
Radiosensitizing Effect of Trabectedin on Human Soft Tissue Sarcoma Cells

 , , , , , ,
, , , , , ,  and
and 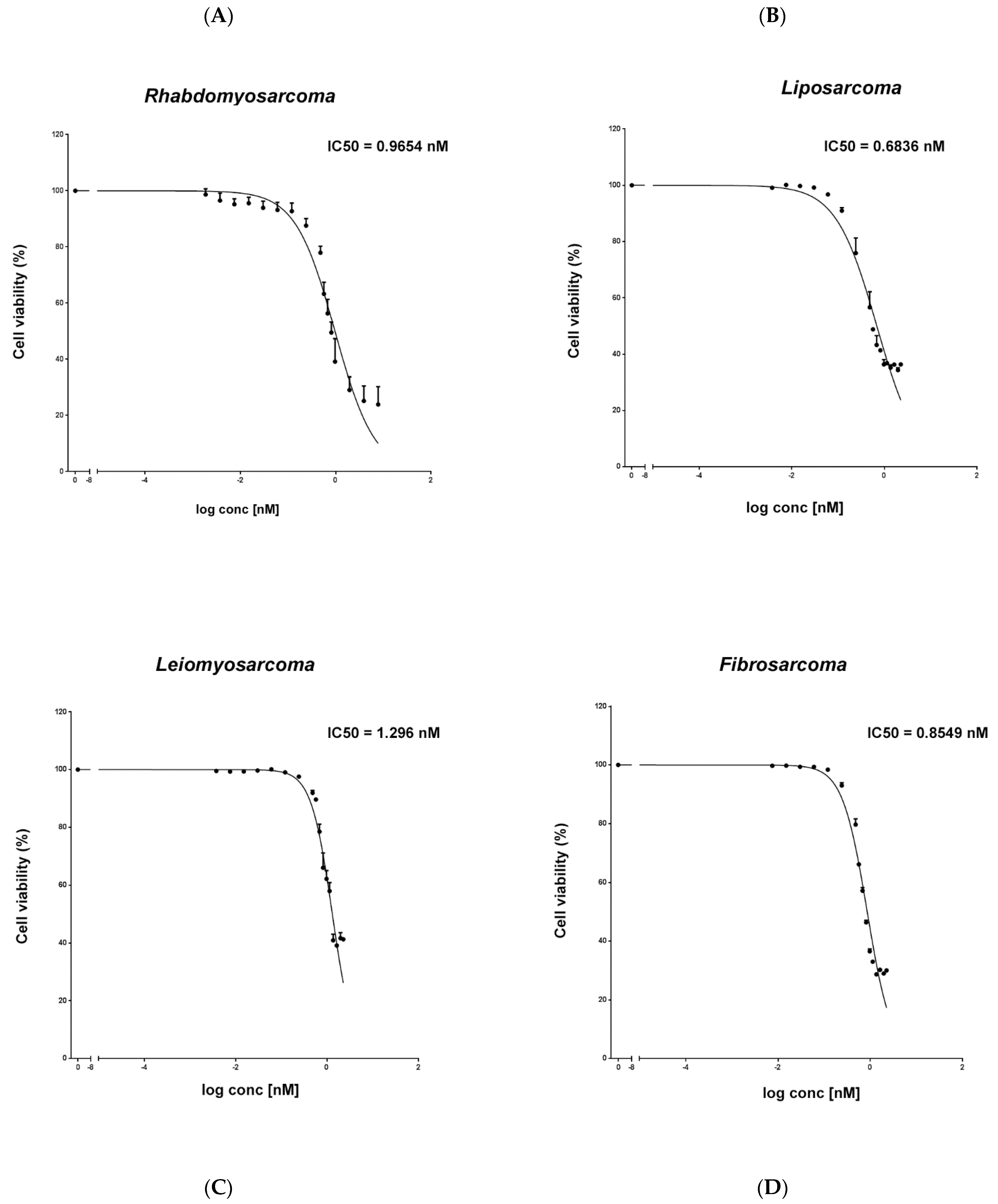{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Trabectedin Is Cytotoxic in the Low Nanomolar Range in STS Cells
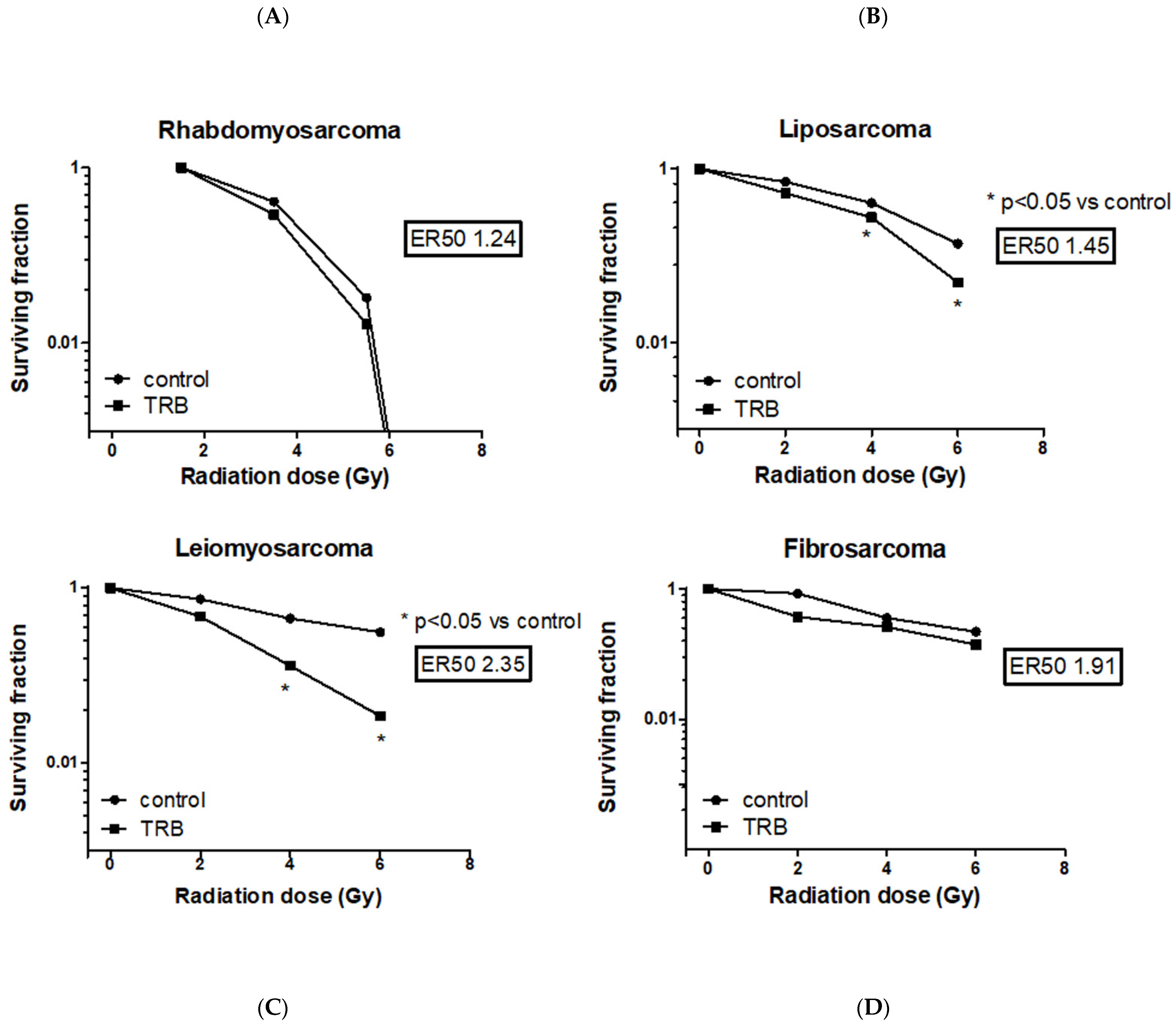
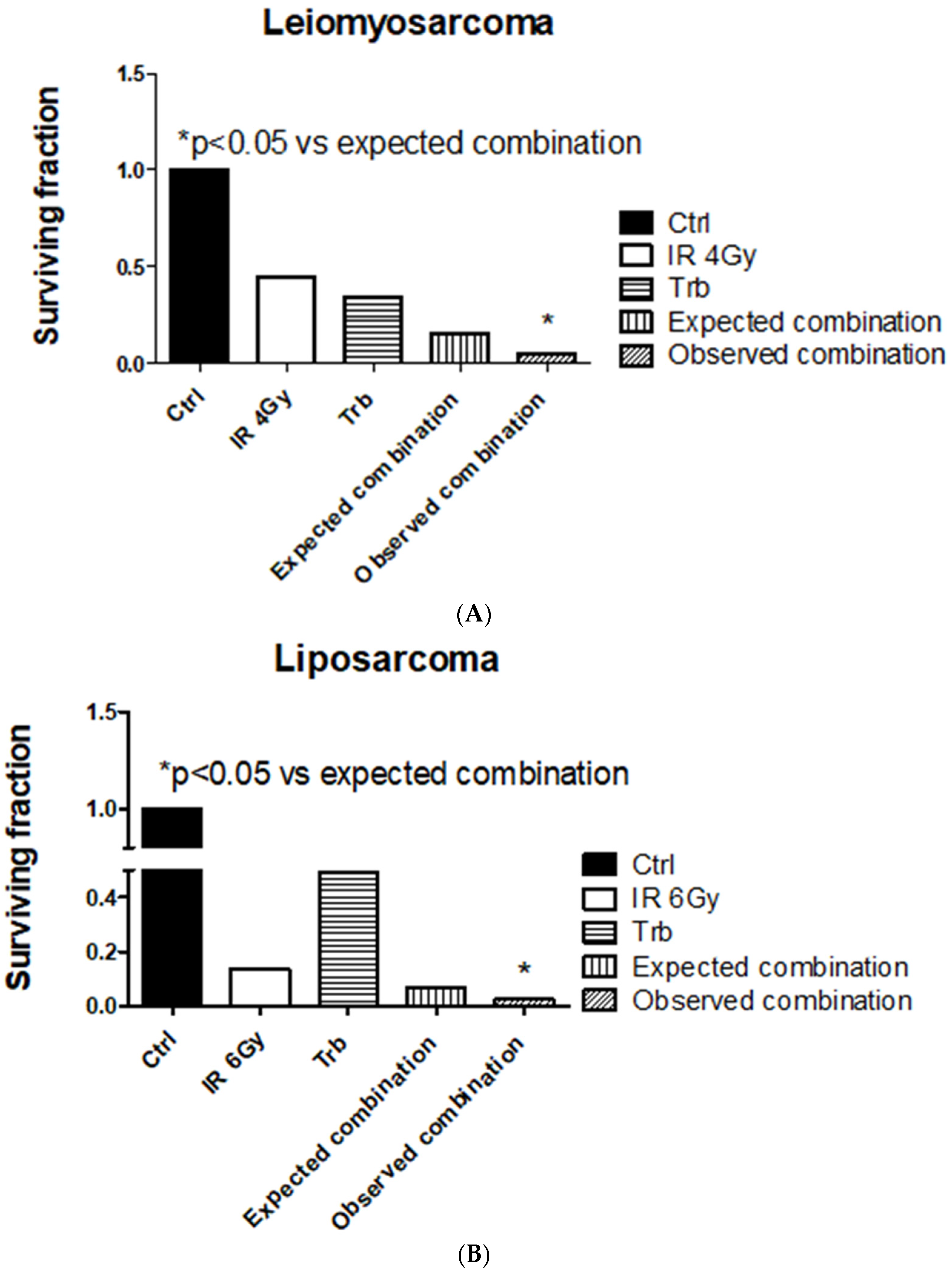
2.2. Trabectedin Improves Cytotoxic Effect of IR
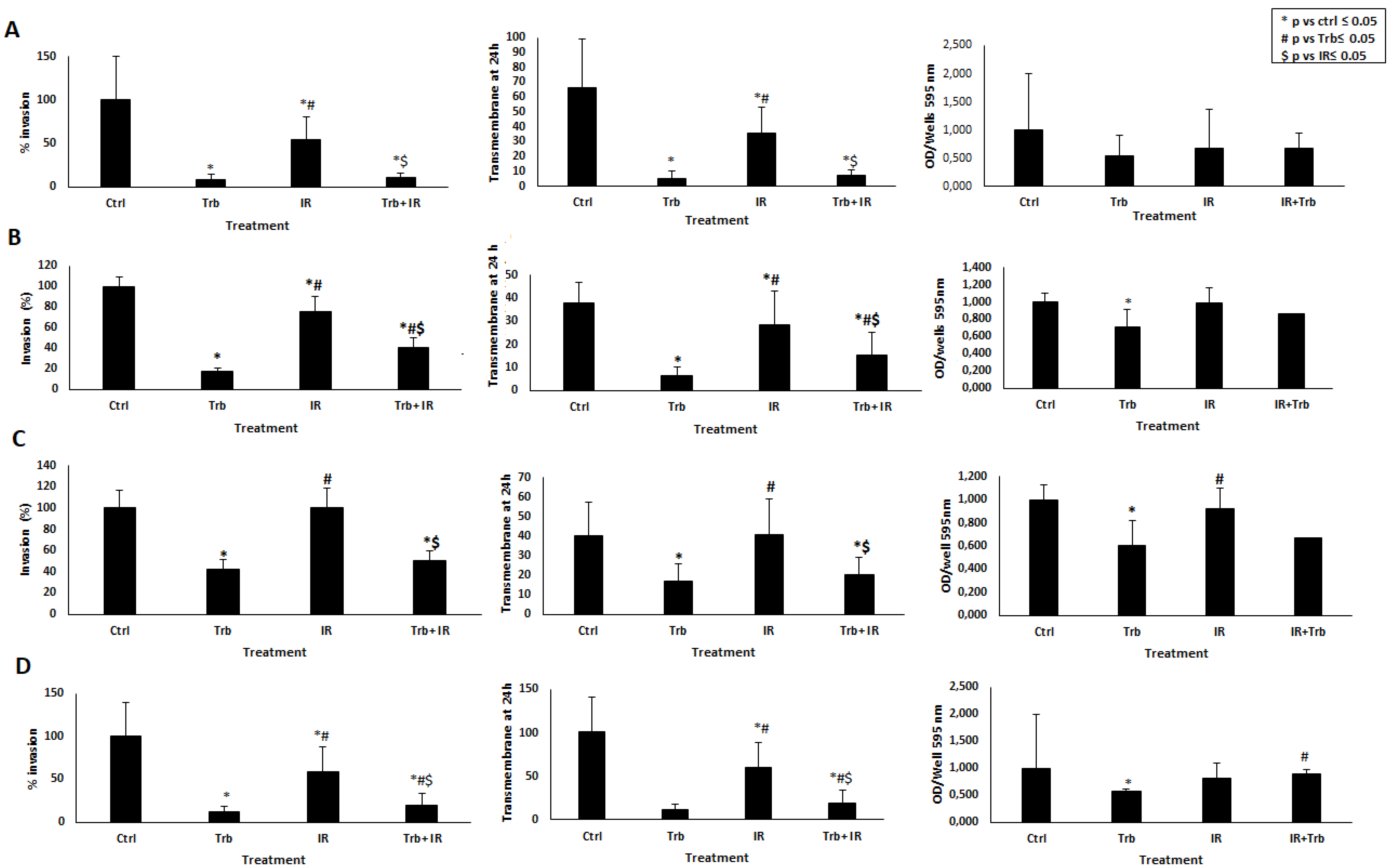
2.3. Trabectedin Significantly Reduces Invasiveness Caused by IR
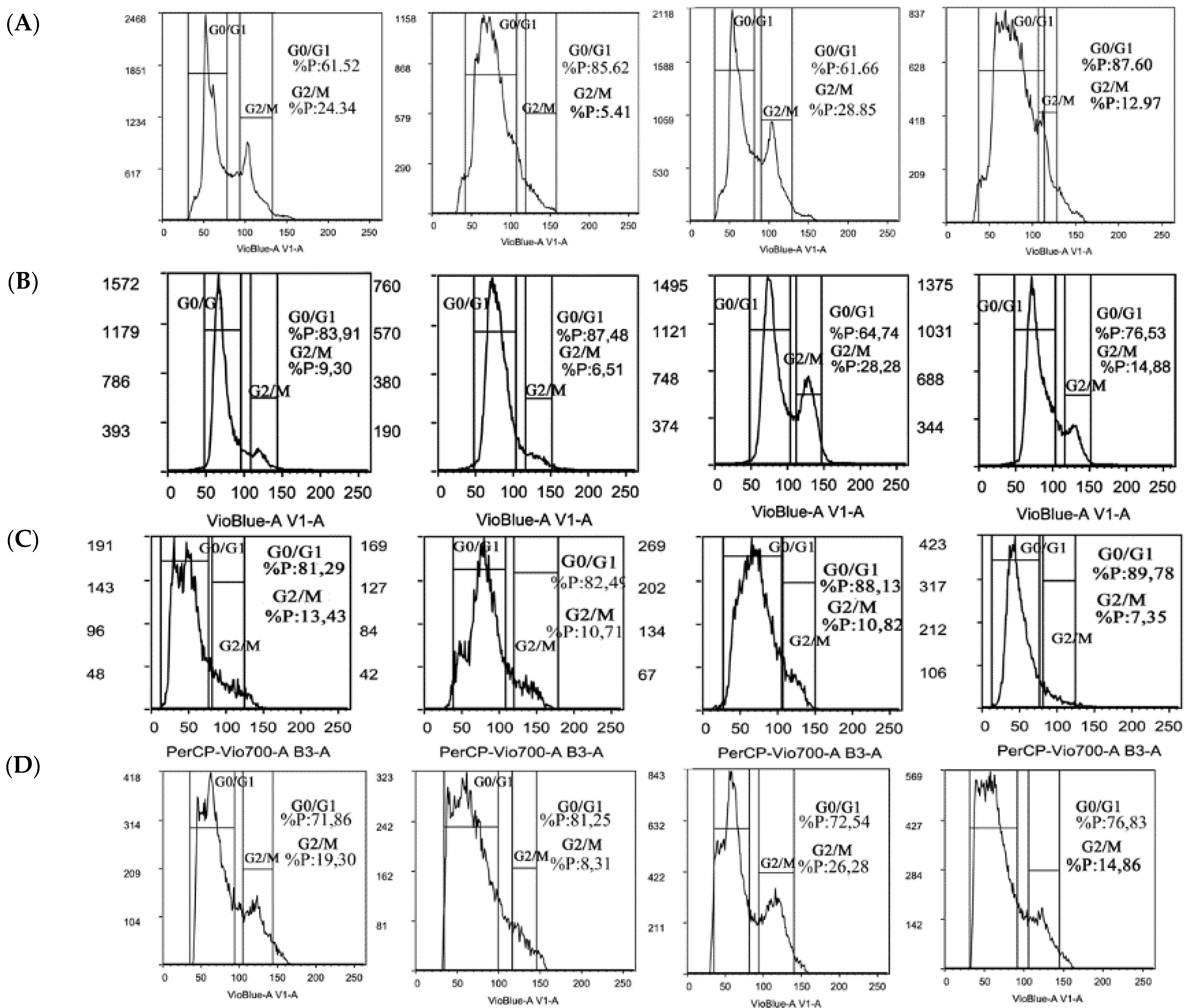
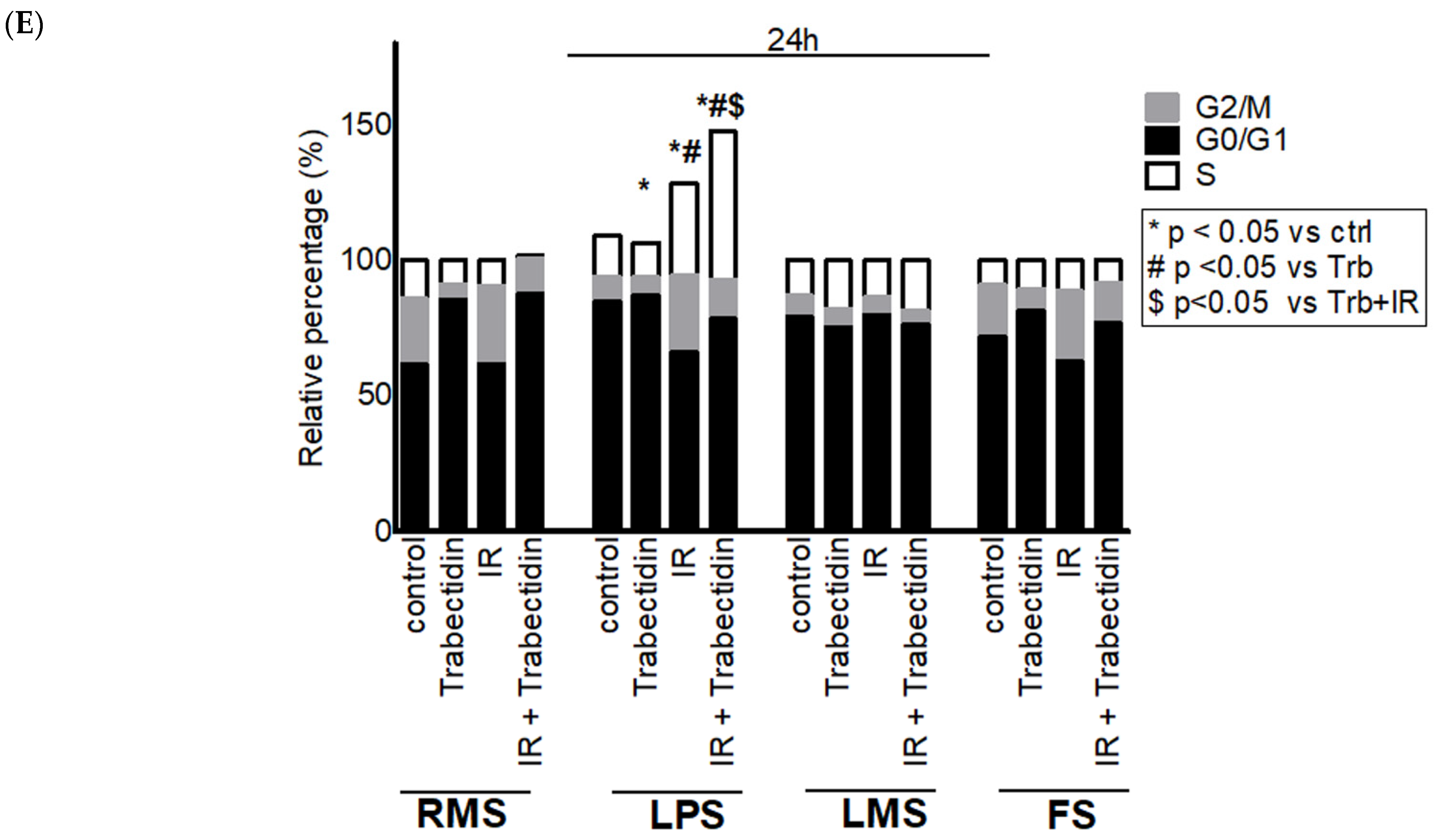
2.4. Trabectedin Affects Cell Cycle Progression of STS Cells
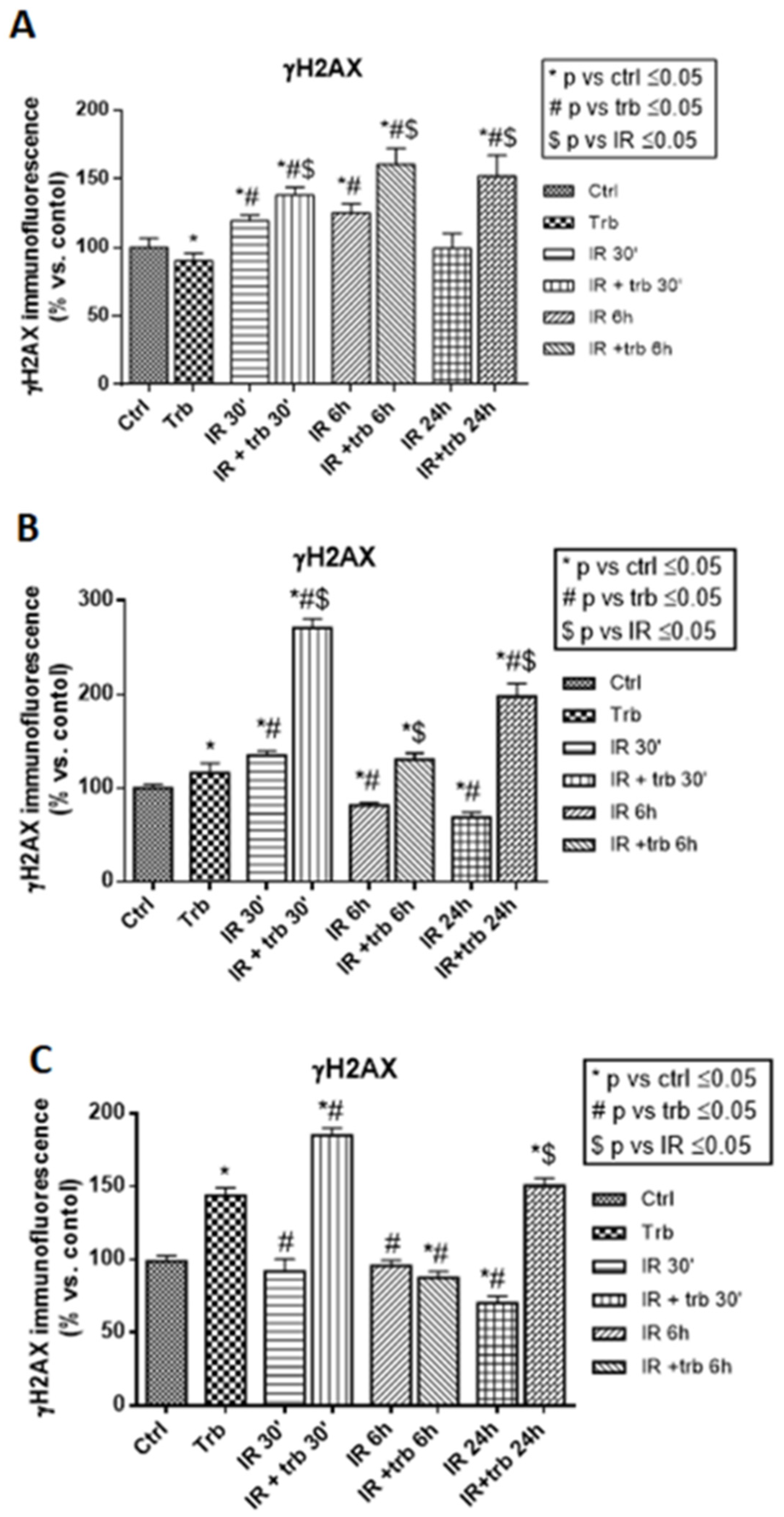
2.5. Trabectedin Combined with IR Significantly Increased the Number of γ-H2AX Foci
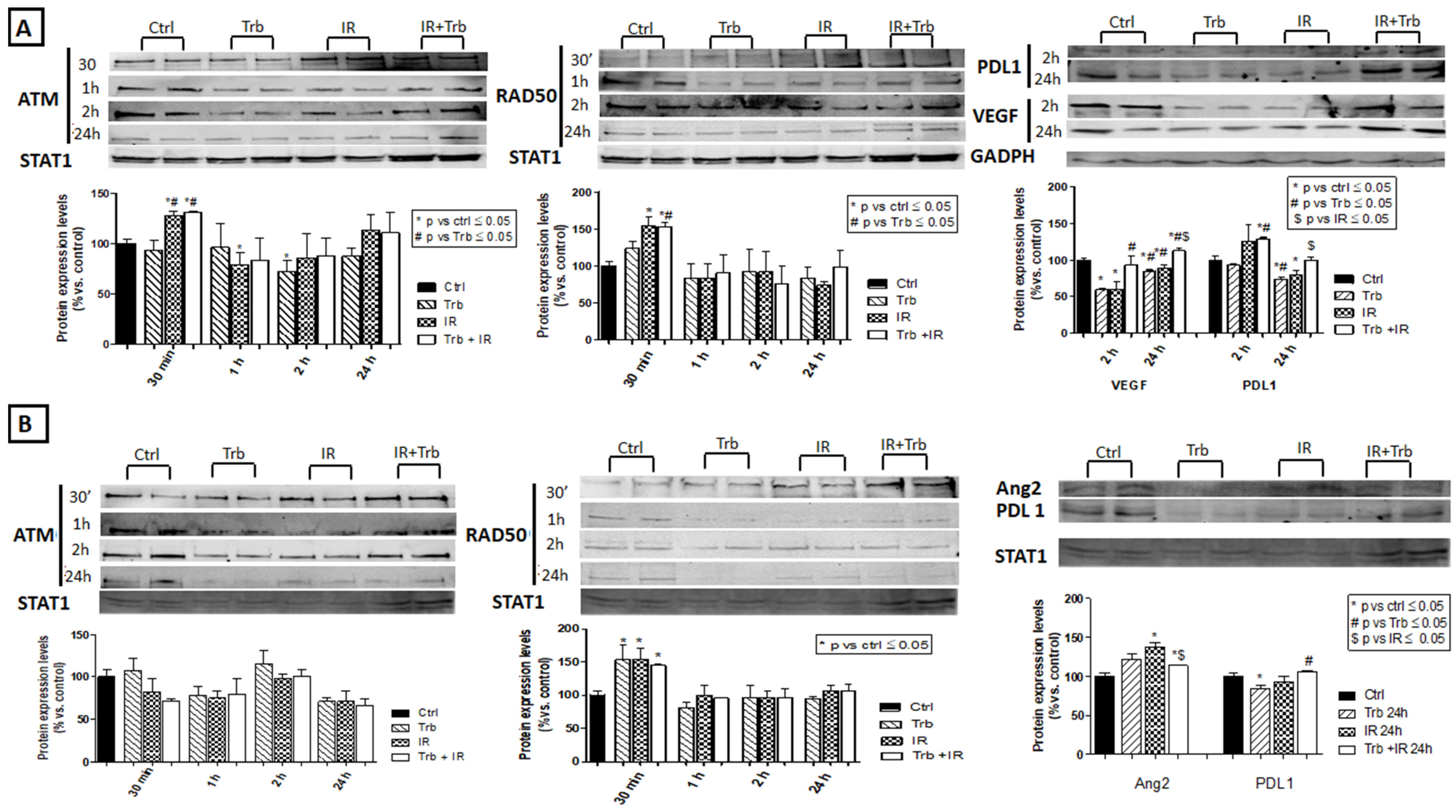
2.6. Trabectedin Combined with IR Affects the Expression of DNA Damage Response Proteins, Angiogenic Factors, and Immune Checkpoint Proteins
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Cultures
4.3. Cytotoxic Assay
4.4. Clonogenic Assays
4.5. Invasion Assay
4.6. Flow Cytometry Analysis
4.7. Immunofluorescence Analysis
4.8. Western Blot
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casali, P.G.; Abecassis, N.; Aro, H.T.; Bauer, S.; Biagini, R.; Bielack, S.; Bonvalot, S.; Boukovinas, I.; Bovee, J.V.M.G.; Brodowicz, T.; et al. Soft tissue and visceral sarcomas: ESMO-EURACAN Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2018, 29 (Suppl. 4), iv51–iv67, Erratum in Ann Oncol. 2018, 29 (Suppl. 4), iv268–iv269. [Google Scholar] [CrossRef] [PubMed]
- Demetri, G.D.; von Mehren, M.; Jones, R.L.; Hensley, M.L.; Schuetze, S.M.; Staddon, A.; Milhem, M.; Elias, A.; Ganjoo, K.; Tawbi, H.; et al. Efficacy and Safety of Trabectedin or Dacarbazine for Metastatic Liposarcoma or Leiomyosarcoma After Failure of Conventional Chemotherapy: Results of a Phase III Randomized Multicenter Clinical Trial. J. Clin. Oncol. 2016, 34, 786–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giandomenico, S.; Frapolli, R.; Bello, E.; Uboldi, S.; Licandro, S.A.; Marchini, S.; Beltrame, L.; Brich, S.; Mauro, V.; Tamborini, E.; et al. Mode of action of trabectedin in myxoid liposarcomas. Oncogene 2014, 33, 5201–5210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allavena, P.; Signorelli, M.; Chieppa, M.; Erba, E.; Bianchi, G.; Marchesi, F.; Olimpio, C.O.; Bonardi, C.; Garbi, A.; Lissoni, A.; et al. Anti-inflammatory properties of the novel antitumor agent yondelis (trabectedin): Inhibition of macrophage differentiation and cytokine production. Cancer Res. 2005, 65, 2964–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germano, G.; Frapolli, R.; Belgiovine, C.; Anselmo, A.; Pesce, S.; Liguori, M.; Erba, E.; Uboldi, S.; Zucchetti, M.; Pasqualini, F.; et al. Role of macrophage targeting in the antitumor activity of trabectedin. Cancer Cell 2013, 23, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Belgiovine, C.; Frapolli, R.; Liguori, M.; Digifico, E.; Colombo, F.S.; Meroni, M.; Allavena, P.; D’Incalci, M. Inhibition of tumor-associated macrophages by trabectedin improves the antitumor adaptive immunity in response to anti-PD-1 therapy. Eur. J. Immunol. 2021, 51, 2677–2686. [Google Scholar] [CrossRef]
- Manda, K.; Präkelt, T.; Schröder, T.; Kriesen, S.; Hildebrandt, G. Radiosensitizing effects of trabectedin on human A549 lung cancer cells and HT-29 colon cancer cells. Investig. New Drugs 2020, 38, 967–976. [Google Scholar] [CrossRef]
- Romero, J.; Zapata, I.; Córdoba, S.; Jimeno, J.M.; López-Martín, J.A.; Tercero, J.C.; De La Torre, A.; Vargas, J.A.; Molerón, R.; Sánchez-Prieto, R. In vitro radiosensitisation by trabectedin in human cancer cell lines. Eur. J. Cancer 2008, 44, 1726–1733. [Google Scholar] [CrossRef]
- Tavecchio, M.; Simone, M.; Erba, E.; Chiolo, I.; Liberi, G.; Foiani, M.; D’Incalci, M.; Damia, G. Role of homologous recombination in trabectedin-induced DNA damage. Eur. J. Cancer 2008, 44, 609–618. [Google Scholar] [CrossRef]
- Damia, G.; Silvestri, S.; Carrassa, L.; Filiberti, L.; Faircloth, G.T.; Liberi, G.; Foiani, M.; D’Incalci, M. Unique pattern of ET-743 activity in different cellular systems with defined deficiencies in DNA-repair pathways. Int. J. Cancer 2001, 92, 583–588. [Google Scholar] [CrossRef]
- Pautier, P.; Italiano, A.; Piperno-Neumann, S.; Chevreau, C.; Penel, N.; Firmin, N.; Boudou-Rouquette, P.; Bertucci, F.; Balleyguier, C.; Lebrun-Ly, V.; et al. Doxorubicin alone versus doxorubicin with trabectedin followed by trabectedin alone as first-line therapy for metastatic or unresectable leiomyosarcoma (LMS-04): A randomised, multicentre, open-label phase 3 trial. Lancet Oncol. 2022, 23, 1044–1054. [Google Scholar] [CrossRef]
- Germano, G.; Frapolli, R.; Simone, M.; Tavecchio, M.; Erba, E.; Pesce, S.; Pasqualini, F.; Grosso, F.; Sanfilippo, R.; Casali, P.G.; et al. Antitumor and anti-inflammatory effects of trabectedin on human myxoid liposarcoma cells. Cancer Res. 2010, 70, 2235–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.W.; Takahashi, N.; Jhanwar, S.; Cordon-Cardo, C.; Elisseyeff, Y.; Jimeno, J.; Faircloth, G.; Bertino, J.R. Sensitivity of soft tissue sarcoma cell lines to chemotherapeutic agents: Identification of ecteinascidin-743 as a potent cytotoxic agent. Clin. Cancer Res. 2001, 7, 2908–2911. [Google Scholar] [PubMed]
- D’Incalci, M.; Colombo, T.; Ubezio, P.; Nicoletti, I.; Giavazzi, R.; Erba, E.; Ferrarese, L.; Meco, D.; Riccardi, R.; Sessa, C.; et al. The combination of yondelis and cisplatin is synergistic against human tumor xenografts. Eur. J. Cancer 2003, 39, 1920–1926. [Google Scholar] [CrossRef]
- Takahashi, N.; Li, W.W.; Banerjee, D.; Scotto, K.W.; Bertino, J.R. Sequence-dependent enhancement of cytotoxicity produced by ecteinascidin 743 (ET-743) with doxorubicin or paclitaxel in soft tissue sarcoma cells. Clin. Cancer Res. 2001, 7, 3251–3257. [Google Scholar]
- Pautier, P.; Floquet, A.; Chevreau, C.; Penel, N.; Guillemet, C.; Delcambre, C.; Cupissol, D.; Selle, F.; Isambert, N.; Piperno-Neumann, S.; et al. A single-arm multicentre phase II trial of doxorubicin in combination with trabectedin in the first-line treatment for leiomyosarcoma with long-term follow-up and impact of cytoreductive surgery. ESMO Open 2021, 6, 100209. [Google Scholar] [CrossRef]
- Simoens, C.; Korst, A.E.; De Pooter, C.M.; Lambrechts, H.A.; Pattyn, G.G.; Faircloth, G.T.; Lardon, F.; Vermorken, J.B. In vitro interaction between ecteinascidin 743 (ET-743) and radiation, in relation to its cell cycle effects. Br. J. Cancer 2003, 89, 2305–2311. [Google Scholar] [CrossRef] [Green Version]
- Jarboe, J.; Zhang, S.; Paz Mejia, A.; Yechieli, R.; Marples, B.; Trent, J. Radiation Sensitization of Sarcoma by Trabectedin. Int. J. Radiat. Oncol. Biol. Phys. 2017, 99, e598. [Google Scholar] [CrossRef] [Green Version]
- Martin-Broto, J.; Hindi, N.; Lopez-Pousa, A.; Peinado-Serrano, J.; Alvarez, R.; Alvarez-Gonzalez, A.; Italiano, A.; Sargos, P.; Cruz-Jurado, J.; Isern-Verdum, J.; et al. Assessment of Safety and Efficacy of Combined Trabectedin and Low-Dose Radiotherapy for Patients With Metastatic Soft-Tissue Sarcomas: A Nonrandomized Phase 1/2 Clinical Trial. JAMA Oncol. 2020, 6, 535–541, Erratum in JAMA Oncol. 2020, 6, 1641. [Google Scholar] [CrossRef]
- Gronchi, A.; Hindi, N.; Cruz, J.; Blay, J.Y.; Lopez-Pousa, A.; Italiano, A.; Alvarez, R.; Gutierrez, A.; Rincón, I.; Sangalli, C.; et al. Trabectedin and RAdiotherapy in Soft Tissue Sarcoma (TRASTS): Results of a Phase I Study in Myxoid Liposarcoma from Spanish (GEIS), Italian (ISG), French (FSG) Sarcoma Groups. EClinicalMedicine 2019, 9, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Rofstad, E.K.; Mathiesen, B.; Henriksen, K.; Kindem, K.; Galappathi, K. The tumor bed effect: Increased metastatic dissemination from hypoxia-induced up-regulation of metastasis-promoting gene products. Cancer Res. 2005, 65, 2387–2396. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, A.D.; Grompe, M. The Fanconi anaemia/BRCA pathway. Nat. Rev. Cancer 2003, 3, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Lavin, M.F.; Kozlov, S.; Gatei, M.; Kijas, A.W. ATM-Dependent Phosphorylation of All Three Members of the MRN Complex: From Sensor to Adaptor. Biomolecules 2015, 5, 2877–2902. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Bhakta, K.S.; Puri, P.L.; Newbury, R.O.; Feramisco, J.R.; Wang, J.Y. Association of ataxia telangiectasia mutated (ATM) gene mutation/deletion with rhabdomyosarcoma. Cancer Biol. Ther. 2003, 2, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Ul-Hassan, A.; Sisley, K.; Hughes, D.; Hammond, D.W.; Robinson, M.H.; Reed, M.W.R. Common genetic changes in leiomyosarcom.a and gastrointestinal stromal tumour: Implication for ataxia telangiectasia mutated involvement. Int. J. Exp. Pathol. 2009, 90, 549–557. [Google Scholar] [CrossRef]
- Bertucci, F.; Pascal Perrot, F.D.; Lerouxd, A.; Collind, F.; le Cesned, A.; Coindred, J.M.; Blayd, J.Y.; Birnbauma, D.; Mamessier, E. PDL1 expression is a poor-prognosis factor in soft-tissue sarcomas. Oncoimmunology 2017, 6, e1278100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocchi, L.; Caraffi, S.; Perris, R.; Mangieri, D. The angiogenic asset of soft tissue sarcomas: A new tool to discover new therapeutic targets. Biosci. Rep. 2014, 34, e00147. [Google Scholar] [CrossRef]
- Tawbi, H.A.; Burgess, M.; Bolejack, V.; Van Tine, B.A.; Schuetze, S.M.; Hu, J.; D’Angelo, S.; Attia, S.; Riedel, R.F.; Priebat, D.A.; et al. Pembrolizumab in advanced soft-tissue sarcoma and bone sarcoma (SARC028): A multicentre, two-cohort, single-arm, open-label, phase 2 trial. Lancet Oncol. 2017, 18, 1493–1501, Erratum in Lancet Oncol. 2017, 18, e711. Erratum in Lancet Oncol. 2018, 19, e8. [Google Scholar] [CrossRef]
- Song, X.; Shao, Y.; Jiang, T.; Ding, Y.; Xu, B.; Zheng, X.; Wang, Q.; Chen, X.; Gu, W.; Wu, C.; et al. Radiotherapy Upregulates Programmed Death Ligand-1 through the Pathways Downstream of Epidermal Growth Factor Receptor in Glioma. EBioMedicine 2018, 28, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.; Sottili, M.; Gerini, C.; Desideri, I.; Bastida, C.; Pallotta, S.; Castiglione, F.; Bonomo, P.; Meattini, I.; Greto, D.; et al. A PPAR-gamma agonist protects from radiation-induced intestinal toxicity. United Eur. Gastroenterol. J. 2017, 5, 218–226. [Google Scholar] [CrossRef]
- Mangoni, M.; Sottili, M.; Salvatore, G.; Meattini, I.; Desideri, I.; Greto, D.; Loi, M.; Becherini, C.; Garlatti, P.; Delli Paoli, C.; et al. Enhancement of Soft Tissue Sarcoma Cell Radiosensitivity by Poly(ADP-ribose) Polymerase-1 Inhibitors. Radiat. Res. 2018, 190, 464–472. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loi, M.; Salvatore, G.; Aquilano, M.; Greto, D.; Talamonti, C.; Salvestrini, V.; Melica, M.E.; Valzano, M.; Francolini, G.; Sottili, M.; et al. Radiosensitizing Effect of Trabectedin on Human Soft Tissue Sarcoma Cells. Int. J. Mol. Sci. 2022, 23, 14305. https://doi.org/10.3390/ijms232214305
Loi M, Salvatore G, Aquilano M, Greto D, Talamonti C, Salvestrini V, Melica ME, Valzano M, Francolini G, Sottili M, et al. Radiosensitizing Effect of Trabectedin on Human Soft Tissue Sarcoma Cells. International Journal of Molecular Sciences. 2022; 23(22):14305. https://doi.org/10.3390/ijms232214305
Chicago/Turabian StyleLoi, Mauro, Giulia Salvatore, Michele Aquilano, Daniela Greto, Cinzia Talamonti, Viola Salvestrini, Maria Elena Melica, Marianna Valzano, Giulio Francolini, Mariangela Sottili, and et al. 2022. "Radiosensitizing Effect of Trabectedin on Human Soft Tissue Sarcoma Cells" International Journal of Molecular Sciences 23, no. 22: 14305. https://doi.org/10.3390/ijms232214305