
Mechanism of Synergy between Piceatannol and Ciprofloxacin against Staphylococcus aureus
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Antibacterial Activity of PIC
2.2. The Combination of PIC and CIP Has a Synergic Effect
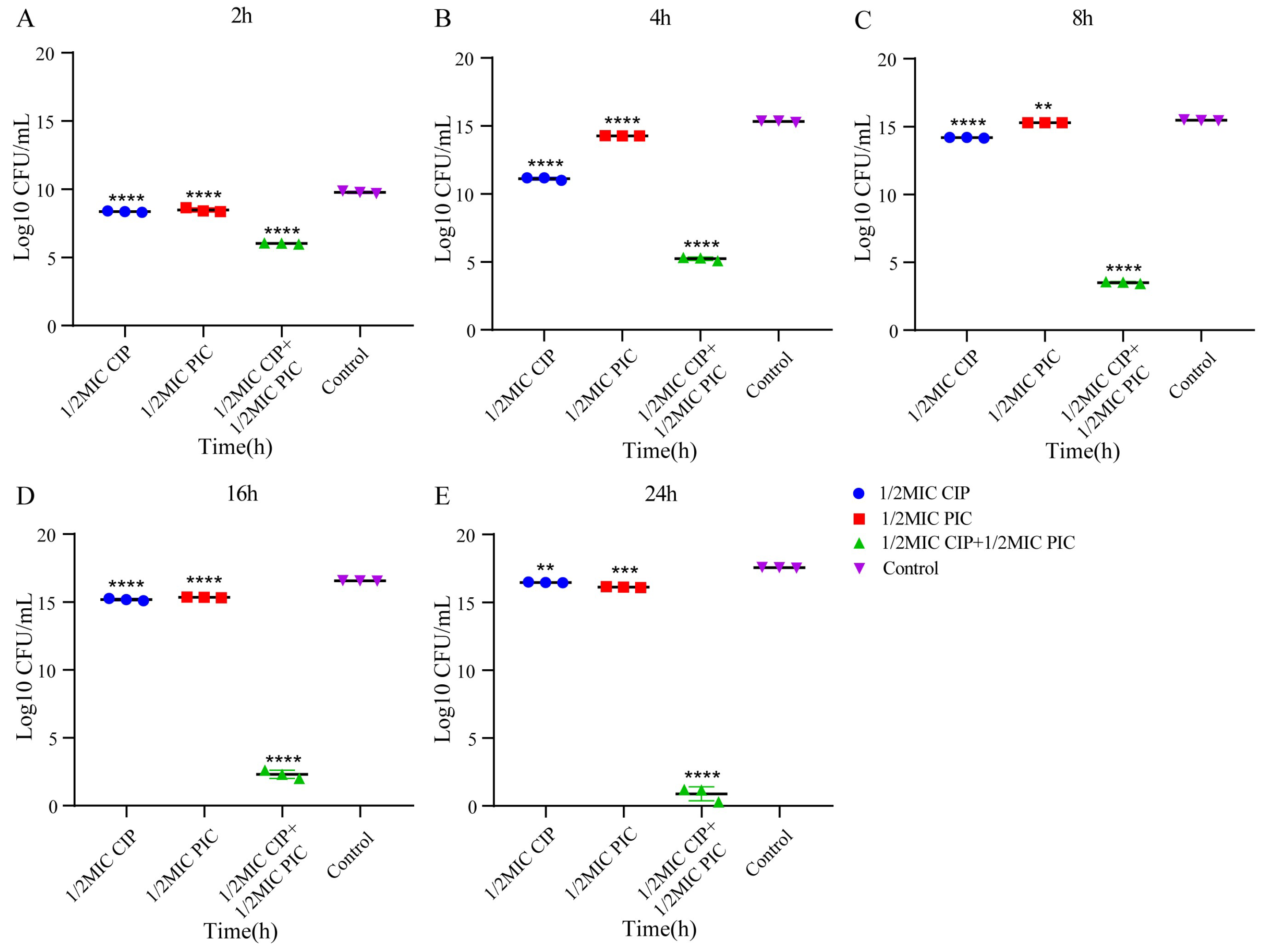
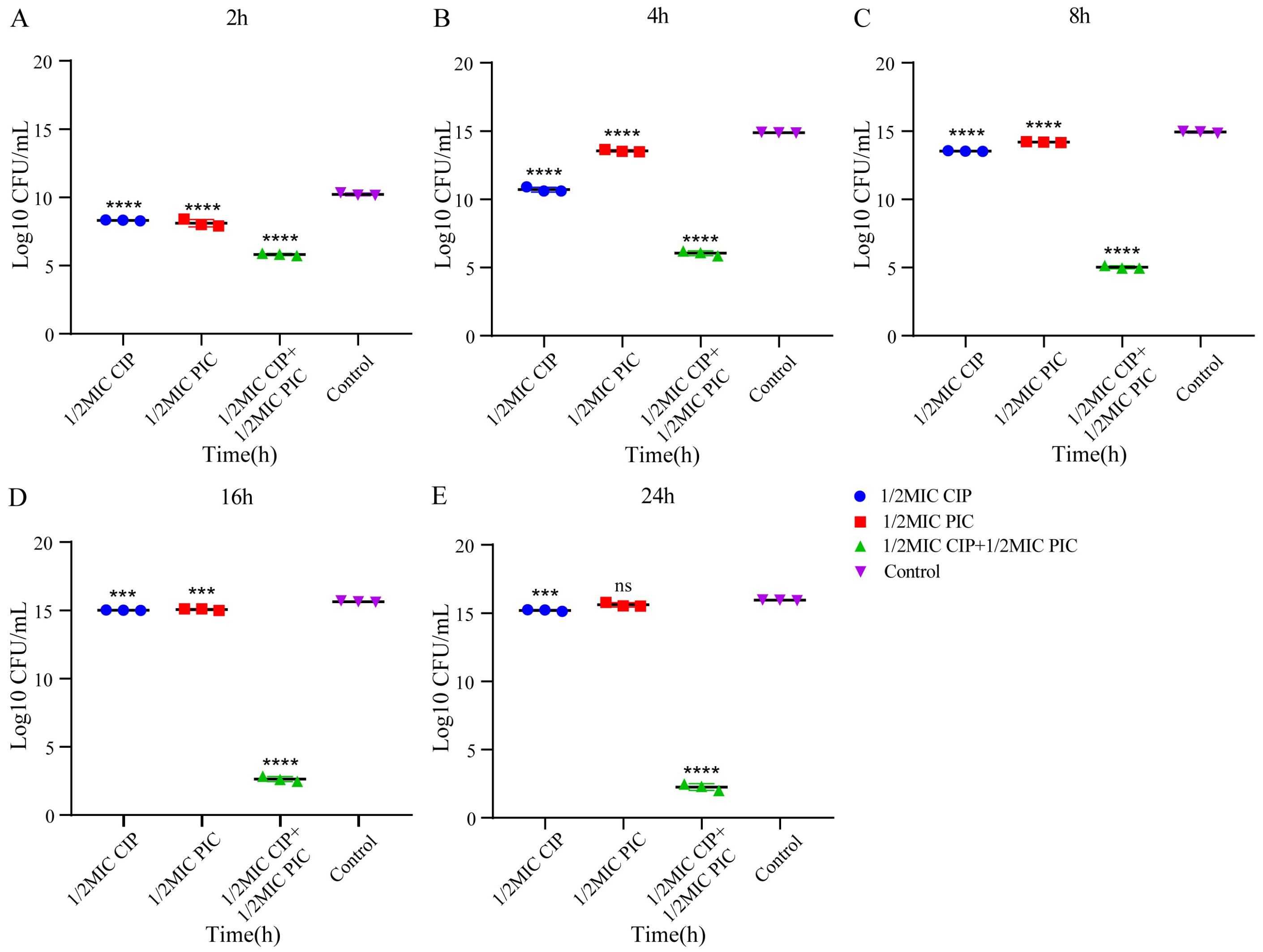
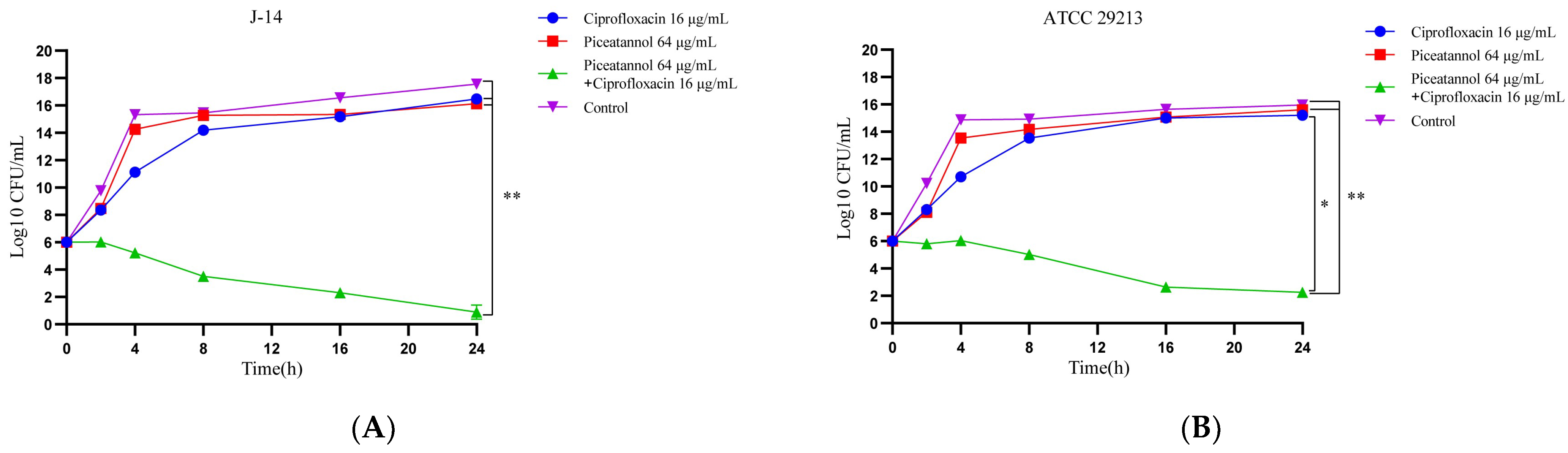
2.3. The Combination of PIC and CIP Shows Potently Bactericidal Activity against S. aureus
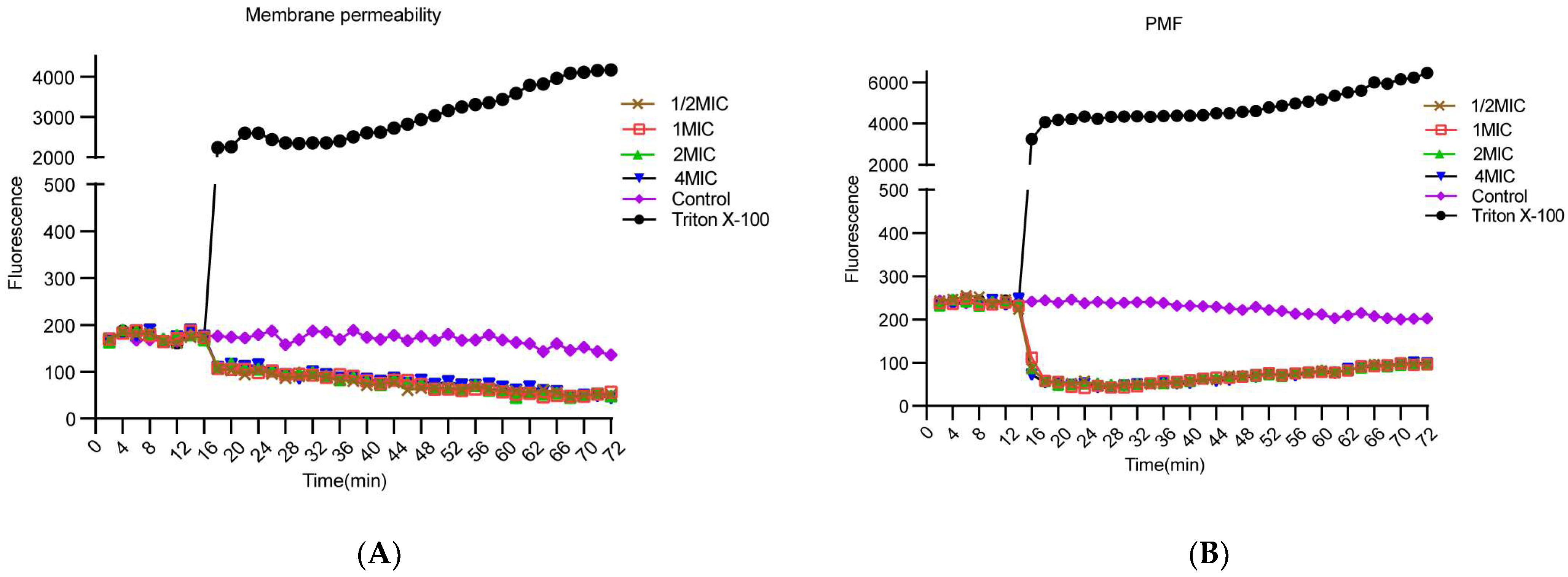
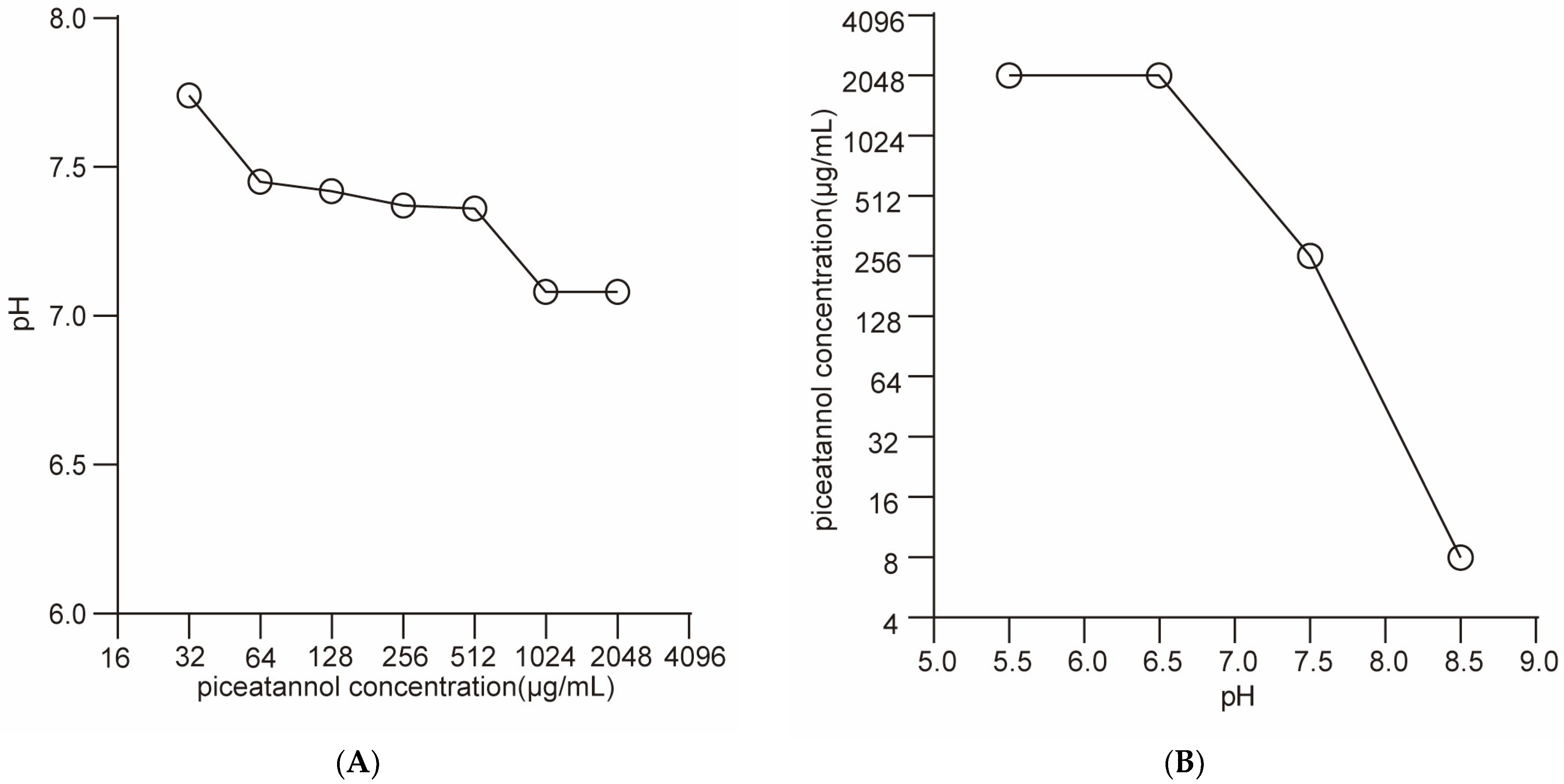
2.4. PIC Dissipates the Proton Motive Force but Does Not Increase the Membrane Permeability of S. aureus
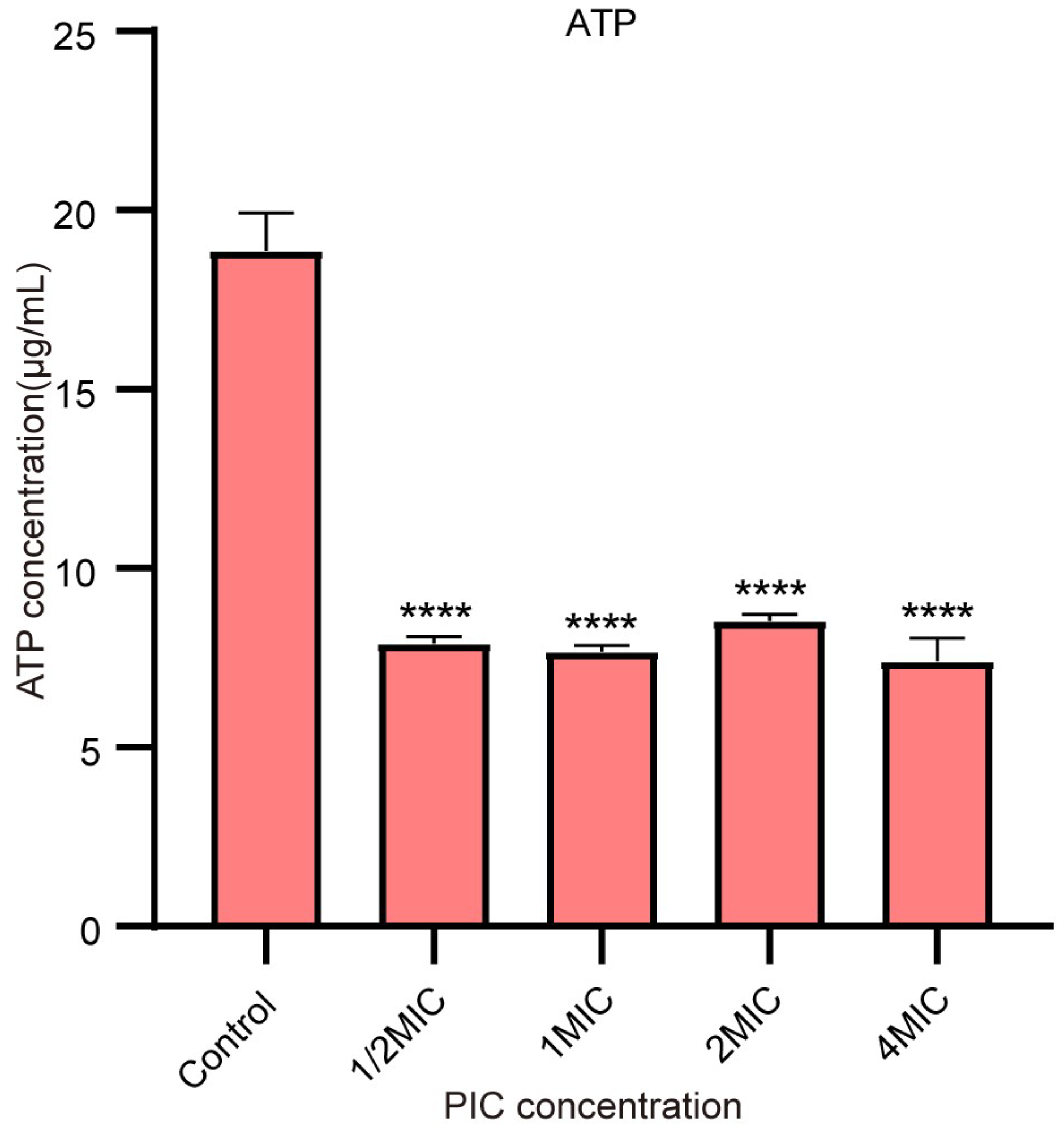
2.5. PIC Reduces the ATP Synthesis of S. aureus
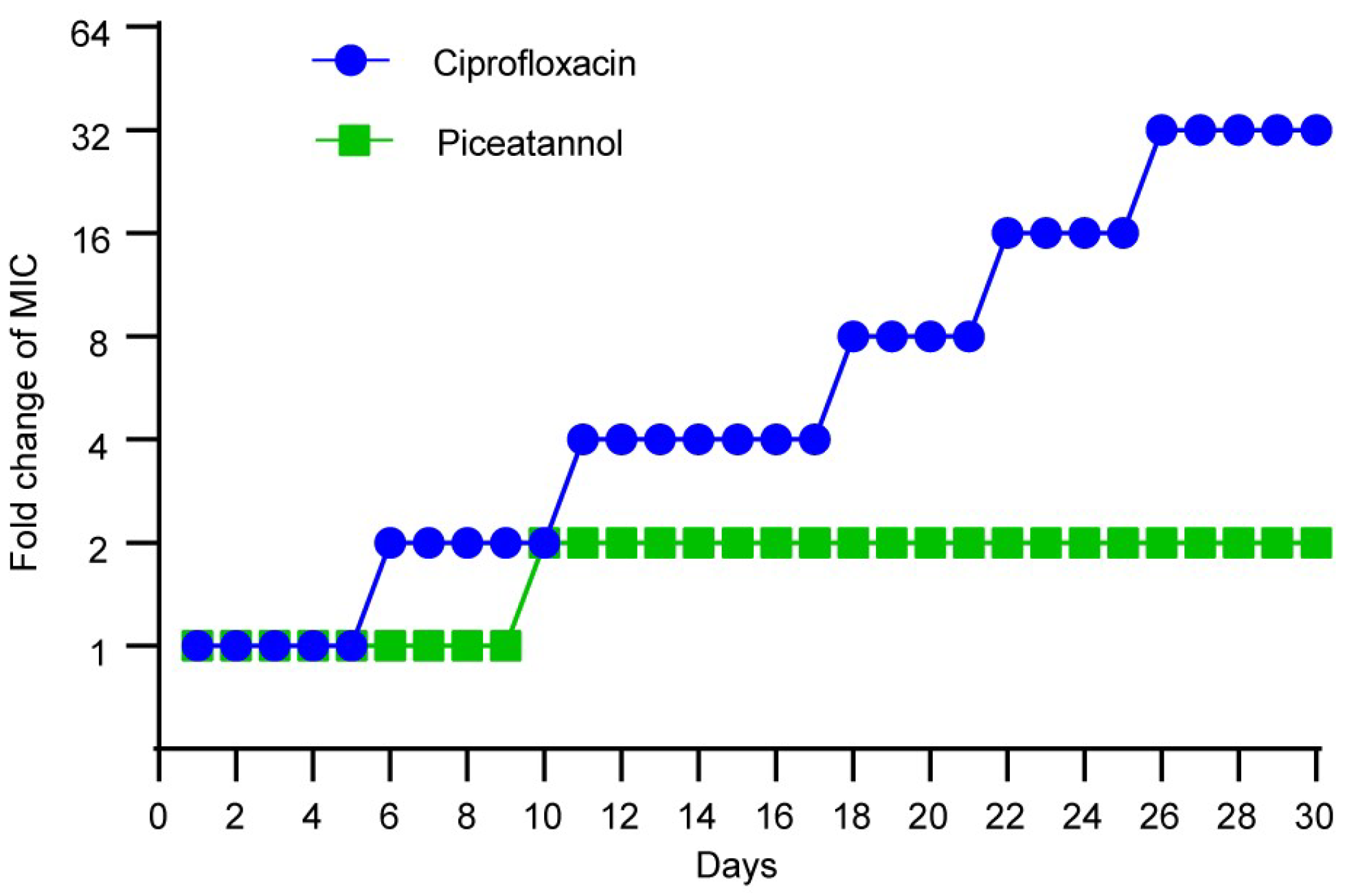
2.6. PIC Displays Low Levels of Resistance Development
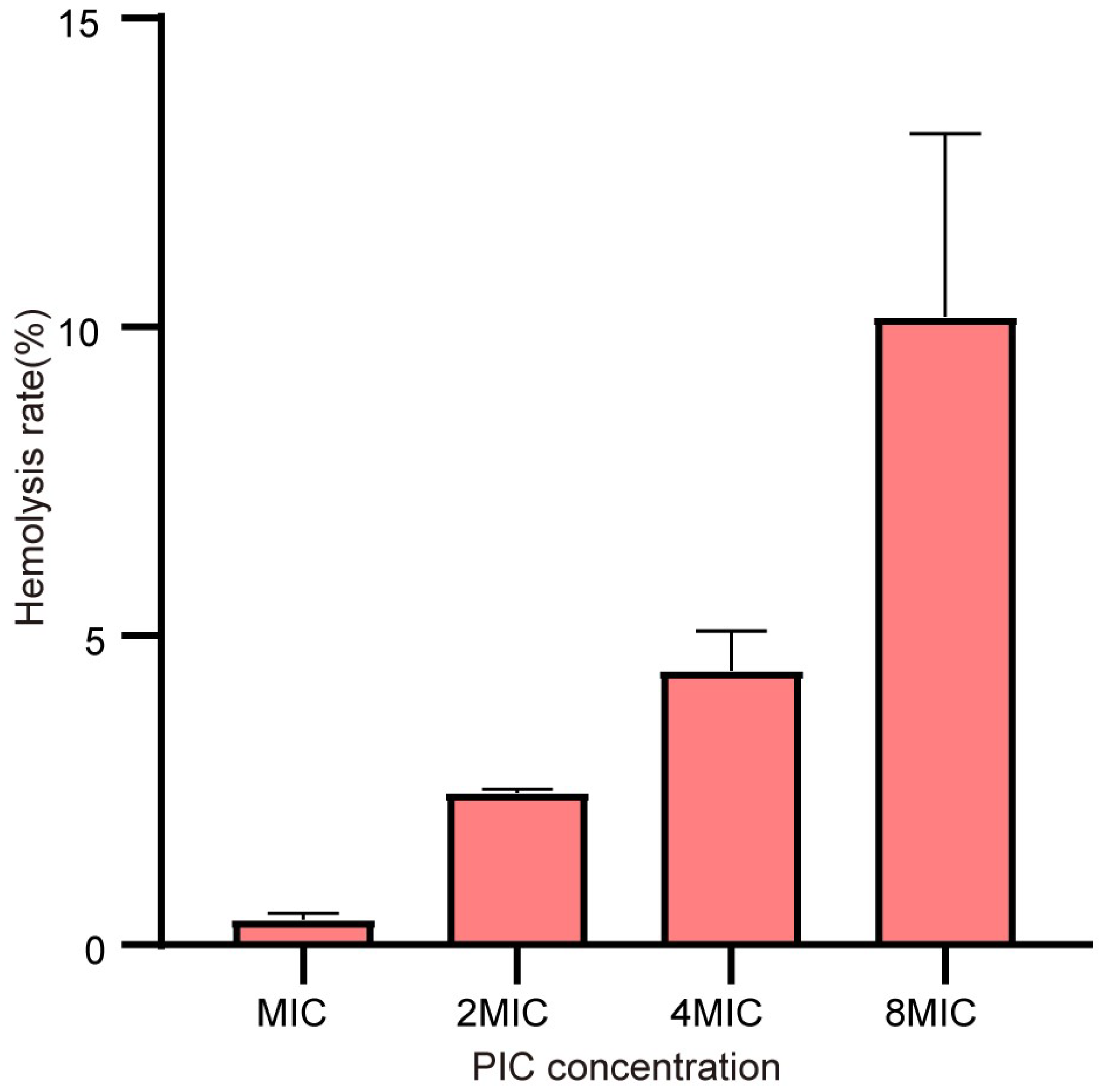
2.7. PIC has Slight Hemolytic Activity on Mammalian Erythrocytes
3. Discussion
4. Materials and Methods
4.1. Materials and Bacterial Strains
4.2. Antimicrobial Activity
4.2.1. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
4.2.2. Antibiotic Synergism Tests
4.2.3. Time–Kill Assay
4.3. The Antibacterial Mechanism
4.3.1. Membrane Permeability Assay
4.3.2. Proton Motive Force Assay
4.3.3. Effects of pH on Antibacterial Activity
4.3.4. ATP Assay
4.4. Resistance Study
4.5. Hemolysis Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreira, B.M.; Daum, R.S. Antimicrobial Resistance in Staphylococci. Pediatr. Clin. N. Am. 1995, 42, 619–648. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Mansour, N.; Loubet, P.; Pouget, C.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.-P.; Molle, V. Staphylococcus aureus Toxins: An Update on Their Pathogenic Properties and Potential Treatments. Toxins 2021, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.A.; Coombs, G.W.; Nimmo, G.R. Staphylococcus aureus ‘Down Under’: Contemporary epidemiology of S. aureus in Australia, New Zealand, and the South West Pacific. Clin. Microbiol. Infect. 2014, 20, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Abril, A.G.; Gonzalez-Villa, T.; Barros-Velázquez, J.; Cañas, B.; Sánchez-Pérez, A.; Calo-Mata, P.; Carrera, M. Staphylococcus aureus Exotoxins and Their Detection in the Dairy Industry and Mastitis. Toxins 2020, 12, 537. [Google Scholar] [CrossRef]
- Wang, M.; Fan, Z.; Han, H. Autophagy in Staphylococcus aureus Infection. Front. Cell. Infect. Microbiol. 2021, 11, 750222. [Google Scholar] [CrossRef]
- Plumet, L.; Ahmad-Mansour, N.; Dunyach-Remy, C.; Kissa, K.; Sotto, A.; Lavigne, J.-P.; Costechareyre, D.; Molle, V. Bacteriophage Therapy for Staphylococcus Aureus Infections: A Review of Animal Models, Treatments, and Clinical Trials. Front. Cell. Infect. Microbiol. 2022, 12, 907314. [Google Scholar] [CrossRef]
- Zhang, G.-F.; Liu, X.; Zhang, S.; Pan, B.; Liu, M.-L. Ciprofloxacin derivatives and their antibacterial activities. Eur. J. Med. Chem. 2018, 146, 599–612. [Google Scholar] [CrossRef]
- Felix, L.; Mishra, B.; Khader, R.; Ganesan, N.; Mylonakis, E. In Vitro and In Vivo Bactericidal and Antibiofilm Efficacy of Alpha Mangostin Against Staphylococcus aureus Persister Cells. Front. Cell. Infect. Microbiol. 2022, 12, 898794. [Google Scholar] [CrossRef]
- Rohinishree, Y.S.; Negi, P.S. Effect of licorice extract on cell viability, biofilm formation and exotoxin production by Staphylococcus aureus. J. Food Sci. Technol. 2015, 53, 1092–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorska, H.; Smoragiewicz, W. Role of probiotics in the prevention and treatment of meticillin-resistant Staphylococcus aureus infections. Int. J. Antimicrob. Agents 2013, 42, 475–481. [Google Scholar] [CrossRef]
- Chen, H.; Yang, N.; Yu, L.; Li, J.; Zhang, H.; Zheng, Y.; Xu, M.; Liu, Y.; Yang, Y.; Li, J. Synergistic Microbicidal Effect of AUR and PEITC Against Staphylococcus aureus Skin Infection. Front. Cell. Infect. Microbiol. 2022, 12, 927289. [Google Scholar] [CrossRef]
- Silva, E.; Monteiro, R.; Grainha, T.; Alves, D.; Pereira, M.O.; Sousa, A.M. Fostering Innovation in the Treatment of Chronic Polymicrobial Cystic Fibrosis-Associated Infections Exploring Aspartic Acid and Succinic Acid as Ciprofloxacin Adjuvants. Front. Cell. Infect. Microbiol. 2020, 10, 441. [Google Scholar] [CrossRef]
- Matsui, Y.; Sugiyama, K.; Kamei, M.; Takahashi, T.; Suzuki, T.; Katagata, Y.; Ito, T. Extract of Passion Fruit (Passiflora edulis) Seed Containing High Amounts of Piceatannol Inhibits Melanogenesis and Promotes Collagen Synthesis. J. Agric. Food Chem. 2010, 58, 11112–11118. [Google Scholar] [CrossRef]
- Sano, S.; Sugiyama, K.; Ito, T.; Katano, Y.; Ishihata, A. Identification of the Strong Vasorelaxing Substance Scirpusin B, a Dimer of Piceatannol, from Passion Fruit (Passiflora edulis) Seeds. J. Agric. Food Chem. 2011, 59, 6209–6213. [Google Scholar] [CrossRef]
- Sáez, V.; Pastene, E.; Vergara, C.; Mardones, C.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S.; Gómez, M.V.; Theoduloz, C.; Riquelme, S.; von Baer, D. Oligostilbenoids in Vitis vinifera L. Pinot Noir grape cane extract: Isolation, characterization, in vitro antioxidant capacity and anti-proliferative effect on cancer cells. Food Chem. 2018, 265, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.; Hsieh, T.-C.; Halicka, H.D.; Darzynkiewicz, Z.; Wu, J.M. Upregulation of PD-L1 expression by resveratrol and piceatannol in breast and colorectal cancer cells occurs via HDAC3/p300-mediated NF-κB signaling. Int. J. Oncol. 2018, 53, 1469–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanapal, J.; Ravindrran, M.B. Chitosan/poly (lactic acid)-coated piceatannol nanoparticles exert an in vitro apoptosis activity on liver, lung and breast cancer cell lines. Artif. Cells Nanomed. Biotechnol. 2018, 46, 274–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoda, R.; Hamada, H.; Uesugi, D.; Iwahara, N.; Nojima, I.; Horio, Y.; Kuno, A. Different Antioxidative and Antiapoptotic Effects of Piceatannol and Resveratrol. J. Pharmacol. Exp. Ther. 2020, 376, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, S.; Kinoshita, Y.; Maruki-Uchida, H.; Yanae, K.; Sai, M.; Ito, T. Piceatannol and Its Metabolite, Isorhapontigenin, Induce SIRT1 Expression in THP-1 Human Monocytic Cell Line. Nutrients 2014, 6, 4794–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijampatnam, B.; Zhang, H.; Cai, X.; Michalek, S.M.; Wu, H.; Velu, S.E. Inhibition of Streptococcus mutans Biofilms by the Natural Stilbene Piceatannol Through the Inhibition of Glucosyltransferases. ACS Omega 2018, 3, 8378–8385. [Google Scholar] [CrossRef] [PubMed]
- Güldaş, N.; Uysal, T.; Ellidokuz, H.; BAŞBINAR, Y. Antimicrobial Effect of Piceatannol, a Resveratrol Metabolite, on Staphylococcus Aureus. J. Basic Clin. Health Sci. 2022, 3, 184–187. [Google Scholar] [CrossRef]
- Ekiert, H.M.; Szopa, A. Biological Activities of Natural Products II. Molecules 2022, 27, 1519. [Google Scholar] [CrossRef]
- Zheng, J.; Ramirez, V.D. Piceatannol, a Stilbene Phytochemical, Inhibits Mitochondrial F0F1-ATPase Activity by Targeting the F1 Complex. Biochem. Biophys. Res. Commun. 1999, 261, 499–503. [Google Scholar] [CrossRef]
- Hawkey, P.M. Pre-clinical experience with daptomycin. J. Antimicrob. Chemother. 2008, 62, iii7–iii14. [Google Scholar] [CrossRef]
- Hazen, K.C. Fungicidal versus fungistatic activity of terbinafine and itraconazole: An in vitro comparison. J. Am. Acad. Dermatol. 1998, 38, S37–S41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qu, X.; Tang, H.; Wang, Y.; Yang, H.; Yuan, W.; Yue, B. Diclofenac Resensitizes Methicillin-Resistant Staphylococcus aureus to β -Lactams and Prevents Implant Infections. Adv. Sci. 2021, 8, 2100681. [Google Scholar] [CrossRef]
- Müller, A.; Wenzel, M.; Strahl, H.; Grein, F.; Saaki, T.N.V.; Kohl, B.; Siersma, T.; Bandow, J.E.; Sahl, H.-G.; Schneider, T.; et al. Daptomycin inhibits cell envelope synthesis by interfering with fluid membrane microdomains. Proc. Natl. Acad. Sci. USA 2016, 113, E7077–E7086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcera, M.J.; Elferink, M.G.L.; Driessen, A.J.M.; Konings, W.N. In vitro pore-forming activity of the lantibiotic nisin. Role of protonmotive force and lipid composition. JBIC J. Biolchem. 1993, 212, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-C.; Han, F.; Song, M.-R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.-Z. Natural Flavones from Morus alba against Methicillin-Resistant Staphylococcus aureus via Targeting the Proton Motive Force and Membrane Permeability. J. Agric. Food Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef] [PubMed]
- Ruhr, E.; Sahl, H.G. Mode of action of the peptide antibiotic nisin and influence on the membrane potential of whole cells and on cytoplasmic and artificial membrane vesicles. Antimicrob. Agents Chemother. 1985, 27, 841–845. [Google Scholar] [CrossRef]
- Bruno, M.E.; Kaiser, A.; Montville, T.J. Depletion of proton motive force by nisin in Listeria monocytogenes cells. Appl. Environ. Microbiol. 1992, 58, 2255–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, C.C. Ciprofloxacin: In Vitro Activity, Mechanism of Action, and Resistance. Clin. Infect. Dis. 1988, 10, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-M. Drug induced hemolysis. Gen. Pharmacol. Vasc. Syst. 1977, 8, 13–19. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, G.-Y.; Zhang, X.-J.; Han, J.; Li, Y.-Q.; Wang, G.-C. In vitro synergism of magnolol and honokiol in combination with antibacterial agents against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). BMC Complement. Altern. Med. 2015, 15, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, S.; Dai, C.; Shen, Z.; Tang, Q.; Wang, H.; Zhai, B.; Zhao, L.; Hao, Z. Mechanism of Synergy Between Tetracycline and Quercetin Against Antibiotic Resistant Escherichia coli. Front. Microbiol. 2019, 10, 2536. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Koh, J.-J.; Aung, T.T.; Sin, W.L.W.; Lim, F.; Wang, L.; Lakshminarayanan, R.; Zhou, L.; Tan, D.T.H.; Cao, D.; et al. Semisynthetic Flavone-Derived Antimicrobials with Therapeutic Potential against Methicillin-Resistant Staphylococcus aureus (MRSA). J. Med. Chem. 2017, 60, 6152–6165. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | PIC | CIP | MET | VAN | CTX | GEN | TCY |
|---|---|---|---|---|---|---|---|
| ATCC 29213 | 128 | 4 | 2 | 4 | 2 | 2 | 0.25 |
| J-28 | 128 | 32 | 2 | 2 | 1 | 16 | 32 |
| J-6 | 64 | 4 | 2 | 4 | 1 | 0.25 | 0.25 |
| J-11 | 64 | 32 | 1 | 1 | 1 | 4 | 0.25 |
| J-14 | 128 | 32 | 2 | 2 | 2 | 32 | 0.25 |
| J-9 | 128 | 32 | 2 | 2 | 1 | 8 | 32 |
| Strains | MBC (μg/mL) | MBC/MIC |
|---|---|---|
| ATCC 29213 | 256 | 2 |
| J-28 | 256 | 2 |
| J-6 | 512 | 8 |
| J-11 | 256 | 4 |
| J-14 | 256 | 2 |
| J-9 | 256 | 2 |
| Antibiotics | ATCC 29213 | J-28 | J-6 | J-11 | J-14 | J-9 |
|---|---|---|---|---|---|---|
| CIP | 0.75 | 0.5 | 0.75 | 0.375 | 0.375 | 1 |
| MET | 2 | 0.75 | 0.75 | 1 | 1 | 0.75 |
| VAN | 1.25 | 1 | 0.5625 | 0.75 | 1 | 1 |
| CTX | 2.5 | 1 | 2 | 1 | 1 | 0.75 |
| GEN | 0.625 | 0.625 | 0.75 | 1.125 | 0.75 | 0.75 |
| TCY | 0.75 | 0.75 | 0.75 | 2 | 2 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.; Bai, Y.; Qiu, Y.; Zhang, X.; Zeng, Z.; Chen, L.; Cheng, F.; Zhang, J. Mechanism of Synergy between Piceatannol and Ciprofloxacin against Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 15341. https://doi.org/10.3390/ijms232315341
Shi M, Bai Y, Qiu Y, Zhang X, Zeng Z, Chen L, Cheng F, Zhang J. Mechanism of Synergy between Piceatannol and Ciprofloxacin against Staphylococcus aureus. International Journal of Molecular Sciences. 2022; 23(23):15341. https://doi.org/10.3390/ijms232315341
Chicago/Turabian StyleShi, Mengyan, Yubin Bai, Yanhua Qiu, Xinxin Zhang, Zikang Zeng, Lingling Chen, Fusheng Cheng, and Jiyu Zhang. 2022. "Mechanism of Synergy between Piceatannol and Ciprofloxacin against Staphylococcus aureus" International Journal of Molecular Sciences 23, no. 23: 15341. https://doi.org/10.3390/ijms232315341