
Effects of Pro-Inflammatory Cytokines on Hepatic Metabolism in Primary Human Hepatocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
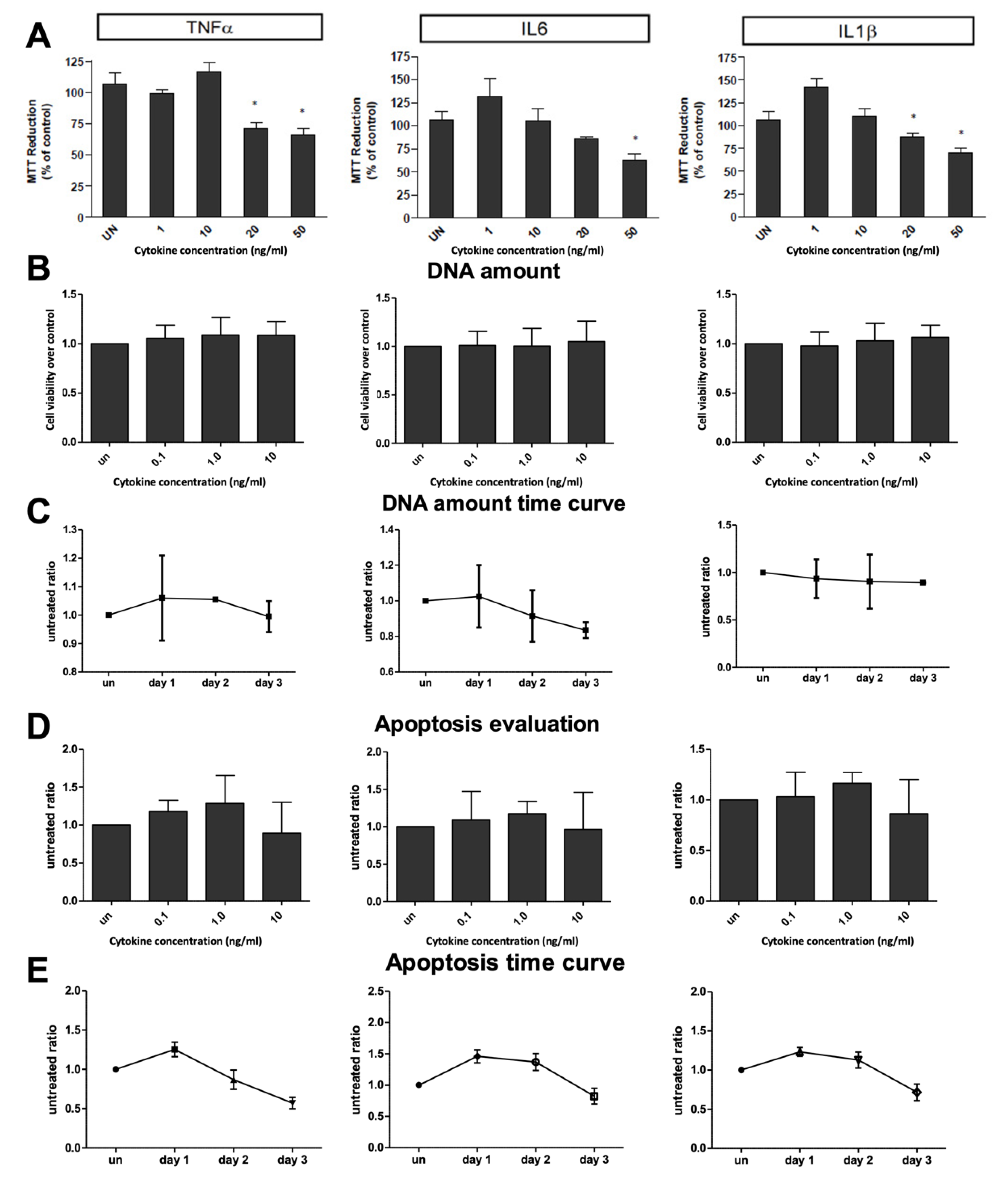
2.1. Inflammatory Cytokines on Ammonia Metabolism
2.2. Inflammatory Cytokines on CYP Activities
2.3. Inflammatory Cytokines on Phase II Enzyme (UGT1A1) Activity
2.4. Inflammatory Cytokines on Drug Transporter (BSEP) Activity
3. Discussion
4. Materials and Methods
4.1. Primary Hepatocyte Cultures
4.2. Cytotoxicity Evaluation
4.3. Apoptosis Evaluation
4.4. Ammonia Metabolism Assay
4.5. Cytochrome P450 Enzyme Activities
4.6. Phase II in Vitro Evaluation
4.7. Evaluation of BSEP Activity
4.8. Protein Amount Quantification
4.9. dsDNA Amount Quantification
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gramignoli, R.; Vosough, M.; Kannisto, K.; Srinivasan, R.C.; Strom, S.C. Clinical hepatocyte transplantation: Practical limits and possible solutions. Eur. Surg. Res. 2015, 54, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Strom, S.C. Human hepatocyte transplantation: Worldwide results. Transplantation 2006, 82, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Siewert, E.; Bort, R.; Kluge, R.; Heinrich, P.C.; Castell, J.; Jover, R. Hepatic cytochrome P450 down-regulation during aseptic inflammation in the mouse is interleukin 6 dependent. Hepatology 2000, 32, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Iber, H.; Chen, Q.; Cheng, P.Y.; Morgan, E.T. Suppression of CYP2C11 gene transcription by interleukin-1 mediated by NF-kappaB binding at the transcription start site. Arch. Biochem. Biophys. 2000, 377, 187–194. [Google Scholar] [CrossRef]
- Morgan, E.T. Regulation of cytochromes P450 during inflammation and infection. Drug Metab. Rev. 1997, 29, 1129–1188. [Google Scholar] [CrossRef]
- Ghonem, N.; Yoshida, J.; Stoltz, D.B.; Humar, A.; Starzl, T.E.; Murase, N.; Venkataramanan, R. Treprostinil, a prostacyclin analog, ameliorates ischemia-reperfusion injury in rat orthotopic liver transplantation. Am. J. Transplant. 2011, 11, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Piquette-Miller, M.; Pak, A.; Kim, H.; Anari, R.; Shahzamani, A. Decreased expression and activity of P-glycoprotein in rat liver during acute inflammation. Pharm. Res. 1998, 15, 706–711. [Google Scholar] [CrossRef]
- Lee, G.; Piquette-Miller, M. Influence of IL-6 on MDR and MRP-mediated multidrug resistance in human hepatoma cells. Can. J. Physiol. Pharmacol. 2001, 79, 876–884. [Google Scholar] [CrossRef]
- Lee, G.; Piquette-Miller, M. Cytokines alter the expression and activity of the multidrug resistance transporters in human hepatoma cell lines; analysis using RT-PCR and cDNA microarrays. J. Pharm. Sci. 2003, 92, 2152–2163. [Google Scholar] [CrossRef]
- Sukhai, M.; Yong, A.; Pak, A.; Piquette-Miller, M. Decreased expression of P-glycoprotein in interleukin-1beta and interleukin-6 treated rat hepatocytes. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2001, 50, 362–370. [Google Scholar]
- Siewert, E.; Dietrich, C.G.; Lammert, F.; Heinrich, P.C.; Matern, S.; Gartung, C.; Geier, A. Interleukin-6 regulates hepatic transporters during acute-phase response. Biochem. Biophys. Res. Commun. 2004, 322, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Shahabi, P.; Siest, G.; Meyer, U.; Visvikis-Siest, S. Human cytochrome P450 epoxygenases: Variability in expression and role in inflammation-related disorders. Pharmacol. Ther. 2014, 144, 134–161. [Google Scholar] [CrossRef] [PubMed]
- Dickmann, L.J.; Patel, S.K.; Wienkers, L.C.; Slatter, J.G. Effects of interleukin 1beta (IL-1beta) and IL-1beta/interleukin 6 (IL-6) combinations on drug metabolizing enzymes in human hepatocyte culture. Curr. Drug Metab. 2012, 13, 930–937. [Google Scholar] [CrossRef]
- Ke, S.; Rabson, A.B.; Germino, J.F.; Gallo, M.A.; Tian, Y. Mechanism of suppression of cytochrome P-450 1A1 expression by tumor necrosis factor-alpha and lipopolysaccharide. J. Biol. Chem. 2001, 276, 39638–39644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beigneux, A.P.; Moser, A.H.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. Reduction in cytochrome P-450 enzyme expression is associated with repression of CAR (constitutive androstane receptor) and PXR (pregnane X receptor) in mouse liver during the acute phase response. Biochem. Biophys. Res. Commun. 2002, 293, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Beier, D.; Gollan, J.L. Regulation of hepatocyte bile salt transporters by endotoxin and inflammatory cytokines in rodents. Gastroenterology 1996, 111, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Whiting, J.F.; Rosenbluth, A.B.; Beier, D.; Gollan, J.L. Interleukin-6 inhibits hepatocyte taurocholate uptake and sodium-potassium-adenosinetriphosphatase activity. Am. J. Physiol. 1994, 267, G1094–G1100. [Google Scholar] [CrossRef]
- Whiting, J.F.; Green, R.M.; Rosenbluth, A.B.; Gollan, J.L. Tumor necrosis factor-alpha decreases hepatocyte bile salt uptake and mediates endotoxin-induced cholestasis. Hepatology 1995, 22, 1273–1278. [Google Scholar]
- Muhlfeld, A.; Kubitz, R.; Dransfeld, O.; Haussinger, D.; Wettstein, M. Taurine supplementation induces multidrug resistance protein 2 and bile salt export pump expression in rats and prevents endotoxin-induced cholestasis. Arch. Biochem. Biophys. 2003, 413, 32–40. [Google Scholar] [CrossRef]
- Koj, A. Initiation of acute phase response and synthesis of cytokines. Biochim. Biophys. Acta 1996, 1317, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Baumann, H.; Gauldie, J. The acute phase response. Immunol. Today 1994, 15, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Puppi, J.; Hughes, R.D.; Mitry, R.R. Human hepatocyte transplantation: Current experience and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Soltys, K.A.; Setoyama, K.; Tafaleng, E.N.; Soto Gutiérrez, A.; Fong, J.; Fukumitsu, K.; Nishikawa, T.; Nagaya, M.; Sada, R.; Haberman, K.; et al. Host conditioning and rejection monitoring in hepatocyte transplantation in humans. J. Hepatol. 2017, 66, 987–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorns, C.; Nowak, G.; Nemeth, A.; Zemack, H.; Mörk, L.M.; Johansson, H.; Gramignoli, R.; Watanabe, M.; Alheim, M.; Hauzenberger, D.; et al. De novo donor-specific HLA antibody formation in two patients with Crigler-Najjar type I following human hepatocyte transplantation with partial hepatectomy preconditioning. Am. J. Transpl. 2016, 16, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Mas, V.R.; Maluf, D.G.; Thompson, M.; Ferreira-Gonzalez, A.; Fisher, R.A. Engraftment measurement in human liver tissue after liver cell transplantation by short tandem repeats analysis. Cell Transplant. 2004, 13, 231–236. [Google Scholar] [CrossRef]
- Abdel-Razzak, Z.; Loyer, P.; Fautrel, A.; Gautier, J.C.; Corcos, L.; Turlin, B.; Beaune, P.; Guillouzo, A. Cytokines down-regulate expression of major cytochrome P-450 enzymes in adult human hepatocytes in primary culture. Mol. Pharmacol. 1993, 44, 707–715. [Google Scholar]
- Gramignoli, R.; Tahan, V.; Dorko, K.; Skvorak, K.; Zhao, W.; Venkataramanan, R.; Geller, D.; Fox, I.J.; Ellis, E.C.S.; Strom, S.C. Rapid-and-sensitive assessment of human hepatocyte function. Cell Transpl. 2014, 23, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U.A.; Zanger, U.M. Molecular mechanisms of genetic polymorphisms of drug metabolism. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 269–296. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.H.; Lu, A.Y. Interindividual variability in inhibition and induction of cytochrome P450 enzymes. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 535–567. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.R. The effects of diet, aging and disease-states on presystemic elimination and oral drug bioavailability in humans. Adv. Drug Deliv. Rev. 1997, 27, 129–159. [Google Scholar] [CrossRef]
- Sunman, J.A.; Hawke, R.L.; LeCluyse, E.L.; Kashuba, A.D. Kupffer cell-mediated IL-2 suppression of CYP3A activity in human hepatocytes. Drug Metab. Dispos. Biol. Fate Chem. 2004, 32, 359–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramignoli, R.; Tahan, V.; Dorko, K.; Skvorak, K.J.; Hansel, M.C.; Zao, W.; Venkataramanan, R.; Ellis, E.C.; Jorns, C.; Ericzon, B.G.; et al. New potential cell source for hepatocyte transplantation: Discarded livers from metabolic disease liver transplants. Stem Cell Res. 2013, 11, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, A.; Yarmush, M.L.; Roth, C.M. Evaluation of an in vitro model of hepatic inflammatory response by gene expression profiling. Tissue Eng. 2005, 11, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Berthiaume, F.; Yarmush, M.L. Long-term stable cultures of rat hepatocytes: An in vitro model to study acute and chronic hepatic inflammation. Tissue Eng. 2002, 8, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Tukey, R.H.; Strassburg, C.P. Human UDP-glucuronosyltransferases: Metabolism, expression, and disease. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Rios, G.R.; King, C.D.; Coffman, B.L.; Green, M.D.; Mojarrabi, B.; Mackenzie, P.I.; Tephly, T.R. Glucuronidation of catechol estrogens by expressed human UDP-glucuronosyltransferases (UGTs) 1A1, 1A3, and 2B7. Toxicol. Sci. Off. J. Soc. Toxicol. 1998, 45, 52–57. [Google Scholar]
- Court, M.H.; Duan, S.X.; von Moltke, L.L.; Greenblatt, D.J.; Patten, C.J.; Miners, J.O.; Mackenzie, P.I. Interindividual variability in acetaminophen glucuronidation by human liver microsomes: Identification of relevant acetaminophen UDP-glucuronosyltransferase isoforms. J. Pharmacol. Exp. Ther. 2001, 299, 998–1006. [Google Scholar] [PubMed]
- Monshouwer, M.; Witkamp, R.F.; Nujmeijer, S.M.; Van Amsterdam, J.G.; Van Miert, A.S. Suppression of cytochrome P450- and UDP glucuronosyl transferase-dependent enzyme activities by proinflammatory cytokines and possible role of nitric oxide in primary cultures of pig hepatocytes. Toxicol. Appl. Pharmacol. 1996, 137, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Congiu, M.; Mashford, M.L.; Slavin, J.L.; Desmond, P.V. UDP glucuronosyltransferase mRNA levels in human liver disease. Drug Metab. Dispos. Biol. Fate Chem. 2002, 30, 129–134. [Google Scholar] [CrossRef]
- Engl, T.; Boost, K.A.; Leckel, K.; Beecken, W.D.; Jonas, D.; Oppermann, E.; Auth, M.K.; Schaudt, A.; Bechstein, W.O.; Blaheta, R.A. Phosphorylation of hepatocyte growth factor receptor and epidermal growth factor receptor of human hepatocytes can be maintained in a (3D) collagen sandwich culture system. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2004, 18, 527–532. [Google Scholar] [CrossRef]
- Sidhu, J.S.; Omiecinski, C.J. Modulation of xenobiotic-inducible cytochrome P450 gene expression by dexamethasone in primary rat hepatocytes. Pharmacogenetics 1995, 5, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Toritsuka, N.; Nonogaki, Y.; Hori, Y.; Sawada, S.; Shoun, H.; Motooka, S. Well maintained expression of CYP genes in sandwich-culturing hepatocytes: Quantitative analysis using real-time PCR method. Res. Commun. Mol. Pathol. Pharmacol. 2001, 110, 311–318. [Google Scholar] [PubMed]
- Kudryavtseva, E.I.; Engelhardt, N.V. Requirement of 3D extracellular network for maintenance of mature hepatocyte morphology and suppression of alpha-fetoprotein synthesis in vitro. Immunol. Lett. 2003, 90, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Kostrubsky, V.E.; Strom, S.C.; Hanson, J.; Urda, E.; Rose, K.; Burliegh, J.; Zocharski, P.; Cai, H.; Sinclair, J.F.; Sahi, J. Evaluation of hepatotoxic potential of drugs by inhibition of bile-acid transport in cultured primary human hepatocytes and intact rats. Toxicol. Sci. Off. J. Soc. Toxicol. 2003, 76, 220–228. [Google Scholar] [CrossRef]
- Sharma, S.; Ellis, E.C.; Gramignoli, R.; Dorko, K.; Tahan, V.; Hansel, M.; Mattison, D.R.; Caritis, S.N.; Hines, R.N.; Venkataramanan, R.; et al. Hepatobiliary Disposition of 17-OHPC and Taurocholate in Fetal Human Hepatocytes: A Comparison with Adult Human Hepatocytes. Drug Metab. Dispos. Biol. Fate Chem. 2013, 41, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Gramignoli, R.; Green, M.L.; Tahan, V.; Dorko, K.; Skvorak, K.J.; Marongiu, F.; Zao, W.; Venkataramanan, R.; Ellis, E.C.S.; Geller, D.; et al. Development and application of purified tissue dissociation enzyme mixtures for human hepatocyte isolation. Cell Transpl. 2012, 21, 1245–1260. [Google Scholar] [CrossRef]
- Gramignoli, R.; Dorko, K.; Tahan, V.; Skvorak, K.J.; Ellis, E.C.; Jorns, C.; Ericzon, B.G.; Fox, I.J.; Strom, S.C. Hypothermic storage of human hepatocytes for transplantation. Cell Transpl. 2014, 23, 1143–1151. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gramignoli, R.; Ranade, A.R.; Venkataramanan, R.; Strom, S.C. Effects of Pro-Inflammatory Cytokines on Hepatic Metabolism in Primary Human Hepatocytes. Int. J. Mol. Sci. 2022, 23, 14880. https://doi.org/10.3390/ijms232314880
Gramignoli R, Ranade AR, Venkataramanan R, Strom SC. Effects of Pro-Inflammatory Cytokines on Hepatic Metabolism in Primary Human Hepatocytes. International Journal of Molecular Sciences. 2022; 23(23):14880. https://doi.org/10.3390/ijms232314880
Chicago/Turabian StyleGramignoli, Roberto, Aarati R. Ranade, Raman Venkataramanan, and Stephen C. Strom. 2022. "Effects of Pro-Inflammatory Cytokines on Hepatic Metabolism in Primary Human Hepatocytes" International Journal of Molecular Sciences 23, no. 23: 14880. https://doi.org/10.3390/ijms232314880