cell Line and Cell Culture
4.6. Microgravity
4.7. Cell Viability Assay
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9. Gene–Gene Interaction Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, R.J.; Averner, M.J.N. Humans in space. Nature 2001, 409, 1115–6828. [Google Scholar] [CrossRef]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the impact of microgravity at the cellular level: Implications for human disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Liu, H.; Zhou, S.; Zhou, X.; Huang, Y.-E.; Hou, F.; Jiang, W. Integrative analysis of regulatory module reveals associations of microgravity with dysfunctions of multi-body systems and tumorigenesis. Int. J. Mol. Sci. 2020, 21, 7585. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.E.; Chouker, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune system dysregulation during spaceflight: Potential countermeasures for deep space exploration missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Nassef, M.Z.; Melnik, D.; Kopp, S.; Sahana, J.; Infanger, M.; Lützenberg, R.; Relja, B.; Wehland, M.; Grimm, D.; Krüger, M. Breast cancer cells in microgravity: New aspects for cancer research. Int. J. Mol. Sci. 2020, 21, 7345. [Google Scholar] [CrossRef]
- Takamatsu, Y.; Koike, W.; Takenouchi, T.; Sugama, S.; Wei, J.; Waragai, M.; Sekiyama, K.; Hashimoto, M. Protection against neurodegenerative disease on Earth and in space. NPJ Microgravity 2016, 2, 16013. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Krüger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schütte, A. Real microgravity influences the cytoskeleton and focal adhesions in human breast cancer cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef] [Green Version]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis induction and alteration of cell adherence in human lung cancer cells under simulated microgravity. Int. J. Mol. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [Green Version]
- Ricci, G.; Cucina, A.; Proietti, S.; Dinicola, S.; Ferranti, F.; Cammarota, M.; Filippini, A.; Bizzarri, M.; Catizone, A. Microgravity induces transient EMT in human keratinocytes by early down-regulation of E-cadherin and cell-adhesion remodeling. J. Appl. Sci. 2020, 11, 110. [Google Scholar] [CrossRef]
- Ahn, C.B.; Lee, J.-H.; Han, D.G.; Kang, H.-W.; Lee, S.-H.; Lee, J.-I.; Son, K.H.; Lee, J.W. Simulated microgravity with floating environment promotes migration of non-small cell lung cancers. Sci. Rep. 2019, 9, 14553. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, M.E.; Noto, A.; De Vitis, C.; Masiello, M.G.; Coluccia, P.; Proietti, S.; Giovagnoli, M.R.; Ricci, A.; Giarnieri, E.; Cucina, A. Lung cancer stem cell lose their stemness default state after exposure to microgravity. BioMed Res. Int. 2014, 2014, 470253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infanger, M.; Kossmehl, P.; Shakibaei, M.; Bauer, J.; Kossmehl-Zorn, S.; Cogoli, A.; Curcio, F.; Oksche, A.; Wehland, M.; Kreutz, R.J.C.; et al. Simulated weightlessness changes the cytoskeleton and extracellular matrix proteins in papillary thyroid carcinoma cells. Cell Tissue Res. 2006, 324, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Microgravity, L. Tumor cells in microgravity. In Into Space: A Journey of How Humans Adapt and Live in Microgravity; IntechOpen: London, UK, 2018; Volume 139, pp. 259–268. [Google Scholar]
- Grimm, D.; Schulz, H.; Krüger, M.; Cortés-Sánchez, J.L.; Egli, M.; Kraus, A.; Sahana, J.; Corydon, T.J.; Hemmersbach, R.; Wise, P.M. The fight against cancer by microgravity: The multicellular spheroid as a metastasis model. Int. J. Mol. Sci. 2022, 23, 3073. [Google Scholar] [CrossRef]
- Schabath, M.B.; Cote, M.L. Cancer progress and priorities: Lung cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.H.; Ahn, C.B.; Son, K.H.; Yi, E.; Son, H.S.; Kim, H.-S.; Lee, S.H. Simulated microgravity effects on nonsmall cell lung cancer cell proliferation and migration. Aerosp. Med. Hum. Perform. 2017, 88, 82–89. [Google Scholar] [CrossRef]
- Chang, D.; Xu, H.; Guo, Y.; Jiang, X.; Liu, Y.; Li, K.; Pan, C.; Yuan, M.; Wang, J.; Li, T.; et al. Simulated microgravity alters the metastatic potential of a human lung adenocarcinoma cell line. Vitr. Cell. Dev. Biol. Anim. 2013, 49, 170–177. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-mesenchymal transition in cancer: A historical overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef]
- Shi, S.; Li, Q.; Cao, Q.; Diao, Y.; Zhang, Y.; Yue, L.; Wei, L. EMT transcription factors are involved in the altered cell adhesion under simulated microgravity effect or overloading by regulation of E-cadherin. Int. J. Mol. Sci. 2020, 21, 1349. [Google Scholar] [CrossRef] [Green Version]
- Topal, U.; Zamur, C. Microgravity, stem cells, and cancer: A new Hope for cancer treatment. Stem Cells Int. 2021, 2021, 5566872. [Google Scholar] [CrossRef]
- Ksiazkiewicz, M.; Markiewicz, A.; Zaczek, A.J. Epithelial-mesenchymal transition: A hallmark in metastasis formation linking circulating tumor cells and cancer stem cells. Pathobiology 2012, 79, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Tian, D.; Chen, J.; Tan, Y.; Cheng, Y.; Ye, L.; Deng, G.; Liu, B.; Yuan, F.; Zhang, S. FCGBP Is a Prognostic Biomarker and Associated With Immune Infiltration in Glioma. Front. Oncol. 2021, 11, 769033. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Ogata, H.; Morikawa, M.; Iijima, S.; Harada, N.; Yoshida, T.; Brown, W.; Inoue, N.; Hamada, Y.; Ishii, H. Distribution and partial characterisation of IgG Fc binding protein in various mucin producing cells and body fluids. Gut 2002, 51, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Y.; Liu, H.; Zhang, Q.; Song, H.; Tang, J.; Fu, J.; Wang, X. FcGBP was upregulated by HPV infection and correlated to longer survival time of HNSCC patients. Oncotarget 2017, 8, 86503–86514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Huo, H.; Mao, Y.; Li, X.; Dong, L. A risk score model for the prediction of osteosarcoma metastasis. FEBS Open Bio 2019, 9, 519–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, T.; Vijayalakshmi, N.; Gopal, G.; Sabitha, K.; Shirley, S.; Raja, U.M.; Ramakrishnan, S. Identification and validation of genes involved in gastric tumorigenesis. Cancer Cell Int. 2010, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Guan, C.; Shang, X.; Ying, X.; Mei, S.; Zhu, H.; Xia, L.; Chai, Z. A bioinformatic analysis: The overexpression and clinical significance of FCGBP in ovarian cancer. Aging 2021, 13, 7416–7429. [Google Scholar] [CrossRef]
- Kobayashi, K.; Blaser, M.; Brown, W.R. Identification of a unique IgG Fc binding site in human intestinal epithelium. J. Immunol. 1989, 143, 2567–2574. [Google Scholar]
- Selbach, M.; Mann, M. Protein interaction screening by quantitative immunoprecipitation combined with knockdown (QUICK). Nat. Methods 2006, 3, 981–983. [Google Scholar] [CrossRef]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 145–153. [Google Scholar] [CrossRef]
- Ferranti, F.; Del Bianco, M.; Pacelli, C. Advantages and limitations of current microgravity platforms for space biology research. Appl. Sci. 2020, 11, 68. [Google Scholar] [CrossRef]
- Brinckmann, E. New facilities and instruments for developmental biology research in space. Adv. Space Biol. Med. 2003, 9, 253–280. [Google Scholar] [PubMed]
- Li, J.; Xu, P.; Wang, L.; Feng, M.; Chen, D.; Yu, X.; Lu, Y. Molecular biology of BPIFB1 and its advances in disease. Ann. Transl. Med. 2020, 8, 651. [Google Scholar] [CrossRef] [PubMed]
- Baines, K.J.; Simpson, J.L.; Wood, L.G.; Scott, R.J.; Fibbens, N.L.; Powell, H.; Cowan, D.C.; Taylor, D.R.; Cowan, J.O.; Gibson, P.G.; et al. Sputum gene expression signature of 6 biomarkers discriminates asthma inflammatory phenotypes. J. Allergy Clin. Immunol. 2014, 133, 997–1007. [Google Scholar] [CrossRef]
- De Smet, E.G.; Seys, L.J.; Verhamme, F.M.; Vanaudenaerde, B.M.; Brusselle, G.G.; Bingle, C.D.; Bracke, K.R. Association of innate defense proteins BPIFA1 and BPIFB1 with disease severity in COPD. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Wu, Y.; Tang, L.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.; Guo, C.; Li, X.; Liao, Q. BPIFB1 (LPLUNC1) inhibits migration and invasion of nasopharyngeal carcinoma by interacting with VTN and VIM. Br. J. Cancer 2018, 118, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Vargas, P.; Speight, P.; Bingle, C.; Barrett, A.; Bingle, L. Expression of PLUNC family members in benign and malignant salivary gland tumours. Oral Dis. 2008, 14, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Zhu, M.; Yin, R.; Shen, W.; Liu, J.; Sun, J.; Wang, C.; Dai, J.; Ma, H.; Wu, C. Low-frequency coding variants at 6p21. 33 and 20q11. 21 are associated with lung cancer risk in Chinese populations. Am. J. Hum. Genet. 2015, 96, 832–840. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Li, Y.; Zhang, R.; Chen, J.; Feng, X.; Duan, Y. Low CFB expression is independently associated with poor overall and diseasefree survival in patients with lung adenocarcinoma. Oncol. Lett. 2021, 21, 478. [Google Scholar] [CrossRef]
- Shimazaki, R.; Takano, S.; Satoh, M.; Takada, M.; Miyahara, Y.; Sasaki, K.; Yoshitomi, H.; Kagawa, S.; Furukawa, K.; Takayashiki, T. Complement factor B regulates cellular senescence and is associated with poor prognosis in pancreatic cancer. Cell. Oncol. 2021, 44, 937–950. [Google Scholar] [CrossRef]
- Conseiller, E.; Debussche, L.; Landais, D.; Venot, C.; Maratrat, M.; Sierra, V.; Tocque, B.; Bracco, L. CTS1: A p53-derived chimeric tumor suppressor gene with enhanced in vitro apoptotic properties. J. Clin. Investig. 1998, 101, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic switch induced by simulated microgravity on MDA-MB-231 breast cancer cells. BioMed Res. Int. 2014, 2014, 652434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitalieri, P.; Marini, M.; Scioli, M.G.; Murdocca, M.; Longo, G.; Orlandi, A.; Novelli, G.; Sangiuolo, F.J.C.; Bioengineering, M. Effects of Simulated Microgravity on Wild Type and Marfan hiPSCs-Derived Embryoid Bodies. Cell. Mol. Bioeng. 2021, 14, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Popper, H. Primary tumor and metastasis—Sectioning the different steps of the metastatic cascade. Transl. Lung Cancer Res. 2020, 9, 2277. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Investig. 2009, 119, 1417–1419. [Google Scholar] [CrossRef] [Green Version]
- Katsuno, Y.; Lamouille, S.; Derynck, R.J. TGF-β signaling and epithelial–mesenchymal transition in cancer progression. Curr. Opin. Oncol. 2013, 25, 76–84. [Google Scholar] [CrossRef]
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: Signaling, therapeutic implications, and challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [Green Version]
- Scheau, C.; Badarau, I.A.; Costache, R.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M.J. The role of matrix metalloproteinases in the epithelial-mesenchymal transition of hepatocellular carcinoma. Anal. Cell. Pathol. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jalaleddine, N.; El-Hajjar, L.; Dakik, H.; Shaito, A.; Saliba, J.; Safi, R.; Zibara, K.; El-Sabban, M.J. Pannexin1 is associated with enhanced epithelial-to-mesenchymal transition in human patient breast cancer tissues and in breast cancer cell lines. Cancers 2019, 11, 1967. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.H.; Gao, B.T.; Goldsmith, Z.K.; Irvine, A.S.; Saleh, N.; Lee, R.P.; Lendermon, J.B.; Bheemreddy, R.; Zhang, Q.; Brennan, R.C. Inhibition of MMP-2 and MMP-9 decreases cellular migration, and angiogenesis in in vitro models of retinoblastoma. BMC Cancer 2017, 17, 434. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Wen, Y.; Miao, X.; Yang, Z.J.C. NT5E and FcGBP as key regulators of TGF-1-induced epithelial–mesenchymal transition (EMT) are associated with tumor progression and survival of patients with gallbladder cancer. Cell Tissue Res. 2014, 355, 365–374. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Lung Cancer Cell Line | Log2 (Fold Change) | Total DEGs between SMG-Exposed Lung Cancer Cells and Controls | Genes Upregulated in SMG | Genes Downregulated in SMG |
|---|---|---|---|---|---|
| GSE36931 | A549 | 4 or −4 | 134 | 47 | 87 |
| GSE78210 | A549 | 3 or −3 | 61 | 21 | 40 |
| GSE78210 | Colo699 | 3 or −3 | 56 | 29 | 27 |
| Geo ID | Title | Lung Cancer Cell Line(s) | Total | Simulated Microgravity Condition | Ground Gravity Condition |
|---|---|---|---|---|---|
| GSE 36931 | Rotating wall vessel (RWV) grown A549s compared with monolayer grown A549s | A549 | 10 | 5 | 5 |
| GSE78210 | 3D cultivation of NSCLC cell lines alters gene expression of key cancer-associated signaling pathways | A549 and Colo699 | 15 | 9 | 6 |
| Total | 2 | 25 | 14 | 11 |
| PRIMER NAME | FORWARD PRIMER | REVERSE PRIMER |
|---|---|---|
| NOX1 | GGAACTCTTGGGGTAGGTGT | GCATCCACAAACAGGAAAAC |
| GDA | AGAGAGTCCCGCTGCGTCT | GGCCAGTTTTTCCTGTTGAG |
| TCN1 | GGTACACTGTTGGAGAGATGA | CGCTGGTTCCCCTGTTATAG |
| FCGBP | AGCCATGGGTGCCCTATGG | GGCTGAGGATGGAGACTGAA |
| BPIFA1 | GGACAGCTGCTGAGACCTC | GGGCTGGATTCACATTCAAG |
| AZGP1 | GCAGACACAATGGTAAGAATGG | AGGTCAGAGAGTAACGACCATC |
| CFB | GGTCTGGAGTTTCAGCTTGG | TCTTGGAGAAGTCGGAAGGA |
| VTCN1 | TCAGCCAGTACCCAGATACG | TGCTCCAGCCAGAATAATGA |
| AGR3 | GGCCAAGTCAGCTTCTTCTG | GTCATCTCCCCATCCTCTTG |
| CTS1 | CAGCTTTGTGCTCTGCCTCT | CACGCTGTACCCACTCATCA |
| F5 | GAGCAGGAAAGGAAGCATGT | TCGGTAGCTCCAACTGATGC |
| CEACAM6 | CAGAAGGAGGAAGGACAGCA | CACCACTGCCAAGCTCACTA |
| BPIFB1 | GTCTGGCATCCTGCACTTG | GACTGAGGGTGGCTTGGAT |
| E-Cadherin | CAGAAAGTTTTCCACCAAAG | AAATGTGAGCAATTCTGCTT |
| N-Cadherin | GGTGGAGGAGAAGAAGAAGACCAG | GGCATCAGGCTCCACAGT |
| Snail | CTTCCAGCAGCCCTACGAC | CGGTGGGGTTGAGGATCT |
| β-catenin | AGGGATTTTCTCAGTCCTTC | CATGCCCTCATCTAATGTCT |
| ZO-1 | CAGCCGGTCACGATCTCCT | TCCGGAGACTGCCATTGC |
| MMP-9 | TTGACAGCGACAAGAAGAAGTGG | GCCATTCACGTCGTCCTTAT |
| MMP-2 | TTGACGGTAAGGACGGACTC | ACTTGCAGTACTCCCATCG |
| GAPDH | TGGTGCTCAGTGTAGCCCAG | GGACCTGACCTGCCGTCTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baghoum, H.; Alahmed, H.; Hachim, M.; Senok, A.; Jalaleddine, N.; Al Heialy, S. Simulated Microgravity Influences Immunity-Related Biomarkers in Lung Cancer. Int. J. Mol. Sci. 2023, 24, 155. https://doi.org/10.3390/ijms24010155
Baghoum H, Alahmed H, Hachim M, Senok A, Jalaleddine N, Al Heialy S. Simulated Microgravity Influences Immunity-Related Biomarkers in Lung Cancer. International Journal of Molecular Sciences. 2023; 24(1):155. https://doi.org/10.3390/ijms24010155
Chicago/Turabian StyleBaghoum, Hend, Hend Alahmed, Mahmood Hachim, Abiola Senok, Nour Jalaleddine, and Saba Al Heialy. 2023. "Simulated Microgravity Influences Immunity-Related Biomarkers in Lung Cancer" International Journal of Molecular Sciences 24, no. 1: 155. https://doi.org/10.3390/ijms24010155