

Effect of Sucrose Concentration on Rhaponticum carthamoides (Willd.) Iljin Transformed Root Biomass, Caffeoylquinic Acid Derivative, and Flavonoid Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
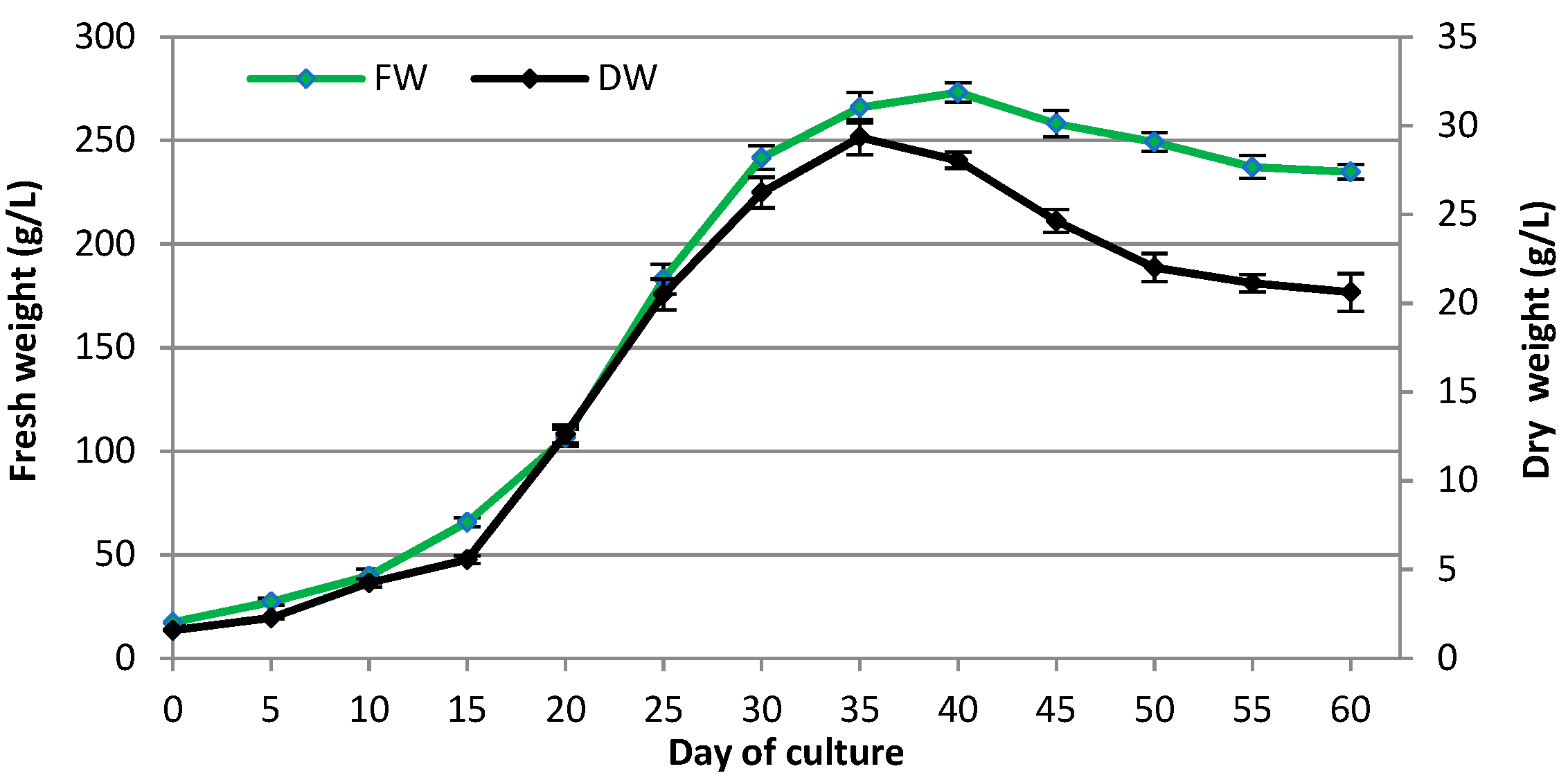
2.1. Influence of Sucrose Concentration on Transformed Root Growth
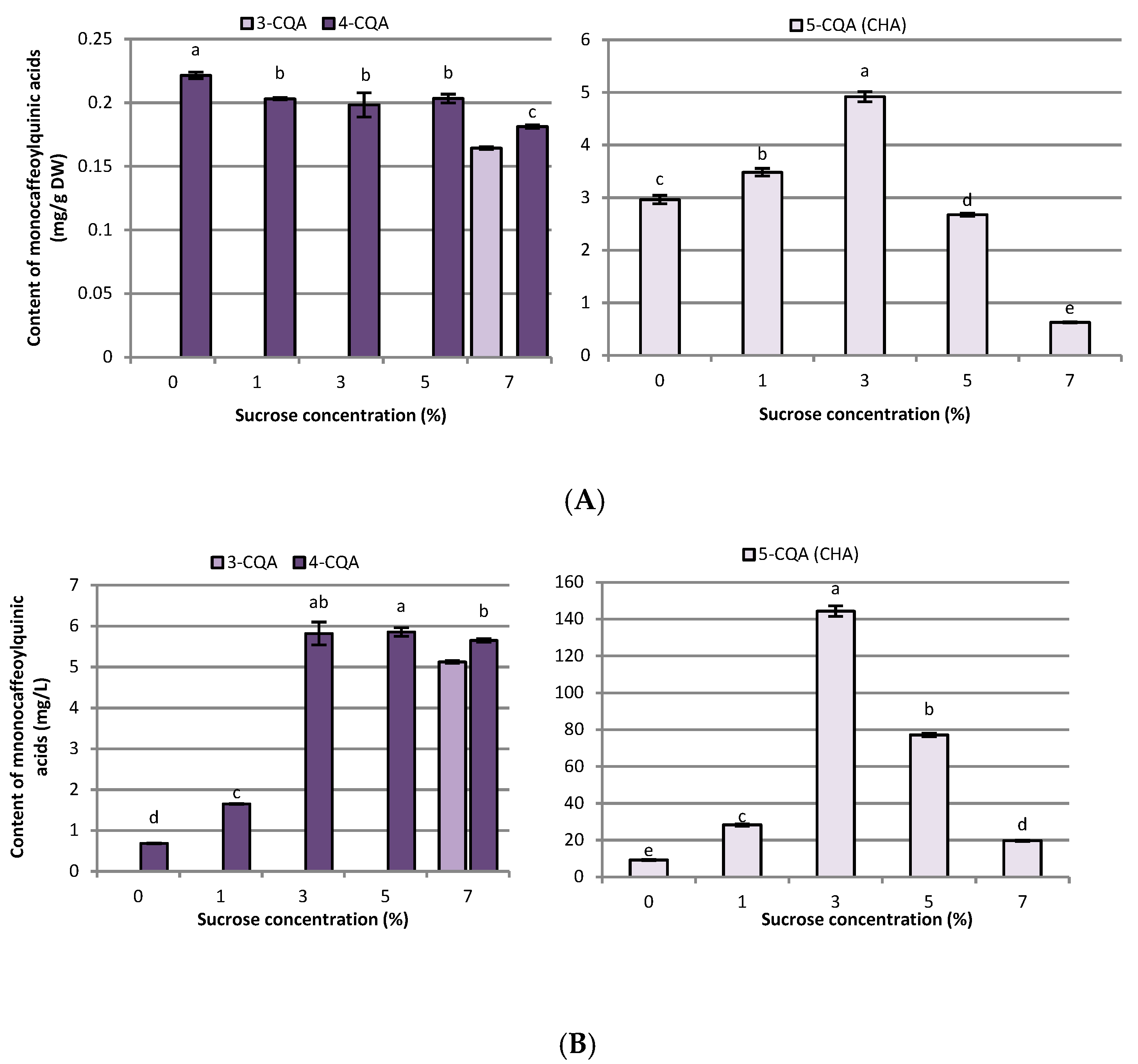
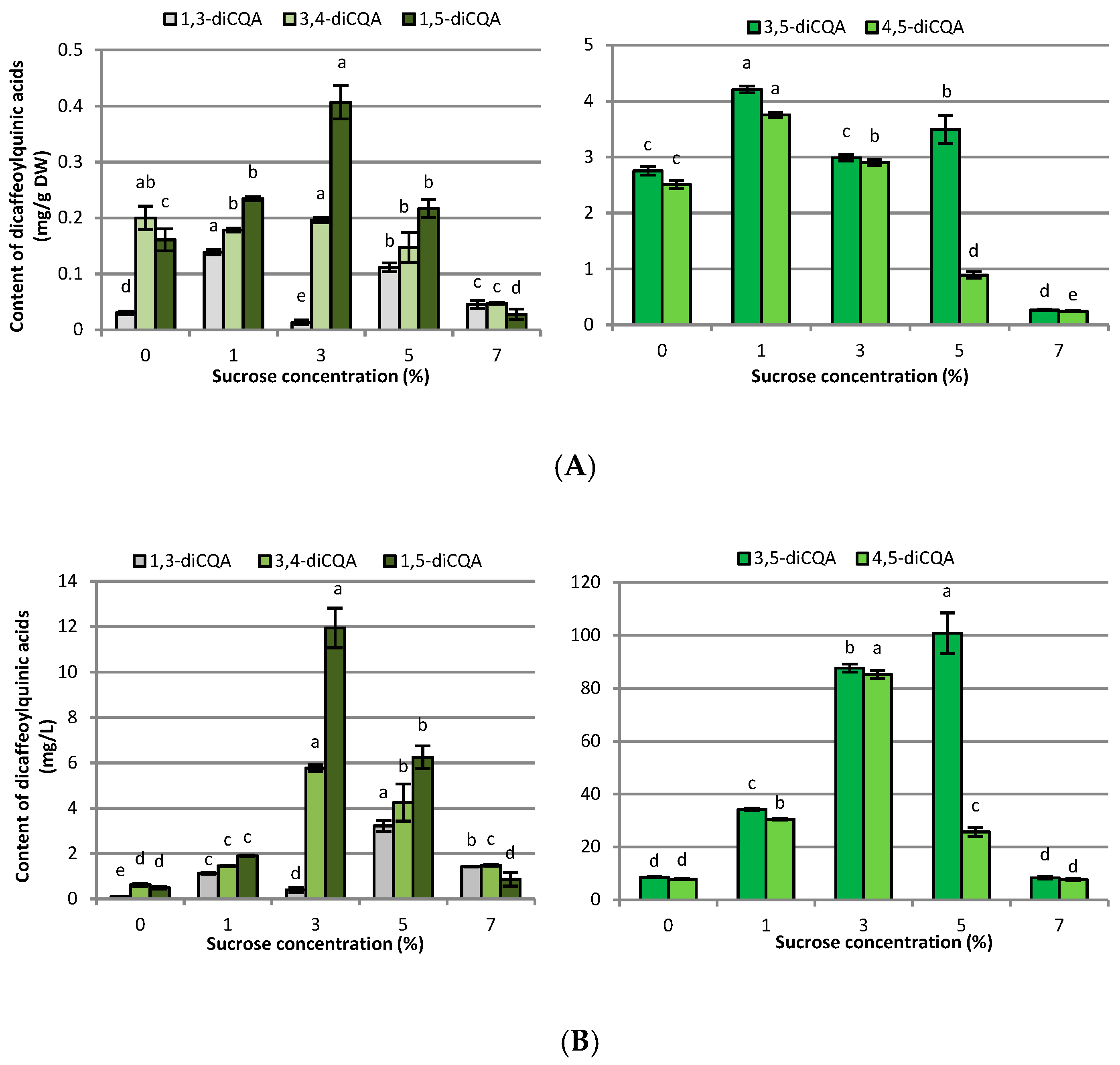
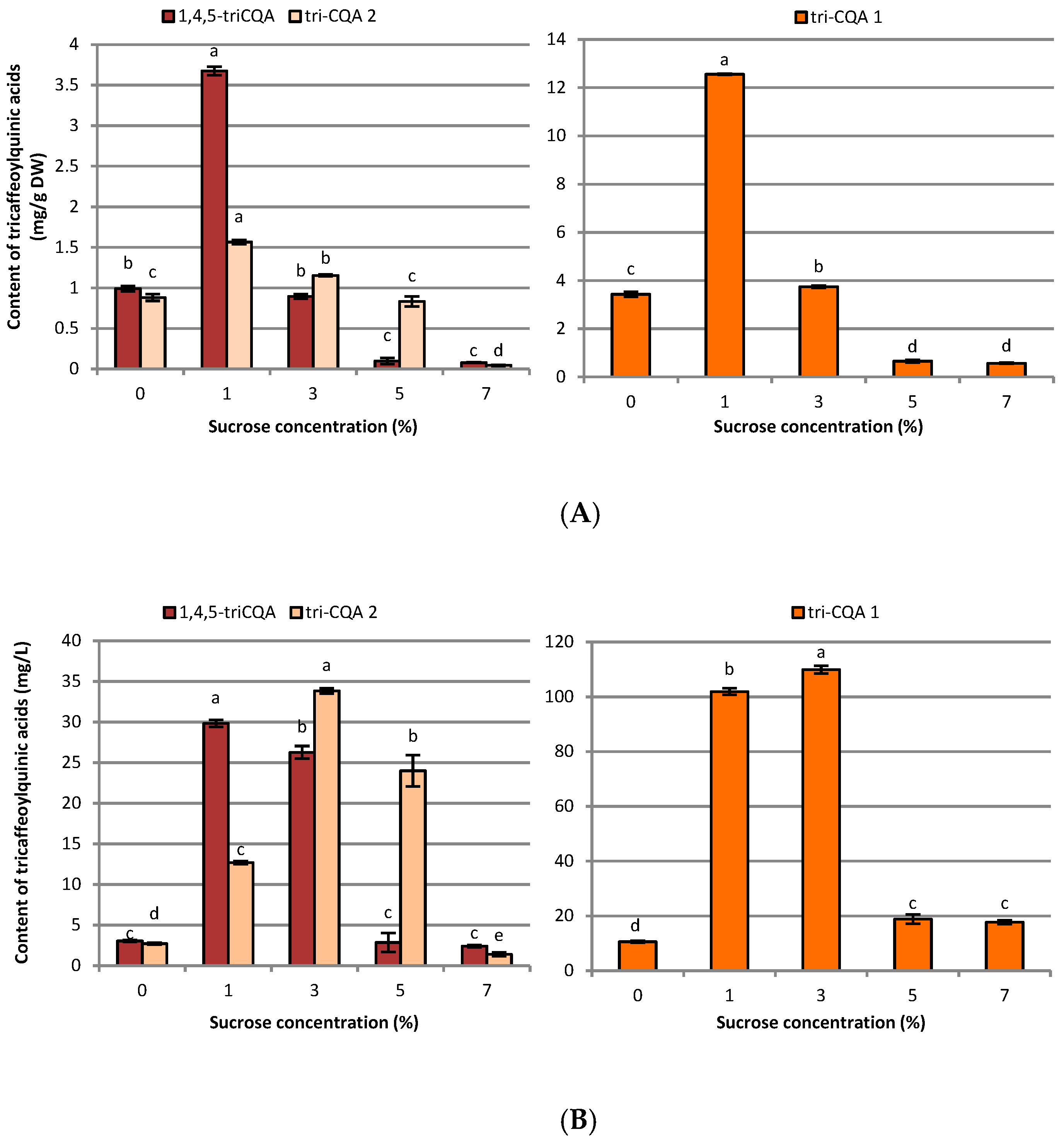
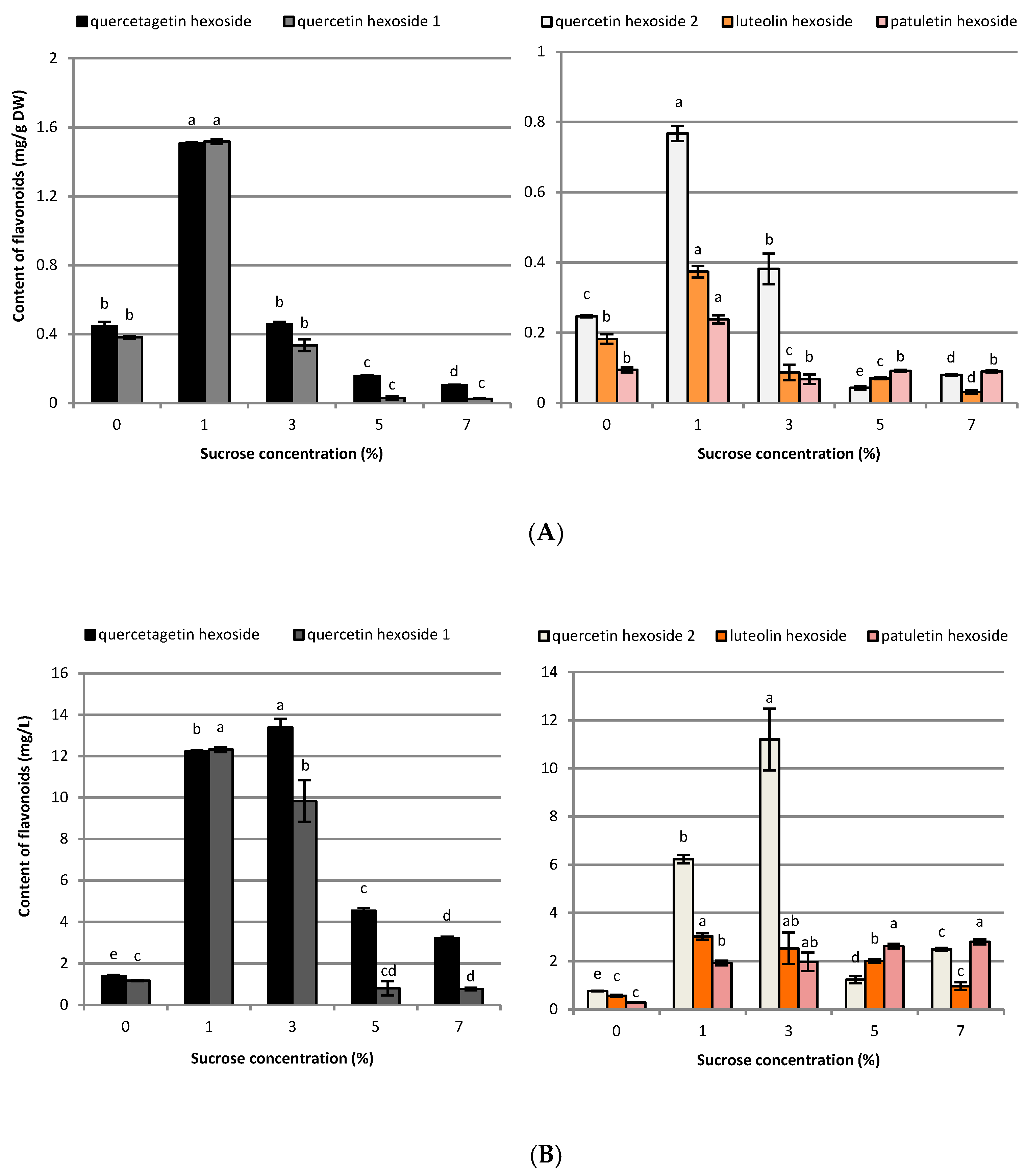
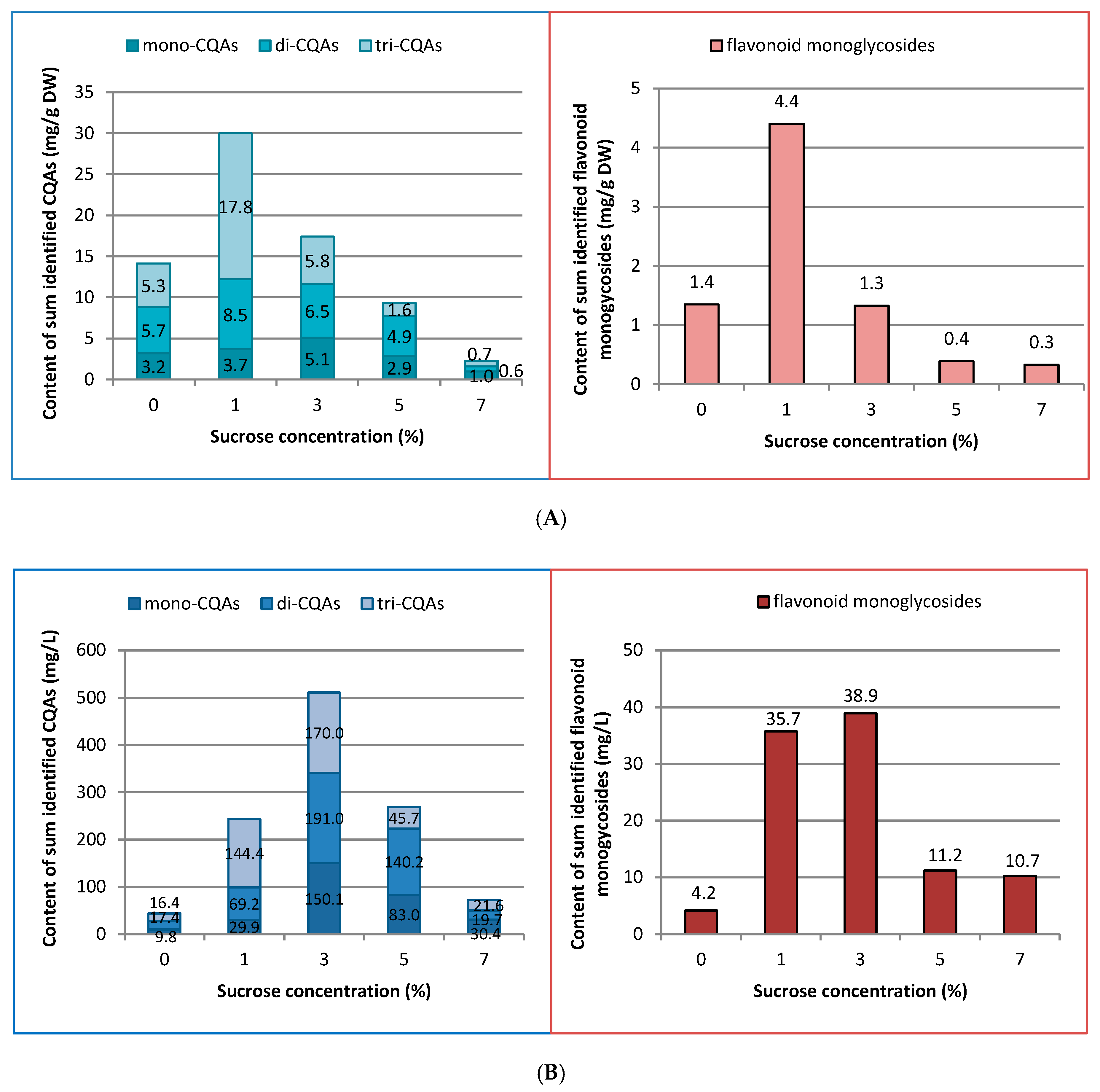
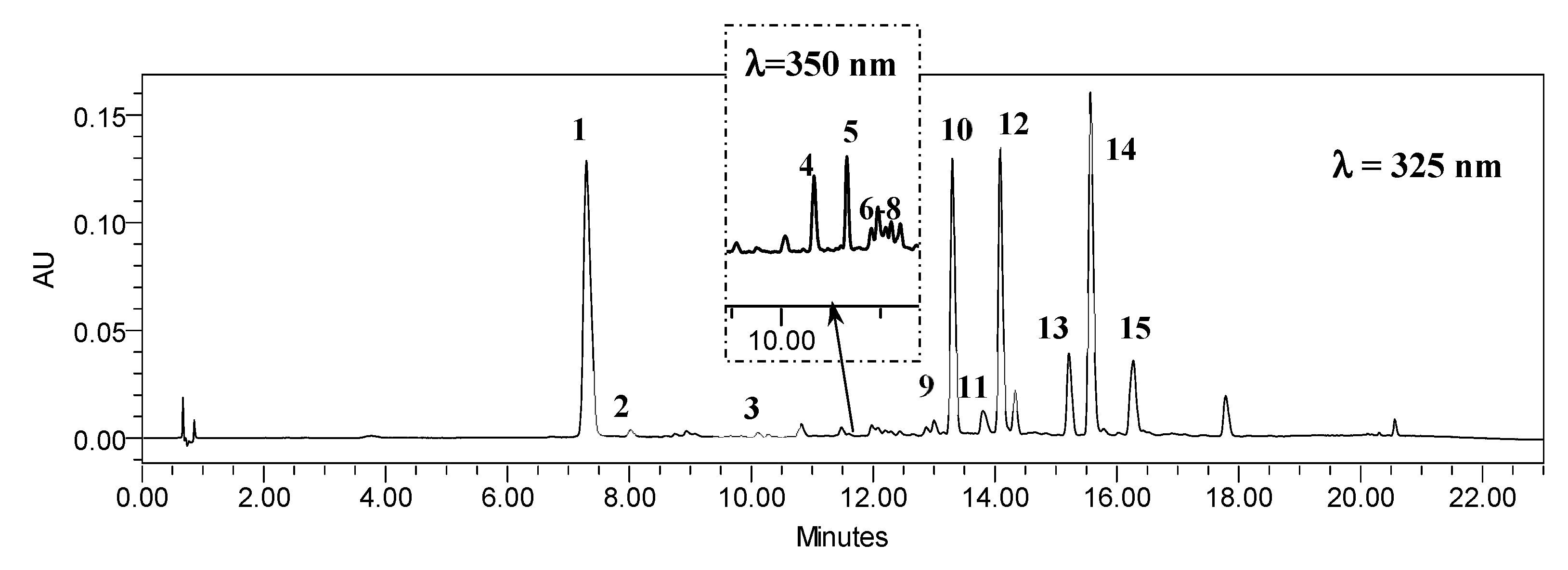
2.2. Influence of Sucrose Concentration on CQA and Flavonoid Production
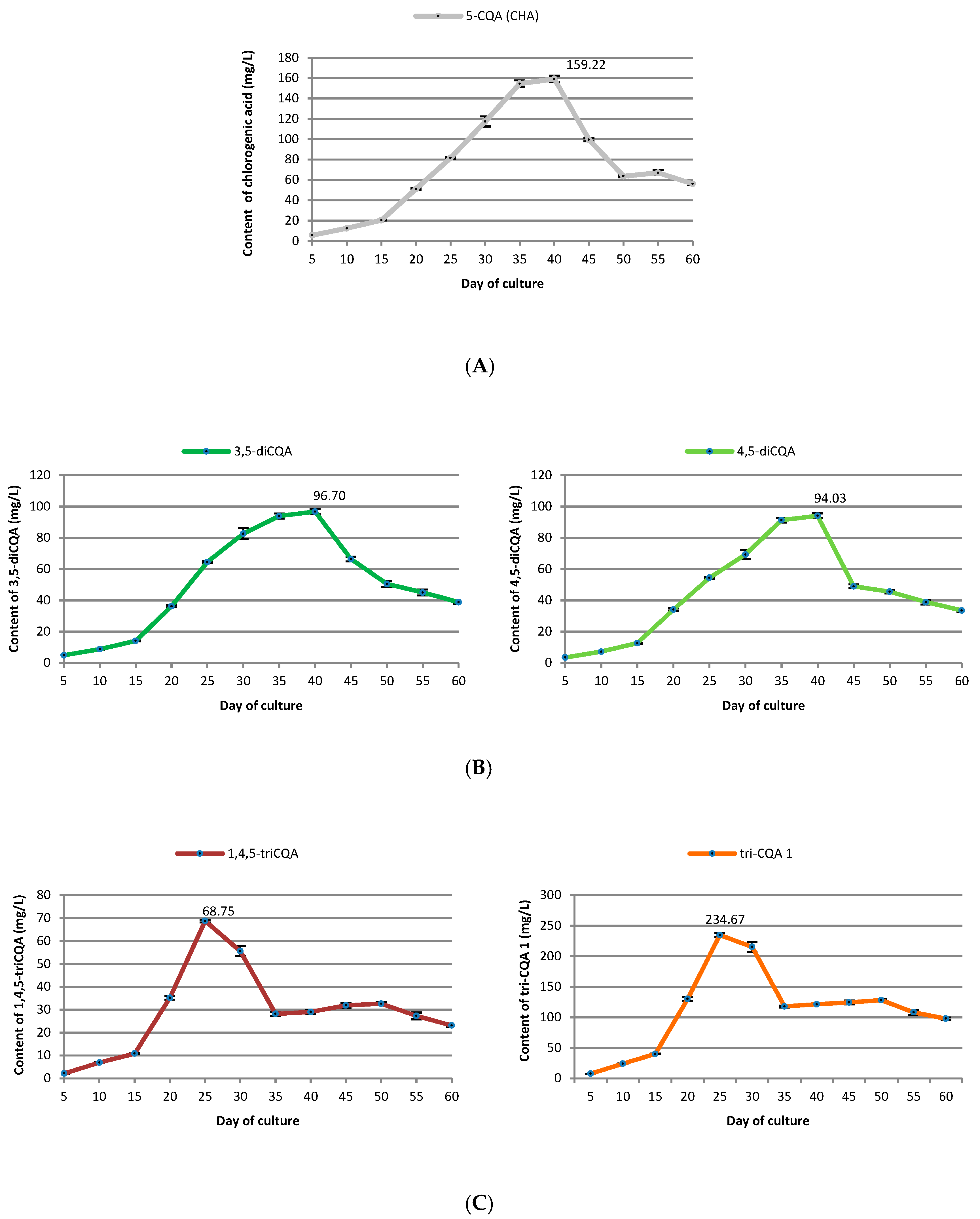
2.3. Growth Curve of Transformed Roots and Accumulation of Various CQAs
3. Discussion
3.1. Influence of Sucrose Concentration on Transformed Root Growth
3.2. Influence of Sucrose Concentration on CQA and Flavonoid Production
3.3. Growth Curve of Transformed Roots and Accumulation of Some CQAs
4. Materials and Methods
4.1. Influence of Sucrose Concentration on the Growth and Time Course of Transformed Roots
4.2. Effect of Sucrose Concentration on Metabolite Production
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greenwell, M.; Rahman, P.K.S.M. Medicinal plants: Their use in anticancer treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [PubMed]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Li, J.; Zhang, X.; Zu, Y.; Yang, Y.; Liu, W.; Xu, Z.; Gao, H.; Sun, X.; Jiang, X.; et al. Current advances in naturally occurring caffeoylquinic acids: Structure, bioactivity and synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef] [PubMed]
- Skała, E.; Makowczyńska, J.; Wieczfinska, J.; Kowalczyk, T.; Sitarek, P. Caffeoylquinic acids with potential biological activity from plant in vitro cultures as alternative sources of valuable natural products. Curr. Pharm. Des. 2020, 26, 2817–2842. [Google Scholar] [CrossRef]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Kokoska, L.; Janovska, D. Chemistry and pharmacology of Rhaponticum carthamoides: A review. Phytochemistry 2009, 70, 842–855. [Google Scholar] [CrossRef]
- Shikov, A.N.; Narkevich, I.A.; Flisyuk, E.V.; Luzhanin, V.G.; Pozharitskaya, O.N. Medicinal plants from the 14th edition of the Russian Pharmacopoeia, recent updates. J. Ethnopharmacol. 2021, 268, 113685. [Google Scholar] [CrossRef]
- Todorova, V.; Ivanov, K.; Ivanova, S. Comparison between the biological active compounds in plants with adaptogenic properties (Rhaponticum carthamoides, Lepidium meyenii, Eleutherococcus senticosus and Panax ginseng). Plants 2022, 11, 64. [Google Scholar] [CrossRef]
- Nekratova, N.A.; Kurovskiy, A.V.; Shurupova, M.N. Impact of elevation and slope exposure on abundance of rare medicinal plant Rhaponticum carthamoides (Maral root). Ukr. J. Ecol. 2020, 10, 210–217. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fub, Y.; Sussmand, M.R.; Wua, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Gantait, S.; Mukherjee, E. Hairy root culture technology: Applications, constraints and prospect. Appl. Microbiol. Biotechnol. 2021, 105, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy root cultures—A versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piątczak, P.; Grąbkowska, R.; Skała, E. pRi-Transformed plants as a source of secondary metabolites. In Hairy Roots: An Effective Tool of Plant Biotechnology; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2018; pp. 45–70. [Google Scholar]
- Bulgakov, V.P.; Vereshchagina, Y.V.; Veremeichik, G.N. Anticancer polyphenols from cultured plant cells: Production and new bioengineering strategies. Curr. Med. Chem. 2018, 25, 4671–4692. [Google Scholar] [CrossRef]
- Skała, E.; Kicel, A.; Olszewska, M.A.; Kiss, A.K.; Wysokińska, H. Establishment of hairy root cultures of Rhaponticum carthamoides (Willd.) Iljin for the production of biomass and caffeic acid derivatives. Biomed. Res. Int. 2015, 2015, 181098. [Google Scholar] [CrossRef] [Green Version]
- Skała, E.; Picot, L.; Bijak, M.; Saluk-Bijak, J.; Szemraj, J.; Kicel, A.; Olszewska, M.A.; Sitarek, P. An efficient plant regeneration from Rhaponticum carthamoides transformed roots, enhanced caffeoylquinic acid derivatives production in pRi-transformed plants and their biological activity. Ind. Crop. Prod. 2019, 129, 327–338. [Google Scholar] [CrossRef]
- Halder, M.; Roychowdhury, D.; Jha, S. A critical review on biotechnological interventions for production and yield enhancement of secondary metabolites in hairy root cultures. In Hairy Roots: An Effective Tool of Plant Biotechnology; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2018; pp. 21–44. [Google Scholar]
- Skała, E.; Sitarek, P. Hairy roots as a source of tropane alkaloids. In Hairy Root Cultures (HRCs) Based Applications. Rhizosphere Biology. Methods and Protocols; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2020; pp. 11–42. [Google Scholar]
- Wawrosch, C.; Schwaiger, S.; Stuppner, H.; Kopp, B. Lignan formation in hairy root cultures of Edelweiss (Leontopodium nivale ssp. alpinum (Cass.) Greuter). Fitoterapia 2014, 97, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Park, J.H.; Kim, Y.K.; Park, N.I.; Lee, S.Y.; Park, S.U. Optimization of growth and pyranocoumarins production in hairy root culture of Angelica gigas Nakai. J. Med. Plant. Res. 2009, 3, 978–981. [Google Scholar]
- Kochan, E.; Szymańska, G.; Szymczyk, P. Effect of sugar concentration on ginsenoside biosynthesis in hairy root cultures of Panax quinquefolium cultivated in shake flasks and nutrient sprinkle bioreactor. Acta Physiol. Plant. 2014, 36, 613–619. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, Y.S.; Li, X.; Kim, H.H.; Arasu, M.V.; Al-Dhabi, N.A.; Lee, S.Y.; Park, S.U. Influence of different carbohydrates on flavonoid accumulation in hairy root cultures of Scutellaria baicalensis. Nat. Prod. Commun. 2016, 11, 799–802. [Google Scholar] [CrossRef] [Green Version]
- Verma, P.C.; Singh, H.; Negi, A.S.; Saxena, G.; Rahman, L.; Banerjee, S. Yield enhancement strategies for the production of picroliv from hairy root culture of Picrorhiza kurroa Royle ex Benth. Plant Signal. Behav. 2015, 10, e1023976. [Google Scholar] [CrossRef] [Green Version]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Naik, P.M.; Al-Khayri, J.M. Impact of abiotic elicitors on in vitro production of plant secondary metabolites: A review. J. Adv. Res. Biotech. 2016, 1, 7. [Google Scholar]
- Praveen, N.; Murthy, H.N. Synthesis of withanolide A depends on carbon source and medium pH in hairy root cultures of Withania somnifera. Ind. Crop. Prod. 2012, 35, 241–243. [Google Scholar] [CrossRef]
- Petrova, M.; Zayova, E.; Dincheva, I.; Badjakov, I.; Vlahova, M. Influence of carbon sources on growth and GC-MS based metabolite profiling of Arnica montana L. hairy roots. Turk. J. Biol. 2015, 39, 469–478. [Google Scholar] [CrossRef]
- Osman, N.I.; Sidik, N.J.; Awal, A. Optimization of sucrose concentration and light treatment in cell suspension culture establishment of Barringtonia racemosa L. Indian J. Sci. Technol. 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Sumaryono, B.; Muslihatin, W.; Ratnadewi, D. Effect of carbohydrate source on growth and performance of in vitro sago palm (Metroxylon sagu Rottb.) plantlets. HAYATI J. Biosci. 2012, 19, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, F.; Xiao, Y.; Wang, W.; Wu, X. Peach PpSnRK1 participates in sucrose-mediated root growth through auxin signaling. Front. Plant Sci. 2020, 11, 409. [Google Scholar] [CrossRef]
- Wang, F.; Ye, Y.; Chen, X.; Wang, J.; Chen, Z.; Zhou, Q. A sucrose non-fermenting-1-related protein kinase 1 gene from potato, StSnRK1, regulates carbohydrate metabolism in transgenic tobacco. Physiol. Mol. Biol. Plants. 2017, 23, 933–943. [Google Scholar] [CrossRef]
- Coleman, H.D.; Beamish, L.; Reid, A.; Park, J.-Y.; Mansfield, S.D. Altered sucrose metabolism impacts plant biomass production and flower development. Transgenic Res. 2010, 19, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Stein, O.; Granot, D. An overview of sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Beigmohamadi, M.; Movafeghi, A.; Jafari, S.; Sharafi, A. Potential of the genetically transformed root cultures of Plumbago europaea for biomass and plumbagin production. Biotechnol. Prog. 2019, 36, e2905. [Google Scholar] [PubMed]
- Makowczyńska, J.; Kalemba, D.; Skała, E. Establishment of Codonopsis pilosula (Franch.) Nannf. transformed roots, influence of the culture conditions on root growth and production of essential oil. Ind. Crop. Prod. 2021, 165, 113446. [Google Scholar] [CrossRef]
- de Paiva Neto, V.B.; Otoni, W.C. Carbon sources and their osmotic potential in plant tissue culture: Does it matter? Sci. Hortic. 2003, 97, 193–202. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Chaki, M.; Begara-Morales, J.C.; Barroso, J.B. Oxidative stress in plants. Antioxidants 2020, 9, 481. [Google Scholar] [CrossRef]
- Lee, E.J.; Mobin, M.; Hahn, E.J.; Paek, K.Y. Effects of sucrose, inoculums density, auxins, and aeration volume on cell growth of Gymnema sylvestre. J. Plant Biol. 2006, 49, 427–431. [Google Scholar] [CrossRef]
- Sari, Y.P.; Kusumawati, E.; Saleh, C.; Kustiawan, W.; Sukartingsih, S. Effect of sucrose and plant growth regulators on callogenesis and preliminary secondary metabolic of different explant Myrmecodia tuberosa. Nus. Biosci. 2018, 10, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, P.M.L.; de Castro, S.; Martins, T.M.; Clemente, A.; Domingos, A. Growth and proteolytic activity of hairy roots from Centaurea calcitrapa: Effect of nitrogen and sucrose. Enzyme Microb. Technol. 2002, 31, 242–249. [Google Scholar] [CrossRef]
- Fu, X.; Yin, Z.-P.; Chen, J.-G.; Shangguan, X.-C.; Wang, X.; Zhang, Q.-F.; Peng, D.-Y. Production of chlorogenic acid and its derivatives in hairy root cultures of Stevia rebaudiana. J. Agric. Food Chem. 2015, 63, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Park, S.Y.; Paek, K.Y. Production of ginsenosides by hairy root cultures of Panax ginseng. In Prduction of Plant Derived Natural Compounds through Hairy Root Culture; Malik, S., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 203–216. [Google Scholar]
- Morkunas, I.; Narożna, D.; Nowak, W.; Samardakiewicz, S.; Remlein-Starosta, D. Cross-talk interactions of sucrose and Fusarium oxysporum in the phenylpropanoid pathway and the accumulation and localization of flavonoids in embryo axes of yellow lupine. J. Plant Physiol. 2011, 168, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Payyavula, R.S.; Shakya, R.; Sengoda, V.G.; Munyaneza, J.E.; Swamy, P.; Navarre, D.A. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flux in HQT-silenced lines. Plant Biotechnol. J. 2015, 13, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, W.; Chen, J.; Jia, R.; Ma, L.; Wang, S.; Wang, J.; Shen, X.; Chu, Z.; Zhu, C.; et al. Development of marker-free transgenic potato tubers enriched in caffeoylquinic acids and flavonols. J. Agric. Food Chem. 2016, 64, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [Green Version]
- Cheevarungnapakul, K.; Khaksar, G.; Panpetch, P.; Boonjing, P.; Sirikantaramas, S. Identification and functional characterization of genes involved in the biosynthesis of caffeoylquinic acids in sunflower (Helianthus annuus L.). Front. Plant Sci. 2019, 10, 968. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.-H.; Murthy, H.N.; Wu, C.-H.; Paek, K.-Y. Sucrose-induced osmotic stress affects biomass, metabolite, and antioxidant levels in root suspension cultures of Hypericum perforatum L. Plant Cell Tiss. Organ Cult. 2010, 103, 7–14. [Google Scholar] [CrossRef]
- Cui, X.-H.; Baque, M.A.; Lee, E.-J.; Paek, K.-Y. Scale-up of adventitious root cultures of Echinacea angustifolia in a pilot-scale bioreactor for the production of biomass and caffeic acid derivatives. Plant Biotechnol. Rep. 2013, 7, 297–308. [Google Scholar] [CrossRef]
- Li, Q.; Tang, M.; Tan, Y.; Ma, D.; Wang, Y.; Zhang, H. Improved production of chlorogenic acid from cell suspension cultures of Lonicera macranthoids. Trop. J. Pharm. Res. 2016, 15, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Grunennvaldt, R.L.; Degenhardt-Goldbach, J.; Brooks, P.; Tomasi, J.C.; Hansel, F.A.; Tran, T.; Gomes, E.N.; Deschamps, C. Callus culture as a new approach for the production of high added value compounds in Ilex paraguariensis: Genotype influence, medium optimization and compounds identification. An. Acad. Bras. Cienc. 2020, 92, e20181251. [Google Scholar] [CrossRef]
- Baque, M.A.; Elgirban, A.; Lee, E.-J.; Paek, K.-Y. Sucrose regulated enhanced induction of anthraquinone, phenolics, flavonoids biosynthesis and activities of antioxidant enzymes in adventitious root suspension cultures of Morinda citrifolia (L.). Acta Physiol. Plant. 2012, 34, 405–415. [Google Scholar] [CrossRef]
- Wu, C.-H.; Dewir, Y.H.; Hahn, E.-J.; Paek, K.-Y. Optimization of culturing conditions for the production of biomass and phenolics from adventitious roots of Echinacea angustifolia. J. Plant Biol. 2006, 49, 193–199. [Google Scholar] [CrossRef]
- Noviyanti, R.; Sari, R.L.K.; Kristanti, A.N.; Yachya, A.; Manuhara, Y.S.W. Biomass and flavonoid production of Gynura procumbens adventitious roots induced by sucrose, phenylalanine and tyrosine. Biosci. Res. 2017, 14, 934–941. [Google Scholar]
- Ali, M.; Abbasi, B.H.; Ahmad, N.; Ali, S.S.; Ali, S.; Ali, G.S. Sucrose-enhanced biosynthesis of medicinally important antioxidant secondary metabolites in cell suspension cultures of Artemisia absinthium L. Bioprocess Biosyst. Eng. 2016, 39, 1945–1954. [Google Scholar] [CrossRef]
- Liu, C.-Z.; Abbasi, B.H.; Gao, M.; Murch, S.J.; Saxena, P.K. Caffeic acid derivatives production by hairy root cultures of Echinacea purpurea. J. Agric. Food Chem. 2006, 54, 8456–8460. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Tian, C.-L.; Murch, S.J.; Saxena, P.K.; Liu, C.-Z. Light-enhanced caffeic acid derivatives biosynthesis in hairy root cultures of Echinacea purpurea. Plant Cell Rep. 2007, 26, 1367–1372. [Google Scholar] [CrossRef]
- Stojakowska, A.; Malarz, J.; Szewczyk, A.; Kisiel, W. Caffeic acid derivatives from a hairy root culture of Lactuca virosa. Acta Physiol. Plant. 2012, 34, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Malarz, J.; Stojakowska, A.; Kisiel, W. Long-term cultured hairy roots of chicory—A rich source of hydroxycinnamates and 8-deoxylactucin glucoside. Appl. Biochem. Biotechnol. 2013, 171, 1589–1601. [Google Scholar] [CrossRef] [Green Version]
- Skała, E.; Grąbkowska, R.; Sitarek, P.; Kuźma, Ł.; Błauż, A.; Wysokińska, H. Rhaponticum carthamoides regeneration through direct and indirect organogenesis, molecular profiles and secondary metabolite production. Plant Cell Tiss. Organ Cult. 2015, 123, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-B.; Chen, J.-G.; Yin, Z.-P.; Shangguan, X.-C.; Peng, D.-Y.; Lu, T.; Lin, P. Methyl jasmonate and salicylic acid elicitation increase content and yield of chlorogenic acid and its derivatives in Gardenia jasminoides cell suspension cultures. Plant Cell Tiss. Organ Cult. 2018, 134, 79–93. [Google Scholar] [CrossRef]
- Bernard, G.; Santos, H.A.D.; Etienne, A.; Samaillie, J.; Neut, C.; Sahpaz, S.; Hilbert, J.-L.; Gagneul, D.; Jullian, N.; Tahrioui, A.; et al. MeJA elicitation of chicory hairy roots promotes efficient increase of 3,5-diCQA accumulation, a potent antioxidant and antibacterial molecule. Antibiotics 2020, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.B.; McCown, B.H. Commercially-feasible micropropagation of mountain lamel Kalmia latifolia by use of shoot tip culture. Proc. Int. Plant Propagator’s Soc. 1980, 30, 421–427. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skała, E.; Olszewska, M.A.; Makowczyńska, J.; Kicel, A. Effect of Sucrose Concentration on Rhaponticum carthamoides (Willd.) Iljin Transformed Root Biomass, Caffeoylquinic Acid Derivative, and Flavonoid Production. Int. J. Mol. Sci. 2022, 23, 13848. https://doi.org/10.3390/ijms232213848
Skała E, Olszewska MA, Makowczyńska J, Kicel A. Effect of Sucrose Concentration on Rhaponticum carthamoides (Willd.) Iljin Transformed Root Biomass, Caffeoylquinic Acid Derivative, and Flavonoid Production. International Journal of Molecular Sciences. 2022; 23(22):13848. https://doi.org/10.3390/ijms232213848
Chicago/Turabian StyleSkała, Ewa, Monika Anna Olszewska, Joanna Makowczyńska, and Agnieszka Kicel. 2022. "Effect of Sucrose Concentration on Rhaponticum carthamoides (Willd.) Iljin Transformed Root Biomass, Caffeoylquinic Acid Derivative, and Flavonoid Production" International Journal of Molecular Sciences 23, no. 22: 13848. https://doi.org/10.3390/ijms232213848