

Methods for the Assessment of NET Formation: From Neutrophil Biology to Translational Research
Abstract
:1. Introduction
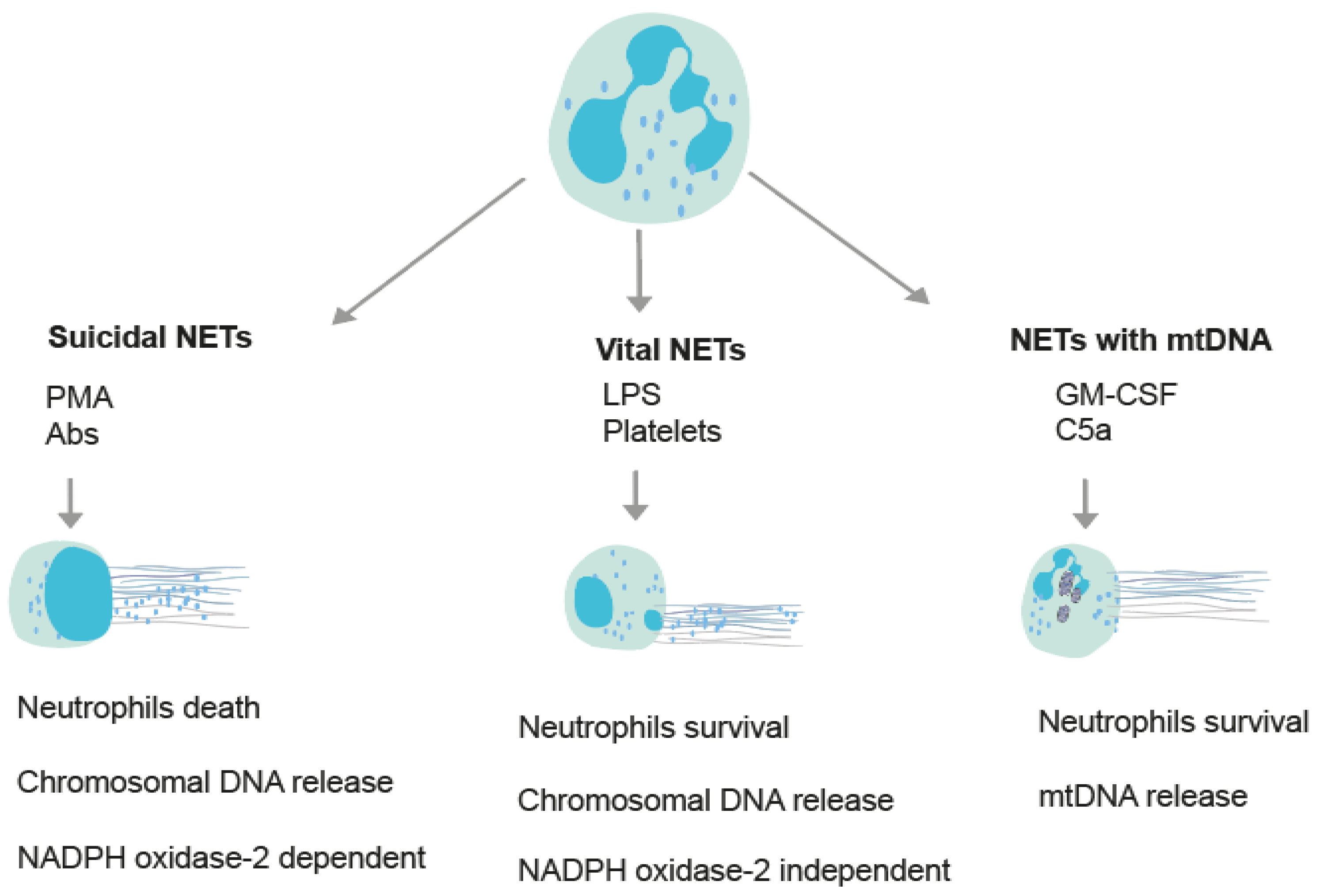
2. Tools for the Assessment of NETs
2.1. Enzyme-Linked Immunosorbent Assay (ELISA)
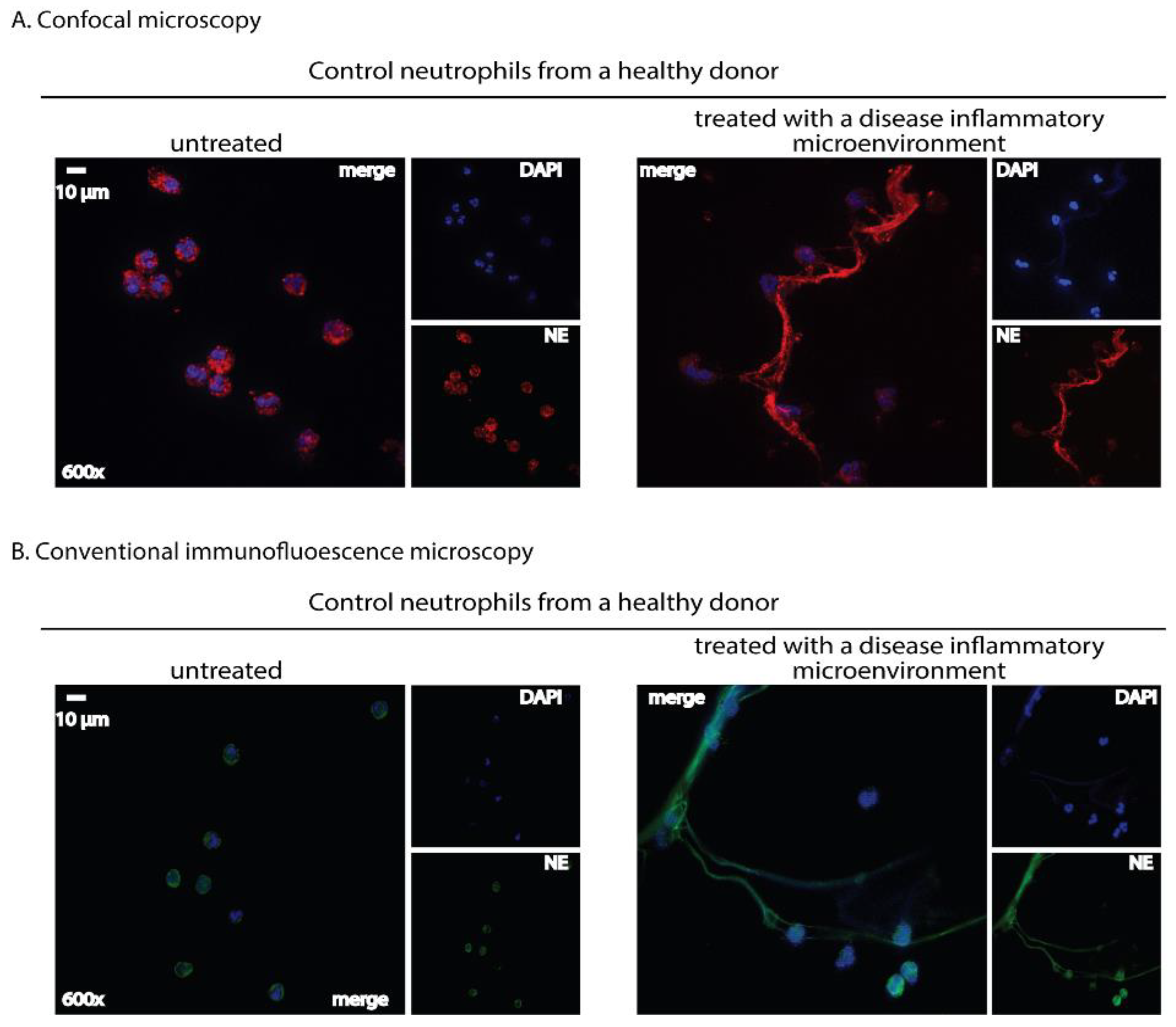
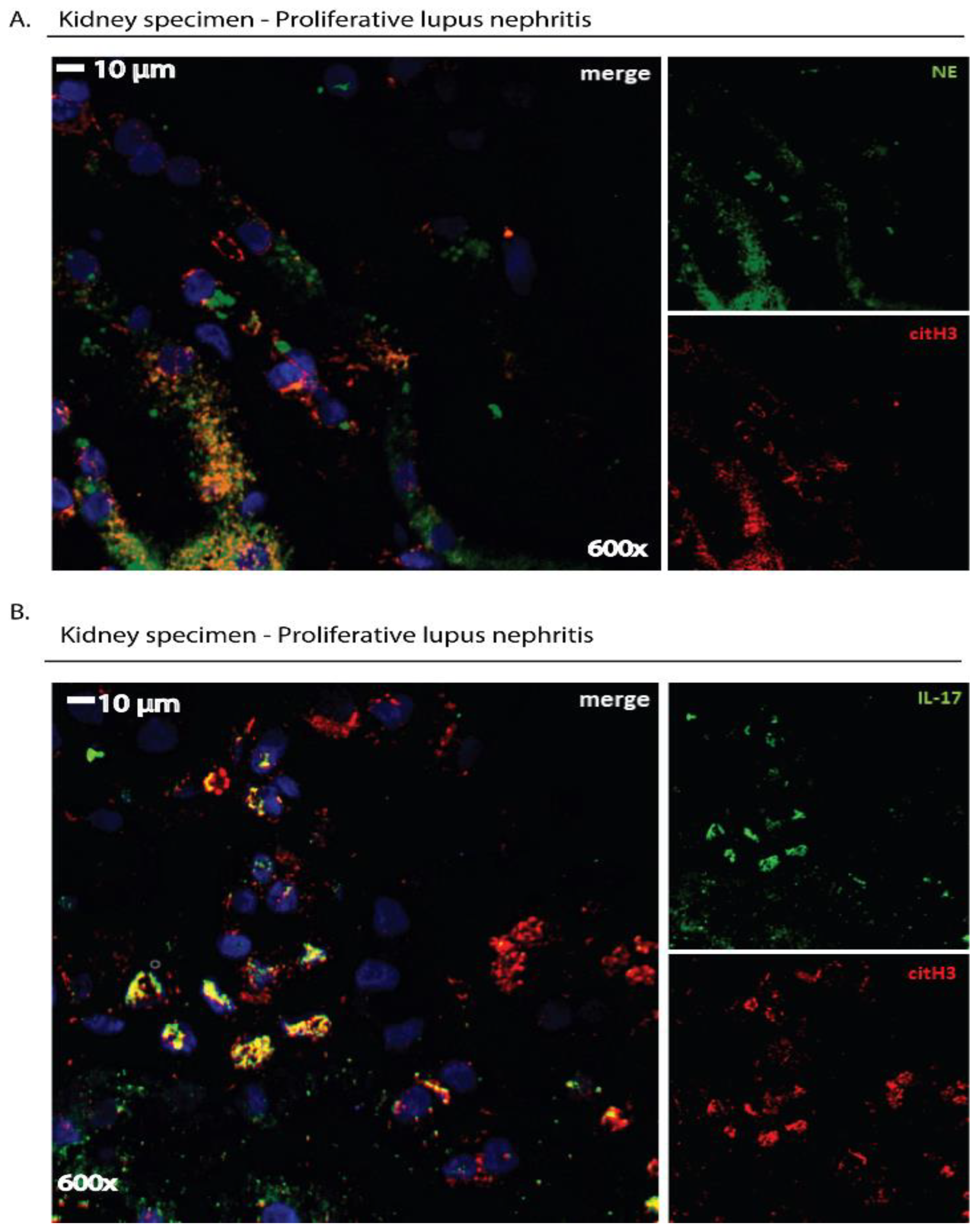
2.2. Immunofluorescence Microscopy
2.3. Electron Microscopy
2.4. Live Imaging
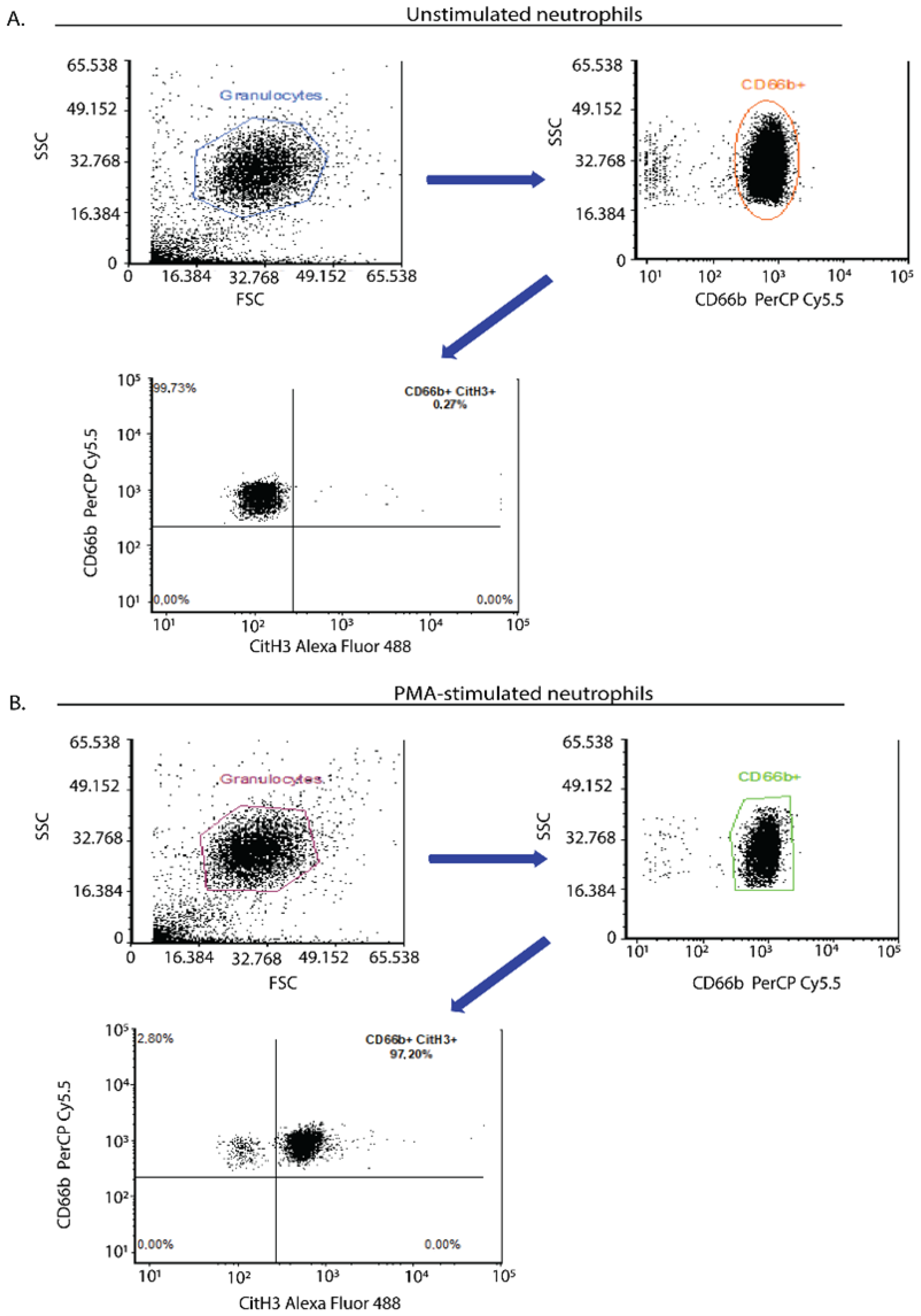
2.5. Flow Cytometry
2.6. Multispectral Image Flow Cytometry (MIFC)
2.7. Western Blot
2.8. DNA Dyes
3. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowland, J.B.; Borregaard, N. Granulopoiesis and Granules of Human Neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Marchal, G.; Bosmans, H.; Van Hecke, P.; Speck, U.; Aerts, P.; Vanhoenacker, P.; Baert, A.L. MR Angiography with Gadopentetate Dimeglumine-Polylysine: Evaluation in Rabbits. Am. J. Roentgenol. 1990, 155, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Heit, B. Armed for Destruction: Formation, Function and Trafficking of Neutrophil Granules. Cell Tissue Res. 2018, 371, 455–471. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, S.M.; Thieblemont, N.; Witko-Sarsat, V. Expanding Neutrophil Horizons: New Concepts in Inflammation. J. Innate Immun. 2018, 10, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Silvestre-Roig, C.; Fridlender, Z.G.; Glogauer, M.; Scapini, P. Neutrophil Diversity in Health and Disease. Trends Immunol. 2019, 40, 565–583. [Google Scholar] [CrossRef]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role during Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef]
- De Bont, C.M.; Boelens, W.C.; Pruijn, G.J.M. NETosis, Complement, and Coagulation: A Triangular Relationship. Cell. Mol. Immunol. 2019, 16, 19–27. [Google Scholar] [CrossRef]
- Tecchio, C.; Cassatella, M.A. Neutrophil-Derived Chemokines on the Road to Immunity. Semin. Immunol. 2016, 28, 119–128. [Google Scholar] [CrossRef]
- Greenlee-Wacker, M.C. Clearance of Apoptotic Neutrophils and Resolution of Inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Boeltz, S.; Amini, P.; Anders, H.-J.; Andrade, F.; Bilyy, R.; Chatfield, S.; Cichon, I.; Clancy, D.M.; Desai, J.; Dumych, T.; et al. To NET or Not to NET:Current Opinions and State of the Science Regarding the Formation of Neutrophil Extracellular Traps. Cell Death Differ. 2019, 26, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable Neutrophils Release Mitochondrial DNA to Form Neutrophil Extracellular Traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochemistry 2020, 85, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- De Buhr, N.; Von Köckritz-Blickwede, M. How Neutrophil Extracellular Traps Become Visible. J. Immunol. Res. 2016, 2016, 4604713. [Google Scholar] [CrossRef] [Green Version]
- Mitsios, A.; Arampatzioglou, A.; Arelaki, S.; Mitroulis, I.; Ritis, K. NETopathies? Unraveling the Dark Side of Old Diseases through Neutrophils. Front. Immunol. 2017, 7, 678. [Google Scholar] [CrossRef]
- Nauseef, W.M.; Borregaard, N. Neutrophils at Work. Nat. Immunol. 2014, 15, 602–611. [Google Scholar] [CrossRef]
- Remijsen, Q.; Berghe, T.V.; Wirawan, E.; Asselbergh, B.; Parthoens, E.; De Rycke, R.; Noppen, S.; Delforge, M.; Willems, J.; Vandenabeele, P. Neutrophil Extracellular Trap Cell Death Requires Both Autophagy and Superoxide Generation. Cell Res. 2011, 21, 290–304. [Google Scholar] [CrossRef] [Green Version]
- Vorobjeva, N.V.; Pinegin, B.V. Neutrophil Extracellular Traps: Mechanisms of Formation and Role in Health and Disease. Biochemistry 2014, 79, 1286–1296. [Google Scholar] [CrossRef]
- Brinkmann, V.; Zychlinsky, A. Neutrophil Extracellular Traps: Is Immunity the Second Function of Chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.; Kienhöfer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated Neutrophil Extracellular Traps Limit Inflammation by Degrading Cytokines and Chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Petretto, A.; Bruschi, M.; Pratesi, F.; Croia, C.; Candiano, G.; Ghiggeri, G.; Migliorini, P. Neutrophil Extracellular Traps (NET) Induced by Different Stimuli: A Comparative Proteomic Analysis. PLoS ONE 2019, 14, e0218946. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.S.; Carmona-Rivera, C.; Kaplan, M.J. Proteins Derived from Neutrophil Extracellular Traps May Serve as Self-Antigens and Mediate Organ Damage in Autoimmune Diseases. Front. Immunol. 2012, 3, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA Traps Promote Thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885. [Google Scholar] [CrossRef] [Green Version]
- Maueröder, C.; Kienhöfer, D.; Hahn, J.; Schauer, C.; Manger, B.; Schett, G.; Herrmann, M.; Hoffmann, M.H. How Neutrophil Extracellular Traps Orchestrate the Local Immune Response in Gout. J. Mol. Med. 2015, 93, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Chrysanthopoulou, A.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Mikroulis, D.; Konstantinidis, T.; Sivridis, E.; Koffa, M.; Giatromanolaki, A.; Boumpas, D.T.; et al. Neutrophil Extracellular Traps Promote Differentiation and Function of Fibroblasts. J. Pathol. 2014, 233, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Thålin, C.; Lundström, S.; Seignez, C.; Daleskog, M.; Lundström, A.; Henriksson, P.; Helleday, T.; Phillipson, M.; Wallén, H.; Demers, M. Citrullinated Histone H3 as a Novel Prognostic Blood Marker in Patients with Advanced Cancer. PLoS ONE 2018, 13, e0191231. [Google Scholar] [CrossRef]
- Kambas, K.; Chrysanthopoulou, A.; Vassilopoulos, D.; Apostolidou, E.; Skendros, P.; Girod, A.; Arelaki, S.; Froudarakis, M.; Nakopoulou, L.; Giatromanolaki, A.; et al. Tissue Factor Expression in Neutrophil Extracellular Traps and Neutrophil Derived Microparticles in Antineutrophil Cytoplasmic Antibody Associated Vasculitis May Promote Thromboinflammation and the Thrombophilic State Associated with the Disease. Ann. Rheum. Dis. 2014, 73, 1854–1863. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Krumbholz, M.; Schönermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Gröne, H.-J.; Brinkmann, V.; Jenne, D.E. Netting Neutrophils in Autoimmune Small-Vessel Vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef]
- Khandpur, R.; Carmona-rivera, C.; Vivekanandan-giri, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; Thompson, P.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra40. [Google Scholar] [CrossRef] [PubMed]
- Angelidou, I.; Chrysanthopoulou, A.; Mitsios, A.; Arelaki, S.; Arampatzioglou, A.; Kambas, K.; Ritis, D.; Tsironidou, V.; Moschos, I.; Dalla, V.; et al. REDD1/Autophagy Pathway Is Associated with Neutrophil-Driven IL-1β Inflammatory Response in Active Ulcerative Colitis. J. Immunol. 2018, 200, 3950–3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangou, E.; Chrysanthopoulou, A.; Mitsios, A.; Kambas, K.; Arelaki, S.; Angelidou, I.; Arampatzioglou, A.; Gakiopoulou, H.; Bertsias, G.K.; Verginis, P.; et al. REDD1/Autophagy Pathway Promotes Thromboinflammation and Fibrosis in Human Systemic Lupus Erythematosus (SLE) through NETs Decorated with Tissue Factor (TF) and Interleukin-17A (IL-17A). Ann. Rheum. Dis. 2019, 78, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Hakkim, A.; Fürnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of Neutrophil Extracellular Trap Degradation Is Associated with Lupus Nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, E.; Hofstra, J.; Berden, J.; Herrmann, M.; Dieker, J.; van der Vlag, J. Acetylated Histones Contribute to the Immunostimulatory Potential of Neutrophil Extracellular Traps in Systemic Lupus Erythematosus. Clin. Exp. Immunol. 2015, 179, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-hussaniy, H.A.; Al-Harcan, N.A.H.; Alexiou, A.; Batiha, G.E.S. Neutrophil Extracellular Traps (NETs) and COVID-19: A New Frontiers for Therapeutic Modality. Int. Immunopharmacol. 2022, 104, 108516. [Google Scholar] [CrossRef] [PubMed]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and Tissue Factor–Enriched Neutrophil Extracellular Traps Are Key Drivers in COVID-19 Immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Aldabbous, L.; Abdul-Salam, V.; Mckinnon, T.; Duluc, L.; Pepke-Zaba, J.; Southwood, M.; Ainscough, A.J.; Hadinnapola, C.; Wilkins, M.R.; Toshner, M.; et al. Neutrophil Extracellular Traps Promote Angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2078–2087. [Google Scholar] [CrossRef] [Green Version]
- Chrysanthopoulou, A.; Gkaliagkousi, E.; Lazaridis, A.; Arelaki, S.; Pateinakis, P.; Ntinopoulou, M.; Mitsios, A.; Antoniadou, C.; Argyriou, C.; Georgiadis, G.S.; et al. Angiotensin II Triggers Release of Neutrophil Extracellular Traps, Linking Thromboinflammation with Essential Hypertension. JCI Insight 2021, 6, e148668. [Google Scholar] [CrossRef]
- Yadav, R.; Yoo, D.G.; Kahlenberg, J.M.; Bridges, S.L.; Oni, O.; Huang, H.; Stecenko, A.; Rada, B. Systemic Levels of Anti-PAD4 Autoantibodies Correlate with Airway Obstruction in Cystic Fibrosis. J. Cyst. Fibros. 2019, 18, 636–645. [Google Scholar] [CrossRef]
- Yoo, D.; Winn, M.; Pang, L.; Moskowitz, S.M.; Malech, H.L.; Leto, T.L.; Rada, B. Release of Cystic Fibrosis Airway Inflammatory Markers from Pseudomonas Aeruginosa—Stimulated Human Neutrophils Involves NADPH Oxidase-Dependent Extracellular DNA Trap Formation. J. Immunol. 2014, 192, 4728–4738. [Google Scholar] [CrossRef]
- Kasprzycka, W.; Homa-Mlak, I.; Mlak, R.; Malecka-Massalska, T. Direct and Indirect Methods of Evaluating the NETosis Process. J. Pre-Clin. Clin. Res. 2019, 13, 50–56. [Google Scholar] [CrossRef]
- Caudrillier, A.; Kessenbrock, K.; Gilliss, B.M.; Nguyen, J.X.; Marques, M.B.; Monestier, M.; Toy, P.; Werb, Z.; Looney, M.R. Platelets Induce Neutrophil Extracellular Traps in Transfusion-Related Acute Lung Injury. J. Clin. Investig. 2012, 122, 2661–2671. [Google Scholar] [CrossRef]
- Mauracher, L.M.; Posch, F.; Martinod, K.; Grilz, E.; Däullary, T.; Hell, L.; Brostjan, C.; Zielinski, C.; Ay, C.; Wagner, D.D.; et al. Citrullinated Histone H3, a Biomarker of Neutrophil Extracellular Trap Formation, Predicts the Risk of Venous Thromboembolism in Cancer Patients. J. Thromb. Haemost. 2018, 16, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Dicker, A.J.; Crichton, M.L.; Pumphrey, E.G.; Cassidy, A.J.; Suarez-Cuartin, G.; Sibila, O.; Furrie, E.; Fong, C.J.; Ibrahim, W.; Brady, G.; et al. Neutrophil Extracellular Traps Are Associated with Disease Severity and Microbiota Diversity in Patients with Chronic Obstructive Pulmonary Disease. J. Allergy Clin. Immunol. 2018, 141, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thålin, C.; Daleskog, M.; Göransson, S.P.; Schatzberg, D.; Lasselin, J.; Laska, A.-C.; Kallner, A.; Helleday, T.; Wallén, H.; Demers, M. Validation of an Enzyme-Linked Immunosorbent Assay for the Quantification of Citrullinated Histone H3 as a Marker for Neutrophil Extracellular Traps in Human Plasma. Immunol. Res. 2017, 65, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Thålin, C.; Hisada, Y.; Lundström, S.; Mackman, N.; Wallén, H. Neutrophil Extracellular Traps: Villains and Targets in Arterial, Venous, and Cancer-Associated Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1724–1738. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.; Ibrahim, N.; Klopf, J.; Zagrapan, B.; Mauracher, L.M.; Hell, L.; Hofbauer, T.M.; Ondracek, A.S.; Schoergenhofer, C.; Jilma, B.; et al. ELISA Detection of MPO-DNA Complexes in Human Plasma Is Error-Prone and Yields Limited Information on Neutrophil Extracellular Traps Formed in vivo. PLoS ONE 2021, 16, e0250265. [Google Scholar] [CrossRef] [PubMed]
- Rada, B. Neutrophil Extracellular Traps. In NADPH Oxidases; Methods in Molecular Biology; Knaus, U.G., Leto, T.L., Eds.; Springer: New York, NY, USA, 2019; Volume 1982, pp. 517–528. ISBN 978-1-4939-9423-6. [Google Scholar]
- Gavillet, M.; Martinod, K.; Renella, R.; Harris, C.; Shapiro, N.I.; Wagner, D.D.; Williams, D.A. Flow Cytometric Assay for Direct Quantification of Neutrophil Extracellular Traps in Blood Samples. Am. J. Hematol. 2015, 90, 1155–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Köckritz-Blickwede, M.; Chow, O.; Ghochani, M.; Nizet, V. Visualization and Functional Evaluation of Phagocyte Extracellular Traps, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2010; Volume 37. [Google Scholar]
- Brinkmann, V.; Goosmann, C.; Kühn, L.I.; Zychlinsky, A. Automatic Quantification of in Vitro NET Formation. Front. Immunol. 2012, 3, 413. [Google Scholar] [CrossRef] [Green Version]
- Coelho, L.P.; Pato, C.; Friães, A.; Neumann, A.; Von Köckritz-Blickwede, M.; Ramirez, M.; Carriço, J.A. Automatic Determination of NET (Neutrophil Extracellular Traps) Coverage in Fluorescent Microscopy Images. Bioinformatics 2015, 31, 2364–2370. [Google Scholar] [CrossRef]
- Lv, D.; Xu, Y.; Cheng, H.; Ke, Y.; Zhang, X.; Ying, K. A Novel Cell-Based Assay for Dynamically Detecting Neutrophil Extracellular Traps-Induced Lung Epithelial Injuries. Exp. Cell Res. 2020, 394, 112101. [Google Scholar] [CrossRef]
- Inkson, B.J. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) for Materials Characterization; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; ISBN 978-0-08-100057-1. [Google Scholar]
- Rajeeve, K.; Das, S.; Prusty, B.K.; Rudel, T. Chlamydia Trachomatis Paralyses Neutrophils to Evade the Host Innate Immune Response. Nat. Microbiol. 2018, 3, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Jorch, S.K.; Deppermann, C. Intravital Imaging Allows Organ-Specific Insights Into Immune Functions. Front. Cell Dev. Biol. 2021, 9, 623906. [Google Scholar] [CrossRef] [PubMed]
- Hoppenbrouwers, T.; Autar, A.S.A.; Sultan, A.R.; Abraham, T.E.; van Cappellen, W.A.; Houtsmuller, A.B.; van Wamel, W.J.B.; van Beusekom, H.M.M.; van Neck, J.W.; de Maat, M.P.M. In Vitro Induction of NETosis: Comprehensive Live Imaging Comparison and Systematic Review. PLoS ONE 2017, 12, e0176472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adan, A.; Alizada, G.; Kiraz, Y.; Baran, Y.; Nalbant, A. Flow Cytometry: Basic Principles and Applications. Crit. Rev. Biotechnol. 2017, 37, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Shimizu, S.; Matsuo, J.; Nishibata, Y.; Kusunoki, Y.; Hattanda, F.; Shida, H.; Nakazawa, D.; Tomaru, U.; Atsumi, T.; et al. Measurement of NET Formation in Vitro and in Vivo by Flow Cytometry. Cytom. Part A 2017, 91, 822–829. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Rivera, C.; Kaplan, M.J. Induction and Quantification of NETosis. Curr. Protoc. Immunol. 2016, 2016, 14–41. [Google Scholar] [CrossRef]
- Pulavendran, S.; Prasanthi, M.; Ramachandran, A.; Grant, R.; Snider, T.A.; Chow, V.T.K.; Malayer, J.R.; Teluguakula, N. Production of Neutrophil Extracellular Traps Contributes to the Pathogenesis of Francisella Tularemia. Front. Immunol. 2020, 11, 379. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.S.; Bardoel, B.W.; Harbort, C.J.; Zychlinsky, A. Induction and Quantification of Neutrophil Extracellular Traps. Methods Mol. Biol. 2014, 1124, 307–318. [Google Scholar] [CrossRef]
- Stakos, D.A.; Kambas, K.; Konstantinidis, T.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Tsironidou, V.; Giatromanolaki, A.; Skendros, P.; Konstantinides, S.; et al. Expression of Functional Tissue Factor by Neutrophil Extracellular Traps in Culprit Artery of Acute Myocardial Infarction. Eur. Heart J. 2015, 36, 1405–1414. [Google Scholar] [CrossRef]
- Apostolidou, E.; Skendros, P.; Kambas, K.; Mitroulis, I.; Konstantinidis, T.; Chrysanthopoulou, A.; Nakos, K.; Tsironidou, V.; Koffa, M.; Boumpas, D.T.; et al. Neutrophil Extracellular Traps Regulate IL-1β-Mediated Inflammation in Familial Mediterranean Fever. Ann. Rheum. Dis. 2016, 75, 269–277. [Google Scholar] [CrossRef]
- Mitroulis, I.; Kambas, K.; Chrysanthopoulou, A.; Skendros, P.; Apostolidou, E.; Kourtzelis, I.; Drosos, G.I.; Boumpas, D.T.; Ritis, K. Neutrophil Extracellular Trap Formation Is Associated with IL-1β and Autophagy-Related Signaling in Gout. PLoS ONE 2011, 6, e29318. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil Elastase and Myeloperoxidase Regulate the Formation of Neutrophil Extracellular Traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone Hypercitrullination Mediates Chromatin Decondensation and Neutrophil Extracellular Trap Formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilley, D.O.; Abuabed, U.; Zimny Arndt, U.; Schmid, M.; Florian, S.; Jungblut, P.R.; Brinkmann, V.; Herzig, A.; Zychlinsky, A. Histone H3 Clipping Is a Novel Signature of Human Neutrophil Extracellular Traps. eLife 2022, 11, e68283. [Google Scholar] [CrossRef]
- Gordon, R.E. Electron Microscopy: A Brief History and Review of Current Clinical Application. Methods Mol. Biol. 2014, 1180, 119–135. [Google Scholar] [CrossRef]
- Condello, M.; Caraglia, M.; Castellano, M.; Arancia, G.; Meschini, S. Structural and Functional Alterations of Cellular Components as Revealed by Electron Microscopy. Microsc. Res. Tech. 2013, 76, 1057–1069. [Google Scholar] [CrossRef]
- Krautgartner, W.D.; Vitkov, L. Visualization of Neutrophil Extracellular Traps in TEM. Micron 2008, 39, 367–372. [Google Scholar] [CrossRef]
- Skendros, P.; Mitroulis, I.; Ritis, K. Autophagy in Neutrophils: From Granulopoiesis to Neutrophil Extracellular Traps. Front. Cell Dev. Biol. 2018, 6, 109. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, Y.; Yin, S.W.; Gao, X.J.; Shi, W.W.; Wang, Y.; Huang, X.; Wang, L.; Zou, L.Y.; Zhao, J.H.; et al. Neutrophil Extracellular Trap Formation Is Associated with Autophagy-Related Signalling in ANCA-Associated Vasculitis. Clin. Exp. Immunol. 2015, 180, 408–418. [Google Scholar] [CrossRef]
- Ermert, D.; Urban, C.F.; Laube, B.; Goosmann, C.; Zychlinsky, A.; Brinkmann, V. Mouse Neutrophil Extracellular Traps in Microbial Infections. J. Innate Immun. 2009, 1, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Manzenreiter, R.; Kienberger, F.; Marcos, V.; Schilcher, K.; Krautgartner, W.D.; Obermayer, A.; Huml, M.; Stoiber, W.; Hector, A.; Griese, M.; et al. Ultrastructural Characterization of Cystic Fibrosis Sputum Using Atomic Force and Scanning Electron Microscopy. J. Cyst. Fibros. 2012, 11, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Chun, H.J.; Kim, E.S.; Cho, B.R. Multiphoton Microscopy: An Introduction to Gastroenterologists. World J. Gastroenterol. 2011, 17, 4456–4460. [Google Scholar] [CrossRef]
- Elliott, A.D. Confocal Microscopy: Principles and Modern Practices. Curr. Protoc. Cytom. 2020, 92, e68. [Google Scholar] [CrossRef]
- Halpern, A.R.; Lee, M.Y.; Howard, M.D.; Woodworth, M.A.; Nicovich, P.R.; Vaughan, J.C. Versatile, Do-It-Yourself, Low-Cost Spinning Disk Confocal Microscope. Biomed. Opt. Express 2022, 13, 1102. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Jenne, C.N.; Surewaard, B.G.J.; Thanabalasuriar, A.; Lee, W.-Y.; Sanz, M.-J.; Mowen, K.; Opdenakker, G.; Kubes, P. Molecular Mechanisms of NET Formation and Degradation Revealed by Intravital Imaging in the Liver Vasculature. Nat. Commun. 2015, 6, 6673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, L.; Yu, H.; Yang, X.; Zhu, Y.; Bai, X.; Wang, R.; Cao, Y.; Xu, H.; Luo, H.; Lu, L.; et al. Neutrophil Extracellular Traps Released by Neutrophils Impair Revascularization and Vascular Remodeling after Stroke. Nat. Commun. 2020, 11, 2488. [Google Scholar] [CrossRef]
- Cichon, I.; Ortmann, W.; Santocki, M.; Opydo-Chanek, M.; Kolaczkowska, E. Scrutinizing Mechanisms of the ‘Obesity Paradox in Sepsis’: Obesity Is Accompanied by Diminished Formation of Neutrophil Extracellular Traps (NETs) Due to Restricted Neutrophil–Platelet Interactions. Cells 2021, 10, 384. [Google Scholar] [CrossRef]
- Jenne, C.N.; Wong, C.H.Y.; Petri, B.; Kubes, P. The Use of Spinning-Disk Confocal Microscopy for the Intravital Analysis of Platelet Dynamics in Response to Systemic and Local Inflammation. PLoS ONE 2011, 6, e25109. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Chan, D.W.; Zaal, K.J.; Kaplan, M.J. A High-Throughput Real-Time Imaging Technique To Quantify NETosis and Distinguish Mechanisms of Cell Death in Human Neutrophils. J. Immunol. 2018, 200, 869–879. [Google Scholar] [CrossRef]
- Zharkova, O.; Tay, S.H.; Lee, H.Y.; Shubhita, T.; Ong, W.Y.; Lateef, A.; MacAry, P.A.; Lim, L.H.K.; Connolly, J.E.; Fairhurst, A.M. A Flow Cytometry-Based Assay for High-Throughput Detection and Quantification of Neutrophil Extracellular Traps in Mixed Cell Populations. Cytom. Part A 2019, 95, 268–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, S.; Nakazawa, D.; Shida, H.; Miyoshi, A.; Kusunoki, Y.; Tomaru, U.; Ishizu, A. NETosis Markers: Quest for Specific, Objective, and Quantitative Markers. Clin. Chim. Acta 2016, 459, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Fogg, D.K.; Kaplan, M.J. A Novel Image-Based Quantitative Method for the Characterization of NETosis. J. Immunol. Methods 2015, 423, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ma, H. Western Blotting and ELISA Techniques. Researcher 2009, 1, 67–86. [Google Scholar]
- Liu, S.; Su, X.; Pan, P.; Zhang, L.; Hu, Y.; Tan, H.; Wu, D.; Liu, B.; Li, H.; Li, H.; et al. Neutrophil Extracellular Traps Are Indirectly Triggered by Lipopolysaccharide and Contribute to Acute Lung Injury. Sci. Rep. 2016, 6, 37252. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.; Gilda, J.E.; Gomes, A.V. Accuracy of Western Blots. Expert Rev. Proteom. 2014, 11, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Yost, C.C.; Cody, M.J.; Harris, E.S.; Thornton, N.L.; McInturff, A.M.; Martinez, M.L.; Chandler, N.B.; Rodesch, C.K.; Albertine, K.H.; Petti, C.A.; et al. Impaired Neutrophil Extracellular Trap (NET) Formation: A Novel Innate Immune Deficiency of Human Neonates. Blood 2009, 113, 6419–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopke, A.; Scherer, A.; Kreuzburg, S.; Abers, M.S.; Zerbe, C.S.; Dinauer, M.C.; Mansour, M.K.; Irimia, D. Neutrophil Swarming Delays the Growth of Clusters of Pathogenic Fungi. Nat. Commun. 2020, 11, 2031. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Techniques | Benefits | Limitations | References |
|---|---|---|---|
| Enzyme-linked immunosorbent assay (ELISA) | Simplicity, specificity, cost-effectiveness, objective and quantitative | Results standardization and differences between in vivo and in vitro NETs | Kasprzycka et al., 2019 [41]; Thålin et al., 2017 [45]; Masuda et al., 2017 [49] |
| Immunofluorescence microscopy (IFM) | Distinguishes cell death mechanisms, objective quantification of NETs with software-based methods, and detects NETs in vitro, ex vivo and in situ | A time-consuming procedure, laborious, limited reproducibility, not available for rapid screening of many cells or samples, observer-dependent, and extensive precaution with sample preparation | Brinkmann et al., 2004 [11]; Von Köckritz-Blickwede et al., 2010 [50]; Brinkmann et al., 2012 [51]; Coelho et al., 2015 [52]; De Buhr and von Köckritz-Blickwede, 2016 [15]; Lv et al., 2020 [53] |
| Electron microscopy | Distinguishes cell death mechanisms and detects NETs in vitro and in situ | Biased based on the field of view and no reliable distinguishing between NETs and fibrin by SEM | Fuchs et al., 2007 [24]; De Buhr and von Köckritz-Blickwede, 2016 [15]; Krautgartner et al., 2008 [54]; Lv et al., 2020 [53] |
| Live imaging | Live in vivo imaging using animal models, close examination of NET structures, and distinguishes NETs from other cell deaths | Staining of many components on NETs, complex equipment and expensive | Cho et al., 2011 [55]; Kolaczkowska et al., 2015 [56]; Gupta et al., 2018 [57]; |
| Flow cytometry | Qualitative, quantitative and objective; detects pre-NETotic cells, both in vivo and in vitro, rapid and sort between cell populations and has the possibility of direct applications in blood samples | Extensive precaution with sample preparation; detects only pre-NETotic cells and is incapable of detecting released NETs/remnants | Masuda et al., 2017 [49]; Kasprzycka et al., 2019 [41]; Gavillet et al., 2015 [58]; Zharkova et al., 2019 [59] |
| Multispectral image flow cytometer (MIFC) | Rapid, semiautomated and studies subcellular morphology and distinguishes cell death mechanisms | Sophisticated equipment and detects fewer NETotic cells than existing | Zhao et al., 2015 [60] |
| Western blot | Semi-quantitative, specificity and sensitivity | Multi-step protocol and optimizes the concentration of the primary antibody | Liu et al., 2016 [61]; Lv et al., 2020 [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoimenou, M.; Tzoros, G.; Skendros, P.; Chrysanthopoulou, A. Methods for the Assessment of NET Formation: From Neutrophil Biology to Translational Research. Int. J. Mol. Sci. 2022, 23, 15823. https://doi.org/10.3390/ijms232415823
Stoimenou M, Tzoros G, Skendros P, Chrysanthopoulou A. Methods for the Assessment of NET Formation: From Neutrophil Biology to Translational Research. International Journal of Molecular Sciences. 2022; 23(24):15823. https://doi.org/10.3390/ijms232415823
Chicago/Turabian StyleStoimenou, Marina, Georgios Tzoros, Panagiotis Skendros, and Akrivi Chrysanthopoulou. 2022. "Methods for the Assessment of NET Formation: From Neutrophil Biology to Translational Research" International Journal of Molecular Sciences 23, no. 24: 15823. https://doi.org/10.3390/ijms232415823