
Complete and Prolonged Inhibition of Herpes Simplex Virus Type 1 Infection In Vitro by CRISPR/Cas9 and CRISPR/CasX Systems
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Characterization of the Genome HSV-1 Strain
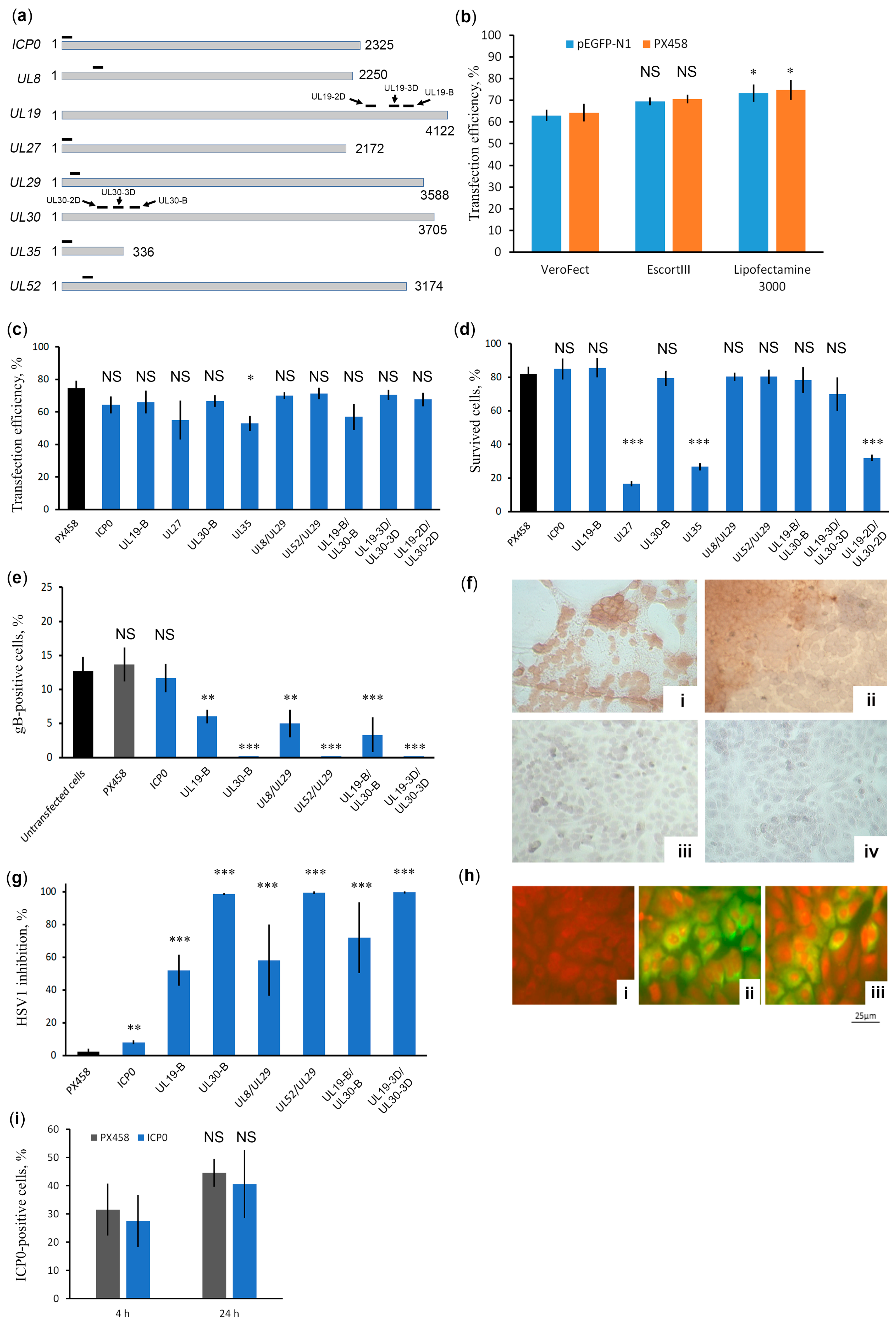
2.2. UL30 Identified as an Effective Anti-HSV-1 Target of the CRISPR/Cas9 System
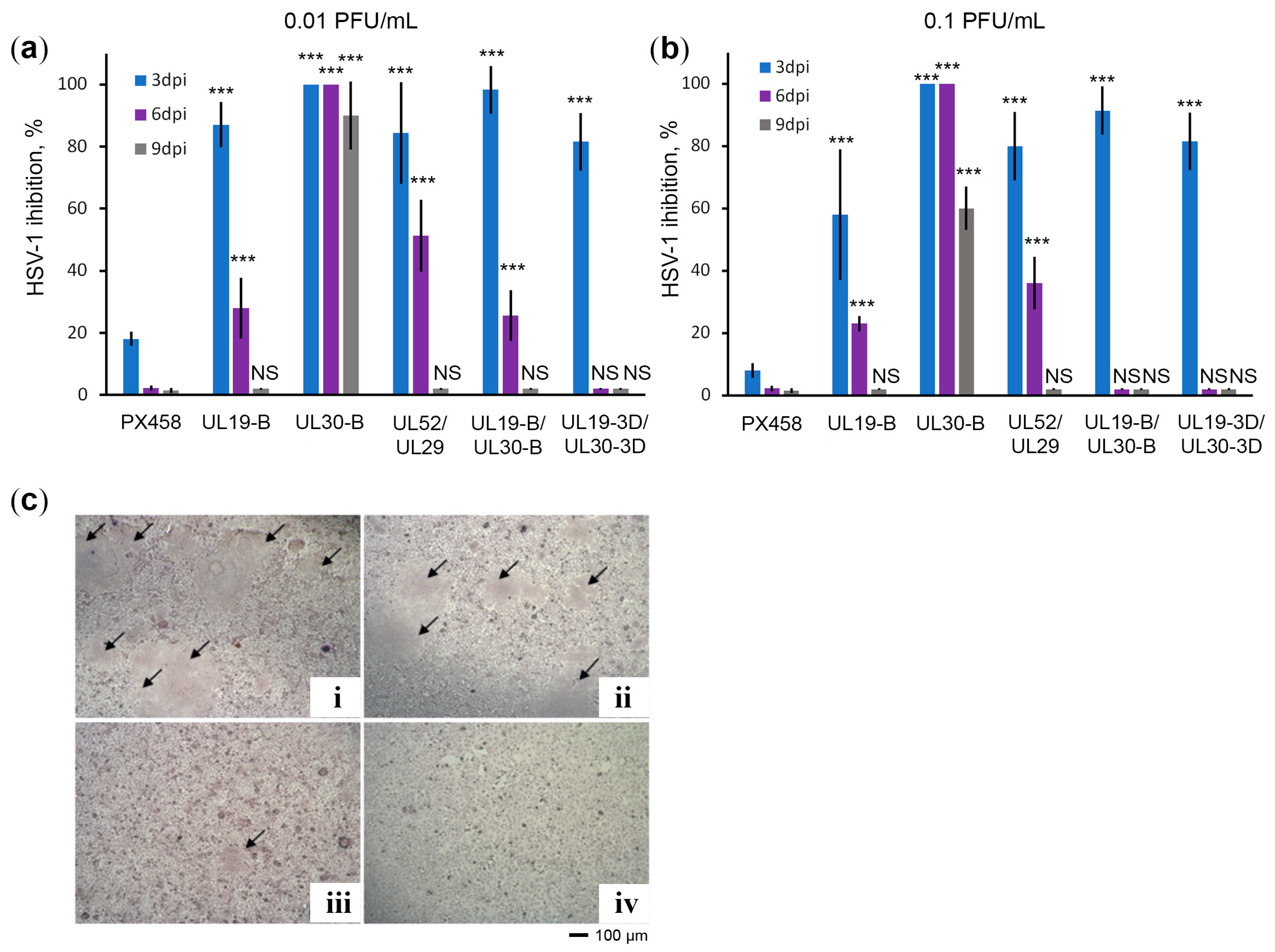
2.3. CRISPR/Cas9 System against UL30 Provides Long-Term Protection against HSV-1 Infection
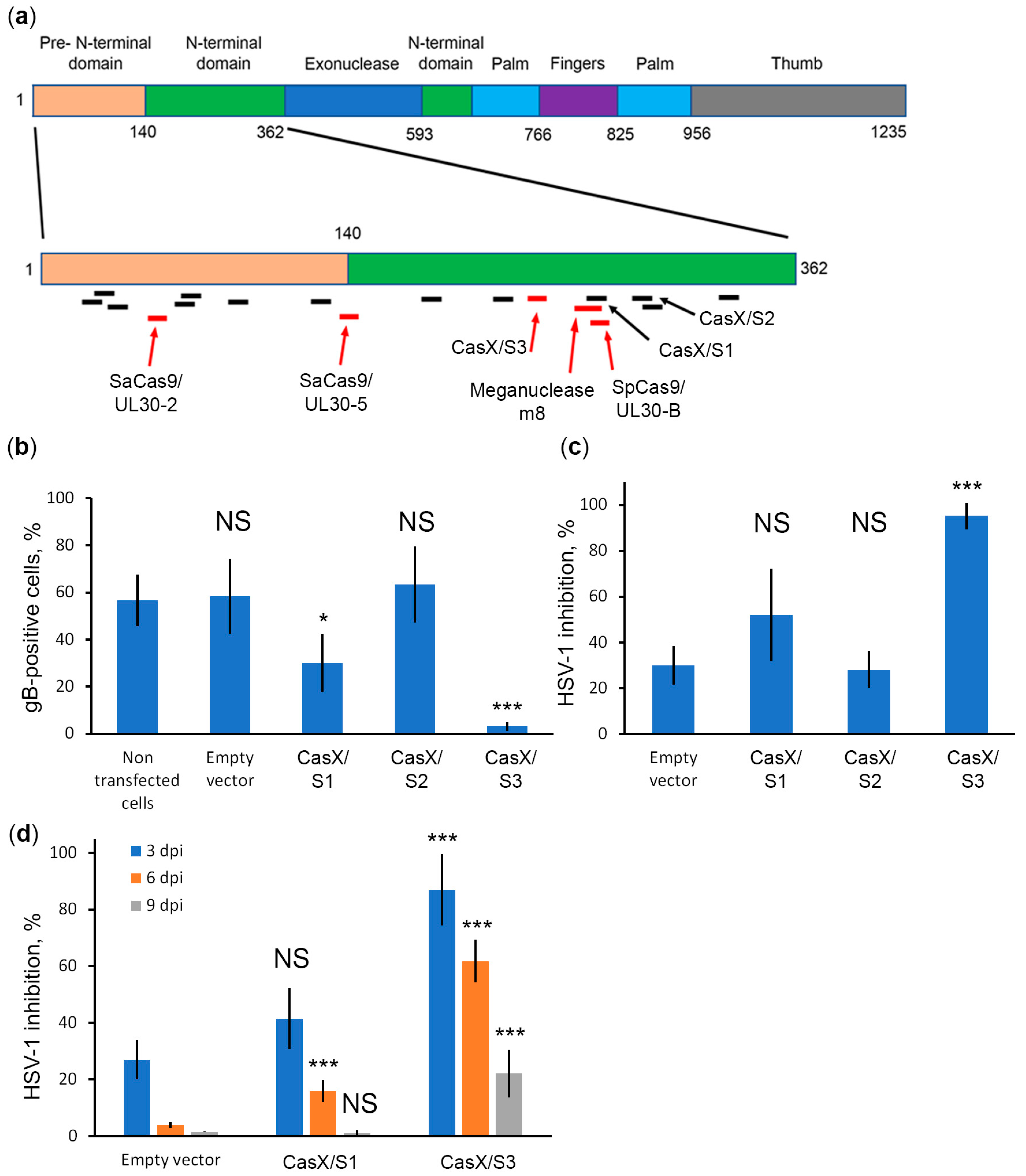
2.4. CRISPR/CasX System against UL30 Efficiently Protects Cells from HSV-1 Infection
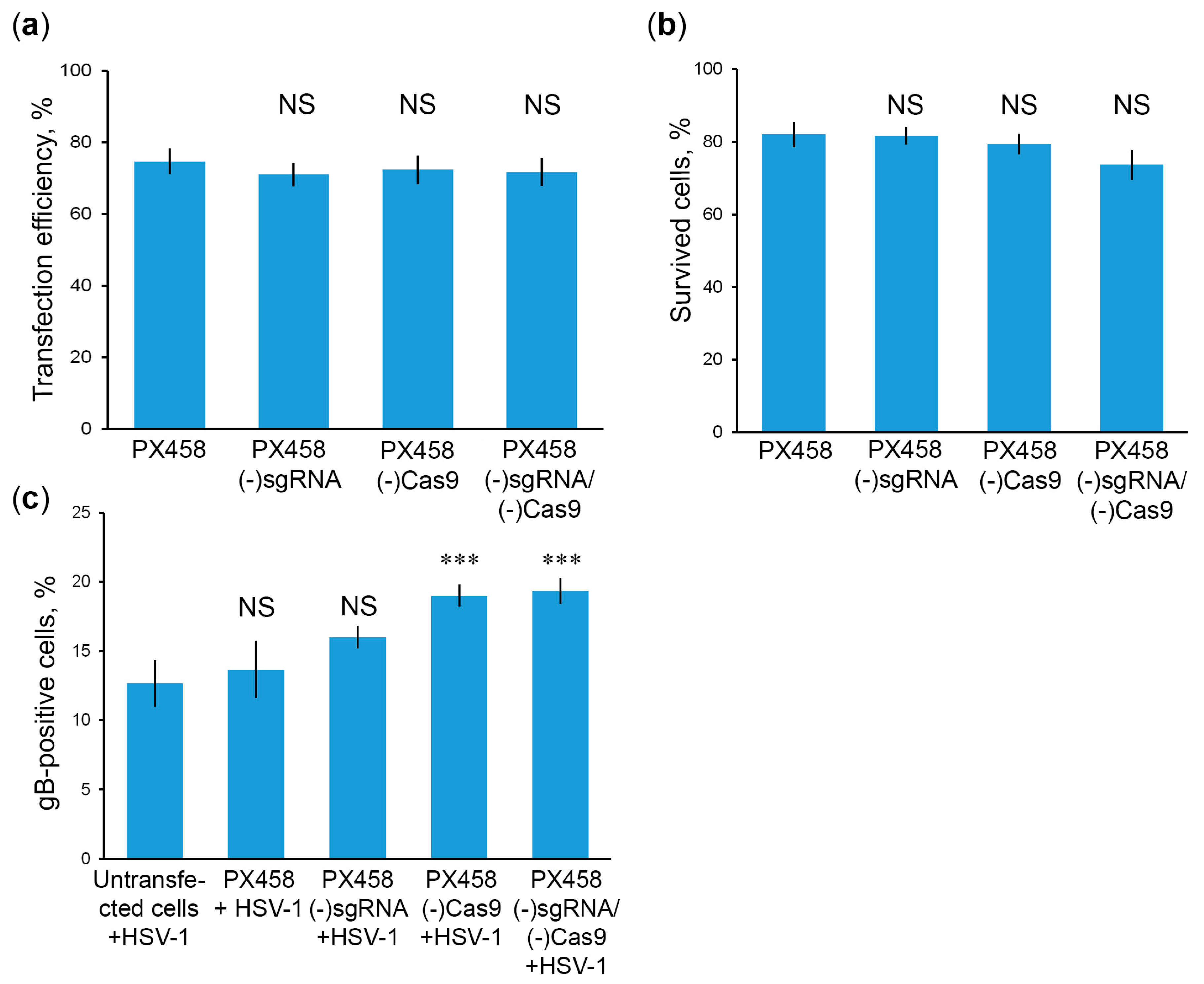
2.5. Cas9 without sgRNA Attenuates HSV-1 Infections
3. Discussion
4. Materials and Methods
4.1. Cells and Viral Strain
4.2. Purification of HSV-1
4.3. DNA Purification from HSV-1 Preparations
4.4. Sequencing of the HSV-1 Genome
4.5. Cloning of Spacers into Plasmids of the CRISPR/Cas System
4.6. Vero Cells Line Transfection
4.7. Evaluation of the Cytotoxicity of CRISPR/Cas Constructs
4.8. Detection of HSV-1
4.9. Estimation of HSV-1 Inhibition Rate
4.10. ICP0 Protein Detection
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining chronic viral infection. Cell 2009, 138, 30–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global and Regional Estimates of Prevalent and Incident Herpes Simplex Virus Type 1 Infections in 2012. PLoS ONE 2015, 10, e0140765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, A.M.; Agelidis, A.M.; Shukla, D. Pathogenesis of herpes simplex keratitis: The host cell response and ocular surface sequelae to infection and inflammation. Ocul. Surf. 2019, 17, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Mielcarska, M.B.; Skowronska, K.; Wyzewski, Z.; Toka, F.N. Disrupting Neurons and Glial Cells Oneness in the Brain-The Possible Causal Role of Herpes Simplex Virus Type 1 (HSV-1) in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 23, 242. [Google Scholar] [CrossRef] [PubMed]
- Jakhmola, S.; Upadhyay, A.; Jain, K.; Mishra, A.; Jha, H.C. Herpesviruses and the hidden links to Multiple Sclerosis neuropathology. J. Neuroimmunol. 2021, 358, 577636. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Viejo-Borbolla, A. Pathogenesis and virulence of herpes simplex virus. Virulence 2021, 12, 2670–2702. [Google Scholar] [CrossRef] [PubMed]
- Tuddenham, S.; Hamill, M.M.; Ghanem, K.G. Diagnosis and Treatment of Sexually Transmitted Infections: A Review. JAMA 2022, 327, 161–172. [Google Scholar] [CrossRef]
- Kushch, A.A.; Kisteneva, L.B.; Klimova, R.R.; Cheshik, S.G. The role of herpesviruses in development of diseases of the urogenital tract and infertility in women. Probl. Virol. 2021, 65, 317–325. [Google Scholar] [CrossRef]
- Busnadiego, I.; Abela, I.A.; Frey, P.M.; Hofmaenner, D.A.; Scheier, T.C.; Schuepbach, R.A.; Buehler, P.K.; Brugger, S.D.; Hale, B.G. Critically ill COVID-19 patients with neutralizing autoantibodies against type I interferons have increased risk of herpesvirus disease. PLoS Biol. 2022, 20, e3001709. [Google Scholar] [CrossRef]
- Chen, J.; Song, J.; Dai, L.; Post, S.R.; Qin, Z. SARS-CoV-2 infection and lytic reactivation of herpesviruses: A potential threat in the postpandemic era? J. Med. Virol. 2022, 94, 5103–5111. [Google Scholar] [CrossRef]
- da Silva Torres, M.K.; Bichara, C.D.A.; de Almeida, M.; Vallinoto, M.C.; Queiroz, M.A.F.; Vallinoto, I.; Dos Santos, E.J.M.; de Carvalho, C.A.M.; Vallinoto, A.C.R. The Complexity of SARS-CoV-2 Infection and the COVID-19 Pandemic. Front. Microbiol. 2022, 13, 789882. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Gantt, S.; AST Infectious Diseases Community of Practice. Human herpesvirus 6, 7 and 8 in solid organ transplantation. Am. J. Transplant. 2013, 13 (Suppl. S4), 128–137. [Google Scholar] [CrossRef] [PubMed]
- Duver, F.; Weissbrich, B.; Eyrich, M.; Wolfl, M.; Schlegel, P.G.; Wiegering, V. Viral reactivations following hematopoietic stem cell transplantation in pediatric patients—A single center 11-year analysis. PLoS ONE 2020, 15, e0228451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Langin, M.; Reichart, B.; Kruger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; et al. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [Google Scholar] [CrossRef] [PubMed]
- Griffith, B.P.; Goerlich, C.E.; Singh, A.K.; Rothblatt, M.; Lau, C.L.; Shah, A.; Lorber, M.; Grazioli, A.; Saharia, K.K.; Hong, S.N.; et al. Genetically Modified Porcine-to-Human Cardiac Xenotransplantation. N. Engl. J. Med. 2022, 387, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.M.; Castillo, E.; Duarte, L.F.; Arriagada, J.; Corrales, N.; Farias, M.A.; Henriquez, A.; Agurto-Munoz, C.; Gonzalez, P.A. Current Antivirals and Novel Botanical Molecules Interfering with Herpes Simplex Virus Infection. Front. Microbiol. 2020, 11, 139. [Google Scholar] [CrossRef]
- Betz, U.A.; Fischer, R.; Kleymann, G.; Hendrix, M.; Rubsamen-Waigmann, H. Potent in vivo antiviral activity of the herpes simplex virus primase-helicase inhibitor BAY 57-1293. Antimicrob. Agents Chemother. 2002, 46, 1766–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gege, C.; Bravo, F.J.; Uhlig, N.; Hagmaier, T.; Schmachtenberg, R.; Elis, J.; Burger-Kentischer, A.; Finkelmeier, D.; Hamprecht, K.; Grunwald, T.; et al. A helicase-primase drug candidate with sufficient target tissue exposure affects latent neural herpes simplex virus infections. Sci. Transl. Med. 2021, 13, eabf8668. [Google Scholar] [CrossRef]
- Gege, C.; Kleymann, G. Helicase-primase inhibitors from Medshine Discovery Inc. (WO2018/127207 and WO2020/007355) for the treatment of herpes simplex virus infections—Structure proposal for Phaeno Therapeutics drug candidate HN0037. Expert Opin. Ther. Pat. 2022, 32, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Wald, A.; Corey, L.; Timmler, B.; Magaret, A.; Warren, T.; Tyring, S.; Johnston, C.; Kriesel, J.; Fife, K.; Galitz, L.; et al. Helicase-primase inhibitor pritelivir for HSV-2 infection. N. Engl. J. Med. 2014, 370, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Majewska, A.; Mlynarczyk-Bonikowska, B. 40 Years after the Registration of Acyclovir: Do We Need New Anti-Herpetic Drugs? Int. J. Mol. Sci. 2022, 23, 3431. [Google Scholar] [CrossRef] [PubMed]
- Shiraki, K.; Yasumoto, S.; Toyama, N.; Fukuda, H. Amenamevir, a Helicase-Primase Inhibitor, for the Optimal Treatment of Herpes Zoster. Viruses 2021, 13, 1547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, I.; Hsiao, Z.; Liu, F. Development of Genome Editing Approaches against Herpes Simplex Virus Infections. Viruses 2021, 13, 338. [Google Scholar] [CrossRef]
- Xiao, J.; Deng, J.; Zhang, Q.; Ma, P.; Lv, L.; Zhang, Y.; Li, C.; Zhang, Y. Targeting human cytomegalovirus IE genes by CRISPR/Cas9 nuclease effectively inhibits viral replication and reactivation. Arch. Virol. 2020, 165, 1827–1835. [Google Scholar] [CrossRef]
- van Diemen, F.R.; Kruse, E.M.; Hooykaas, M.J.; Bruggeling, C.E.; Schurch, A.C.; van Ham, P.M.; Imhof, S.M.; Nijhuis, M.; Wiertz, E.J.; Lebbink, R.J. CRISPR/Cas9-Mediated Genome Editing of Herpesviruses Limits Productive and Latent Infections. PLoS Pathog. 2016, 12, e1005701. [Google Scholar] [CrossRef] [Green Version]
- Bokhoven, M.; Stephen, S.L.; Knight, S.; Gevers, E.F.; Robinson, I.C.; Takeuchi, Y.; Collins, M.K. Insertional gene activation by lentiviral and gammaretroviral vectors. J. Virol. 2009, 83, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranzani, M.; Cesana, D.; Bartholomae, C.C.; Sanvito, F.; Pala, M.; Benedicenti, F.; Gallina, P.; Sergi, L.S.; Merella, S.; Bulfone, A.; et al. Lentiviral vector-based insertional mutagenesis identifies genes associated with liver cancer. Nat. Methods 2013, 10, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Roehm, P.C.; Shekarabi, M.; Wollebo, H.S.; Bellizzi, A.; He, L.; Salkind, J.; Khalili, K. Inhibition of HSV-1 Replication by Gene Editing Strategy. Sci. Rep. 2016, 6, 23146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Ling, S.; Wang, D.; Dai, Y.; Jiang, H.; Zhou, X.; Paludan, S.R.; Hong, J.; Cai, Y. Targeting herpes simplex virus with CRISPR-Cas9 cures herpetic stromal keratitis in mice. Nat. Biotechnol. 2021, 39, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Tee, L.Y.; Wang, X.G.; Huang, Q.S.; Yang, S.H. Off-target Effects in CRISPR/Cas9-mediated Genome Engineering. Mol. Ther. Nucleic Acids 2015, 4, e264. [Google Scholar] [CrossRef] [PubMed]
- Karpov, D.S.; Karpov, V.L.; Klimova, R.R.; Demidova, N.A.; Kushch, A.A. A Plasmid-Expressed CRISPR/Cas9 System Suppresses Replication of HSV Type I in a Vero Cell Culture. Mol. Biol. 2019, 53, 91–100. [Google Scholar] [CrossRef]
- Khodadad, N.; Fani, M.; Jamehdor, S.; Nahidsamiei, R.; Makvandi, M.; Kaboli, S.; Teimoori, A.; Thekkiniath, J. A knockdown of the herpes simplex virus type-1 gene in all-in-one CRISPR vectors. Folia Histochem. Cytobiol. 2020, 58, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoblov, M.Y.; Lavrov, A.V.; Bragin, A.G.; Zubtsov, D.A.; Andronova, V.L.; Galegov, G.A.; Skoblov, Y.S. The genome nucleotide sequence of herpes simplex virus 1 strain L2. Russ. J. Bioorganic Chem. 2017, 43, 140–142. [Google Scholar] [CrossRef]
- Aubert, M.; Strongin, D.E.; Roychoudhury, P.; Loprieno, M.A.; Haick, A.K.; Klouser, L.M.; Stensland, L.; Huang, M.L.; Makhsous, N.; Tait, A.; et al. Gene editing and elimination of latent herpes simplex virus in vivo. Nat. Commun. 2020, 11, 4148. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.S.; Neuhausser, W.M.; Eggan, P.; Angelova, M.; Kirchner, R.; Eggan, K.C.; Knipe, D.M. Herpesviral lytic gene functions render the viral genome susceptible to novel editing by CRISPR/Cas9. Elife 2019, 8, 51662. [Google Scholar] [CrossRef] [PubMed]
- Dogrammatzis, C.; Waisner, H.; Kalamvoki, M. “Non-Essential” Proteins of HSV-1 with Essential Roles In Vivo: A Comprehensive Review. Viruses 2020, 13, 17. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, N.; Berkhout, B.; Das, A.T. CRISPR-Cas9 Can Inhibit HIV-1 Replication but NHEJ Repair Facilitates Virus Escape. Mol. Ther. 2016, 24, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Fan, M.; Das, A.T.; Herrera-Carrillo, E.; Berkhout, B. Extinction of all infectious HIV in cell culture by the CRISPR-Cas12a system with only a single crRNA. Nucleic Acids Res. 2020, 48, 5527–5539. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Dybas, J.M.; Hughes, J.; Weitzman, M.D.; Boutell, C. The HSV-1 ubiquitin ligase ICP0: Modifying the cellular proteome to promote infection. Virus Res. 2020, 285, 198015. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Rinsma, M.; Janssen, J.M.; Liu, J.; Maggio, I.; Goncalves, M.A. Probing the impact of chromatin conformation on genome editing tools. Nucleic Acids Res. 2016, 44, 6482–6492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daer, R.M.; Cutts, J.P.; Brafman, D.A.; Haynes, K.A. The Impact of Chromatin Dynamics on Cas9-Mediated Genome Editing in Human Cells. ACS Synth. Biol. 2017, 6, 428–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Orlova, N.; Oakes, B.L.; Ma, E.; Spinner, H.B.; Baney, K.L.M.; Chuck, J.; Tan, D.; Knott, G.J.; Harrington, L.B.; et al. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nature 2019, 566, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Rube, H.T.; Vakulskas, C.A.; Behlke, M.A.; Bussemaker, H.J.; Pufall, M.A. Systematic in vitro profiling of off-target affinity, cleavage and efficiency for CRISPR enzymes. Nucleic Acids Res. 2020, 48, 5037–5053. [Google Scholar] [CrossRef] [Green Version]
- Hayes, R.P.; Heo, M.R.; Mason, M.; Reid, J.; Burlein, C.; Armacost, K.A.; Tellers, D.M.; Raheem, I.; Shaw, A.W.; Murray, E.; et al. Structural understanding of non-nucleoside inhibition in an elongating herpesvirus polymerase. Nat. Commun. 2021, 12, 3040. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Koo, T.; Jee, H.G.; Cho, H.Y.; Lee, G.; Lim, D.G.; Shin, H.S.; Kim, J.S. CRISPR RNAs trigger innate immune responses in human cells. Genome Res. 2018, 28, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. CRISPR/Cas9-Based Antiviral Strategy: Current Status and the Potential Challenge. Molecules 2019, 24, 1349. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Berkhout, B.; Herrera-Carrillo, E. A combinatorial CRISPR-Cas12a attack on HIV DNA. Mol. Ther. Methods Clin. Dev. 2022, 25, 43–51. [Google Scholar] [CrossRef]
- Artusi, S.; Nadai, M.; Perrone, R.; Biasolo, M.A.; Palu, G.; Flamand, L.; Calistri, A.; Richter, S.N. The Herpes Simplex Virus-1 genome contains multiple clusters of repeated G-quadruplex: Implications for the antiviral activity of a G-quadruplex ligand. Antiviral Res. 2015, 118, 123–131. [Google Scholar] [CrossRef]
- Hoque, M.E.; Mustafa, G.; Basu, S.; Balci, H. Encounters between Cas9/dCas9 and G-Quadruplexes: Implications for Transcription Regulation and Cas9-Mediated DNA Cleavage. ACS Synth. Biol. 2021, 10, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Danastas, K.; Miranda-Saksena, M.; Cunningham, A.L. Herpes Simplex Virus Type 1 Interactions with the Interferon System. Int. J. Mol. Sci. 2020, 21, 5150. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qian, F.; Yang, J.; Liu, Y.; Dong, F.; Xu, C.; Sun, B.; Chen, B.; Xu, X.; Li, Y.; et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat. Commun. 2017, 8, 15179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, J.M.; Hopkins, F.F.; Chavez, A.; Diallo, M.; Chase, M.R.; Gerrick, E.R.; Pritchard, J.R.; Church, G.M.; Rubin, E.J.; Sassetti, C.M.; et al. Programmable transcriptional repression in mycobacteria using an orthogonal CRISPR interference platform. Nat. Microbiol. 2017, 2, 16274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Voigt, C.A. Engineered dCas9 with reduced toxicity in bacteria: Implications for genetic circuit design. Nucleic Acids Res. 2018, 46, 11115–11125. [Google Scholar] [CrossRef] [Green Version]
- Rondelli, V.; Helmy, S.; Passignani, G.; Parisse, P.; Di Silvestre, D. Integrated Strategies for a Holistic View of Extracellular Vesicles. ACS Omega 2022, 7, 19058–19069. [Google Scholar] [CrossRef] [PubMed]
- Lainscek, D.; Kadunc, L.; Keber, M.M.; Bratkovic, I.H.; Romih, R.; Jerala, R. Delivery of an Artificial Transcription Regulator dCas9-VPR by Extracellular Vesicles for Therapeutic Gene Activation. ACS Synth. Biol. 2018, 7, 2715–2725. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, Z.H. Purification of Herpesvirus Virions and Capsids. Bio Protoc. 2014, 4, e1193. [Google Scholar] [CrossRef] [Green Version]
- Shipley, M.M.; Rathbun, M.M.; Szpara, M.L. Oligonucleotide Enrichment of HSV-1 Genomic DNA from Clinical Specimens for Use in High-Throughput Sequencing. Methods Mol. Biol. 2020, 2060, 199–217. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Description | References |
|---|---|---|
| ICP0 | RING-type E3 ubiquitin ligase, acts on initial stages of infection, or during HSV-1 reactivation, helps to evade cellular antiviral response | [29] |
| UL8 | DNA helicase/primase complex-associated protein | [26,30] |
| UL19 | Major capsid protein, forms an icosahedral capsid with a T = 16 symmetry consisting of 162 capsomers. | [36] |
| UL27 | gB, envelope glycoprotein that forms spikes at the surface of virion envelope, essential for the initial attachment to the host cell receptors, involved in fusion of viral and cellular membranes, together with gK induces syncytia formation | [26,33] |
| UL29 | ssDNA-binding protein | [26,30,37] |
| UL30 | Large catalytic subunit of DNA-directed DNA polymerase, essential for HSV-1 genome replication | [26,36,37] |
| UL35 | Small capsomere-interacting protein, participates in the assembly of the infectious particles, forms a layer between the capsid and the tegument | [38] |
| UL52 | DNA primase, essential for HSV-1 genome replication | [26] |
| Name of the Oligonucleotide | Sequence (5′→3′) | Purpose |
|---|---|---|
| ISP0_F | CACCTCCCTGCGACCGAGACCTGC | Cloning spacers for CRISPR/Cas9 system |
| ISP0_R | AAACGCAGGTCTCGGTCGCAGGGA | |
| ICP0_ch_F | AACTCGTGGGCGCTGATTGA | |
| ICP0_ch_R | TCGTCGCTCCCCCCGTCCTCT | |
| UL27_F | CACCGGTGCCGGTGGTTCGTCGTA | |
| UL27_R | AAACTACGACGAACCACCGGCACC | |
| UL35_F | CACCGTGAAATTGCGGGACGGCCAT | |
| UL35_R | AAACATGGCCGTCCCGCAATTTCAC | |
| UL35_ch_F | AAGGACGCACCGCCGCCCTA | |
| UL35_ch_R | CGGCCCCTTGGGTGCCCTGG | |
| UL27_ch_F | GGAGCCGCCGACGCCACCAGG | |
| UL27_ch_R | CGTACGACTCCGACTGTCCGCT | |
| U6-ch-F | CGATACAAGGCTGTTAGAGAGA | |
| UL19-B-F | ATATAGAAGACCTCACCGTAGTTGACGTCGGTCGACACGTTTTAGAGCTAGAAATAGCAAG | |
| UL19-2D-F | ATATAGAAGACCTCACCGGACCGCGTTCCGCAGGTACAGTTTTAGAGCTAGAAATAGCAAG | |
| UL19-3D-F | ATATAGAAGACCTCACCGTAAACTCACACACGGCATCCGTTTTAGAGCTAGAAATAGCAAG | |
| UL30-B-R | TAGAGGAAGACCCAAACCTTCGGACGTAGACGCGGTACGGTGTTTCGTCCTTTCCAC | |
| UL30-2D-R | TAGAGGAAGACCCAAACCGTGCCGTAAACGTGAACGGCGGTGTTTCGTCCTTTCCAC | |
| UL30-3D-R | TAGAGGAAGACCCAAACGGCGCGTCGTTCCGCGGCATCGGTGTTTCGTCCTTTCCAC | |
| UL30-BsgRNA-F | CACCGTACCGCGTCTACGTCCGAAG | |
| UL30-BsgRNA-R | AAACCTTCGGACGTAGACGCGGTAC | |
| UL19-B-sgRNA-F | CACCGTAGTTGACGTCGGTCGACAC | |
| UL19-B-sgRNA-R | AAACGTGTCGACCGACGTCAACTAC | |
| U6-PciI-F | TTTTGCTCACATGTGAGGGCCTATTTCC | Cloning spacers for CRISPR/CasX system |
| CasX-S1-sgRNA-R | TATGTAACGGGTACCAAAAAAAATGTTTTACCGCGTCTACGTCCTTTGATGCGTTTTACTTATCGGTTTC | |
| CasX-S2-sgRNA-R | TATGTAACGGGTACCAAAAAAAACAACTTCTGCCCGGCCATCACTTTGATGCGTTTTACTTATCGGTTTC | |
| CasX-S3-sgRNA-R | TATGTAACGGGTACCAAAAAAAAGCGCTCCACCACCTCCGCCTCTTTGATGCGTTTTACTTATCGGTTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpov, D.S.; Demidova, N.A.; Kulagin, K.A.; Shuvalova, A.I.; Kovalev, M.A.; Simonov, R.A.; Karpov, V.L.; Snezhkina, A.V.; Kudryavtseva, A.V.; Klimova, R.R.; et al. Complete and Prolonged Inhibition of Herpes Simplex Virus Type 1 Infection In Vitro by CRISPR/Cas9 and CRISPR/CasX Systems. Int. J. Mol. Sci. 2022, 23, 14847. https://doi.org/10.3390/ijms232314847
Karpov DS, Demidova NA, Kulagin KA, Shuvalova AI, Kovalev MA, Simonov RA, Karpov VL, Snezhkina AV, Kudryavtseva AV, Klimova RR, et al. Complete and Prolonged Inhibition of Herpes Simplex Virus Type 1 Infection In Vitro by CRISPR/Cas9 and CRISPR/CasX Systems. International Journal of Molecular Sciences. 2022; 23(23):14847. https://doi.org/10.3390/ijms232314847
Chicago/Turabian StyleKarpov, Dmitry S., Natalia A. Demidova, Kirill A. Kulagin, Anastasija I. Shuvalova, Maxim A. Kovalev, Ruslan A. Simonov, Vadim L. Karpov, Anastasiya V. Snezhkina, Anna V. Kudryavtseva, Regina R. Klimova, and et al. 2022. "Complete and Prolonged Inhibition of Herpes Simplex Virus Type 1 Infection In Vitro by CRISPR/Cas9 and CRISPR/CasX Systems" International Journal of Molecular Sciences 23, no. 23: 14847. https://doi.org/10.3390/ijms232314847