
Elevated Expression of CCN3 in Articular Cartilage Induces Osteoarthritis in Hip Joints Irrespective of Age and Weight Bearing
 ,
,
Abstract
:1. Introduction
2. Results
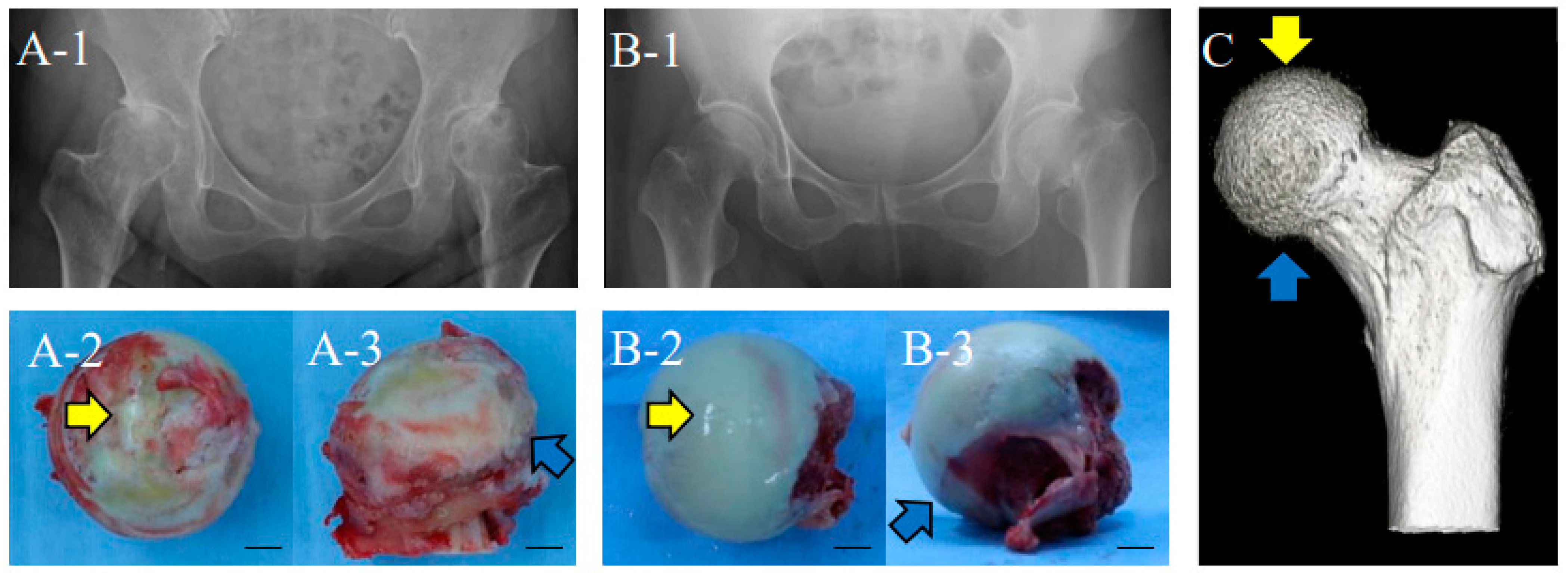
2.1. Hip Femoral Articular Cartilage from Human Patients
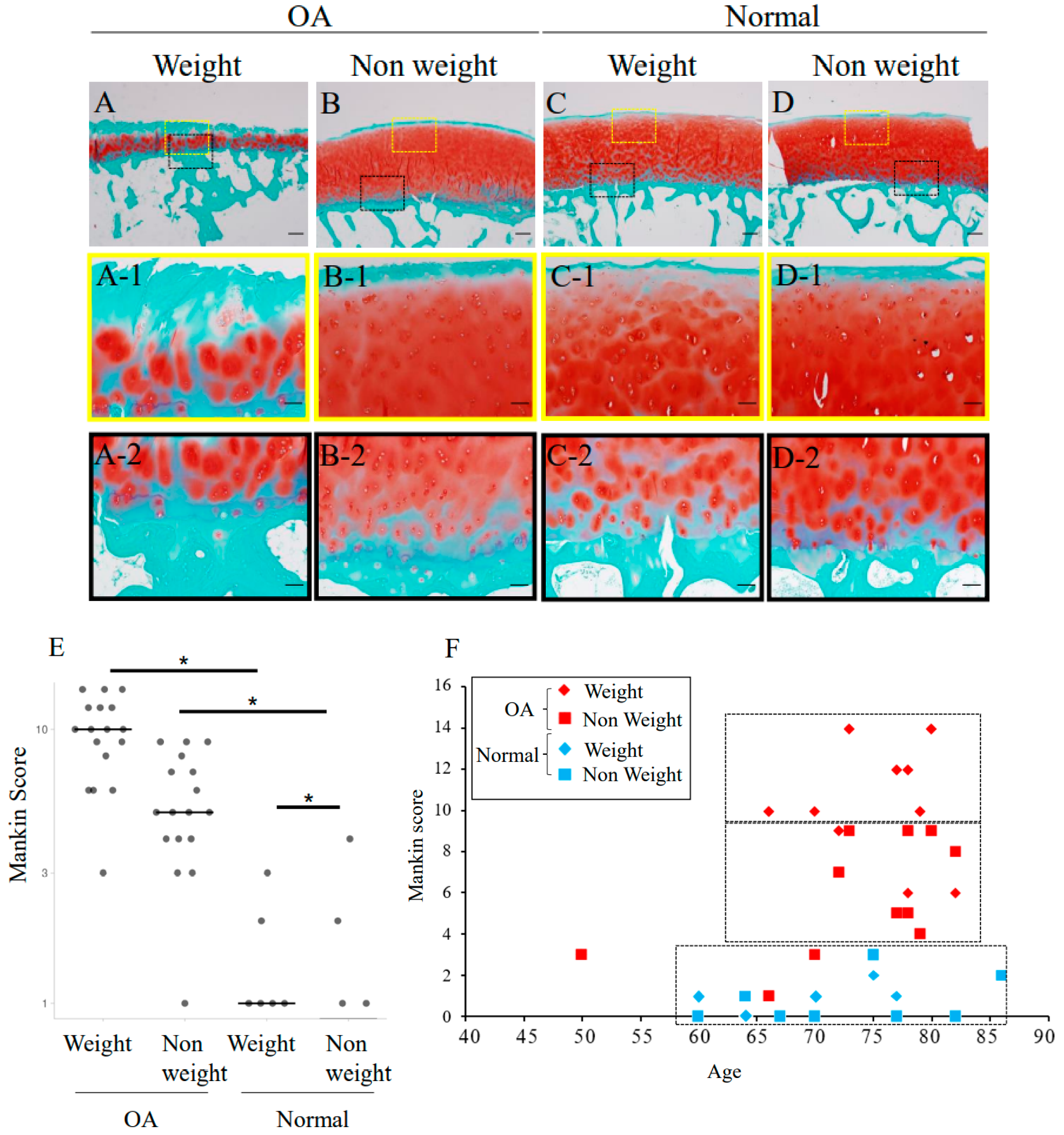
2.2. Safranin-O Staining and Mankin Score Evaluation of Degeneration Grade of Human Femoral Cartilage
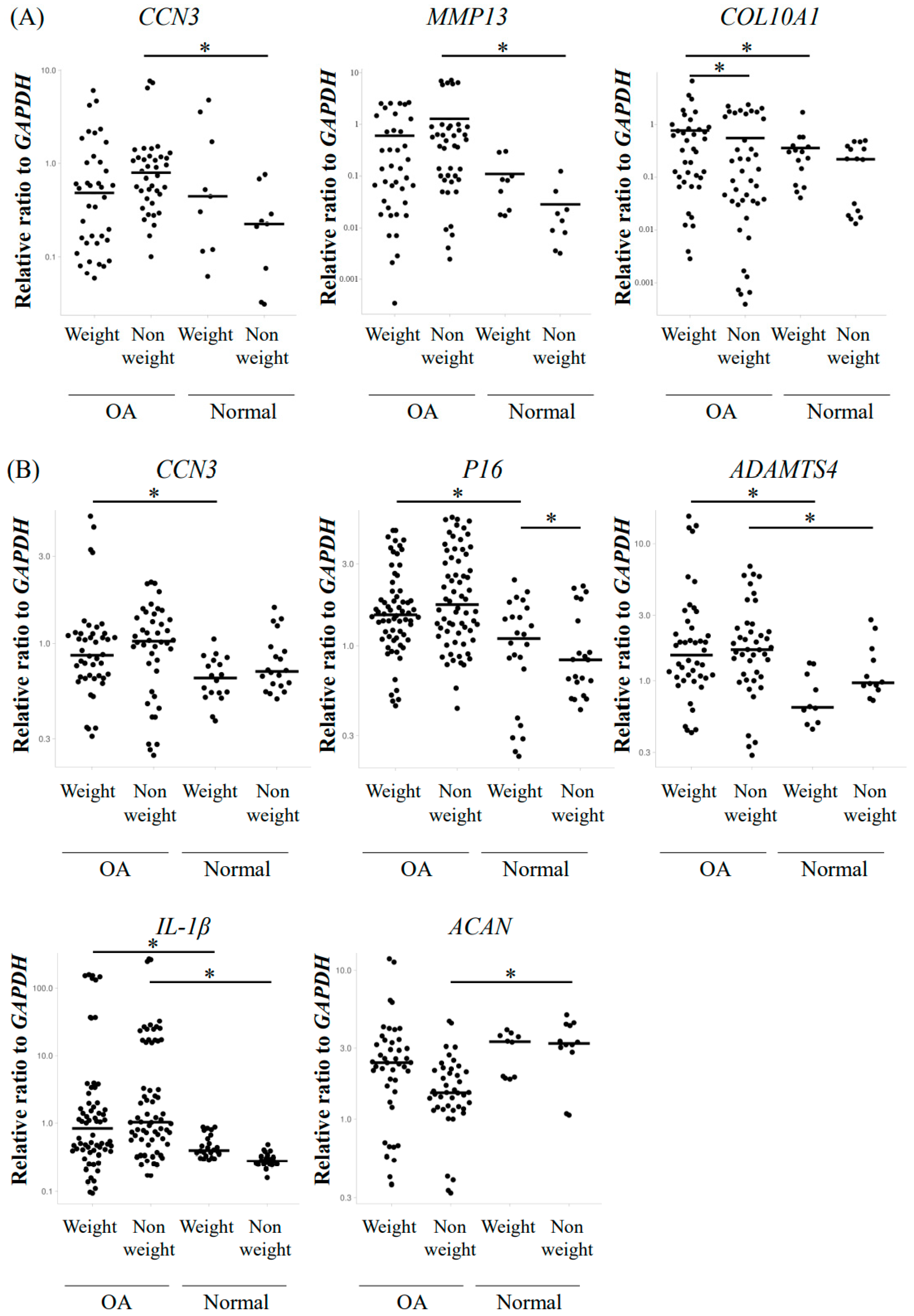
2.3. Elevated Gene Expression of CCN3 and OA-Related Factors in Osteoarthritic Human Hip Femoral Articular Cartilage
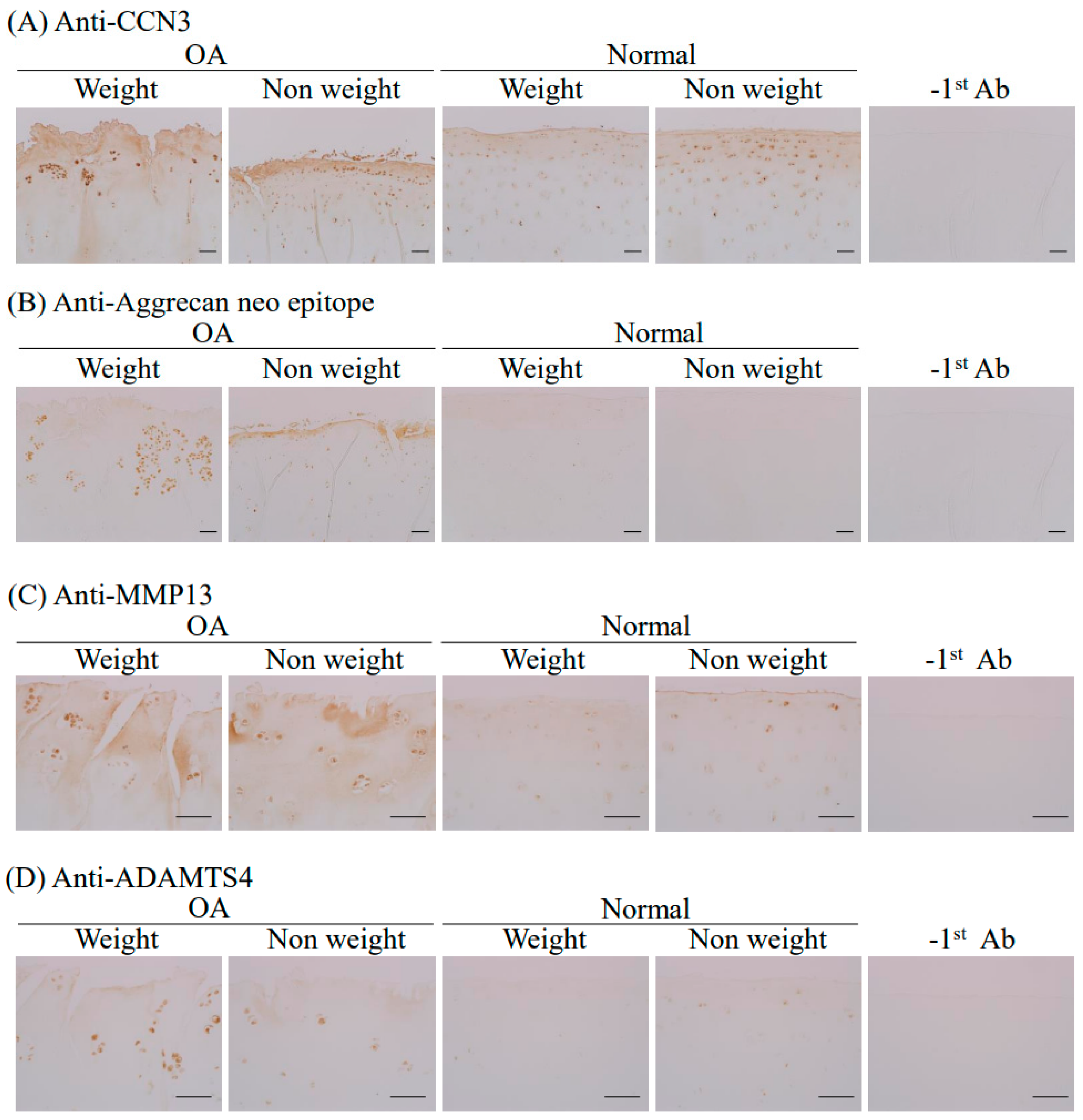
2.4. Immunohistochemical Analysis of CCN3, Aggrecan Neoepitope, MMP13, and ADAMTS4 in Human Femoral Articular Cartilage
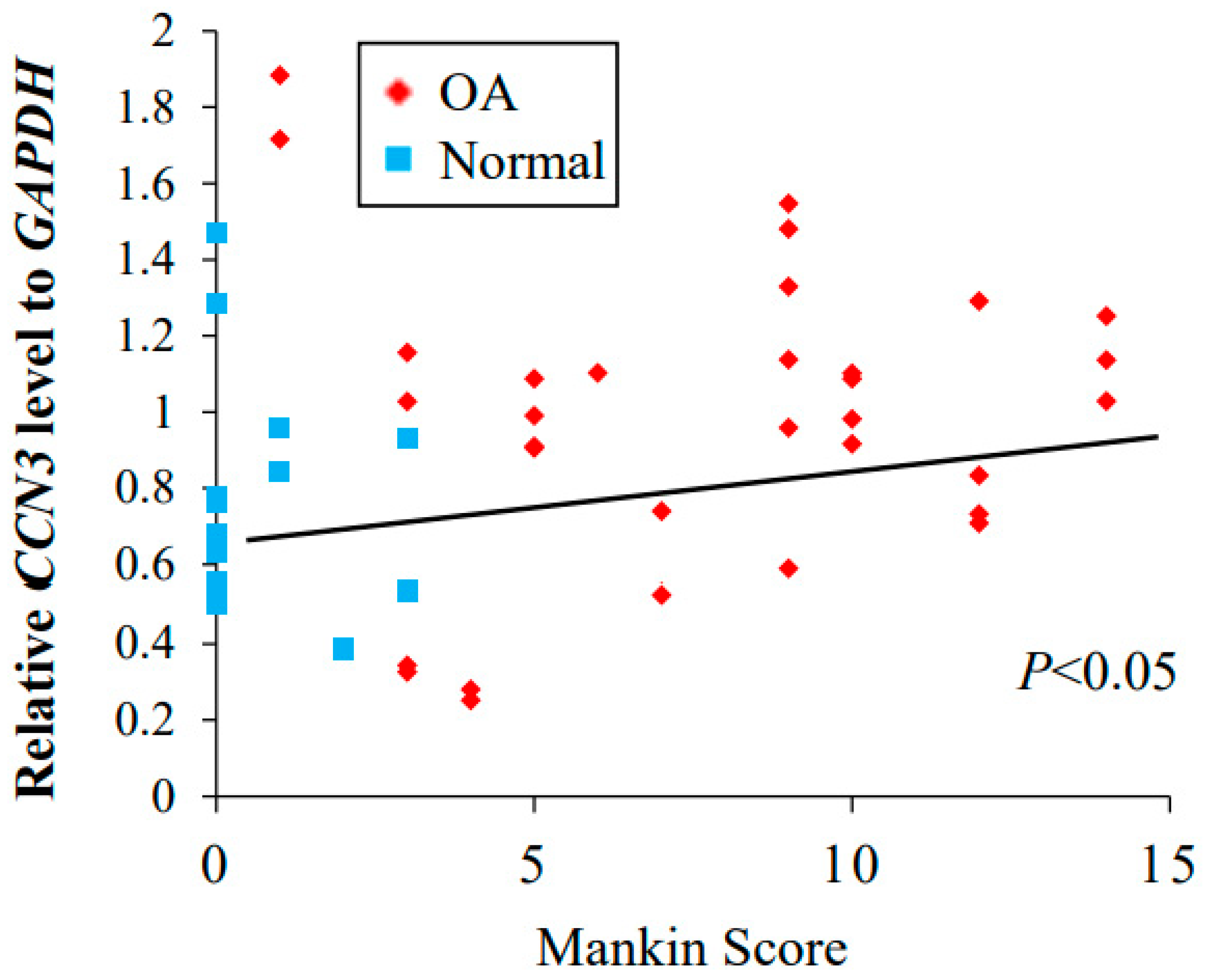
2.5. Correlation between the Expression of CCN3 and Mankin Score
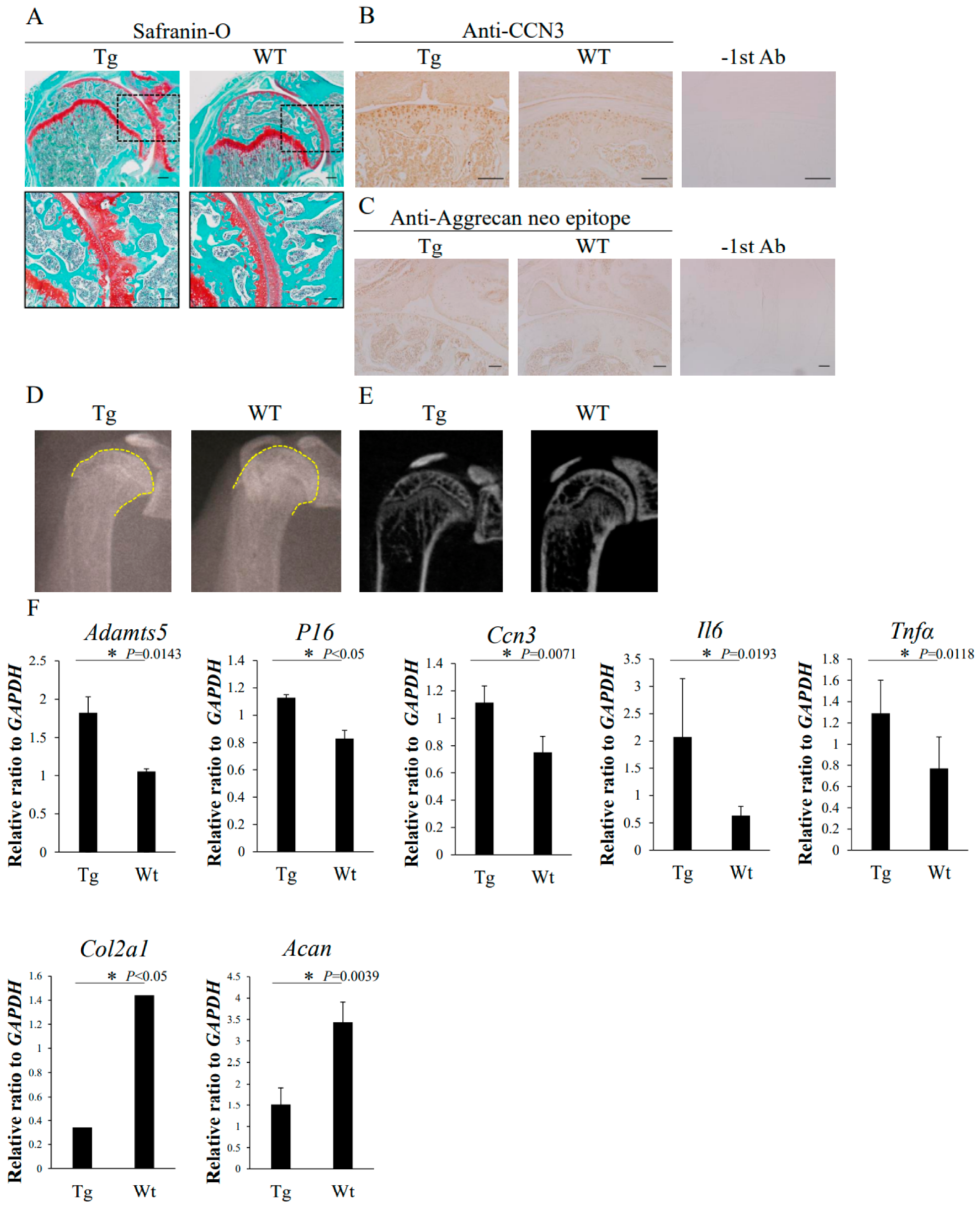
2.6. Cartilage-Specific Overexpression of Ccn3 Induced OA-like Degenerative Changes in Particular Hip Joint In Vivo
3. Discussion
4. Materials and Methods
4.1. Population and Sample Collection
4.2. Treatment of Femoral Heads
4.3. Cell Culture
4.4. Isolation of Total RNA
4.5. Real-Time Polymerase Chain Reaction
4.6. Histology
4.7. Analyzing Hip Joints of Ccn3 Transgenic Mice
4.8. Mankin Score
4.9. X-ray Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leichtenberg, C.S.; van Tol, F.R.; Gademan, M.G.J.; Krom, T.; Tilbury, C.; Horemans, H.L.D.; Bussmann, J.B.J.; Verdegaal, S.H.M.; Marijnissen, W.; Nelissen, R.; et al. Are pain, functional limitations and quality of life associated with objectively measured physical activity in patients with end-stage osteoarthritis of the hip or knee? Knee 2021, 29, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Papalia, R.; Campi, S.; Vorini, F.; Zampogna, B.; Vasta, S.; Papalia, G.; Fossati, C.; Torre, G.; Denaro, V. The Role of Physical Activity and Rehabilitation Following Hip and Knee Arthroplasty in the Elderly. J. Clin. Med. 2020, 9, 1401. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, M.R.; Murtha, A.S.; Clohisy, J.C.; Group, A.S. Developmental Dysplasia of the Hip in Adolescents and Young Adults. J. Am. Acad. Orthop. Surg. 2020, 28, 91–101. [Google Scholar] [CrossRef]
- Aki, T.; Hashimoto, K.; Ogasawara, M.; Itoi, E. A whole-genome transcriptome analysis of articular chondrocytes in secondary osteoarthritis of the hip. PLoS ONE 2018, 13, e0199734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnusson, K.; Scurrah, K.; Ystrom, E.; Orstavik, R.E.; Nilsen, T.; Steingrimsdottir, O.A.; Ferreira, P.; Fenstad, A.M.; Furnes, O.; Hagen, K.B. Genetic factors contribute more to hip than knee surgery due to osteoarthritis—A population-based twin registry study of joint arthroplasty. Osteoarthr. Cartil. 2017, 25, 878–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funck-Brentano, T.; Nethander, M.; Moverare-Skrtic, S.; Richette, P.; Ohlsson, C. Causal Factors for Knee, Hip, and Hand Osteoarthritis: A Mendelian Randomization Study in the UK Biobank. Arthritis Rheumatol. 2019, 71, 1634–1641. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, P.A.; Wells, J.; Usheva, E.; Nakonezny, P.A.; Barati, Z.; Gonzalez, R.; Kassem, L.; Henson, F.M.D. Early-Onset Osteoarthritis originates at the chondrocyte level in Hip Dysplasia. Sci. Rep. 2020, 10, 627. [Google Scholar] [CrossRef] [Green Version]
- Kawaki, H.; Kubota, S.; Suzuki, A.; Lazar, N.; Yamada, T.; Matsumura, T.; Ohgawara, T.; Maeda, T.; Perbal, B.; Lyons, K.M.; et al. Cooperative regulation of chondrocyte differentiation by CCN2 and CCN3 shown by a comprehensive analysis of the CCN family proteins in cartilage. J. Bone Miner Res. 2008, 23, 1751–1764. [Google Scholar] [CrossRef]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of Osteoarthritis of the Hand, Hip, and Knee. Arthritis Care Res. 2020, 72, 149–162. [Google Scholar] [CrossRef]
- Aweid, O.; Haider, Z.; Saed, A.; Kalairajah, Y. Treatment modalities for hip and knee osteoarthritis: A systematic review of safety. J. Orthop. Surg. 2018, 26, 1–12. [Google Scholar] [CrossRef]
- Grynpas, M.D.; Alpert, B.; Katz, I.; Lieberman, I.; Pritzker, K.P. Subchondral bone in osteoarthritis. Calcif. Tissue Int. 1991, 49, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Holbourn, K.P.; Acharya, K.R.; Perbal, B. The CCN family of proteins: Structure-function relationships. Trends Biochem. Sci. 2008, 33, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, M. CCN2: A master regulator of the genesis of bone and cartilage. J. Cell Commun. Signal. 2013, 7, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leask, A.; Abraham, D.J. All in the CCN family: Essential matricellular signaling modulators emerge from the bunker. J. Cell Sci. 2006, 119 Pt 23, 4803–4810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, M.; Nakamura, Y.; Maruyama, M.; Abe, K.; Watanapokasin, R.; Kato, H. Expression profiles of human CCN genes in patients with osteoarthritis or rheumatoid arthritis. J. Orthop. Sci. 2015, 20, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, M.; Kadoya, K.; Kondo, S.; Fu, S.; Miyake, Y.; Ogo, A.; Ono, M.; Furumatsu, T.; Nakata, E.; Sasaki, T.; et al. CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage. Int. J. Mol. Sci. 2020, 21, 7556. [Google Scholar] [CrossRef]
- Pauli, C.; Whiteside, R.; Heras, F.L.; Nesic, D.; Koziol, J.; Grogan, S.P.; Matyas, J.; Pritzker, K.P.; D’Lima, D.D.; Lotz, M.K. Comparison of cartilage histopathology assessment systems on human knee joints at all stages of osteoarthritis development. Osteoarthr. Cartil. 2012, 20, 476–485. [Google Scholar] [CrossRef] [Green Version]
- Mankin, H.J. Biochemical and metabolic aspects of osteoarthritis. Orthop. Clin. N. Am. 1971, 2, 19–31. [Google Scholar] [CrossRef]
- Roddy, K.A.; Boulter, C.A. Targeted mutation of NOV/CCN3 in mice disrupts joint homeostasis and causes osteoarthritis-like disease. Osteoarthr. Cartil. 2015, 23, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Summanen, M.; Ukkola-Vuoti, L.; Kurki, S.; Tuominen, S.; Madanat, R. The burden of hip and knee osteoarthritis in Finnish occupational healthcare. BMC Musculoskelet. Disord. 2021, 22, 501. [Google Scholar] [CrossRef]
- Koseki, H.; Sunagawa, S.; Noguchi, C.; Yonekura, A.; Matsumura, U.; Watanabe, K.; Nishiyama, Y.; Osaki, M. Incidence of and risk factors for hip fracture in Nagasaki, Japan from 2005 to 2014. Arch. Osteoporos. 2021, 16, 111. [Google Scholar] [CrossRef] [PubMed]
- Litwic, A.; Edwards, M.H.; Dennison, E.M.; Cooper, C. Epidemiology and burden of osteoarthritis. Br. Med. Bull. 2013, 105, 185–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, E.; Gonzalez-Garcia, C.; Gomez Ribelles, J.L.; Salmeron-Sanchez, M. Maintenance of chondrocyte phenotype during expansion on PLLA microtopographies. J. Tissue Eng. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schild, C.; Trueb, B. Three members of the connective tissue growth factor family CCN are differentially regulated by mechanical stress. Biochim. Biophys. Acta 2004, 1691, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, S.; Kawaki, H.; Perbal, B.; Kawata, K.; Hattori, T.; Nishida, T. Cellular communication network factor 3 in cartilage development and maintenance. J. Cell Commun. Signal. 2021, 15, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Jayasuriya, C.T.; Hu, N.; Li, J.; Lemme, N.; Terek, R.; Ehrlich, M.G.; Chen, Q. Molecular characterization of mesenchymal stem cells in human osteoarthritis cartilage reveals contribution to the OA phenotype. Sci. Rep. 2018, 8, 7044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnik, S.; Gabler, J.; Dreher, S.I.; Hecht, N.; Hofmann, N.; Grossner, T.; Richter, W. MiR-218 affects hypertrophic differentiation of human mesenchymal stromal cells during chondrogenesis via targeting RUNX2, MEF2C, and COL10A1. Stem. Cell Res. Ther. 2020, 11, 532. [Google Scholar] [CrossRef]
- Coryell, P.R.; Diekman, B.O.; Loeser, R.F. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat. Rev. Rheumatol. 2021, 17, 47–57. [Google Scholar] [CrossRef]
- Boulestreau, J.; Maumus, M.; Jorgensen, C.; Noel, D. Extracellular vesicles from mesenchymal stromal cells: Therapeutic perspectives for targeting senescence in osteoarthritis. Adv. Drug Deliv. Rev. 2021, 175, 113836. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Mori, D.; Kobayashi, H.; Mori, Y.; Nakamoto, H.; Okada, K.; Taniguchi, Y.; Sugita, S.; Yano, F.; Chung, U.I.; et al. Excessive mechanical loading promotes osteoarthritis through the gremlin-1-NF-kappaB pathway. Nat. Commun. 2019, 10, 1442. [Google Scholar] [CrossRef]
- Deng, Z.H.; Li, Y.S.; Gao, X.; Lei, G.H.; Huard, J. Bone morphogenetic proteins for articular cartilage regeneration. Osteoarthr. Cartil. 2018, 26, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetsunaga, T.; Nishida, K.; Furumatsu, T.; Naruse, K.; Hirohata, S.; Yoshida, A.; Saito, T.; Ozaki, T. Regulation of mechanical stress-induced MMP-13 and ADAMTS-5 expression by RUNX-2 transcriptional factor in SW1353 chondrocyte-like cells. Osteoarthr. Cartil. 2011, 19, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wu, X.; Lu, J.; Huang, G.; Dang, L.; Zhang, H.; Zhong, C.; Zhang, Z.; Li, D.; Li, F.; et al. Exosomal transfer of osteoclast-derived miRNAs to chondrocytes contributes to osteoarthritis progression. Nat. Aging 2021, 1, 368–384. [Google Scholar] [CrossRef]
- Backenkohler, U.; Halata, Z.; Strasmann, T.J. The sensory innervation of the shoulder joint of the mouse. Ann. Anat. 1996, 178, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Oki, S.; Shirasawa, H.; Yoda, M.; Matsumura, N.; Tohmonda, T.; Yuasa, K.; Nakamura, M.; Matsumoto, M.; Horiuchi, K. Generation and characterization of a novel shoulder contracture mouse model. J. Orthop. Res. 2015, 33, 1732–1738. [Google Scholar] [CrossRef]
- Joliot, V.; Martinerie, C.; Dambrine, G.; Plassiart, G.; Brisac, M.; Crochet, J.; Perbal, B. Proviral rearrangements and overexpression of a new cellular gene (nov) in myeloblastosis-associated virus type 1-induced nephroblastomas. Mol. Cell Biol. 1992, 12, 10–21. [Google Scholar]
- Chevalier, G.; Yeger, H.; Martinerie, C.; Laurent, M.; Alami, J.; Schofield, P.N.; Perbal, B. novH: Differential expression in developing kidney and Wilm’s tumors. Am. J. Pathol. 1998, 152, 1563–1575. [Google Scholar]
- Yu, C.; Le, A.T.; Yeger, H.; Perbal, B.; Alman, B.A. NOV (CCN3) regulation in the growth plate and CCN family member expression in cartilage neoplasia. J. Pathol. 2003, 201, 609–615. [Google Scholar] [CrossRef]
- Mizukawa, T.; Nishida, T.; Akashi, S.; Kawata, K.; Kikuchi, S.; Kawaki, H.; Takigawa, M.; Kamioka, H.; Kubota, S. RFX1-mediated CCN3 induction that may support chondrocyte survival under starved conditions. J. Cell Physiol. 2021, 236, 6884–6896. [Google Scholar] [CrossRef]
- Huang, X.; Ni, B.; Mao, Z.; Xi, Y.; Chu, X.; Zhang, R.; Ma, X.; You, H. NOV/CCN3 induces cartilage protection by inhibiting PI3K/AKT/mTOR pathway. J. Cell Mol. Med. 2019, 23, 7525–7534. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| human samples for direct RNA purification from cartilage | |||
| Variables | OA | Normal | p value |
| Males: Females | 1:11 | 1:03 | NS |
| Age (years) | 71.9 ± 10.0 | 74.0 ± 6.8 | NS |
| BMI (kg/m2) | 25.8 ± 5.3 | 19.8 ± 4.0 | NS |
| human samples for direct RNA purification from primary cultured chondrocytes | |||
| Variables | OA | Normal | p value |
| Males: Females | 1:10 | 1:03 | NS |
| Age (years) | 70.2 ± 10.1 | 73.5 ± 6.6 | NS |
| BMI (kg/m2) | 25.7 ± 5.5 | 21.1 ± 1.6 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirose, K.; Kuwahara, M.; Nakata, E.; Tetsunaga, T.; Yamada, K.; Saiga, K.; Takigawa, M.; Ozaki, T.; Kubota, S.; Hattori, T. Elevated Expression of CCN3 in Articular Cartilage Induces Osteoarthritis in Hip Joints Irrespective of Age and Weight Bearing. Int. J. Mol. Sci. 2022, 23, 15311. https://doi.org/10.3390/ijms232315311
Hirose K, Kuwahara M, Nakata E, Tetsunaga T, Yamada K, Saiga K, Takigawa M, Ozaki T, Kubota S, Hattori T. Elevated Expression of CCN3 in Articular Cartilage Induces Osteoarthritis in Hip Joints Irrespective of Age and Weight Bearing. International Journal of Molecular Sciences. 2022; 23(23):15311. https://doi.org/10.3390/ijms232315311
Chicago/Turabian StyleHirose, Kazuki, Miho Kuwahara, Eiji Nakata, Tomonori Tetsunaga, Kazuki Yamada, Kenta Saiga, Masaharu Takigawa, Toshifumi Ozaki, Satoshi Kubota, and Takako Hattori. 2022. "Elevated Expression of CCN3 in Articular Cartilage Induces Osteoarthritis in Hip Joints Irrespective of Age and Weight Bearing" International Journal of Molecular Sciences 23, no. 23: 15311. https://doi.org/10.3390/ijms232315311