
The Prognostic Significance of FKBP1A and Its Related Immune Infiltration in Liver Hepatocellular Carcinoma
Abstract
:1. Introduction
2. Results
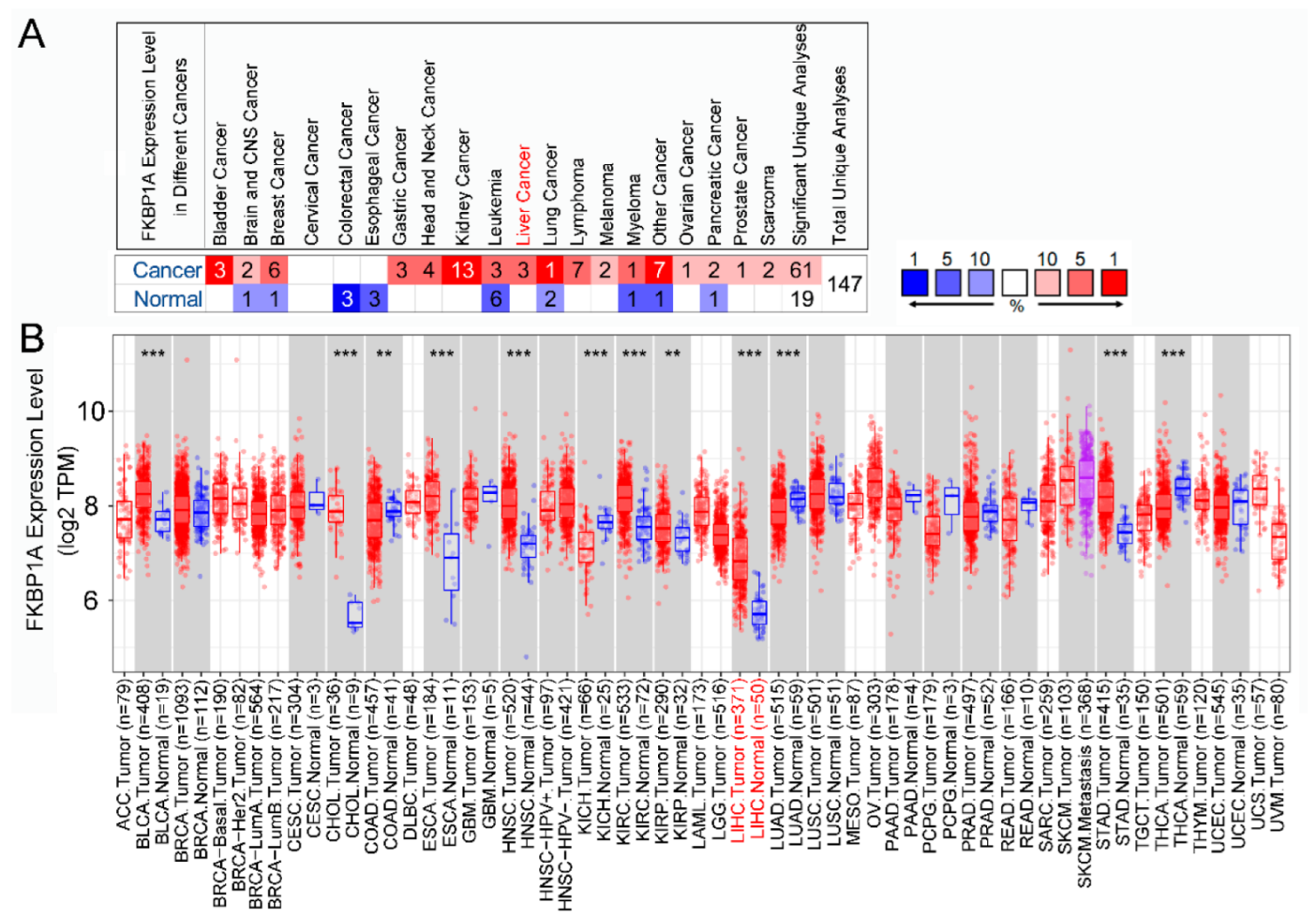
2.1. Differential Expression of FKBP1A in Different Cancers
2.2. Expression of FKBP1A Is Upregulated in Liver Hepatocellular Carcinoma
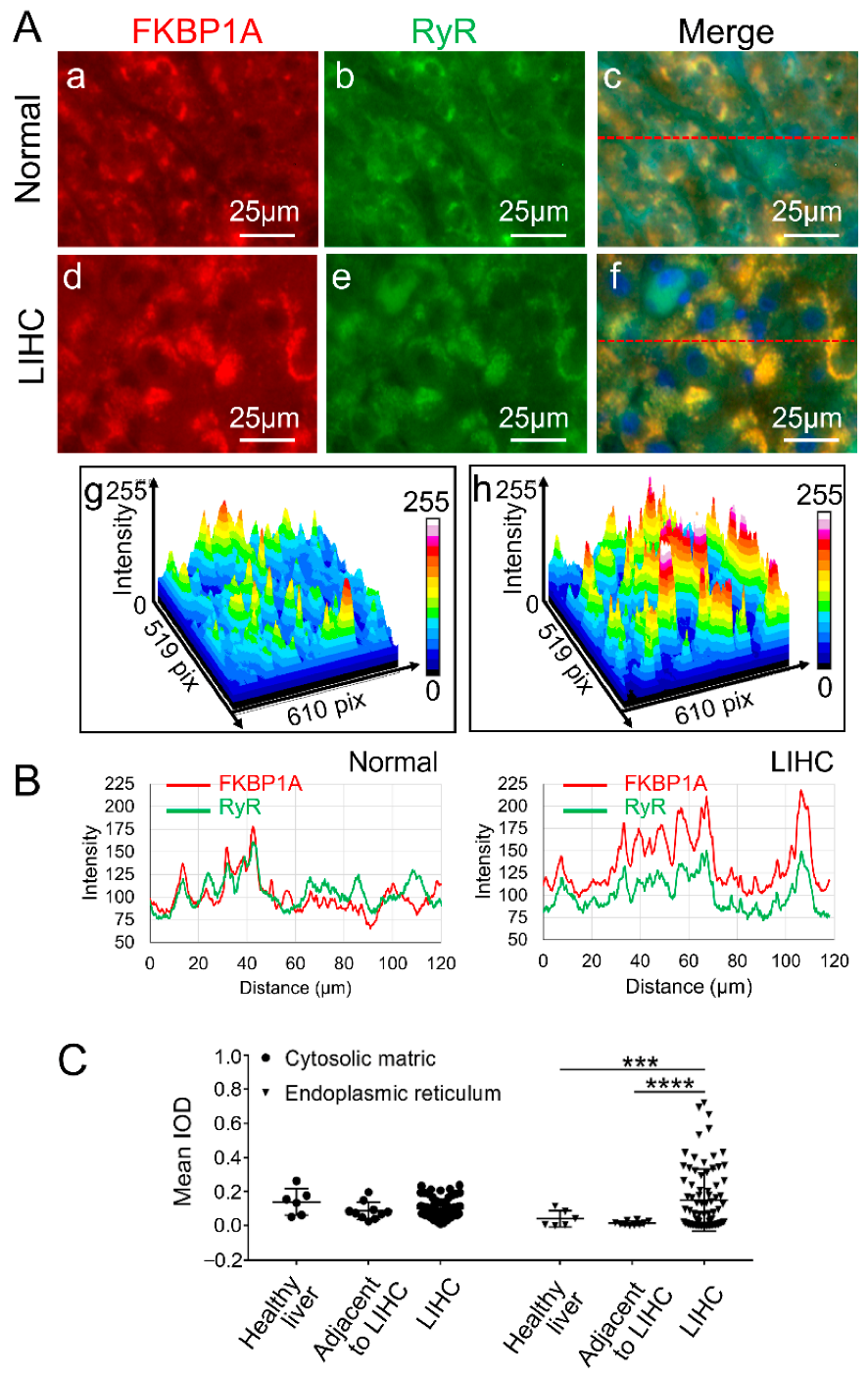
2.3. Upregulated FKBP1A Protein Is Accumulated in Endoplasmic Reticulum in Liver Hepatocellular Carcinoma
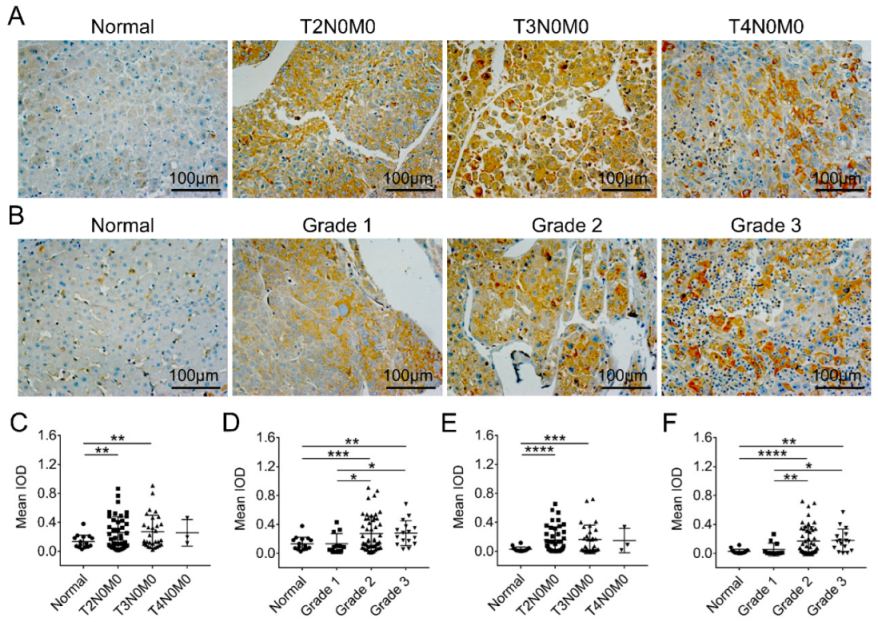
2.4. Intracellular Abnormality of FKBP1A Expression Distribution Is Accompanied by ER-Accumulation during Developing Severity of Hepatocellular Carcinoma
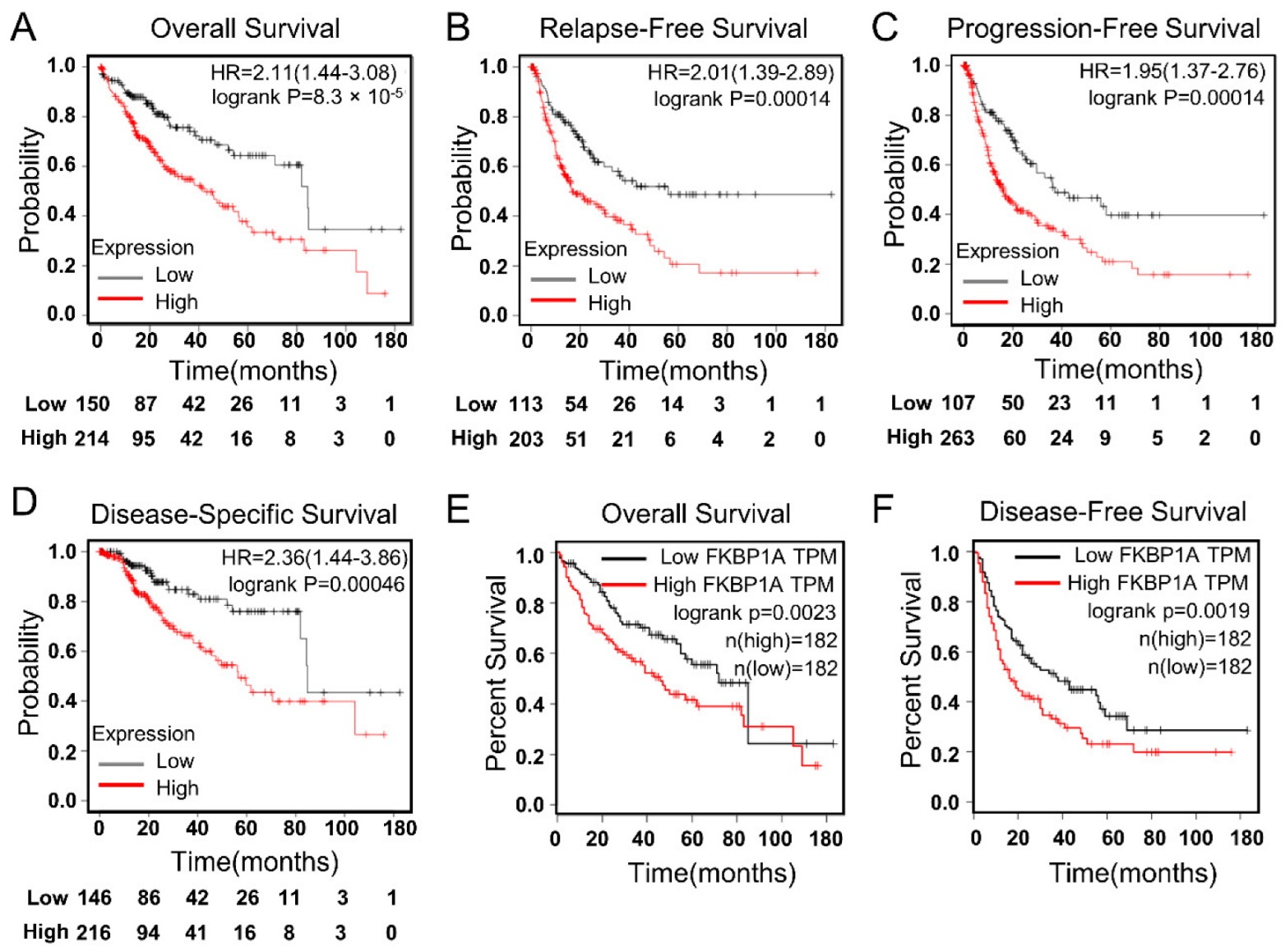
2.5. FKBP1A Expression Could Be Critical Factor for the Prognosis of LIHC Patients
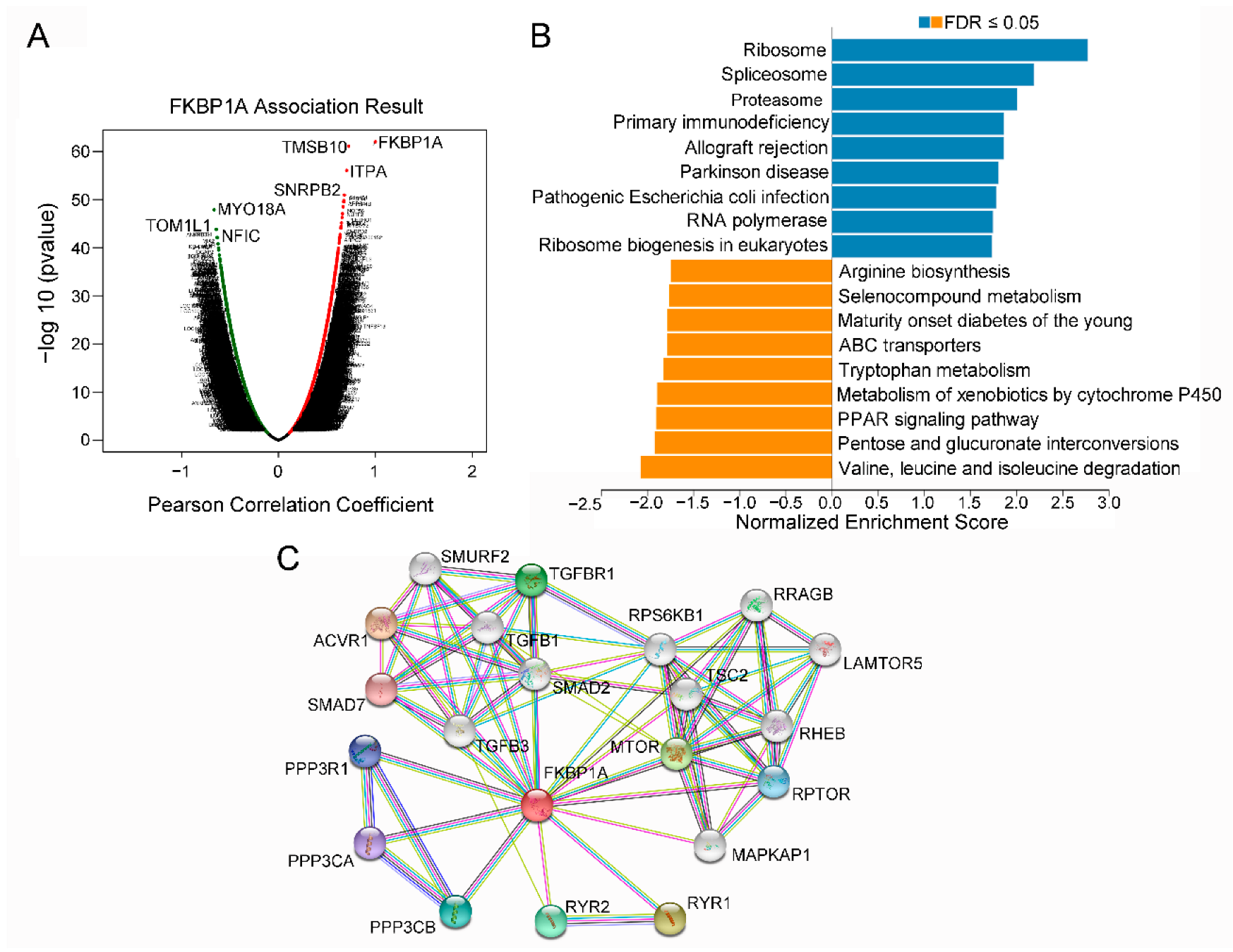
2.6. The Co-Expression Networks of FKBP1A Directs to Immunodeficiency Pathway in LIHC
2.7. Immune Infiltration Analysis of FKBP1A in LIHC
3. Discussion
4. Materials and Methods
4.1. Oncomine Database Analysis
4.2. TIMER Database Analysis
4.3. GEPIA Database Analysis
4.4. HCCDB Database Analysis
4.5. Kaplan–Meier Plotter Analysis
4.6. LinkedOmics Database Analysis
4.7. UALCAN Database Analysis
4.8. STRING Database Analysis
4.9. Tissue Microarrays (TMA) and Immunohistochemistry (IHC)
4.9.1. Information on Human Liver Tissue Microarray
4.9.2. The Process of Immunohistochemical and Immunofluorescence Staining
4.9.3. Statistical Analysis of IHC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, C.S.; Lin, Y.R.; Teng, T.H.; Lin, P.Y.; Tu, S.J.; Chou, C.H.; Huang, Y.R.; Huang, W.C.; Weng, S.L.; Huang, H.D.; et al. Haptoglobin expression correlates with tumor differentiation and five-year overall survival rate in hepatocellular carcinoma. PLoS ONE 2017, 12, e0171269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Yin, T.; Xu, Y.; Lu, X.J. Therapeutics for advanced hepatocellular carcinoma: Recent advances, current dilemma, and future directions. J. Cell. Physiol. 2019, 234, 12122–12132. [Google Scholar] [CrossRef]
- Sangro, B.; Sarobe, P.; Hervás-Stubbs, S.; Melero, I. Advances in immunotherapy for hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 525–543. [Google Scholar] [CrossRef]
- Ghartey-Kwansah, G.; Li, Z.; Feng, R.; Wang, L.; Xu, M.M.; Jones, O.; Bryant, J.; Isaacs, W.B.; Ma, J.; Xu, X.H.; et al. Comparative analysis of FKBP family protein: Evaluation, structure, and function in mammals and Drosophila melanogaster. BMC Dev. Biol. 2018, 18, 7. [Google Scholar] [CrossRef] [Green Version]
- Annett, S.; Moore, G.; Robson, T. FK506 binding proteins and inflammation related signalling pathways; basic biology, current status and future prospects for pharmacological intervention. Pharmacol. Ther. 2020, 215, 107623. [Google Scholar] [CrossRef]
- Tong, M.; Jiang, Y. FK506-Binding Proteins and Their Diverse Functions. Curr. Mol. Pharmacol. 2015, 9, 48–65. [Google Scholar] [CrossRef]
- Hoeffer, C.A.; Tang, W.; Wong, H.; Santillan, A.; Patterson, R.J.; Martinez, L.A.; Tejada-Simon, M.V.; Paylor, R.; Hamilton, S.L.; Klann, E. Removal of FKBP12 enhances mTOR-Raptor interactions, LTP, memory, and perseverative/repetitive behavior. Neuron 2008, 60, 832–845. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.M.; Steiner, J.P.; Roskams, A.J.; Ali, S.M.; Ronnett, G.V.; Snyder, S.H. Calcineurin associated with the inositol 1,4,5-trisphosphate receptor-FKBP12 complex modulates Ca2+ flux. Cell 1995, 83, 463–472. [Google Scholar] [CrossRef]
- Aizpurua, J.M.; Miranda, J.I.; Irastorza, A.; Torres, E.; Eceiza, M.; Sagartzazu-Aizpurua, M.; Ferrón, P.; Aldanondo, G.; Lasa-Fernández, H.; Marco-Moreno, P.; et al. Discovery of a novel family of FKBP12 “reshapers” and their use as calcium modulators in skeletal muscle under nitro-oxidative stress. Eur. J. Med. Chem. 2021, 213, 113160. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Wang, J.; Yang, Q.; Wang, Y.; Li, J.; Xiong, J.; Zhou, S. FKBP12 is a predictive biomarker for efficacy of anthracycline-based chemotherapy in breast cancer. Cancer Chemother. Pharmacol. 2019, 84, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Xiong, J.; Yi, S.; Zhang, H.; Zhou, S.; Gu, L.; Zhou, M. FKBP12 enhances sensitivity to chemotherapy-induced cancer cell apoptosis by inhibiting MDM2. Oncogene 2017, 36, 1678–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.H.; Balk, S.P.; Isaacs, W.B.; Ma, J. Calcium signaling: An underlying link between cardiac disease and carcinogenesis. Cell Biosci. 2018, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xu, M.M.; Li, Z.; Zhou, X.; Jiang, X.; Bryant, J.; Balk, S.; Ma, J.; Isaacs, W.B.; Xu, X.H.; et al. Calcium and CaSR/IP3R in prostate cancer development. Cell Biosci. 2018, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, Z.; Wang, Z.; Chen, E.; Jones, O.; Tan, T.; Takeshima, H.; Bryant, J.; Ma, J.; Xu, X.H.; et al. Syncytium calcium signaling and macrophage function in the heart. Cell Biosci. 2018, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Leone, P.; Solimando, A.G.; Fasano, R.; Argentiero, A.; Malerba, E.; Buonavoglia, A.; Lupo, L.G.; De Re, V.; Silvestris, N.; Racanelli, V. The evolving role of immune checkpoint inhibitors in hepatocellular carcinoma treatment. Vaccines 2021, 9, 532. [Google Scholar] [CrossRef]
- Xing, R.; Gao, J.; Cui, Q.; Wang, Q. Strategies to improve the antitumor effect of immunotherapy for hepatocellular carcinoma. Front. Immunol. 2021, 12, 783236. [Google Scholar] [CrossRef]
- Patel, D.; Dabhi, A.M.; Dmello, C.; Seervi, M.; Sneha, K.M.; Agrawal, P.; Sahani, M.H.; Kanojia, D. FKBP1A upregulation correlates with poor prognosis and increased metastatic potential of HNSCC. Cell Biol. Int. 2022, 46, 443–453. [Google Scholar] [CrossRef]
- Leng, W.; Liu, Q.; Zhang, S.; Sun, D.; Guo, Y. LncRNA AFAP1-AS1 modulates the sensitivity of paclitaxel-resistant prostate cancer cells to paclitaxel via miR-195-5p/FKBP1A axis. Cancer Biol. Ther. 2020, 21, 1072–1080. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, K.; Zhao, Z.; Qin, Z.; Tang, H. Prognosis-related autophagy genes in female lung adenocarcinoma. Medicine 2022, 101, e28500. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Chen, F.; Wu, C.; Wang, J.; Chen, S.S.; Li, X.R.; Wang, J.; Wu, L.; Ding, J.P.; Wang, J.C.; et al. Rapamycin recruits SIRT2 for FKBP12 deacetylation during mTOR activity modulation in innate immunity. iScience 2021, 24, 103177. [Google Scholar] [CrossRef] [PubMed]
- Boulch, M.; Grandjean, C.L.; Cazaux, M.; Bousso, P. Tumor Immunosurveillance and Immunotherapies: A Fresh Look from Intravital Imaging. Trends Immunol. 2019, 40, 1022–1034. [Google Scholar] [CrossRef] [PubMed]
- Kalbasi, A.; Ribas, A. Tumour-intrinsic resistance to immune checkpoint blockade. Nat. Rev. Immunol. 2020, 20, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Shi, Z.; Zhou, Y.; Zhang, Z.; Zhou, Y.; Chen, B.; Zhang, Q. Autophagy-related signatures as prognostic indicators for hepatocellular carcinoma. Front. Oncol. 2022, 12, 654449. [Google Scholar] [CrossRef] [PubMed]
- Shou, W.; Aghdasi, B.; Armstrong, D.L.; Guo, Q.; Bao, S.; Charng, M.J.; Mathews, L.M.; Schneider, M.D.; Hamilton, S.L.; Matzuk, M.M. Cardiac defects and altered ryanodine receptor function in mice lacking FKBP12. Nature 1998, 391, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, M.; Li, B.Y.; Chen, H.; Xu, X.H.; Song, L.S.; Guatimosim, S.; Zhu, W.; Yong, W.; Zhang, W.; Bu, G.; et al. FKBP12 is a critical regulator of the heart rhythm and the cardiac voltage-gated sodium current in mice. Circ. Res. 2011, 108, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. Nat. Rev. Immunol. 2019, 19, 369–382. [Google Scholar] [CrossRef]
- Ning, J.; Ye, Y.; Bu, D.; Zhao, G.; Song, T.; Liu, P.; Yu, W.; Wang, H.; Li, H.; Ren, X.; et al. Imbalance of TGF-β1/BMP-7 pathways induced by M2-polarized macrophages promotes hepatocellular carcinoma aggressiveness. Mol. Ther. 2021, 29, 2067–2087. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; He, Y.; Luo, N.; Patel, S.J.; Han, Y.; Gao, R.; Modak, M.; Carotta, S.; Haslinger, C.; Kind, D.; et al. Landscape and dynamics of single immune cells in hepatocellular carcinoma. Cell 2019, 179, 829–845.e820. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ge, W.; Zhou, J.; Gao, B.; Qian, X.; Wang, W. The role of tumor associated macrophages in hepatocellular carcinoma. J. Cancer 2021, 12, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Iñarrairaegui, M.; Melero, I.; Sangro, B. Immunotherapy of hepatocellular carcinoma: Facts and hopes. Clin. Cancer Res. 2018, 24, 1518–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef]
- Fasano, R.; Malerba, E.; Prete, M.; Solimando, A.G.; Buonavoglia, A.; Silvestris, N.; Leone, P.; Racanelli, V. Impact of antigen presentation mechanisms on immune response in autoimmune hepatitis. Front. Immunol. 2021, 12, 814155. [Google Scholar] [CrossRef] [PubMed]
- Silvestris, N.; Argentiero, A.; Brunetti, O.; Sonnessa, M.; Colonna, F.; Delcuratolo, S.; Luchini, C.; Scarpa, A.; Lonardi, S.; Nappo, F.; et al. PD-L1 and Notch as novel biomarkers in pancreatic sarcomatoid carcinoma: A pilot study. Expert Opin. Ther. Targets 2021, 25, 1007–1016. [Google Scholar] [CrossRef]
- Solimando, A.G.; Summa, S.; Vacca, A.; Ribatti, D. Cancer-associated angiogenesis: The endothelial cell as a checkpoint for immunological patrolling. Cancers 2020, 12, 3380. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Brunetti, O.; Gnoni, A.; Licchetta, A.; Delcuratolo, S.; Memeo, R.; Solimando, A.G.; Argentiero, A. Emerging role of immune checkpoint inhibitors in hepatocellular carcinoma. Medicina 2019, 55, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Fu, M.; Wang, M.; Wei, Y.; Wei, X. Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol. Cancer 2022, 21, 104. [Google Scholar] [CrossRef]
- Aghdasi, B.; Ye, K.; Resnick, A.; Huang, A.; Ha, H.C.; Guo, X.; Dawson, T.M.; Dawson, V.L.; Snyder, S.H. FKBP12, the 12-kDa FK506-binding protein, is a physiologic regulator of the cell cycle. Proc. Natl. Acad. Sci. USA 2001, 98, 2425–2430. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Donahoe, P.K. The immunophilin FKBP12: A molecular guardian of the TGF-beta family type I receptors. Front. Biosci. 2004, 9, 619–631. [Google Scholar] [CrossRef]
- Chen, J.; Zaidi, S.; Rao, S.; Chen, J.S.; Phan, L.; Farci, P.; Su, X.; Shetty, K.; White, J.; Zamboni, F.; et al. Analysis of genomes and transcriptomes of hepatocellular carcinomas identifies mutations and gene expression changes in the transforming growth Factor-β pathway. Gastroenterology 2018, 154, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gingold, J.A.; Su, X. Immunomodulatory TGF-β signaling in hepatocellular carcinoma. Trends Mol. Med. 2019, 25, 1010–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llovet, J.M.; Montal, R.; Sia, D.; Finn, R.S. Molecular therapies and precision medicine for hepatocellular carcinoma. Nat. Rev. Clin. Oncol. 2018, 15, 599–616. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A web server for comprehensive analysis of tumor-Infiltrating immune cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Severson, E.; Pignon, J.C.; Zhao, H.; Li, T.; Novak, J.; Jiang, P.; Shen, H.; Aster, J.C.; Rodig, S.; et al. Comprehensive analyses of tumor immunity: Implications for cancer immunotherapy. Genome Biol. 2016, 17, 174. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Lian, Q.; Wang, S.; Zhang, G.; Wang, D.; Luo, G.; Tang, J.; Chen, L.; Gu, J. HCCDB: A database of hepatocellular carcinoma expression atlas. Genom. Proteom. Bioinform. 2018, 16, 269–275. [Google Scholar] [CrossRef]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef]
- Menyhárt, O.; Nagy, Á.; Győrffy, B. Determining consistent prognostic biomarkers of overall survival and vascular invasion in hepatocellular carcinoma. R. Soc. Open Sci. 2018, 5, 181006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Han, Y.; Cai, C.; Liu, P.; Chen, Y.; Shen, H.; Xu, X.; Zeng, S. CLEC10A is a prognostic biomarker and correlated with clinical pathologic features and immune infiltrates in lung adenocarcinoma. J. Cell. Mol. Med. 2021, 25, 3391–3399. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Factors | Overall Survival (n = 364) | Progression-Free Survival (n = 366) | |||||
|---|---|---|---|---|---|---|---|
| N | Hazard Ratio | p Value | N | Hazard Ratio | p Value | ||
| Sex | Female | 118 | 1.82 (1.04–3.18) | 0.033 | 121 | 1.87 (1.08–3.22) | 0.022 |
| Male | 246 | 2.55 (1.5–4.32) | 0.00034 | 249 | 2.2 (1.4–3.45) | 0.00048 | |
| Stage | 1 | 170 | 2.54 (1.38–4.66) | 0.0019 | 171 | 2.03 (1.13–3.65) | 0.016 |
| 2 | 83 | 0.62 (0.25–1.54) | 0.2978 | 85 | 1.88 (0.94–3.73) | 0.0678 | |
| 3 | 83 | 3.52 (1.55–7.97) | 0.0014 | 85 | 1.98 (1.14–3.44) | 0.0142 | |
| 4 | 4 | - | - | 5 | - | - | |
| Grade | 1 | 55 | 3.8 (1.41–10.25) | 0.0048 | 55 | 2.69 (1.23–5.88) | 0.0099 |
| 2 | 174 | 1.79 (1.07–2.99) | 0.0251 | 177 | 1.84 (1.14–2.95) | 0.0107 | |
| 3 | 118 | 2.18 (1.18–4.02) | 0.0106 | 121 | 1.55 (0.84–2.85) | 0.1612 | |
| 4 | 12 | - | - | 12 | - | - | |
| AJCC-T | 1 | 180 | 2.34 (1.31–4.19) | 0.0032 | 181 | 1.94 (1.11–3.38) | 0.0175 |
| 2 | 90 | 0.61 (0.27–1.37) | 0.2278 | 93 | 1.93 (0.98–3.78) | 0.0519 | |
| 3 | 78 | 2.79 (1.45–5.37) | 0.0014 | 80 | 2.2 (1.12–4.29) | 0.0187 | |
| 4 | 13 | - | - | 13 | - | - | |
| Vascular invasion | None | 203 | 1.88 (1.1–3.19) | 0.0182 | 205 | 1.67 (1.04–2.67) | 0.0322 |
| Micro | 90 | 2.27 (0.78–6.62) | 0.1211 | 92 | 2.5 (1.24–5.04) | 0.0079 | |
| Race | White | 181 | 1.76 (1.1–2.8) | 0.0163 | 184 | 2.22 (1.37–3.59) | 0.0008 |
| Asian | 155 | 4.12 (1.74–9.75) | 0.0005 | 157 | 2.72 (1.45–5.08) | 0.0012 | |
| Alcohol consumption | Yes | 115 | 2.31 (1.13–4.74) | 0.0191 | 116 | 1.55 (0.91–2.67) | 0.1059 |
| None | 202 | 2.09 (1.31–3.32) | 0.0016 | 205 | 2.83 (1.65–4.87) | 8.8 × 10−5 | |
| Viral hepatitis | Yes | 160 | 2.27 (1.07–4.82) | 0.0276 | 153 | 1.63 (0.93–2.84) | 0.086 |
| None | 167 | 1.92 (1.19–3.08) | 0.0063 | 169 | 2.56 (1.51–4.34) | 0.0003 | |
| Factor | Univariable Analysis | Multivariable Analysis | ||
|---|---|---|---|---|
| Hazard Ratio (95% CI) | p-Value | Hazard Ratio (95% CI) | p-Value | |
| FKBP1A | 1.766 (1.375–2.268) | <0.001 | 1.546 (1.185–2.017) | 0.001 |
| Age | 1.012 (0.999–1.026) | 0.078 | 1.008 (0.994–1.022) | 0.261 |
| Gender | 0.816 (0.573–1.163) | 0.260 | 0.907 (0.623–1.320) | 0.611 |
| pT stage | 1.675 (1.397–2.007) | <0.001 | 1.804 (1.411–2.306) | <0.001 |
| pTNM stage | 1.376 (1.145–1.654) | <0.001 | 0.870 (0.687–1.101) | 0.246 |
| Grade | 1.121 (0.887–1.417) | 0.339 | 1.079 (0.836–1.392) | 0.559 |
| #Term ID | Term Description | Count in Network | Strength | False Discovery Rate |
|---|---|---|---|---|
| hsa04659 | Th17 cell differentiation | 5/101 | 1.94 | 6.40 × 10−7 |
| hsa04218 | Cellular senescence | 5/150 | 1.77 | 2.11 × 10−6 |
| hsa04921 | Oxytocin signaling pathway | 5/149 | 1.78 | 2.11 × 10−6 |
| hsa04020 | Calcium signaling pathway | 5/193 | 1.66 | 3.72 × 10−6 |
| hsa05235 | PD-L1 expression and PD-1 checkpoint pathway in cancer | 4/88 | 1.91 | 9.88 × 10−6 |
| hsa05020 | Prion disease | 5/265 | 1.53 | 1.16 × 10−5 |
| hsa04380 | Osteoclast differentiation | 4/122 | 1.77 | 2.50 × 10−5 |
| hsa04371 | Apelin signaling pathway | 4/131 | 1.73 | 2.89 × 10−5 |
| hsa05167 | Kaposi sarcoma-associated herpesvirus infection | 4/187 | 1.58 | 0.0001 |
| hsa04370 | VEGF signaling pathway | 3/57 | 1.97 | 0.00013 |
| hsa05163 | Human cytomegalovirus infection | 4/218 | 1.51 | 0.00013 |
| hsa05166 | Human T-cell leukemia virus 1 infection | 4/211 | 1.53 | 0.00013 |
| hsa05170 | Human immunodeficiency virus 1 infection | 4/204 | 1.54 | 0.00013 |
| hsa04720 | Long-term potentiation | 3/64 | 1.92 | 0.00015 |
| hsa04924 | Renin secretion | 3/66 | 1.91 | 0.00015 |
| hsa05031 | Amphetamine addiction | 3/66 | 1.91 | 0.00015 |
| hsa04662 | B cell receptor signaling pathway | 3/78 | 1.84 | 0.00022 |
| hsa04010 | MAPK signaling pathway | 4/288 | 1.39 | 0.00028 |
| hsa04658 | Th1 and Th2 cell differentiation | 3/87 | 1.79 | 0.00028 |
| hsa04350 | TGF-beta signaling pathway | 3/91 | 1.77 | 0.00029 |
| hsa04625 | C-type lectin receptor signaling pathway | 3/102 | 1.72 | 0.00037 |
| hsa04660 | T-cell receptor signaling pathway | 3/101 | 1.72 | 0.00037 |
| hsa04922 | Glucagon signaling pathway | 3/101 | 1.72 | 0.00037 |
| hsa04724 | Glutamatergic synapse | 3/111 | 1.68 | 0.00043 |
| hsa05010 | Alzheimer disease | 4/355 | 1.3 | 0.00043 |
| hsa05014 | Amyotrophic lateral sclerosis | 4/352 | 1.31 | 0.00043 |
| hsa04114 | Oocyte meiosis | 3/120 | 1.65 | 0.00048 |
| hsa04650 | Natural killer cell mediated cytotoxicity | 3/121 | 1.64 | 0.00048 |
| hsa04310 | Wnt signaling pathway | 3/154 | 1.54 | 0.00092 |
| hsa04022 | cGMP-PKG signaling pathway | 3/162 | 1.52 | 0.001 |
| hsa05152 | Tuberculosis | 3/168 | 1.5 | 0.0011 |
| hsa04136 | Autophagy—other | 2/29 | 2.09 | 0.0013 |
| hsa04360 | Axon guidance | 3/177 | 1.48 | 0.0013 |
| hsa04213 | Longevity regulating pathway—multiple species | 2/61 | 1.77 | 0.0054 |
| hsa05212 | Pancreatic cancer | 2/73 | 1.69 | 0.0075 |
| hsa05210 | Colorectal cancer | 2/82 | 1.64 | 0.0091 |
| hsa04211 | Longevity regulating pathway | 2/87 | 1.61 | 0.0099 |
| hsa04713 | Circadian entrainment | 2/92 | 1.59 | 0.0108 |
| hsa04152 | AMPK signaling pathway | 2/120 | 1.47 | 0.0176 |
| hsa04728 | Dopaminergic synapse | 2/128 | 1.44 | 0.0194 |
| hsa04140 | Autophagy—animal | 2/130 | 1.44 | 0.0195 |
| hsa04910 | Insulin signaling pathway | 2/133 | 1.43 | 0.0199 |
| hsa05017 | Spinocerebellar ataxia | 2/135 | 1.42 | 0.02 |
| hsa05226 | Gastric cancer | 2/144 | 1.39 | 0.0222 |
| hsa04150 | mTOR signaling pathway | 2/151 | 1.37 | 0.0238 |
| hsa04390 | Hippo signaling pathway | 2/153 | 1.37 | 0.0239 |
| hsa05206 | MicroRNAs in cancer | 2/160 | 1.35 | 0.0255 |
| hsa05225 | Hepatocellular carcinoma | 2/160 | 1.35 | 0.0255 |
| hsa05131 | Shigellosis | 2/218 | 1.21 | 0.0443 |
| hsa04714 | Thermogenesis | 2/229 | 1.19 | 0.0478 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Cui, Y.; Duan, Q.; Zhang, J.; Shao, D.; Cao, X.; Gao, Y.; Wang, S.; Li, J.; Jones, O.D.; et al. The Prognostic Significance of FKBP1A and Its Related Immune Infiltration in Liver Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 12797. https://doi.org/10.3390/ijms232112797
Li Z, Cui Y, Duan Q, Zhang J, Shao D, Cao X, Gao Y, Wang S, Li J, Jones OD, et al. The Prognostic Significance of FKBP1A and Its Related Immune Infiltration in Liver Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2022; 23(21):12797. https://doi.org/10.3390/ijms232112797
Chicago/Turabian StyleLi, Zhongguang, Ying Cui, Qinchun Duan, Jianfei Zhang, Danyang Shao, Xixi Cao, Yuru Gao, Shulin Wang, Jiali Li, Odell D. Jones, and et al. 2022. "The Prognostic Significance of FKBP1A and Its Related Immune Infiltration in Liver Hepatocellular Carcinoma" International Journal of Molecular Sciences 23, no. 21: 12797. https://doi.org/10.3390/ijms232112797