
Highly Charged Ru(II) Polypyridyl Complexes as Photosensitizer Agents in Photodynamic Therapy of Epithelial Ovarian Cancer Cells

 , , , ,
, , , ,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
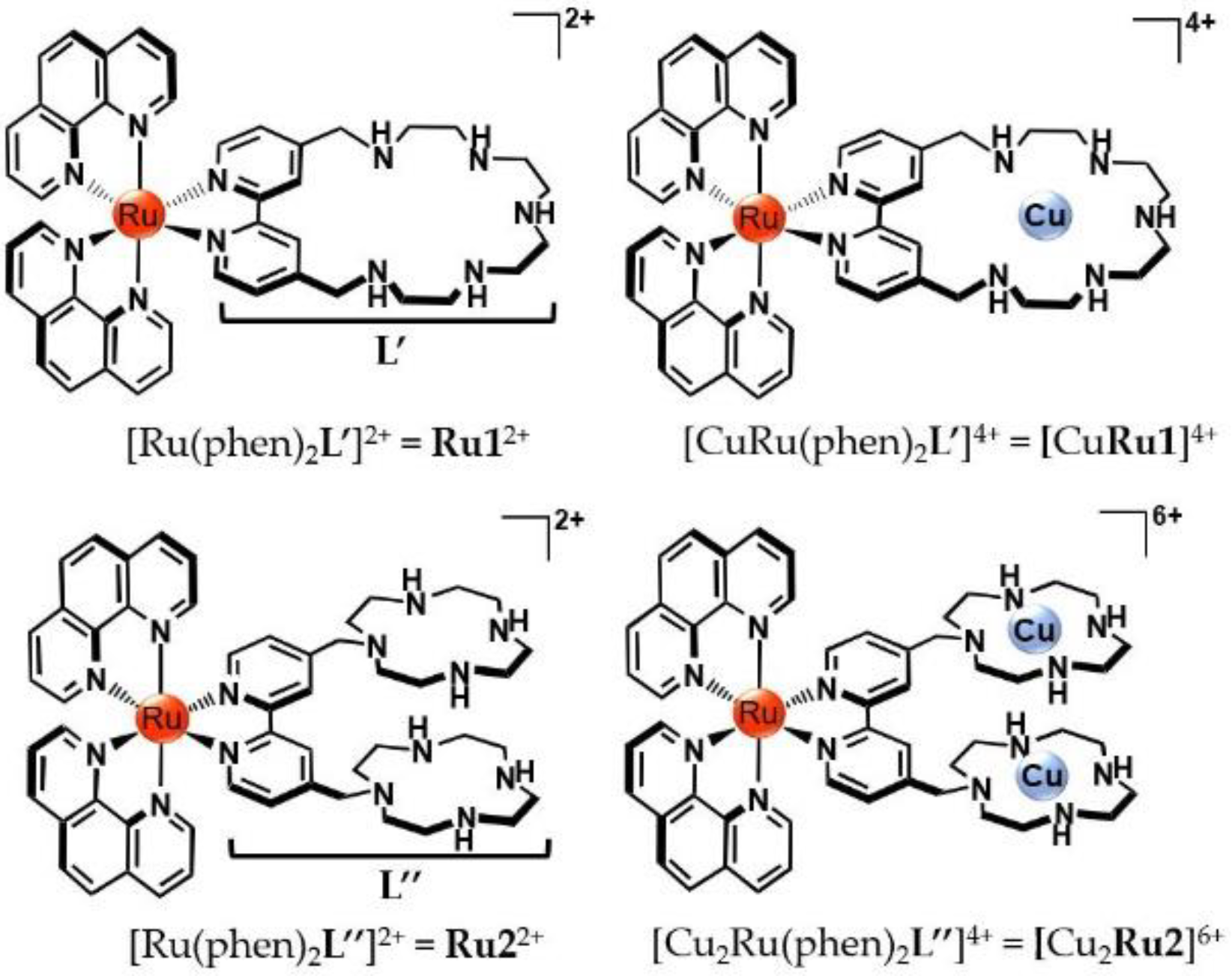
2.1. Protonation, Metal Binding of Ruthenium Compounds and Stability of the Mixed Ruthenium/Copper Complexed Species
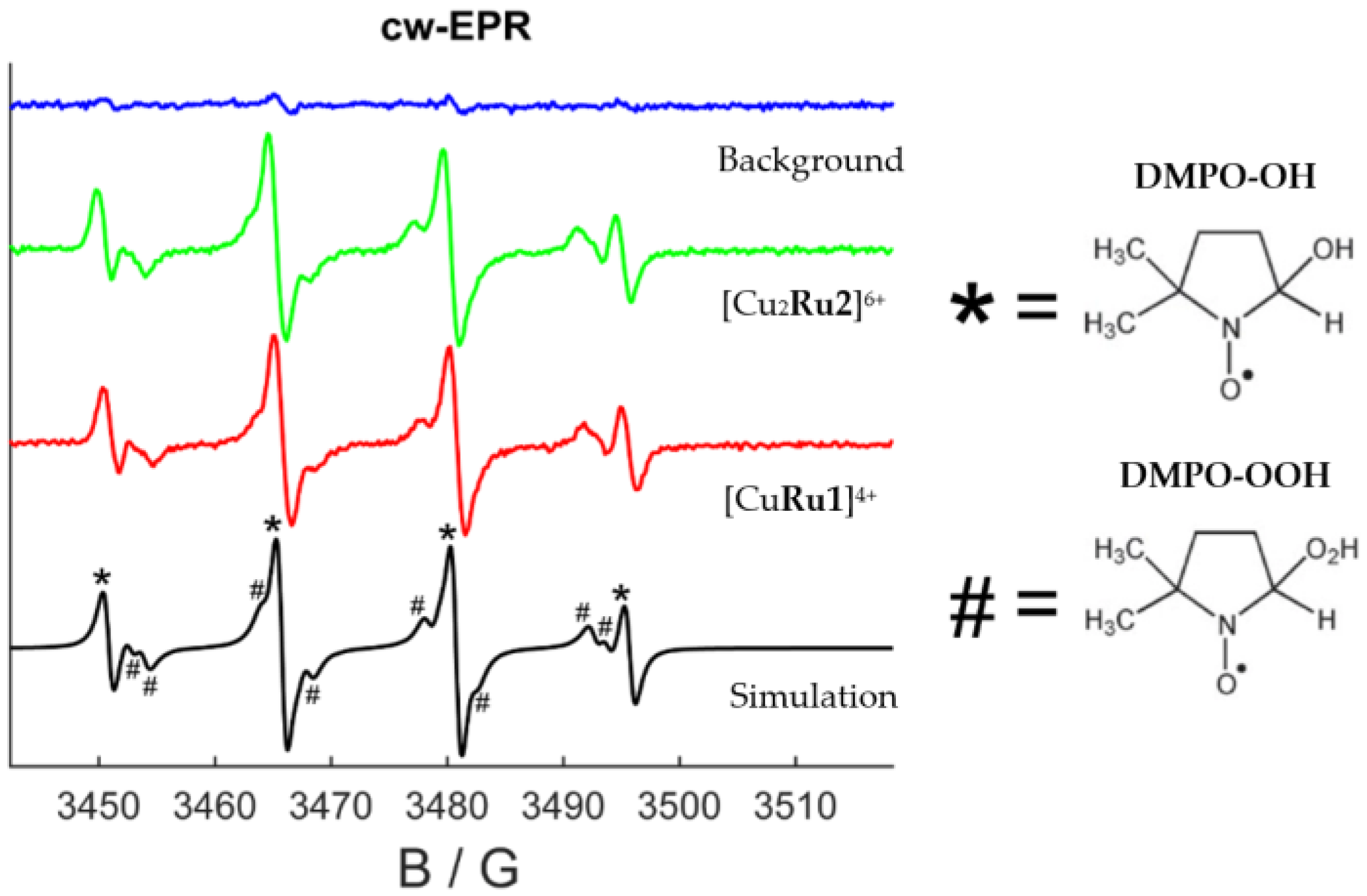
2.2. Reactive Oxygen Species (ROS) Production by Ruthenium Compounds
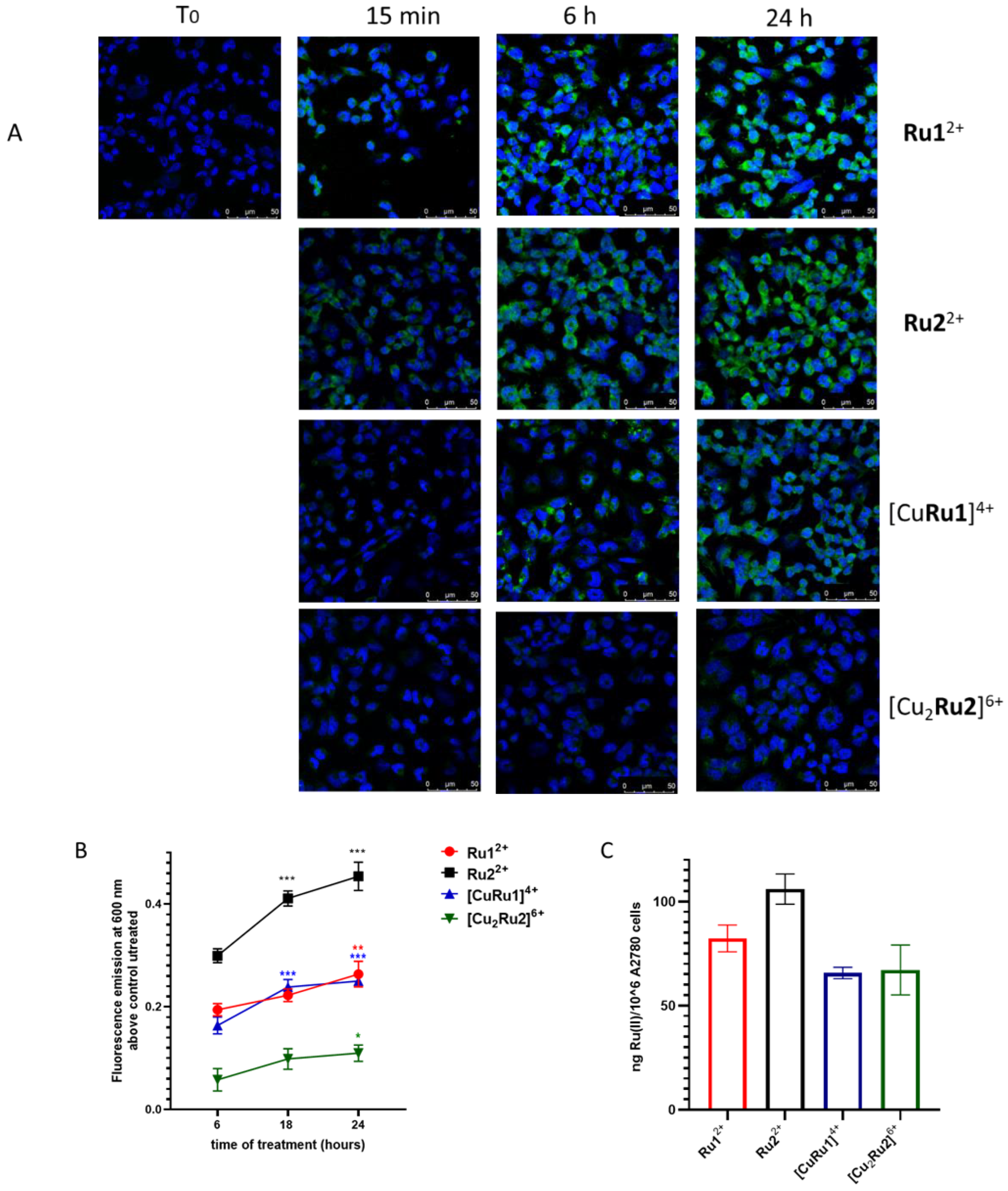
2.3. Internalization of Ru(II) Complexes
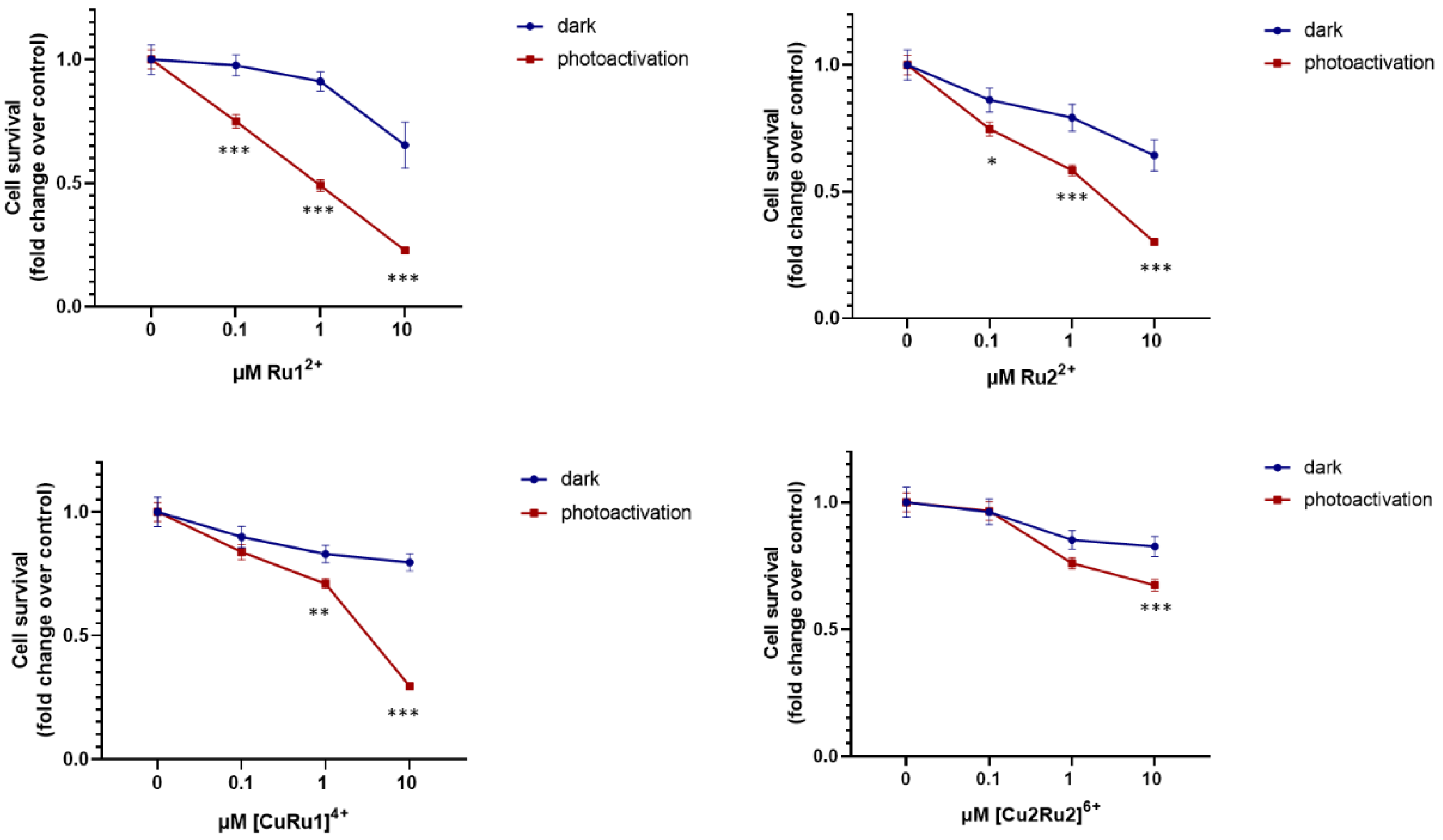
2.4. Effect of Ru(II) Complexes on A2780 Cell Survival after Photosensitization
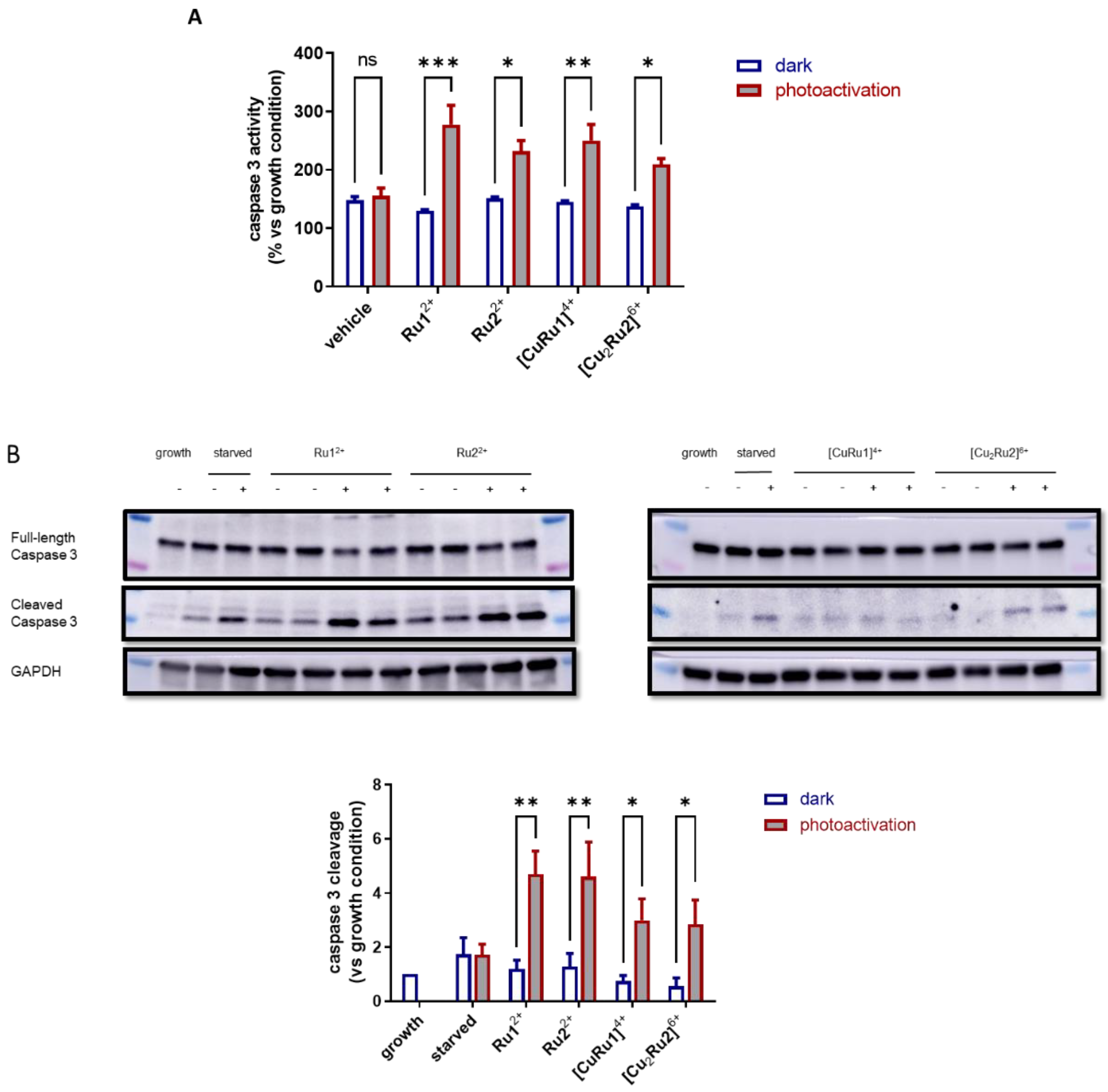
2.5. Effect of Ru(II) Complexes on Apoptosis of A2780 Cells after Photoactivation
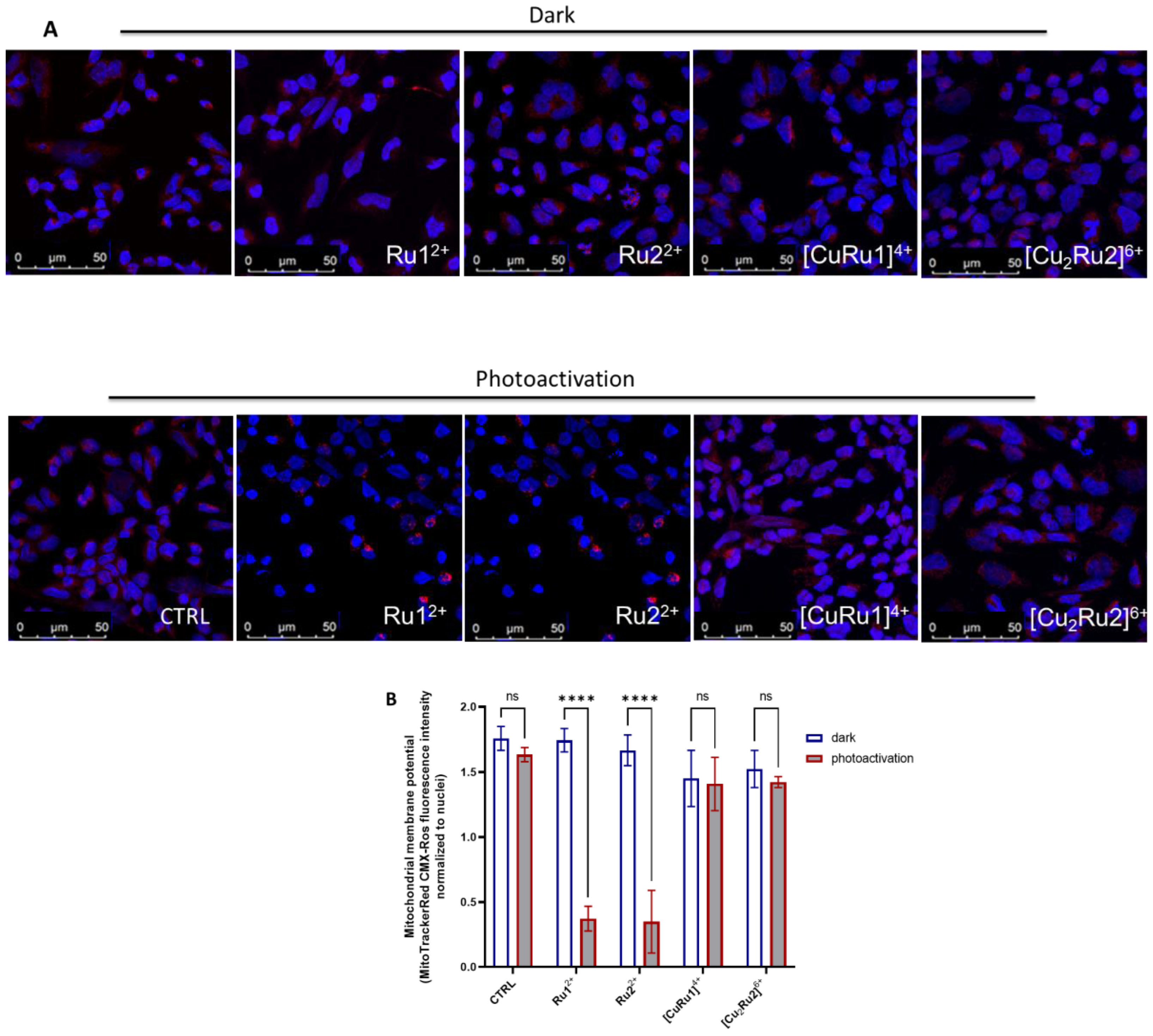
2.6. Mitochondrial Membrane Potential Is Lost after Photosensitization of Ru(II) Complexes
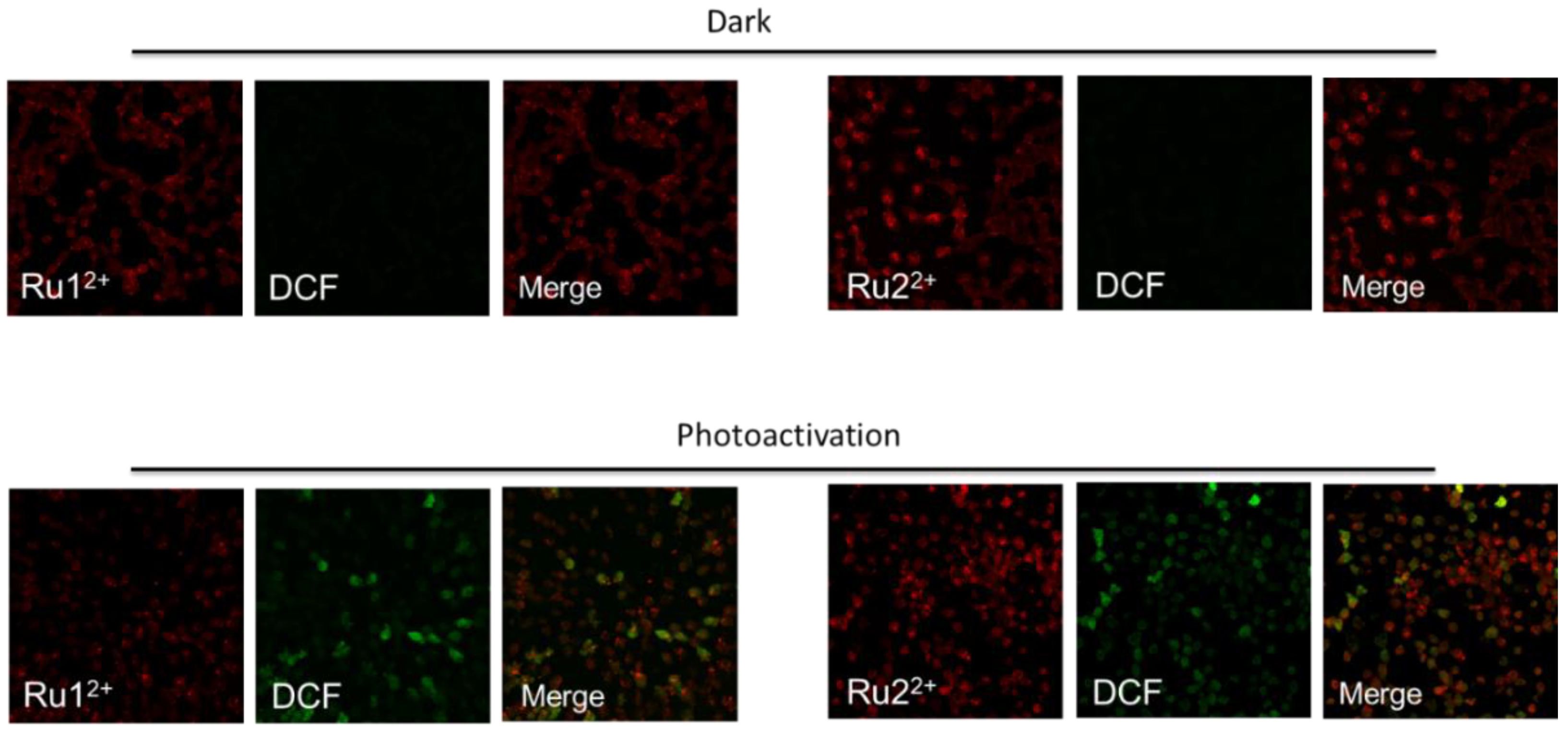
2.7. Cytosolic ROS Production after Photosensitization of Ru(II) Complexes Ru12+ and Ru22+
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of Ru(II)-Complexes
4.3. Potentiometric Measurements
4.4. EPR Measurements
4.5. Cell Culture
4.6. ICP-AES Measurements
4.7. MTT Reduction Assay for Cell Survival
4.8. Caspase-3 Activity Assay
4.9. Western Blot Analysis
4.10. Laser-Scanning Confocal Microscopy
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Coburn, S.B.; Bray, F.; Sherman, M.E.; Trabert, B. International patterns and trends in ovarian cancer incidence, overall and by histologic subtype. Int. J. Cancer 2017, 140, 2451–2460. [Google Scholar] [CrossRef] [Green Version]
- Buys, S.S.; Partridge, E.; Black, A.; Johnson, C.C.; Lamerato, L.; Isaacs, C.; Reding, D.J.; Greenlee, R.T.; Yokochi, L.A.; Kessel, B.; et al. Effect of Screening on Ovarian Cancer Mortality. JAMA 2011, 305, 2295–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikuła-Pietrasik, J.; Witucka, A.; Pakuła, M.; Uruski, P.; Begier-Krasińska, B.; Niklas, A.; Tykarski, A.; Książek, K. Comprehensive review on how platinum- and taxane-based chemotherapy of ovarian cancer affects biology of normal cells. Cell. Mol. Life Sci. 2019, 76, 681–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simović, A.R.; Masnikosa, R.; Bratsos, I.; Alessio, E. Chemistry and reactivity of ruthenium(II) complexes: DNA/protein binding mode and anticancer activity are related to the complex structure. Co-ord. Chem. Rev. 2019, 398, 113011. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, C.Y.; Nam, T.-G. Ruthenium Complexes as Anticancer Agents: A Brief History and Perspectives. Drug Des. Dev. Ther. 2020, 14, 5375–5392. [Google Scholar] [CrossRef]
- Kostova, I. Ruthenium Complexes as Anticancer Agents. Curr. Med. Chem. 2006, 13, 1085–1107. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, C.; Rees, T.W.; Ke, L.; Ji, L.; Chao, H. Harnessing ruthenium(II) as photodynamic agents: Encouraging advances in cancer therapy. Co-ord. Chem. Rev. 2018, 363, 17–28. [Google Scholar] [CrossRef]
- Poynton, F.E.; Bright, S.A.; Blasco, S.; Williams, D.C.; Kelly, J.M.; Gunnlaugsson, T. The development of ruthenium(II) polypyridyl complexes and conjugates for in vitro cellular and in vivo applications. Chem. Soc. Rev. 2017, 46, 7706–7756. [Google Scholar] [CrossRef]
- Monro, S.; Colón, K.L.; Yin, H.; Roque, J., III; Konda, P.; Gujar, S.; Thummel, R.P.; Lilge, L.; Cameron, C.G.; McFarland, S.A. Transition Metal Complexes and Photodynamic Therapy from a Tumor-Centered Approach: Challenges, Opportunities, and Highlights from the Development of TLD1433. Chem. Rev. 2019, 119, 797–828. [Google Scholar] [CrossRef]
- Lin, K.; Zhao, Z.-Z.; Bo, H.-B.; Hao, X.-J.; Wang, J.-Q. Applications of Ruthenium Complex in Tumor Diagnosis and Therapy. Front. Pharmacol. 2018, 9, 1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mari, C.; Pierroz, V.; Ferrari, S.; Gasser, G. Combination of Ru(II) complexes and light: New frontiers in cancer therapy. Chem. Sci. 2015, 6, 2660–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gall, T.; Lemercier, G.; Chevreux, S.; Tucking, K.-S.; Ravel, J.; Thetiot, F.; Jonas, U.; Schonherr, H.; Montier, T. Ruthenium(II) Polypyridyl Complexes as Photosensitizers for Antibacterial Photodynamic Therapy: A Structure-Activity Study on Clinical Bacterial Strains. ChemMedChem 2018, 13, 2229–2239. [Google Scholar] [CrossRef]
- Giacomazzo, G.E.; Conti, L.; Guerri, A.; Pagliai, M.; Fagorzi, C.; Sfragano, P.S.; Palchetti, I.; Pietraperzia, G.; Mengoni, A.; Valtancoli, B.; et al. Nitroimidazole-Based Ruthenium(II) Complexes: Playing with Structural Parameters to Design Photostable and Light-Responsive Antibacterial Agents. Inorg. Chem. 2021, 61, 6689–6694. [Google Scholar] [CrossRef] [PubMed]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in photodynamic therapy: Part one—Photosensitizers, photochemistry and cellular localization. Photodiagn. Photodyn. Ther. 2004, 1, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Conti, L.; Bencini, A.; Ferrante, C.; Gellini, C.; Paoli, P.; Parri, M.; Pietraperzia, G.; Valtancoli, B.; Giorgi, C. Highly Charged Ruthenium(II) Polypyridyl Complexes as Effective Photosensitizer in Photodynamic Therapy. Chem.–A Eur. J. 2019, 25, 10606–10615. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Mengoni, A.; Giacomazzo, G.E.; Mari, L.; Perfetti, M.; Fagorzi, C.; Sorace, L.; Valtancoli, B.; Giorgi, C. Exploring the potential of highly charged Ru(II)- and heteronuclear Ru(II)/Cu(II)-polypyridyl complexes as antimicrobial agents. J. Inorg. Biochem. 2021, 220, 111467. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Flore, N.; Formica, M.; Giorgi, L.; Pagliai, M.; Mancini, L.; Fusi, V.; Valtancoli, B.; Giorgi, C. Glyphosate and AMPA binding by two polyamino-phenolic ligands and their dinuclear Zn(II) complexes. Inorg. Chim. Acta 2021, 519, 120261. [Google Scholar] [CrossRef]
- Juarranz, Á.; Jaén, P.; Sanz-Rodríguez, F.; Cuevas, J.; González, S. Photodynamic therapy of cancer. Basic principles and applications. Clin. Transl. Oncol. 2008, 10, 148–154. [Google Scholar] [CrossRef]
- Ochsner, M. Photophysical and photobiological processes in the photodynamic therapy of tumours. J. Photochem. Photobiol. B Biol. 1997, 39, 1–18. [Google Scholar] [CrossRef]
- Pineiro, M.; Pereira, M.M.; Gonsalves, A.M.D.R.; Arnaut, L.; Formosinho, S. Singlet oxygen quantum yields from halogenated chlorins: Potential new photodynamic therapy agents. J. Photochem. Photobiol. A Chem. 2001, 138, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Yu, H.; Dong, Y.; Tian, R.; Huang, G.; Boothman, D.A.; Sumer, B.D.; Gao, J. Photoactivation switch from type II to type I reactions by electron-rich micelles for improved photodynamic therapy of cancer cells under hypoxia. J. Control. Release 2011, 156, 276–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucková, L.; Jomová, K.; Švorcová, A.; Valko, M.; Segľa, P.; Moncoľ, J.; Kožíšek, J. Synthesis, Crystal Structure, Spectroscopic Properties and Potential Biological Activities of Salicylate–Neocuproine Ternary Copper(II) Complexes. Molecules 2015, 20, 2115–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, K.L.; Franz, K.J. Application of Metal Coordination Chemistry To Explore and Manipulate Cell Biology. Chem. Rev. 2009, 109, 4921–4960. [Google Scholar] [CrossRef] [Green Version]
- Kakar, S.S.; Jala, V.R.; Fong, M.Y. Synergistic cytotoxic action of cisplatin and withaferin A on ovarian cancer cell lines. Biochem. Biophys. Res. Commun. 2012, 423, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Ricci, J.E.; Waterhouse, N.; Green, D.R. Mitochondrial functions during cell death, a complex (I–V) dilemma. Cell Death Differ. 2003, 10, 488–492. [Google Scholar] [CrossRef]
- Hu, X.-M.; Li, Z.-X.; Lin, R.-H.; Shan, J.-Q.; Yu, Q.-W.; Wang, R.-X.; Liao, L.-S.; Yan, W.-T.; Wang, Z.; Shang, L.; et al. Guidelines for Regulated Cell Death Assays: A Systematic Summary, A Categorical Comparison, A Prospective. Front. Cell Dev. Biol. 2021, 9, 634690. [Google Scholar] [CrossRef]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- Rocconi, R.P.; Case, A.S.; Michael Straughn, J., Jr.; Estes, J.M.; Partridge, E.E. Role of chemotherapy for patients with recurrent platinum-resistant advanced epithelial ovarian cancer. Cancer 2006, 107, 536–543. [Google Scholar] [CrossRef]
- Au, K.K.; Josahkian, J.A.; Francis, J.A.; Squire, J.A.; Koti, M. Current state of biomarkers in ovarian cancer prognosis. Futur. Oncol. 2015, 11, 3187–3195. [Google Scholar] [CrossRef]
- Kurman, R.J.; Shih, I.-M. The Dualistic Model of Ovarian Carcinogenesis: Revisited, revised, and expanded. Am. J. Pathol. 2016, 186, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Thota, S.; Rodrigues, D.A.; Crans, D.; Barreiro, E.J. Ru(II) Compounds: Next-Generation Anticancer Metallotherapeutics? J. Med. Chem. 2018, 61, 5805–5821. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Gupta, P.; Chen, Y.; Wang, E.; Ji, L.; Chao, H.; Chen, Z.-S. The development of anticancer ruthenium(II) complexes: From single molecule compounds to nanomaterials. Chem. Soc. Rev. 2017, 46, 5771–5804. [Google Scholar] [CrossRef]
- Berndsen, R.H.; Weiss, A.; Abdul, U.K.; Wong, T.J.; Meraldi, P.; Griffioen, A.W.; Dyson, P.; Nowak-Sliwinska, P. Combination of ruthenium(II)-arene complex [Ru(η6-p-cymene)Cl2(pta)] (RAPTA-C) and the epidermal growth factor receptor inhibitor erlotinib results in efficient angiostatic and antitumor activity. Sci. Rep. 2017, 7, srep43005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.-Y.; Chen, J.; Shao, J.; Jiao, Z.-Q.; Tang, T.-S.; Tang, M.; Sheng, Z.-G.; Mao, L.; Huang, R.; Huang, C.-H.; et al. The cell-impermeable Ru(II) polypyridyl complex as a potent intracellular photosensitizer under visible light irradiation via ion-pairing with suitable lipophilic counter-anions. Free Radic. Biol. Med. 2021, 171, 69–79. [Google Scholar] [CrossRef]
- Spreckelmeyer, S.; Orvig, C.; Casini, A. Cellular Transport Mechanisms of Cytotoxic Metallodrugs: An Overview beyond Cisplatin. Molecules 2014, 19, 15584–15610. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (deltapsi(m)) in apoptosis; an update. Apoptosis Int. J. Program. Cell Death 2003, 8, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Mummolo, L.; Romano, G.; Giorgi, C.; Giacomazzo, G.; Prodi, L.; Bencini, A. Exploring the Ability of Luminescent Metal Assemblies to Bind and Sense Anionic or Ionizable Analytes A Ru(phen)2bipy-Based Dizinc Complex for Bisphenol A (BPA) Recognition. Molecules 2021, 26, 527. [Google Scholar] [CrossRef]
- Aragoni, M.C.; Arca, M.; Bencini, A.; Caltagirone, C.; Conti, L.; Garau, A.; Valtancoli, B.; Isaia, F.; Lippolis, V.; Palomba, F.; et al. Zn2+/Cd2+ optical discrimination by fluorescent acridine-based bis-macrocylic receptors. Supramol. Chem. 2017, 29, 912–921. [Google Scholar] [CrossRef]
- Conti, L.; Giorgi, C.; Valtancoli, B.; Paoli, P.; Rossi, P.; Marchionni, A.; Faggi, E.; Bencini, A. Switching on the Fluorescence Emission of Polypyridine Ligands by Simultaneous Zinc(II) Binding and Protonation. ChemPlusChem 2020, 85, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Garau, A.; Bencini, A.; Blake, A.J.; Caltagirone, C.; Conti, L.; Isaia, F.; Lippolis, V.; Montis, R.; Mariani, P.; Scorciapino, M.A. [9]aneN3-based fluorescent receptors for metal ion sensing, featuring urea and amide functional groups. Dalton Trans. 2019, 48, 4949–4960. [Google Scholar] [CrossRef] [PubMed]
- Becatti, M.; Bencini, A.; Nistri, S.; Conti, L.; Fabbrini, M.G.; Lucarini, L.; Ghini, V.; Severi, M.; Fiorillo, C.; Giorgi, C.; et al. Different Antioxidant Efficacy of Two MnII-Containing Superoxide Anion Scavengers on Hypoxia/Reoxygenation-Exposed Cardiac Muscle Cells. Sci. Rep. 2019, 9, 10320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernacchioni, C.; Ghini, V.; Cencetti, F.; Japtok, L.; Donati, C.; Bruni, P.; Turano, P. NMR metabolomics highlights sphingosine kinase-1 as a new molecular switch in the orchestration of aberrant metabolic phenotype in cancer cells. Mol. Oncol. 2017, 11, 517–533. [Google Scholar] [CrossRef] [Green Version]
- Conti, L.; Ciambellotti, S.; Giacomazzo, G.E.; Ghini, V.; Cosottini, L.; Puliti, E.; Severi, M.; Fratini, E.; Cencetti, F.; Bruni, P.; et al. Ferritin nanocomposites for the selective delivery of photosensitizing ruthenium-polypyridyl compounds to cancer cells. Inorg. Chem. Front. 2022, 9, 1070–1081. [Google Scholar] [CrossRef]
- Donati, C.; Cencetti, F.; De Palma, C.; Rapizzi, E.; Brunelli, S.; Cossu, G.; Clementi, E.; Bruni, P. TGFβ protects mesoangioblasts from apoptosis via sphingosine kinase-1 regulation. Cell. Signal. 2009, 21, 228–236. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Rosen, G.M.; Beselman, A.; Tsai, P.; Pou, S.; Mailer, C.; Ichikawa, K.; Robinson, B.H.; Nielsen, R.; Halpern, H.J.; MacKerell, A.D. Influence of Conformation on the EPR Spectrum of 5,5-Dimethyl-1-hydroperoxy-1-pyrrolidinyloxyl: A Spin Trapped Adduct of Superoxide. J. Org. Chem. 2004, 69, 1321–1330. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction | LogK | |
|---|---|---|
| L = Ru1 | L = Ru2 | |
| L2+ + Cu2+ = CuL4+ | 15.34 (6) a | 16.72 (8) |
| CuL4+ + H+ = CuHL5+ | 5.60 (6) | 7.58 (7) |
| CuHL5+ + H+ = CuH2L6+ | 3.91 (4) | 5.89 (5) |
| CuL4+ + OH− = CuL(OH)3+ | 6.06 (4) | |
| L2+ + 2Cu2+ + OH− = Cu2L(OH)5+ | 12.95 (5) | |
| CuL4+ + Cu2+ = Cu2L6+ | 10.88 (7) | |
| Cu2L6+ + 2OH− = Cu2L(OH)24+ | 9.48 (6) | |
| L2+ + Zn2+ = ZnL4+ | 8.90 (7) | 14.91 (5) |
| ZnL4+ + H+ = ZnHL5+ | 6.36 (7) | 8.65 (4) |
| ZnHL5+ + H+ = ZnH2L6+ | 5.56 (6) | 7.16 (5) |
| ZnH2L6+ + H+ = ZnH3L7+ | 4.75 (8) | |
| ZnL4+ + 2OH− = ZnL(OH)22+ | 11.55 (4) | |
| ZnL4+ + Zn2+ = Zn2L6+ | 5.42 (7) | |
| Zn2L6+ + 3OH- = Zn2L(OH)33+ | 15.48 (6) | |
| L2+ + Ca2+ = CaL4+ | 2.62 (5) | 2.55 (6) |
| L2+ + Mg2+ = MgL4+ | 2.23 (5) | 2.06 (6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, L.; Giacomazzo, G.E.; Valtancoli, B.; Perfetti, M.; Privitera, A.; Giorgi, C.; Sfragano, P.S.; Palchetti, I.; Pecchioli, S.; Bruni, P.; et al. Highly Charged Ru(II) Polypyridyl Complexes as Photosensitizer Agents in Photodynamic Therapy of Epithelial Ovarian Cancer Cells. Int. J. Mol. Sci. 2022, 23, 13302. https://doi.org/10.3390/ijms232113302
Conti L, Giacomazzo GE, Valtancoli B, Perfetti M, Privitera A, Giorgi C, Sfragano PS, Palchetti I, Pecchioli S, Bruni P, et al. Highly Charged Ru(II) Polypyridyl Complexes as Photosensitizer Agents in Photodynamic Therapy of Epithelial Ovarian Cancer Cells. International Journal of Molecular Sciences. 2022; 23(21):13302. https://doi.org/10.3390/ijms232113302
Chicago/Turabian StyleConti, Luca, Gina Elena Giacomazzo, Barbara Valtancoli, Mauro Perfetti, Alberto Privitera, Claudia Giorgi, Patrick Severin Sfragano, Ilaria Palchetti, Sara Pecchioli, Paola Bruni, and et al. 2022. "Highly Charged Ru(II) Polypyridyl Complexes as Photosensitizer Agents in Photodynamic Therapy of Epithelial Ovarian Cancer Cells" International Journal of Molecular Sciences 23, no. 21: 13302. https://doi.org/10.3390/ijms232113302