
Human Dental Pulp Stem Cells Differentiate into Cementoid-Like-Secreting Cells on Decellularized Teeth Scaffolds
 , and
, and 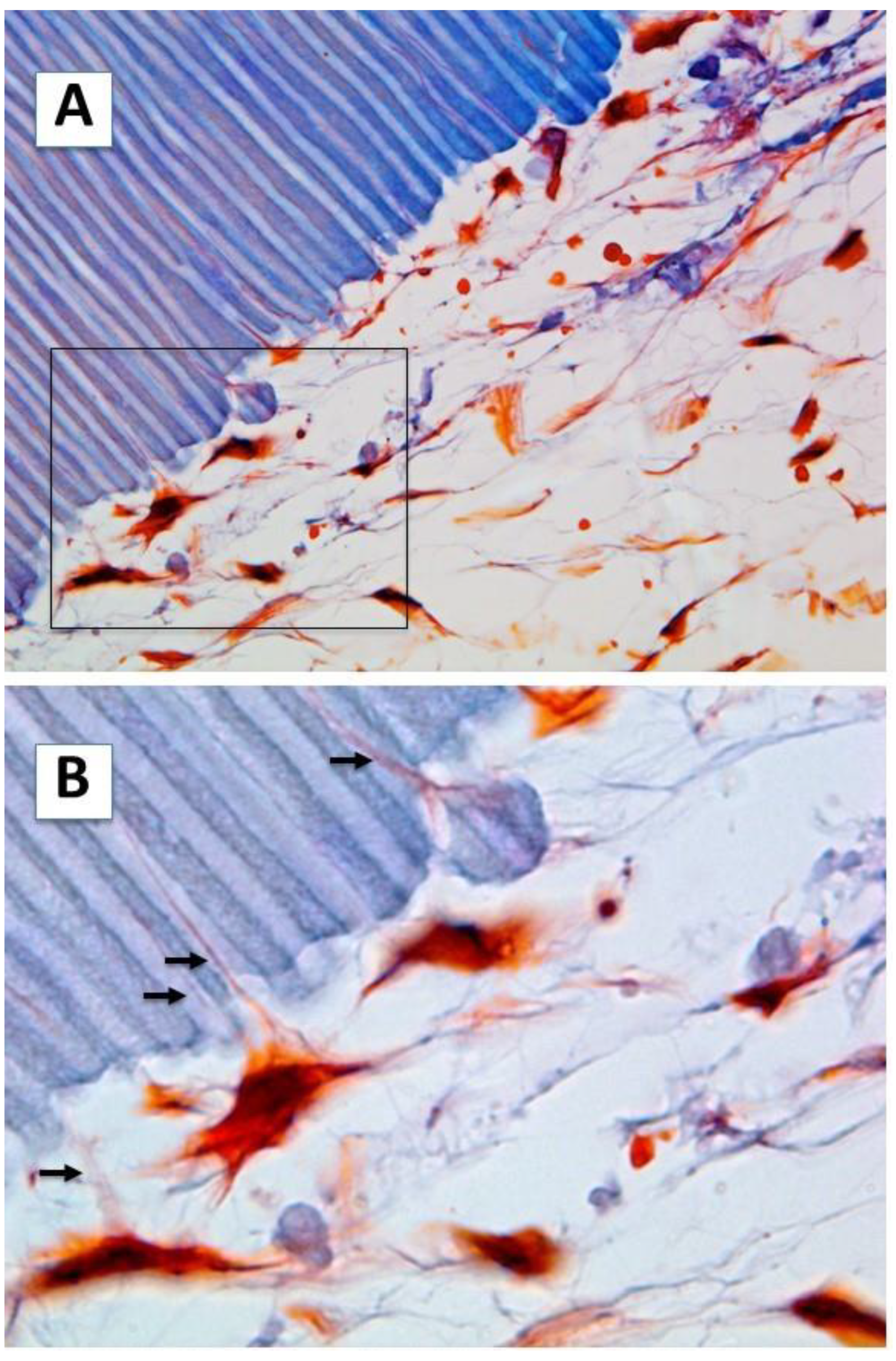{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
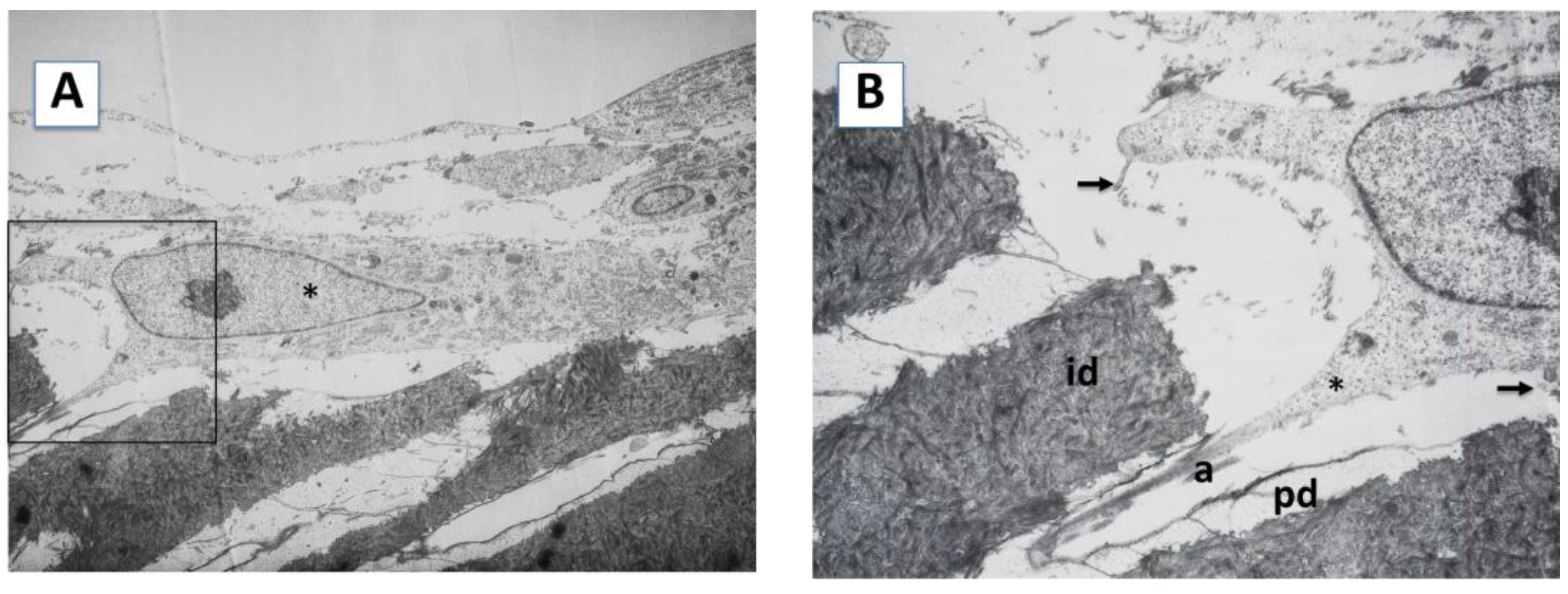
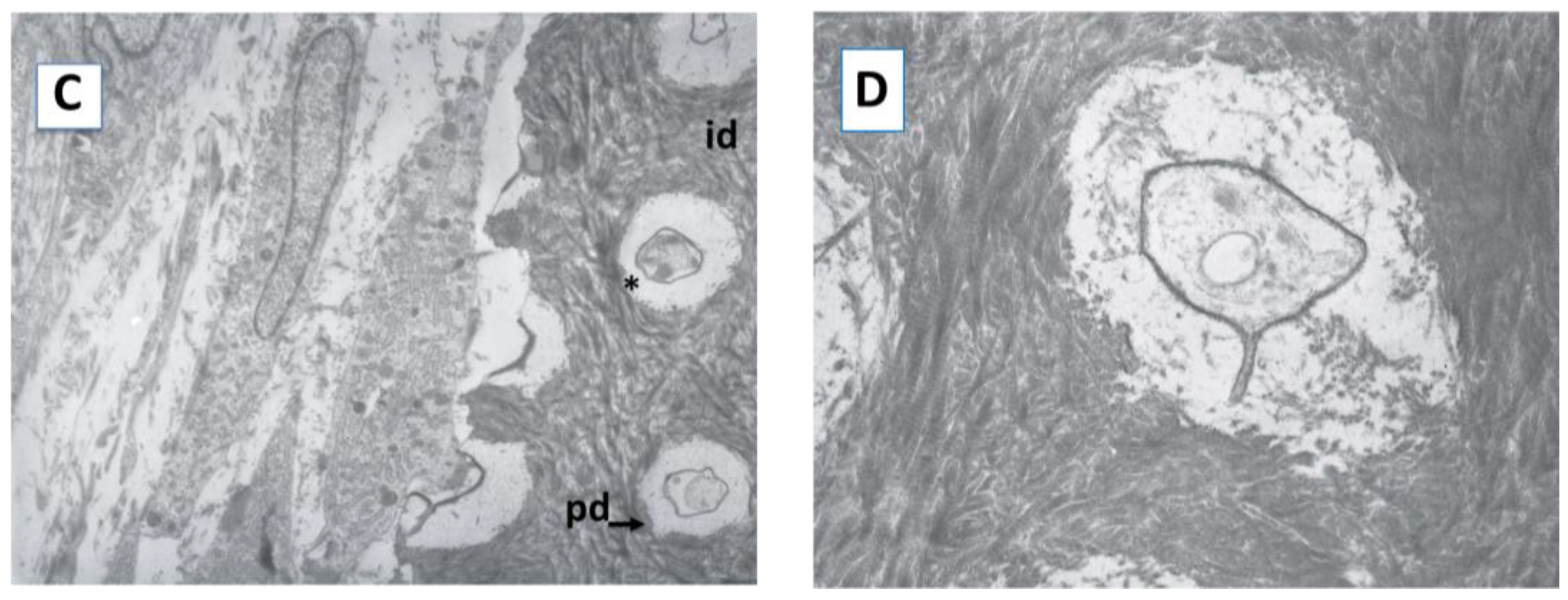
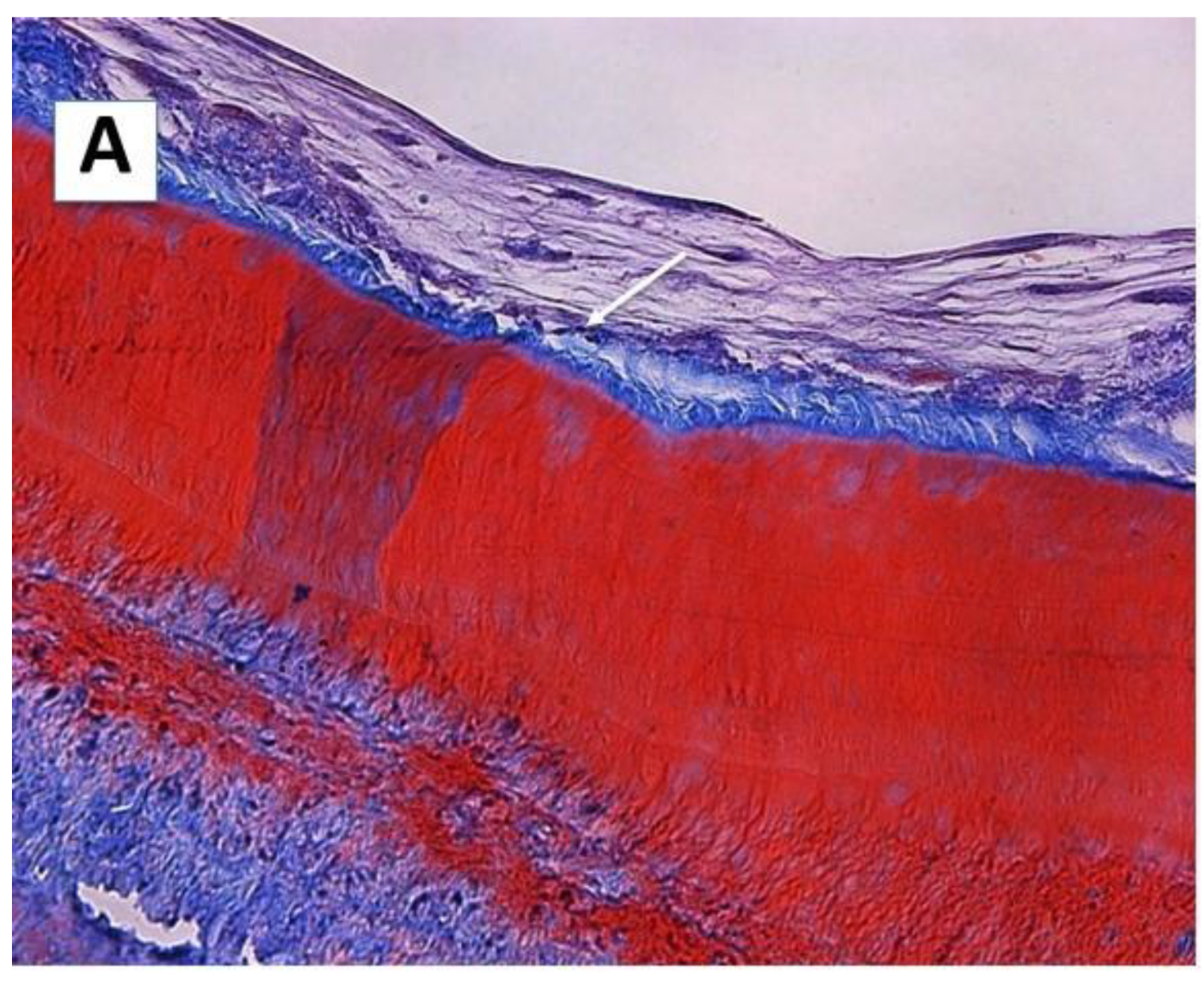
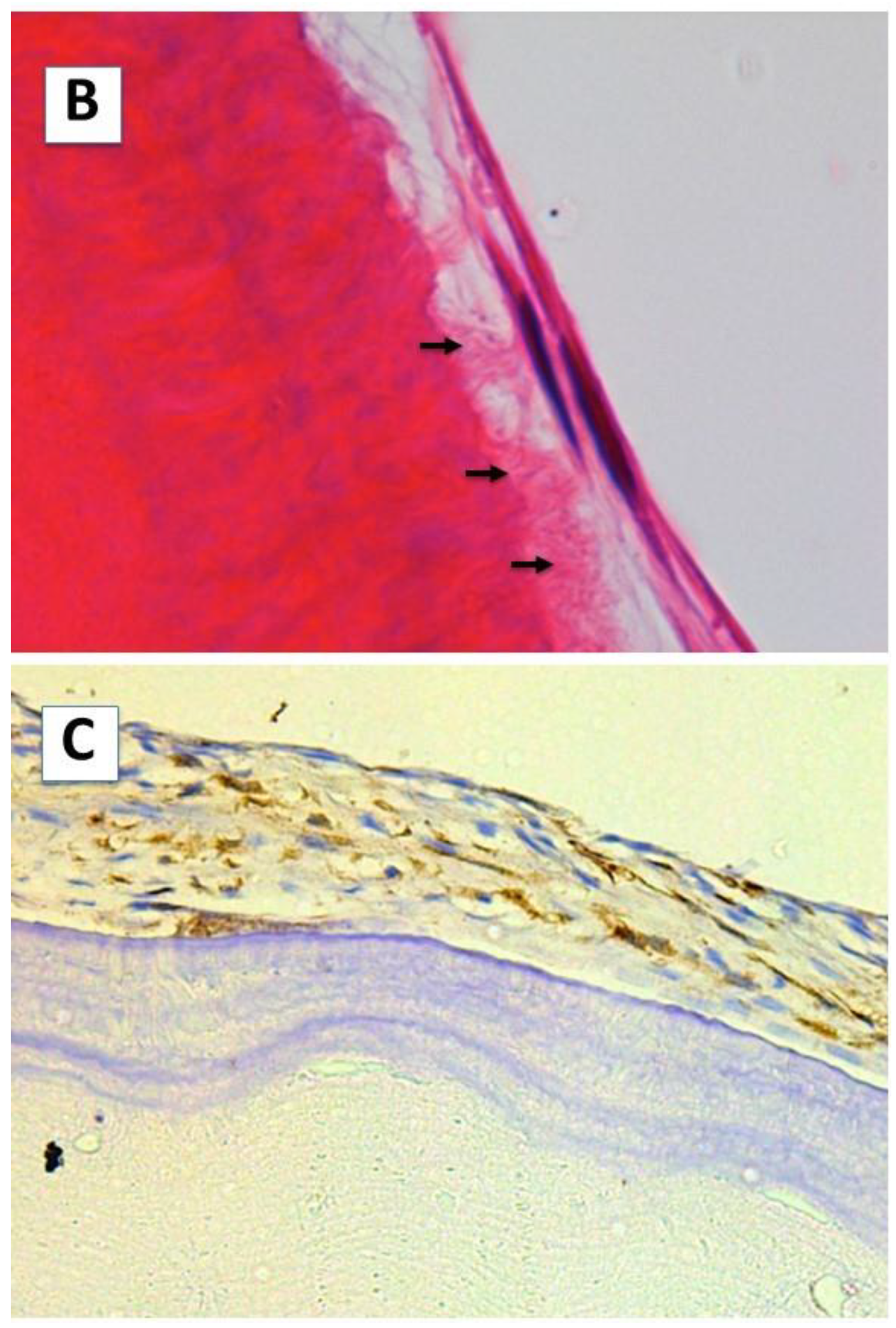
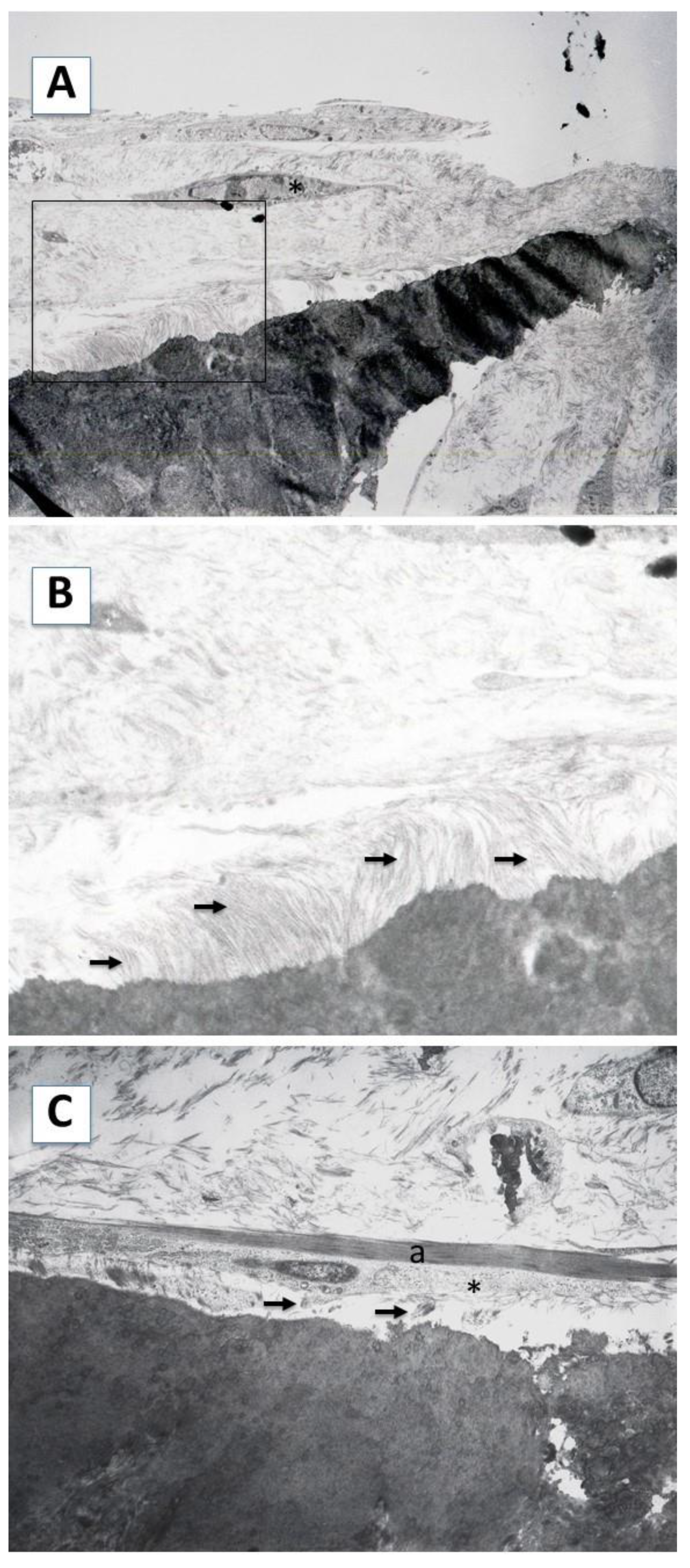
2.1. Cell Morphology and Extracellular Matrix Characteristics of hDPSCs Cultured on Surface A (SA)
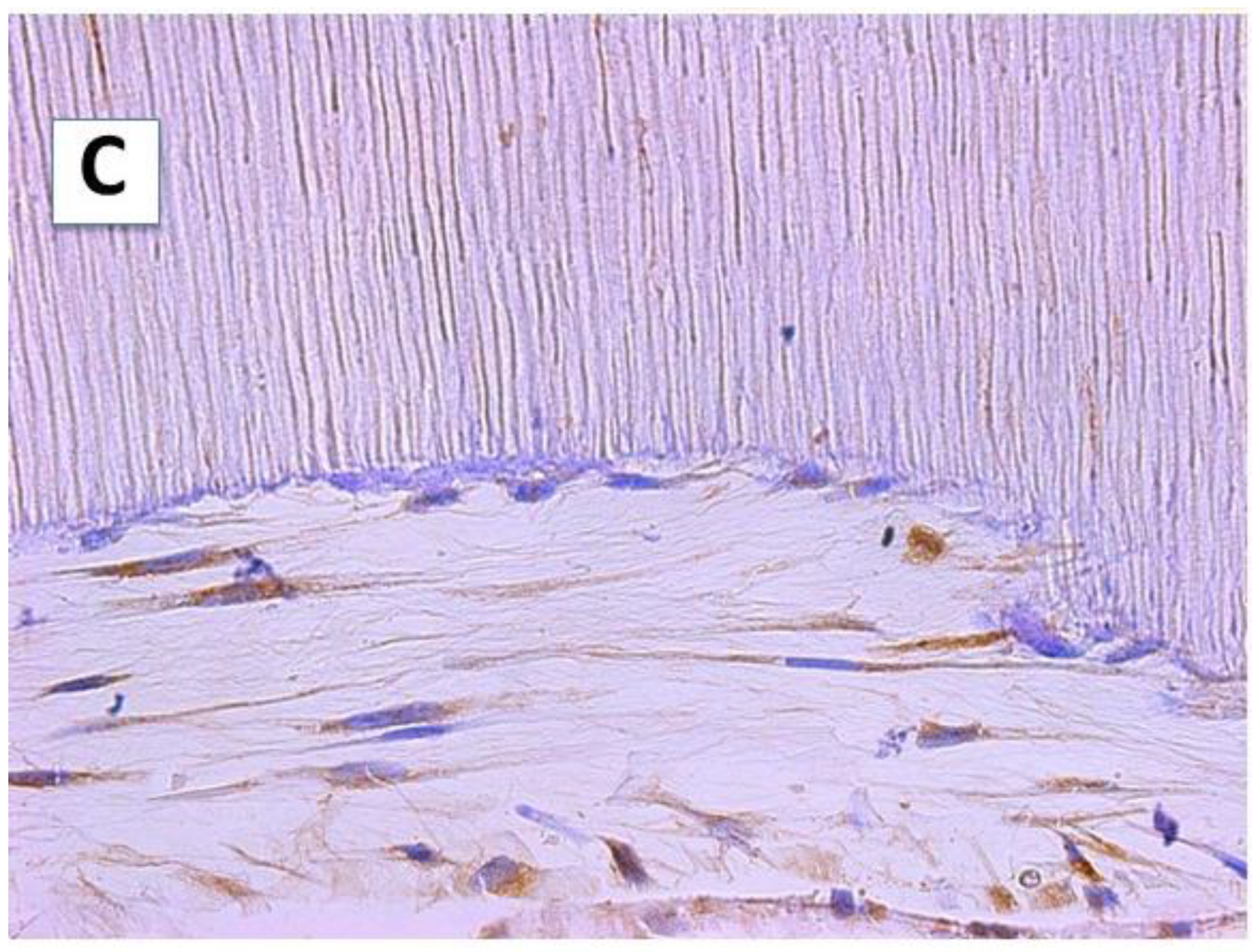
2.2. Cellular Morphology and Extracellular Matrix Characteristics of hDPSCs Cultured on Surface B (SB)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Flow Cytometric Characterization of hDPSCs
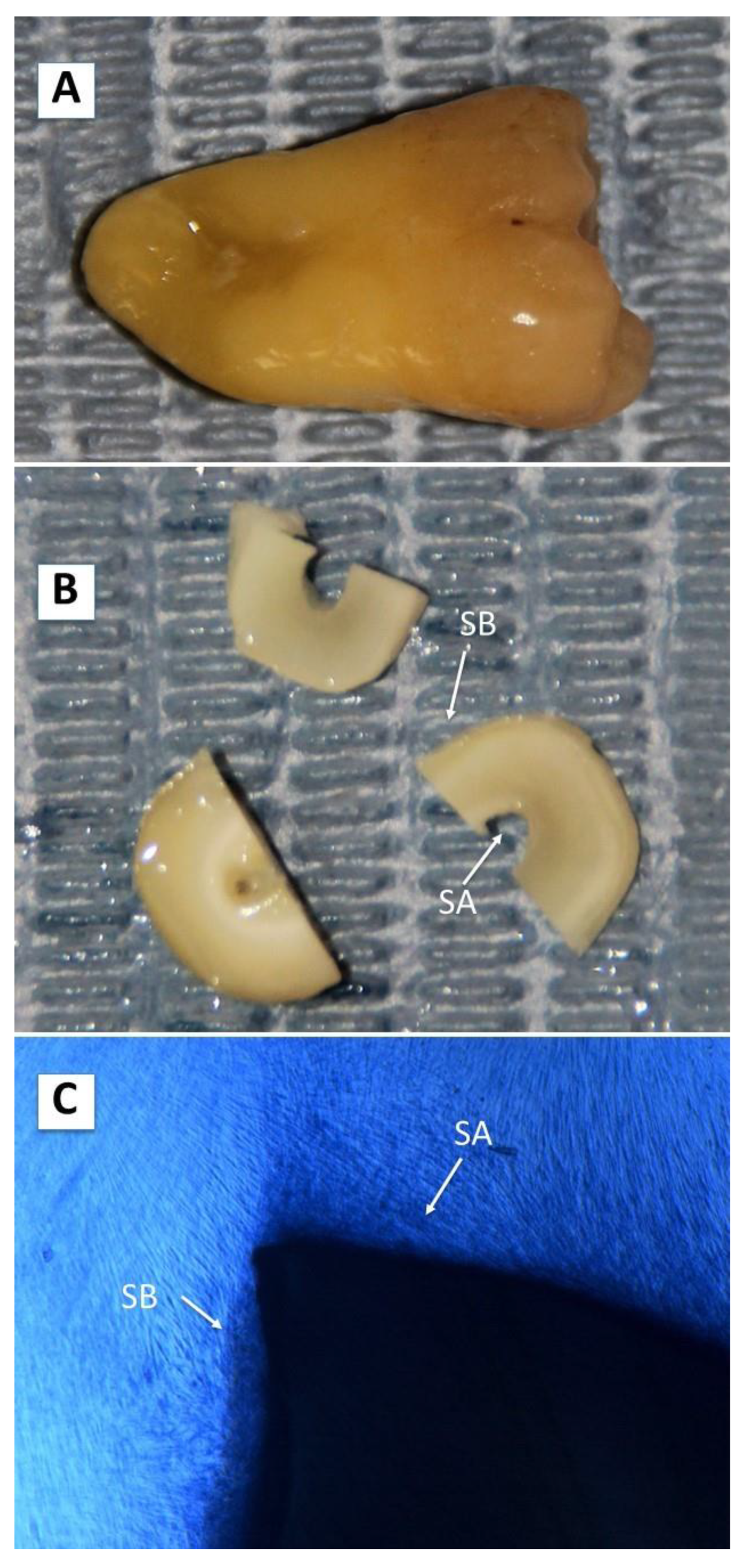
4.3. Scaffold Preparation and Study Design
4.4. Light Microscopy: Determination of Dentin Sialoprotein Expression
4.5. Transmission Electron Microscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harvey, C.E. Management of periodontal disease: Understanding the options. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 819–836. [Google Scholar] [CrossRef] [PubMed]
- Ward, E. A Review of Tissue Engineering for Periodontal Tissue Regeneration. J. Vet. Dent. 2022, 39, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Shue, L.; Yufeng, Z.; Mony, U. Biomaterials for periodontal regeneration: A review of ceramics and polymers. Biomatter 2012, 2, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Choi, S.H.; Moon, I.S.; Cho, K.S.; Chai, J.K.; Kim, C.K. Eight-week histological analysis on the effect of chitosan on surgically created one-wall intrabony defects in beagle dogs. J. Clin. Periodontol. 2003, 30, 443–453. [Google Scholar] [CrossRef]
- Hayashi, C.; Kinoshita, A.; Oda, S.; Mizutani, K.; Shirakata, Y.; Ishikawa, I. Injectable calcium phosphate bone cement provides favorable space and a scaffold for periodontal regeneration in dogs. J. Periodontol. 2006, 77, 940–946. [Google Scholar] [CrossRef]
- Yang, C.; Lee, J.S.; Jung, U.W.; Seo, Y.K.; Park, J.K.; Choi, S.H. Periodontal regeneration with nano-hyroxyapatite-coated silk scaffolds in dogs. J. Periodontal Implant Sci. 2013, 43, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Hyun, S.Y.; Lee, J.H.; Kang, K.J.; Jang, Y.J. Effect of FGF-2, TGF-β-1, and BMPs on Teno/Ligamentogenesis and Osteo/Cementogenesis of Human Periodontal Ligament Stem Cells. Mol. Cells. 2017, 40, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Arzate, H.; Zeichner-David, M.; Mercado-Celis, G. Cementum proteins: Role in cementogenesis, biomineralization, periodontium formation and regeneration. Periodontol. 2000 2015, 67, 211–233. [Google Scholar] [CrossRef]
- Bernhard Gottlieb, M.D. Biology of the Cementum. J. Periodontol. 1942, 13, 13–17. [Google Scholar] [CrossRef]
- Bernhard Gottlieb, M.D. The new concept of periodontoclasia. J. Periodontol. 1946, 13, 7–23. [Google Scholar] [CrossRef]
- Gómez de Ferraris, M.E.; Campos Muñoz, A. Histología y embriología bucodental; Editorial Médica Panamericana: Madrid, España, 1999. [Google Scholar]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbanová, J.; Soukup, T.; Suchánek, J.; Pytlík, R.; Corbeil, D.; Mokrý, J. Characterization of dental pulp stem cells from impacted third molars cultured in low serum-containing medium. Cells Tissues Organs 2011, 193, 344–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoyama, W.; Liu, Y.; Fang, D.; Yamaza, T.; Seo, B.M.; Zhang, C.; Liu, H.; Gronthos, S.; Wang, C.Y.; Wang, S.; et al. Mesenchymal stem cell-mediated functional tooth regeneration in swine. PLoS ONE 2006, 1, e79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, K.; Saito, M.; Tsunoda, A.; Yamauchi, M.; Hattori, S.; Sato, S.; Toyoda, M.; Teranaka, T.; Narayanan, A.S. Progenitor cells from dental follicle are able to form cementum matrix in vivo. Connect. Tissue Res. 2002, 43, 406–408. [Google Scholar] [CrossRef]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Yamamoto, A.; Matsubara, K.; Nakamura, S.; Naruse, M.; Yamagata, M.; Sakamoto, K.; Tauchi, R.; Wakao, N.; Imagama, S.; et al. Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J. Clin. Invest. 2012, 122, 80–90. [Google Scholar] [CrossRef]
- Chen, K.; Xiong, H.; Xu, N.; Shen, Y.; Huang, Y.; Liu, C. Chondrogenic potential of stem cells from human exfoliated deciduous teeth in vitro and in vivo. Acta Odontol. Scand. 2014, 72, 664–672. [Google Scholar] [CrossRef]
- Dai, J.; Wang, J.; Lu, J.; Zou, D.; Sun, H.; Dong, Y.; Yu, H.; Zhang, L.; Yang, T.; Zhang, X.; et al. The effect of co-culturing costal chondrocytes and dental pulp stem cells combined with exogenous FGF9 protein on chondrogenesis and ossification in engineered cartilage. Biomaterials 2012, 33, 7699–7711. [Google Scholar] [CrossRef]
- Needleman, I.G.; Worthington, H.V.; Giedrys-Leeper, E.; Tucker, R.J. Guided tissue regeneration for periodontal infra-bony defects. Cochrane Database Syst. Rev. 2006, 2, CD001724. [Google Scholar] [CrossRef] [PubMed]
- Kao, R.T.; Nares, S.; Reynolds, M.A. Periodontal regeneration—Intrabony defects: A systematic review from the AAP Regeneration Workshop. J. Periodontol. 2015, 86 (Suppl. 2), S77–S104. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Monjaraz, B.; Santiago-Osorio, E.; Monroy-García, A.; Ledesma-Martínez, E.; Mendoza-Núñez, V.M. Mesenchymal Stem Cells of Dental Origin for Inducing Tissue Regeneration in Periodontitis: A Mini-Review. Int. J. Mol. Sci. 2018, 19, 944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-de-Llano, J.J.; Mata, M.; Peydró, S.; Peydró, A.; Carda, C. Dentin tubule orientation determines odontoblastic differentiation in vitro: A morphological study. PLoS ONE 2019, 14, e0215780. [Google Scholar] [CrossRef]
- Bègue-Kirn, C.; Krebsbach, P.H.; Bartlett, J.D.; Butler, W.T. Dentin sialoprotein, dentin phosphoprotein, enamelysin and ameloblastin: Tooth-specific molecules that are distinctively expressed during murine dental differentiation. Eur. J. Oral Sci. 1998, 106, 963–970. [Google Scholar] [CrossRef]
- Butler, W.T.; Bhown, M.; Brunn, J.C.; D’Souza, R.N.; Farach-Carson, M.C.; Happonen, R.P.; Schrohenloher, R.E.; Seyer, J.M.; Somerman, M.J.; Foster, R.A.; et al. Isolation, characterization and immunolocalization of a 53-kDal dentin sialoprotein (DSP). Matrix 1992, 12, 343–351. [Google Scholar] [CrossRef]
- D’Souza, R.N.; Bronckers, A.L.; Happonen, R.P.; Doga, D.A.; Farach-Carson, M.C.; Butler, W.T. Developmental expression of a 53 KD dentin sialoprotein in rat tooth organs. J. Histochem. Cytochem. 1992, 40, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Bronckers, A.L.; D’Souza, R.N.; Butler, W.T.; Lyaruu, D.M.; van Dijk, S.; Gay, S.; Wöltgens, J.H. Dentin sialoprotein: Biosynthesis and developmental appearance in rat tooth germs in comparison with amelogenins, osteocalcin and collagen type-I. Cell Tissue Res. 1993, 272, 237–247. [Google Scholar] [CrossRef]
- Bronckers, A.L.; Farach-Carson, M.C.; Van Waveren, E.; Butler, W.T. Immunolocalization of osteopontin, osteocalcin, and dentin sialoprotein during dental root formation and early cementogenesis in the rat. J. Bone Miner Res. 1994, 9, 833–841. [Google Scholar] [CrossRef]
- Butler, W.T.; Ritchie, H. The nature and functional significance of dentin extracellular matrix proteins. Int. J. Dev. Biol. 1995, 39, 169–179. [Google Scholar]
- Baba, O.; Qin, C.; Brunn, J.C.; Jones, J.E.; Wygant, J.N.; McIntyre, B.W.; Butler, W.T. Detection of dentin sialoprotein in rat periodontium. Eur. J. Oral Sci. 2004, 112, 163–170. [Google Scholar] [CrossRef] [PubMed]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 9, 7–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.C.; Kornman, K.S. The pathogenesis of human periodontitis: An introduction. Periodontol. 2000 1997, 14, 9–11. [Google Scholar] [CrossRef]
- Dabra, S.; Chhina, K.; Soni, N.; Bhatnagar, R. Tissue engineering in periodontal regeneration: A brief review. Dent. Res. J. 2012, 9, 671–680. [Google Scholar]
- Yamamoto, T.; Hasegawa, T.; Yamamoto, T.; Hongo, H.; Amizuka, N. Histology of human cementum: Its structure, function, and development. Jpn. Dent. Sci. Rev. 2016, 52, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faltin, R.M.; Faltin, K.; Sander, F.G.; Arana-Chavez, V.E. Ultrastructure of cementum and periodontal ligament after continuous intrusion in humans: A transmission electron microscopy study. Eur. J. Orthod. 2001, 23, 35–49. [Google Scholar] [CrossRef] [PubMed]
- McKee, M.D.; Zalzal, S.; Nanci, A. Extracellular matrix in tooth cementum and mantle dentin: Localization of osteopontin and other noncollagenous proteins, plasma proteins, and glycoconjugates by electron microscopy. Anat. Rec. 1996, 245, 293–312. [Google Scholar] [CrossRef]
- Gibson, M.P.; Jani, P.; Liu, Y.; Wang, X.; Lu, Y.; Feng, J.Q.; Qin, C. Failure to process dentin sialophosphoprotein into fragments leads to periodontal defects in mice. Eur. J. Oral Sci. 2013, 121, 545–550. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata, M.; Peydró, S.; de Llano, J.J.M.; Sancho-Tello, M.; Carda, C. Human Dental Pulp Stem Cells Differentiate into Cementoid-Like-Secreting Cells on Decellularized Teeth Scaffolds. Int. J. Mol. Sci. 2022, 23, 15588. https://doi.org/10.3390/ijms232415588
Mata M, Peydró S, de Llano JJM, Sancho-Tello M, Carda C. Human Dental Pulp Stem Cells Differentiate into Cementoid-Like-Secreting Cells on Decellularized Teeth Scaffolds. International Journal of Molecular Sciences. 2022; 23(24):15588. https://doi.org/10.3390/ijms232415588
Chicago/Turabian StyleMata, Manuel, Santiago Peydró, José Javier Martín de Llano, María Sancho-Tello, and Carmen Carda. 2022. "Human Dental Pulp Stem Cells Differentiate into Cementoid-Like-Secreting Cells on Decellularized Teeth Scaffolds" International Journal of Molecular Sciences 23, no. 24: 15588. https://doi.org/10.3390/ijms232415588