
Sex- and Neuropsychiatric-Dependent Circadian Alterations in Daily Voluntary Physical Activity Engagement and Patterns in Aged 3xTg-AD Mice


Abstract
:1. Introduction
2. Results
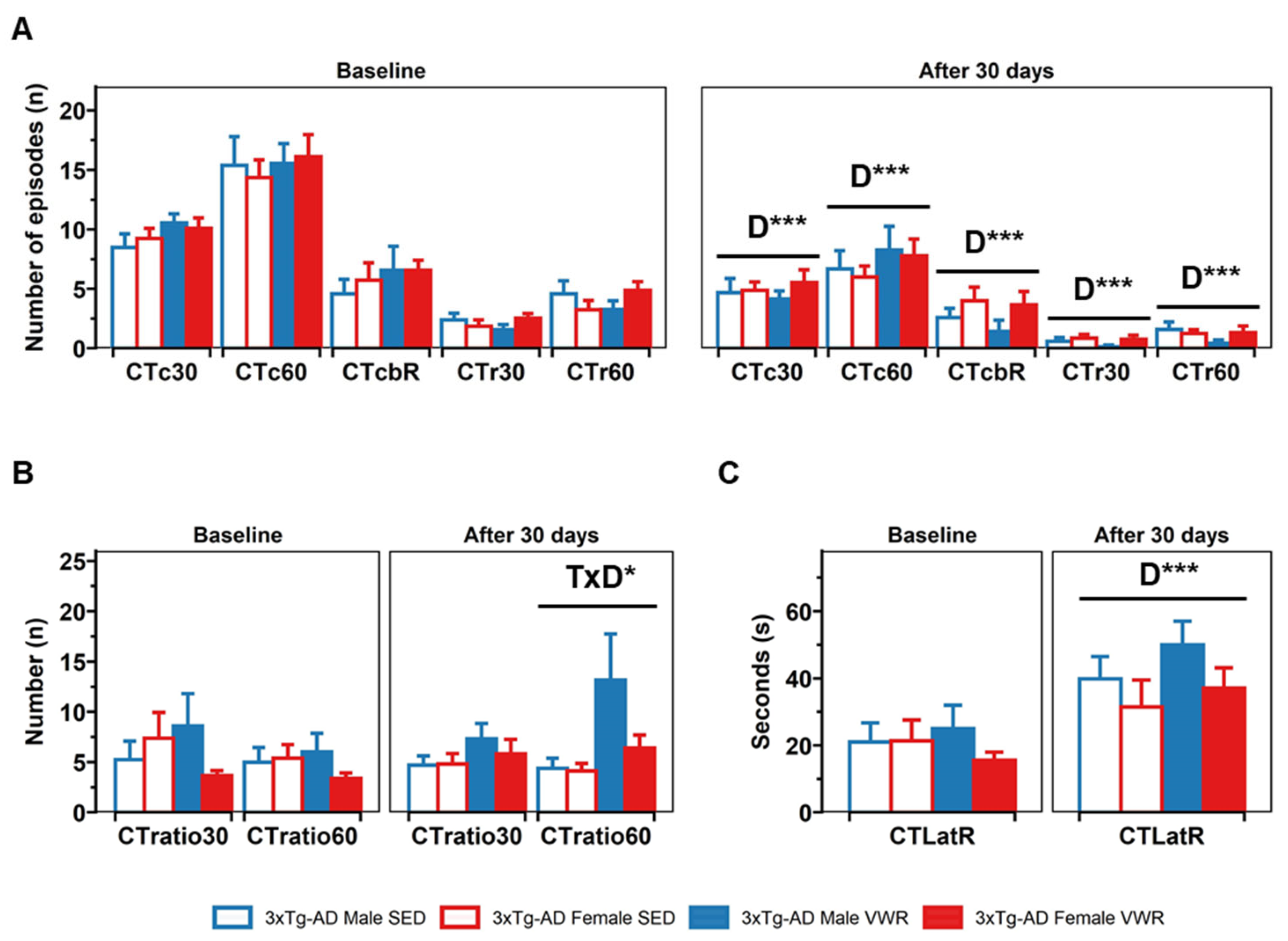
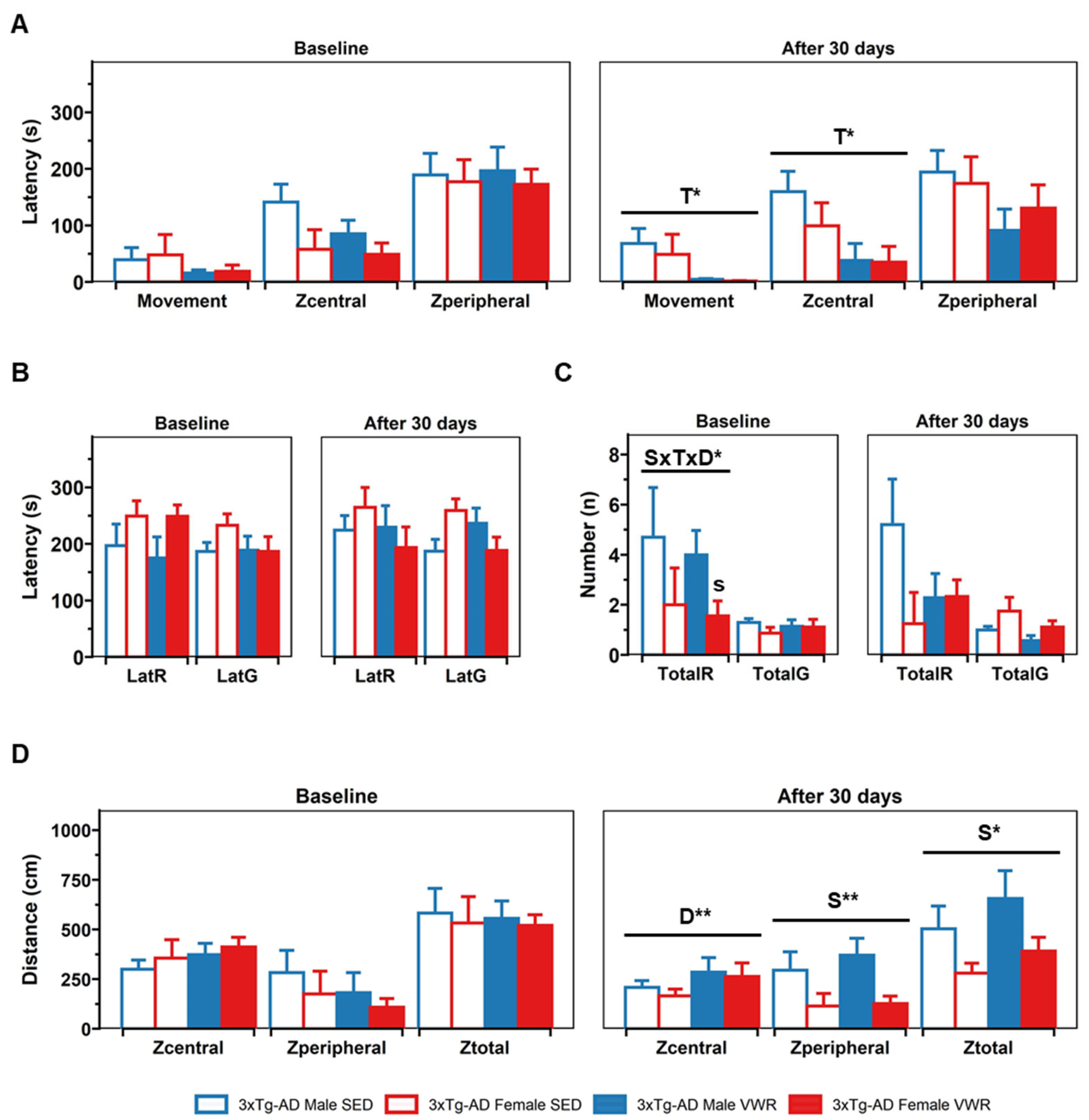
2.1. Corner (CT) and Open Field (OF) Tests
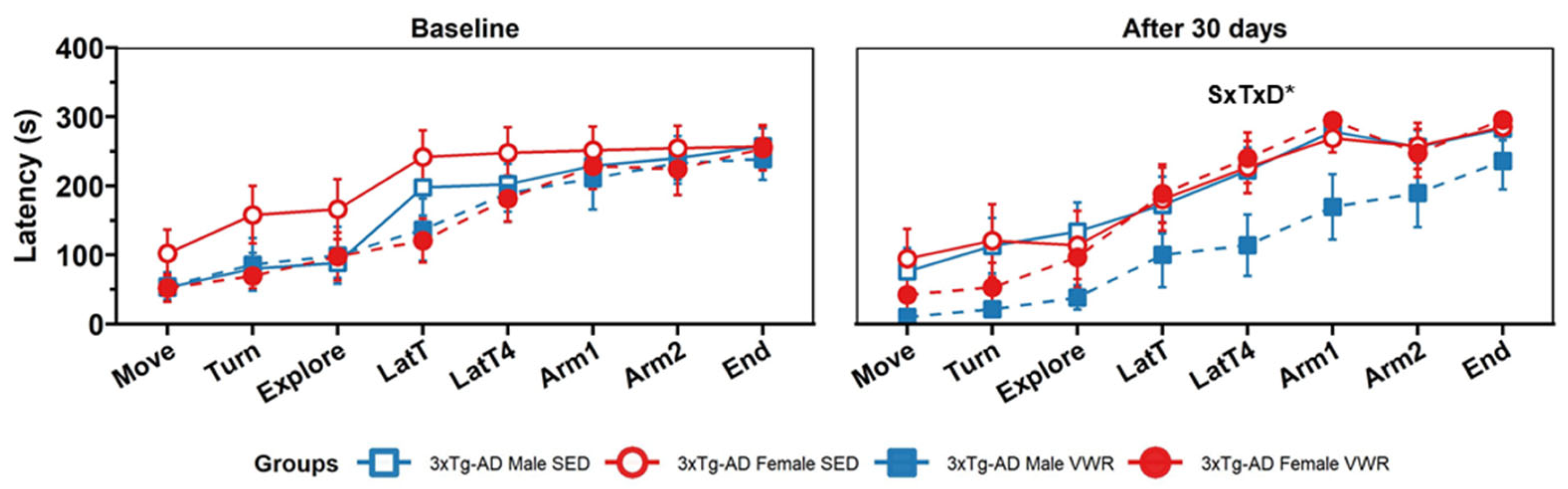
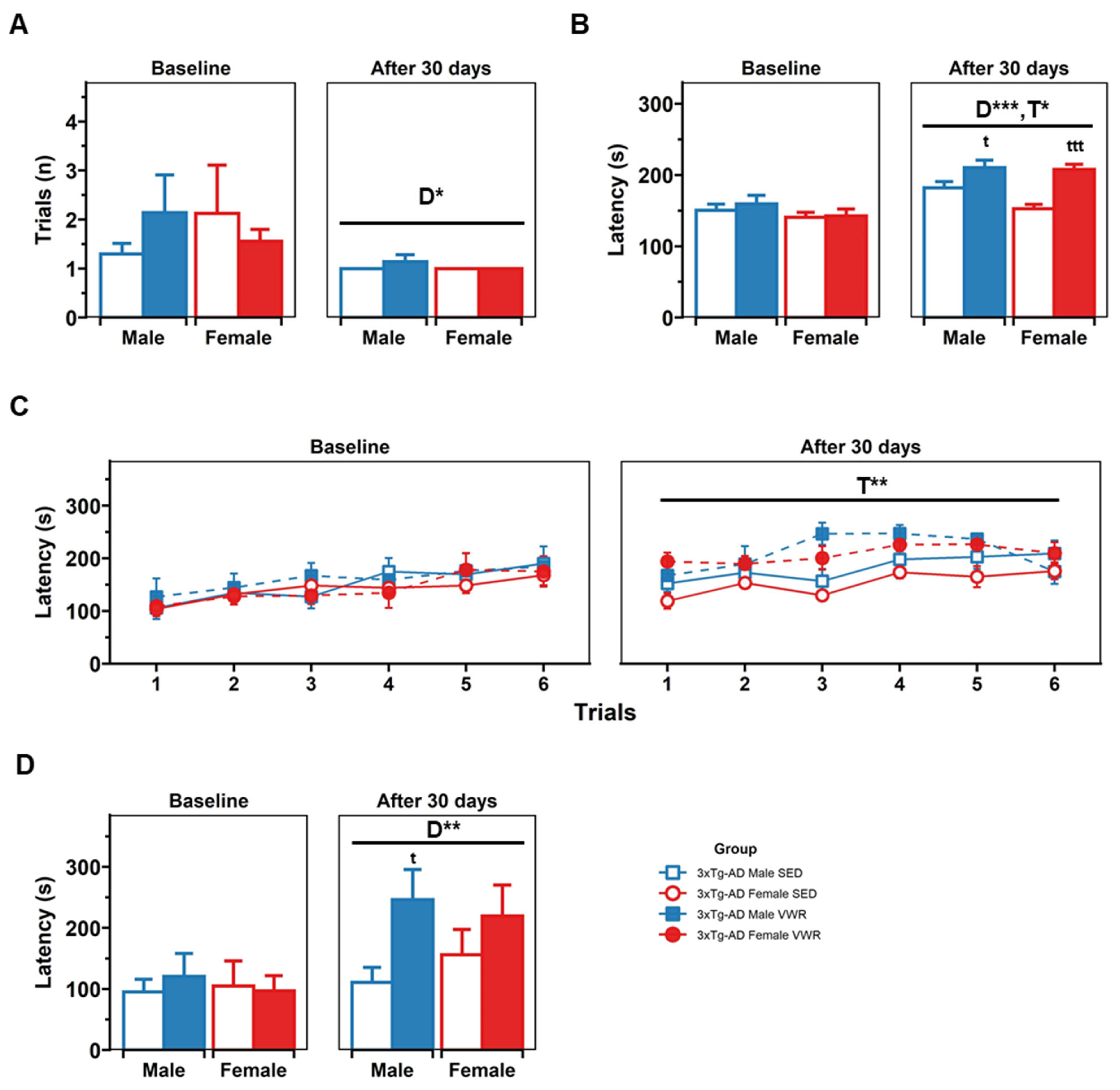
2.2. T-Maze Spontaneous Alternation (TMSA)
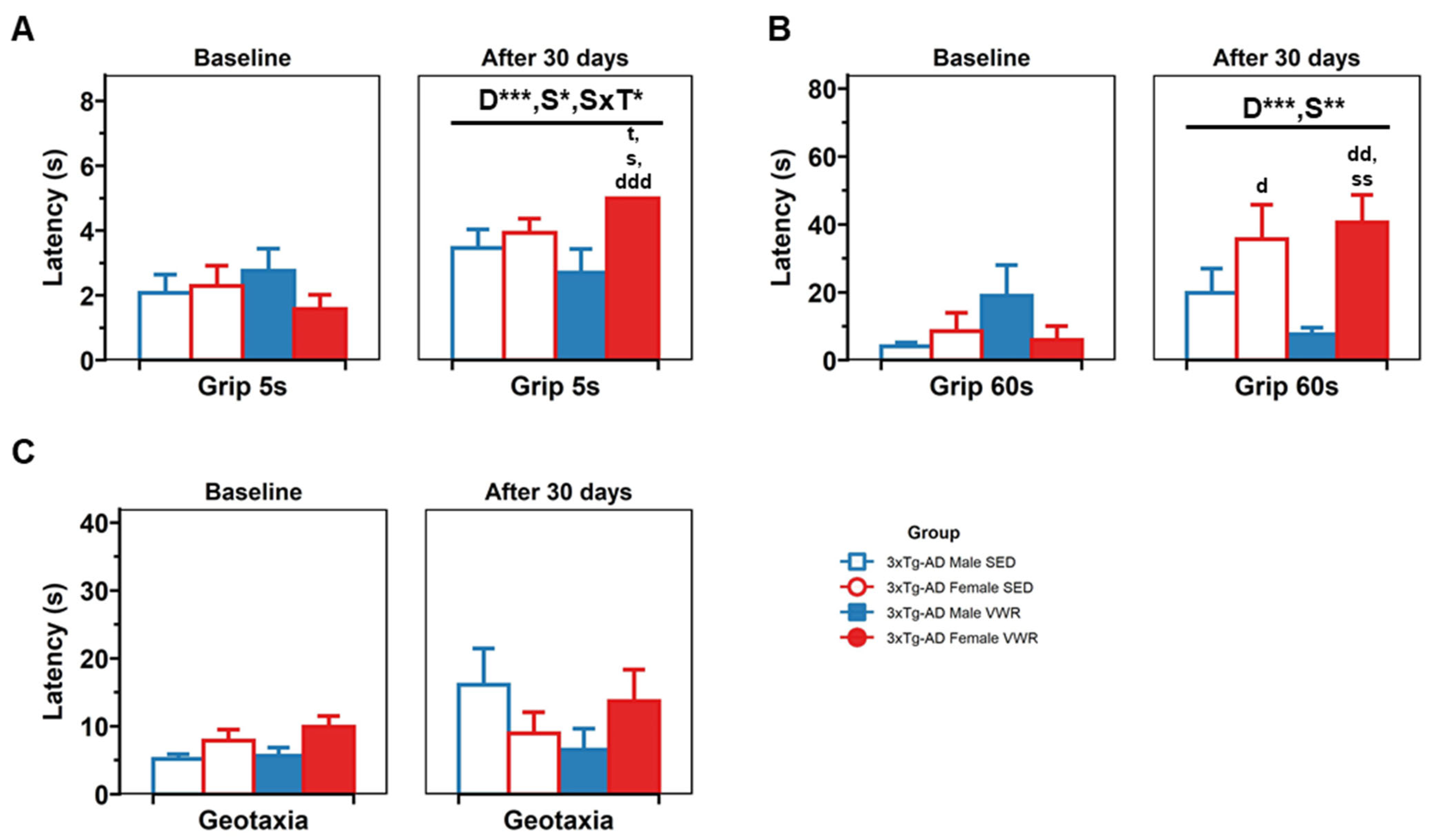
2.3. Sensorimotor Assessment
2.4. Rotarod
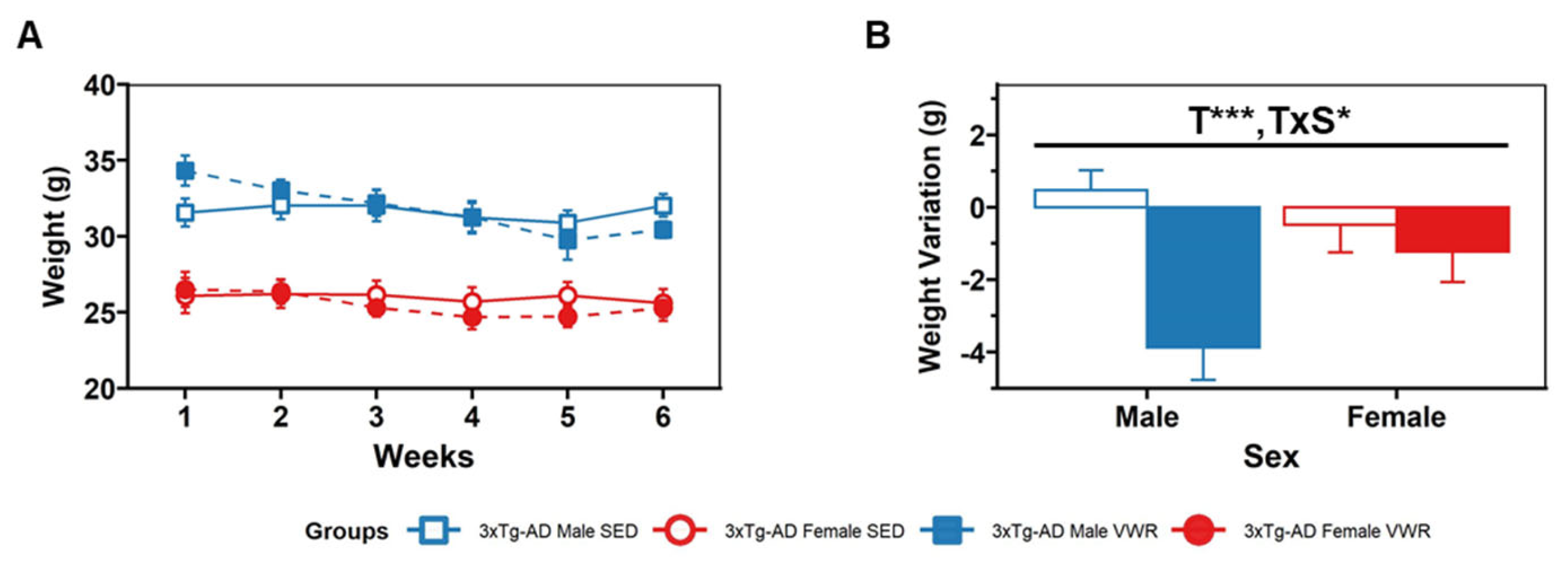
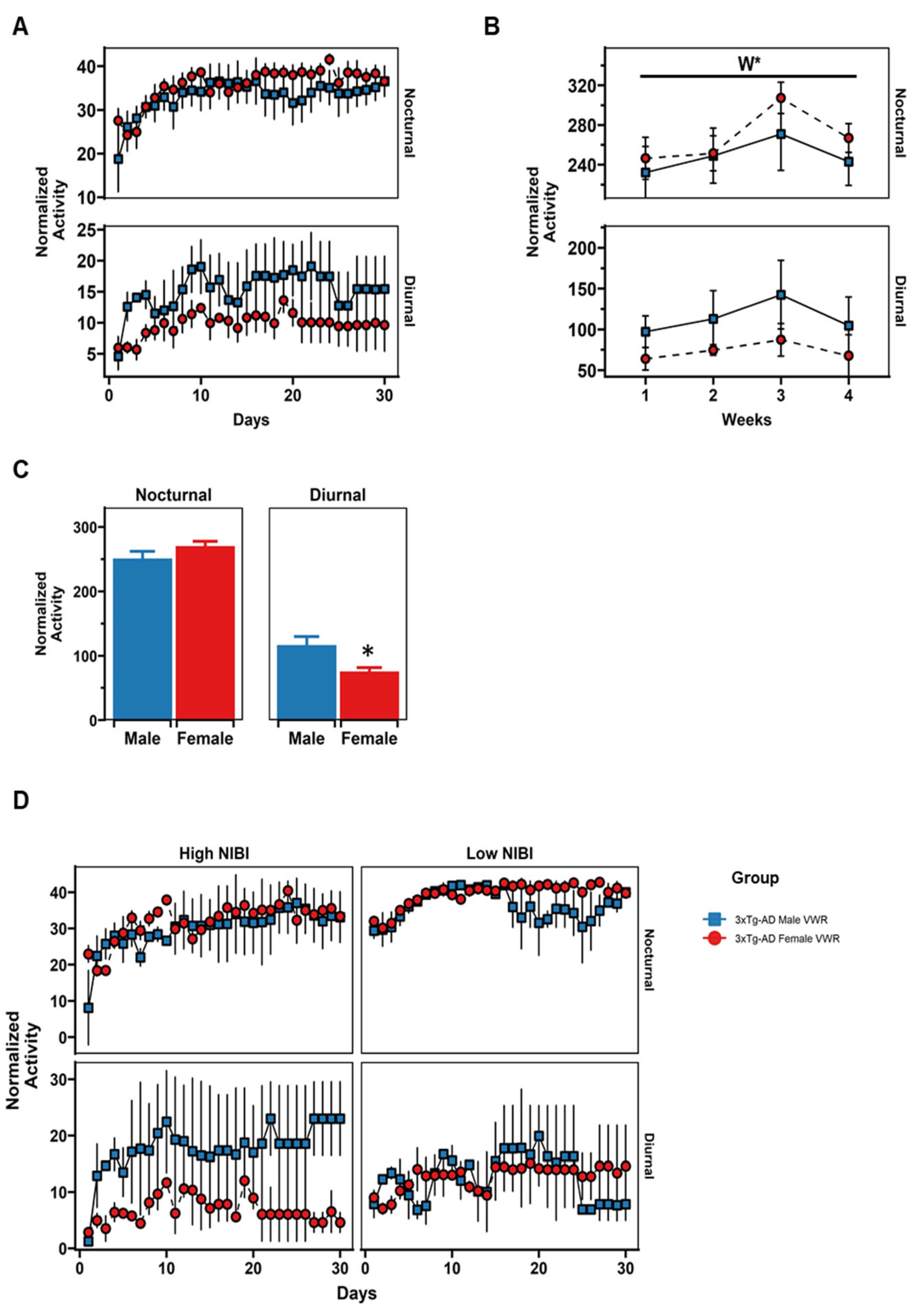
2.5. Daily Patterns of Voluntary Wheel Running (VWR)
2.6. Survival
3. Discussion
4. Methods
4.1. Animals
4.2. Behavioral Assessment
4.2.1. Corner Test (CT)
4.2.2. Open Field (OF) Test
4.2.3. T-Maze Spontaneous Alternation (TMSA)
4.2.4. Sensorimotor Assessment
4.2.5. Rotarod
4.3. Intervention Protocol
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The Diagnosis of Dementia Due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimers Dement. 2011, 7, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, J.L.; McPherson, S. Neuropsychiatric Assessment of Alzheimer’s Disease and Related Dementias. Aging 2001, 13, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, K.; Lovestone, S. The Dementias. Lancet 2002, 360, 1759–1766. [Google Scholar] [CrossRef]
- Bianchetti, A.; Scuratti, A.; Zanetti, O.; Binetti, G.; Frisoni, G.B.; Magni, E.; Trabucchi, M. Predictors of Mortality and Institutionalization in Alzheimer Disease Patients 1 Year after Discharge from an Alzheimer Dementia Unit. Dementia 1995, 6, 108–112. [Google Scholar] [CrossRef]
- Pollak, C.P.; Perlick, D. Sleep Problems and Institutionalization of the Elderly. J. Geriatr. Psychiatry Neurol. 1991, 4, 204–210. [Google Scholar] [CrossRef]
- Reisberg, B.; Borenstein, J.; Salob, S.P.; Ferris, S.H.; Franssen, E.; Georgotas, A. Behavioral Symptoms in Alzheimer’s Disease: Phenomenology and Treatment. J. Clin. Psychiatry 1987, 48 Suppl, 9–15. [Google Scholar]
- Cerejeira, J.; Lagarto, L.; Mukaetova-Ladinska, E.B. Behavioral and Psychological Symptoms of Dementia. Front. Neurol. 2012, MAY, 73. [Google Scholar] [CrossRef] [Green Version]
- Videnovic, A.; Lazar, A.S.; Barker, R.A.; Overeem, S. ‘The Clocks That Time Us’—Circadian Rhythms in Neurodegenerative Disorders. Nat. Rev. Neurol. 2014, 10, 683. [Google Scholar] [CrossRef] [Green Version]
- Esquiva, G.; Hannibal, J. Melanopsin-Expressing Retinal Ganglion Cells in Aging and Disease. Histol. Histopathol. 2019, 34, 1299–1311. [Google Scholar] [CrossRef]
- Stopa, E.G.; Volicer, L.; Kuo-Leblanc, V.; Harper, D.; Lathi, D.; Tate, B.; Satlin, A. Pathologic Evaluation of the Human Suprachiasmatic Nucleus in Severe Dementia. J. Neuropathol. Exp. Neurol. 1999, 58, 29–39. [Google Scholar] [CrossRef]
- Tranah, G.J.; Blackwell, T.; Stone, K.L.; Ancoli-Israel, S.; Paudel, M.L.; Ensrud, K.E.; Cauley, J.A.; Redline, S.; Hillier, T.A.; Cummings, S.R.; et al. Circadian Activity Rhythms and Risk of Incident Dementia and Mild Cognitive Impairment in Older Women. Ann. Neurol. 2011, 70, 722–732. [Google Scholar] [CrossRef]
- Tolppanen, A.-M.; Solomon, A.; Kulmala, J.; Kåreholt, I.; Ngandu, T.; Rusanen, M.; Laatikainen, T.; Soininen, H.; Kivipelto, M. Leisure-Time Physical Activity from Mid- to Late Life, Body Mass Index, and Risk of Dementia. Alzheimer’s Dement. 2015, 11, 434–443.e6. [Google Scholar] [CrossRef]
- Stephen, R.; Hongisto, K.; Solomon, A.; Lönnroos, E. Physical Activity and Alzheimer’s Disease: A Systematic Review. J. Gerontol. A. Biol. Sci. Med. Sci. 2017, 72, 733–739. [Google Scholar] [CrossRef] [Green Version]
- Honea, R.A.; Thomas, G.P.; Harsha, A.; Anderson, H.S.; Donnelly, J.E.; Brooks, W.M.; Burns, J.M. Cardiorespiratory Fitness and Preserved Medial Temporal Lobe Volume in Alzheimer Disease. Alzheimer Dis. Assoc. Disord. 2009, 23, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Watts, A.S.; Mortby, M.E.; Burns, J.M. Depressive Symptoms as a Barrier to Engagement in Physical Activity in Older Adults with and without Alzheimer’s Disease. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Cote, A.C.; Phelps, R.J.; Kabiri, N.S.; Bhangu, J.S.; Thomas, K. Evaluation of Wearable Technology in Dementia: A Systematic Review and Meta-Analysis. Front. Med. 2021, 7, 501104. [Google Scholar] [CrossRef]
- Lu, Z.; Harris, T.B.; Shiroma, E.J.; Leung, J.; Kwok, T. Patterns of Physical Activity and Sedentary Behavior for Older Adults with Alzheimer’s Disease, Mild Cognitive Impairment, and Cognitively Normal in Hong Kong. J. Alzheimer’s Dis. 2018, 66, 1453–1462. [Google Scholar] [CrossRef]
- Moyle, W.; Jones, C.; Murfield, J.; Draper, B.; Beattie, E.; Shum, D.; Thalib, L.; O’Dwyer, S.; Mervin, C.M. Levels of Physical Activity and Sleep Patterns among Older People with Dementia Living in Long-Term Care Facilities: A 24-h Snapshot. Maturitas 2017, 102, 62–68. [Google Scholar] [CrossRef] [Green Version]
- García-Mesa, Y.; López-Ramos, J.C.; Giménez-Llort, L.; Revilla, S.; Guerra, R.; Gruart, A.; Laferla, F.M.; Cristòfol, R.; Delgado-García, J.M.; Sanfeliu, C. Physical Exercise Protects against Alzheimer’s Disease in 3xTg-AD Mice. J. Alzheimer’s Dis. 2011, 24, 421–454. [Google Scholar] [CrossRef] [Green Version]
- Kosel, F.; Pelley, J.M.S.; Franklin, T.B. Behavioural and Psychological Symptoms of Dementia in Mouse Models of Alzheimer’s Disease-Related Pathology. Neurosci. Biobehav. Rev. 2020, 112, 634–647. [Google Scholar] [CrossRef]
- Ingram, D.K. Age-Related Decline in Physical Activity: Generalization to Nonhumans. Med. Sci. Sports Exerc. 2000, 32, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- De Bono, J.P.; Adlam, D.; Paterson, D.J.; Channon, K.M. Novel Quantitative Phenotypes of Exercise Training in Mouse Models. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2006, 290, 926–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightfoot, J.T.; Turner, M.J.; Daves, M.; Vordermark, A.; Kleeberger, S.R. Genetic Influence on Daily Wheel Running Activity Level. Physiol. Genomics 2005, 19, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, J.T. Sex Hormones’ Regulation of Rodent Physical Activity: A Review. Int. J. Biol. Sci. 2008, 4, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-Transgenic Model of Alzheimer’s Disease with Plaques and Tangles: Intracellular Aβ and Synaptic Dysfunction. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Bilkei-Gorzo, A. Genetic Mouse Models of Brain Ageing and Alzheimer’s Disease. Pharmacol. Ther. 2014, 142, 244–257. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Kitazawa, M.; Tseng, B.P.; Laferla, F.M. Amyloid Deposition Precedes Tangle Formation in a Triple Transgenic Model of Alzheimer’ s Disease. Neurobiol. Aging 2003, 24, 1063–1070. [Google Scholar] [CrossRef]
- Belfiore, R.; Rodin, A.; Ferreira, E.; Velazquez, R.; Branca, C.; Caccamo, A.; Oddo, S. Temporal and Regional Progression of Alzheimer’s Disease-like Pathology in 3xTg-AD Mice. Aging Cell 2019, 18. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Johansson, B.; Oddo, S.; Tobeña, A.; LaFerla, F.M.; Fernández-Teruel, A. Modeling Behavioral and Neuronal Symptoms of Alzheimer’s Disease in Mice: A Role for Intraneuronal Amyloid. Neurosci. Biobehav. Rev. 2007, 31, 125–147. [Google Scholar] [CrossRef]
- Blázquez, G.; Cañete, T.; Tobeña, A.; Giménez-Llort, L.; Fernández-Teruel, A. Cognitive and Emotional Profiles of Aged Alzheimer’s Disease (3×TgAD) Mice: Effects of Environmental Enrichment and Sexual Dimorphism. Behav. Brain Res. 2014, 268, 185–201. [Google Scholar] [CrossRef]
- Cañete, T.; Blázquez, G.; Tobeña, A.; Giménez-Llort, L.; Fernández-Teruel, A. Cognitive and Emotional Alterations in Young Alzheimer’s Disease (3xTgAD) Mice: Effects of Neonatal Handling Stimulation and Sexual Dimorphism. Behav. Brain Res. 2015, 281, 156–171. [Google Scholar] [CrossRef]
- Stover, K.R.; Campbell, M.A.; Van Winssen, C.M.; Brown, R.E. Analysis of Motor Function in 6-Month-Old Male and Female 3xTg-AD Mice. Behav. Brain Res. 2015, 281, 16–23. [Google Scholar] [CrossRef]
- Wu, M.; Zhou, F.; Cao, X.; Yang, J.; Bai, Y.; Yan, X.; Cao, J.; Qi, J. Abnormal Circadian Locomotor Rhythms and Per Gene Expression in Six-Month-Old Triple Transgenic Mice Model of Alzheimer’s Disease. Neurosci. Lett. 2018, 676, 13–18. [Google Scholar] [CrossRef]
- Schrack, J.A.; Zipunnikov, V.; Goldsmith, J.; Bai, J.; Simonsick, E.M.; Crainiceanu, C.; Ferrucci, L. Assessing the Physical Cliff: Detailed Quantification of Age-Related Differences in Daily Patterns of Physical Activity. J. Gerontol.-Ser. A Biol. Sci. Med. Sci. 2014, 69, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.P.; Curtis, T.S.; Turner, M.J.; Lightfoot, J.T. Influence of Age of Exposure to a Running Wheel on Activity in Inbred Mice. Med. Sci. Sports Exerc. 2006, 38, 51–56. [Google Scholar] [CrossRef]
- Nassan, M.; Videnovic, A. Circadian Rhythms in Neurodegenerative Disorders. Nat. Rev. Neurol. 2022, 18, 7–24. [Google Scholar] [CrossRef]
- Bedrosian, T.A.; Herring, K.L.; Weil, Z.M.; Nelson, R.J. Altered Temporal Patterns of Anxiety in Aged and Amyloid Precursor Protein (APP) Transgenic Mice. Proc. Natl. Acad. Sci. USA 2011, 108, 11686–11691. [Google Scholar] [CrossRef] [Green Version]
- Volicer, L.; Harper, D.G.; Manning, B.C.; Goldstein, R.; Satlin, A. Sundowning and Circadian Rhythms in Alzheimer’s Disease. Am. J. Psychiatry 2001, 158, 704–711. [Google Scholar] [CrossRef]
- Mahlberg, R.; Kunz, D.; Sutej, I.; Kühl, K.-P.; Hellweg, R. Melatonin Treatment of Day-Night Rhythm Disturbances and Sundowning in Alzheimer Disease: An Open-Label Pilot Study Using Actigraphy. J. Clin. Psychopharmacol. 2004, 24, 456–459. [Google Scholar] [CrossRef]
- Cornish, E.R.; Mrosovsky, N. Activity during Food Deprivation and Satiation of Six Species of Rodent. Anim. Behav. 1965, 13, 242–248. [Google Scholar] [CrossRef]
- Koteja, P.; Swallow, J.G.; Carter, P.A.; Garland, T. Energy Cost of Wheel Running in House Mice: Implications for Coadaptation of Locomotion and Energy Budgets. Physiol. Biochem. Zool. 1999, 72, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Pietropaolo, S.; Sun, Y.; Li, R.; Brana, C.; Feldon, J.; Yee, B.K. The Impact of Voluntary Exercise on Mental Health in Rodents: A Neuroplasticity Perspective. Behav. Brain Res. 2008, 192, 42–60. [Google Scholar] [CrossRef] [PubMed]
- James, B.D.; Boyle, P.A.; Bennett, D.A.; Buchman, A.S. Total Daily Activity Measured with Actigraphy and Motor Function in Community-Dwelling Older Persons with and without Dementia. Alzheimer Dis. Assoc. Disord. 2012, 26, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Torres-Lista, V.; De la Fuente, M.; Giménez-Llort, L. Survival Curves and Behavioral Profiles of Female 3xTg-AD Mice Surviving to 18-Months of Age as Compared to Mice with Normal Aging. J. Alzheimer’s Dis. Rep. 2017, 1, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mesa, Y.; Giménez-Llort, L.; López, L.C.; Venegas, C.; Cristòfol, R.; Escames, G.; Acuña-Castroviejo, D.; Sanfeliu, C. Melatonin plus Physical Exercise Are Highly Neuroprotective in the 3xTg-AD Mouse. Neurobiol. Aging 2012, 33, 1124.e13–1124.e29. [Google Scholar] [CrossRef] [Green Version]
- Torres-Lista, V.; Giménez-Llort, L. Early Postnatal Handling and Environmental Enrichment Improve the Behavioral Responses of 17-Month-Old 3xTg-AD and Non-Transgenic Mice in the Forced Swim Test in a Gender-Dependent Manner. Behav. Processes 2015, 120, 120–127. [Google Scholar] [CrossRef]
- Roda, A.R.; Villegas, S.; Esquerda-Canals, G.; Martí-Clúa, J. Cognitive Impairment in the 3xTg-AD Mouse Model of Alzheimer’s Disease Is Affected by Aβ-ImmunoTherapy and Cognitive Stimulation. Pharmaceutics 2020, 12, 1–21. [Google Scholar] [CrossRef]
- Castillo-Mariqueo, L.; Giménez-Llort, L. Impact of Behavioral Assessment and Re-Test as Functional Trainings That Modify Survival, Anxiety and Functional Profile (Physical Endurance and Motor Learning) of Old Male and Female 3xTg-AD Mice and NTg Mice with Normal Aging. Biomedicines 2022, 10, 973. [Google Scholar] [CrossRef]
- Santana-Santana, M.; Bayascas, J.-R.; Giménez-Llort, L. Sex-Dependent Signatures, Time Frames and Longitudinal Fine-Tuning of the Marble Burying Test in Normal and AD-Pathological Aging Mice. Biomedicines 2021, 9, 994. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The Open Field as a Paradigm to Measure the Effects of Drugs on Anxiety-like Behaviors: A Review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Torres-Lista, V.; Parrado-Fernández, C.; Alvarez-Montón, I.; Frontiñán-Rubio, J.; Durán-Prado, M.; Peinado, J.R.; Johansson, B.; Alcaín, F.J.; Giménez-Llort, L. Neophobia, NQO1 and SIRT1 as Premorbid and Prodromal Indicators of AD in 3xTg-AD Mice. Behav. Brain Res. 2014, 271, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Careau, V.; Bininda-Emonds, O.R.P.; Ordonez, G.; Garland Jr, T. Are Voluntary Wheel Running and Open-Field Behavior Correlated in Mice? Different Answers from Comparative and Artificial Selection Approaches. Behav. Genet. 2012, 42, 830–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guayerbas, N.; Puerto, M.; Ferrández, M.D.; De La Fuente, M. A Diet Supplemented with Thiolic Anti-Oxidants Improves Leucocyte Function in Two Strains of Prematurely Ageing Mice. Clin. Exp. Pharmacol. Physiol. 2002, 29, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Roemers, P.; Hulst, Y.; van Heijningen, S.; van Dijk, G.; van Heuvelen, M.J.G.; De Deyn, P.P.; van der Zee, E.A. Inducing Physical Inactivity in Mice: Preventing Climbing and Reducing Cage Size Negatively Affect Physical Fitness and Body Composition. Front. Behav. Neurosci. 2019, 13, 221. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Estimate | SE | Z Value | p-Value |
|---|---|---|---|---|
| Model 1 | Revolutions~I (Days-1) ×Sex × Period + (1|Cage) | |||
| β0 | 29.71 | 2.68 | 11.09 | <0.001 (***) |
| βDays | 0.24 | 0.06 | 4.05 | <0.001 (***) |
| βSex | 1.11 | 3.79 | 0.29 | n.s. |
| βPeriod | −16.59 | 1.40 | −11.81 | <0.001 (***) |
| βDays,Sex | 0.10 | 0.08 | 1.20 | n.s. |
| βDays,Period | −0.09 | 0.08 | −1.12 | n.s. |
| βSex,Period | −5.61 | 1.99 | −2.82 | 0.004 (**) |
| βDays,Sex,Period | −0.17 | 0.12 | −1.41 | n.s. |
| Model 2 | Revolutions~I (Days-1) × Sex × NIBI × Period + (1|Cage) | |||
| β0 | 22.97 | 3.83 | 5.99 | <0.001 (***) |
| βDays | 0.48 | 0.07 | 6.47 | <0.001 (***) |
| βSex | 3.12 | 5.42 | 0.58 | n.s. |
| βNIBI | 13.48 | 5.42 | 2.49 | 0.013 (*) |
| βPeriod | −9.57 | 1.78 | −5.38 | <0.001 (***) |
| βDays,Sex | −0.08 | 0.11 | −0.73 | n.s. |
| βDays,NIBI | −0.49 | 0.11 | −4.62 | <0.001 (***) |
| βSex,NIBI | −4.02 | 7.67 | −0.52 | n.s. |
| βDays,Period | −0.17 | 0.11 | −1.63 | n.s. |
| βSex,Period | −9.59 | 2.52 | −3.81 | <0.001 (***) |
| βNIBI,Period | −14.04 | 2.52 | −5.58 | <0.001 (***) |
| βDays,Sex,NIBI | 0.35 | 0.15 | 2.38 | 0.018 (*) |
| βDays,Sex,Period | −0.24 | 0.15 | −1.60 | n.s. |
| βDays,NIBI,Period | 0.16 | 0.15 | 1.05 | n.s. |
| βSex,NIBI,Period | 7.96 | 3.56 | 2.24 | 0.025 (*) |
| βDays,Sex,NIBI,Period | 0.15 | 0.21 | 0.69 | n.s. |
| Rearing Latency (CT) | Center Latency (OF) | Total Grooming (OF) | Rocking (RR) | Grip 60 | Initial Weight | |
|---|---|---|---|---|---|---|
| Total activity (in 30 days) | 0.511 | 0.737 * | 0.669 | 0.719 * | 0.371 | 0.712 * |
| Mean nocturnal activity | 0.354 | 0.583 | 0.714 * | 0.635 | 0.482 | 0.746 * |
| Mean diurnal activity | 0.745 * | 0.644 | −0.101 | 0.557 | 0.007 | 0.308 |
| Mean daily activity | 0.491 | 0.681 | 0.642 | 0.711 * | 0.450 | 0.760 * |
| Mean nocturnal activity (days 1–10) | −0.114 | 0.229 | 0.605 | 0.101 | 0.721 * | 0.730 * |
| Mean diurnal activity (days 1–10) | 0.781 * | 0.592 | −0.013 | 0.688 | −0.038 | 0.215 |
| Ratio days 1–10 | −0.800 * | −0.174 | −0.582 | −0.674 | −0.048 | −0.051 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alveal-Mellado, D.; Castillo-Mariqueo, L.; Giménez-Llort, L. Sex- and Neuropsychiatric-Dependent Circadian Alterations in Daily Voluntary Physical Activity Engagement and Patterns in Aged 3xTg-AD Mice. Int. J. Mol. Sci. 2022, 23, 13671. https://doi.org/10.3390/ijms232213671
Alveal-Mellado D, Castillo-Mariqueo L, Giménez-Llort L. Sex- and Neuropsychiatric-Dependent Circadian Alterations in Daily Voluntary Physical Activity Engagement and Patterns in Aged 3xTg-AD Mice. International Journal of Molecular Sciences. 2022; 23(22):13671. https://doi.org/10.3390/ijms232213671
Chicago/Turabian StyleAlveal-Mellado, Daniel, Lidia Castillo-Mariqueo, and Lydia Giménez-Llort. 2022. "Sex- and Neuropsychiatric-Dependent Circadian Alterations in Daily Voluntary Physical Activity Engagement and Patterns in Aged 3xTg-AD Mice" International Journal of Molecular Sciences 23, no. 22: 13671. https://doi.org/10.3390/ijms232213671