
Correlation between Sperm Micro Ribonucleic Acid-34b and -34c Levels and Clinical Outcomes of Intracytoplasmic Sperm Injection in Men with Male Factor Infertility

Abstract
:1. Introduction
2. Results
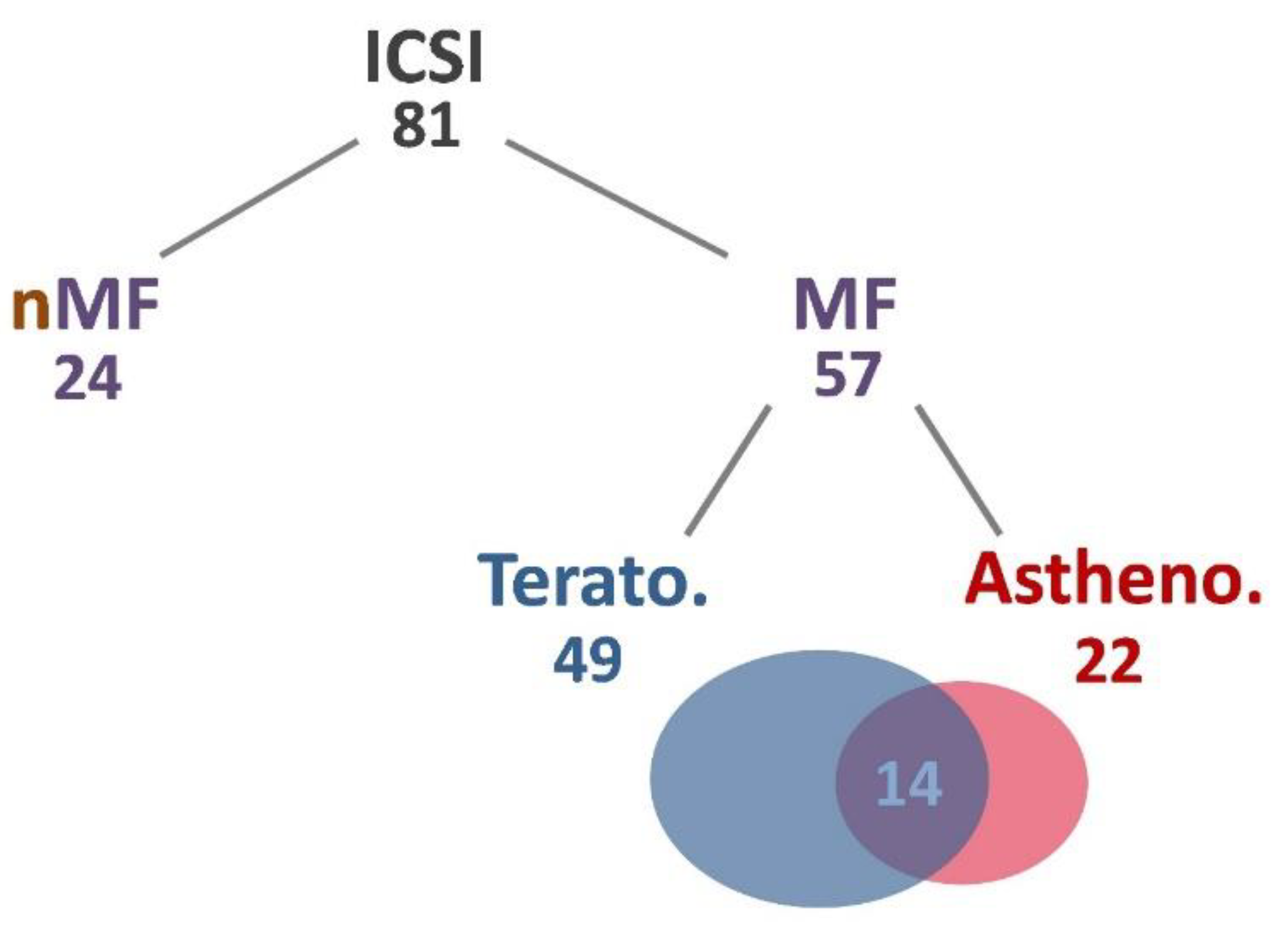
2.1. Physiological Data Analysis of Subjects
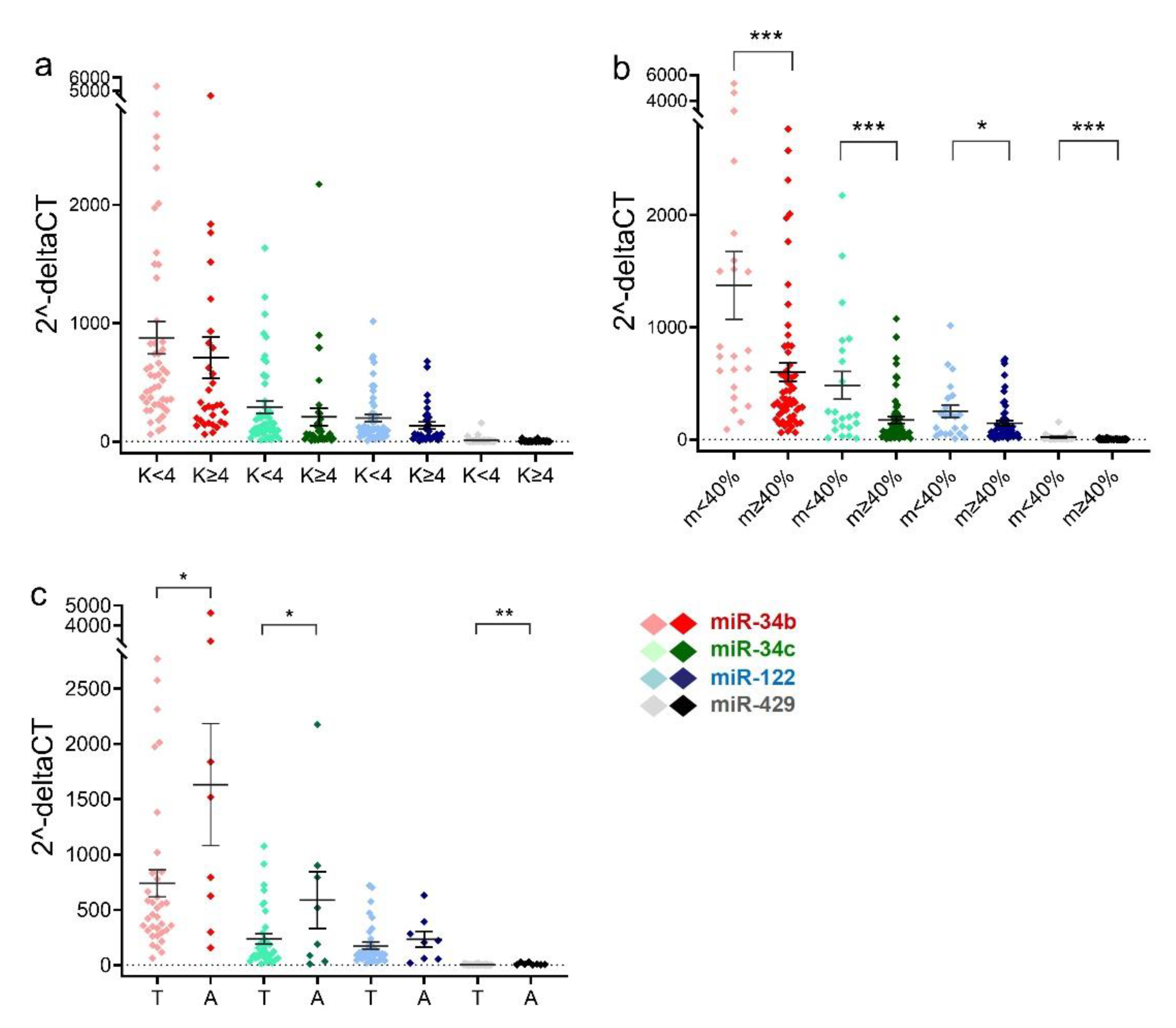
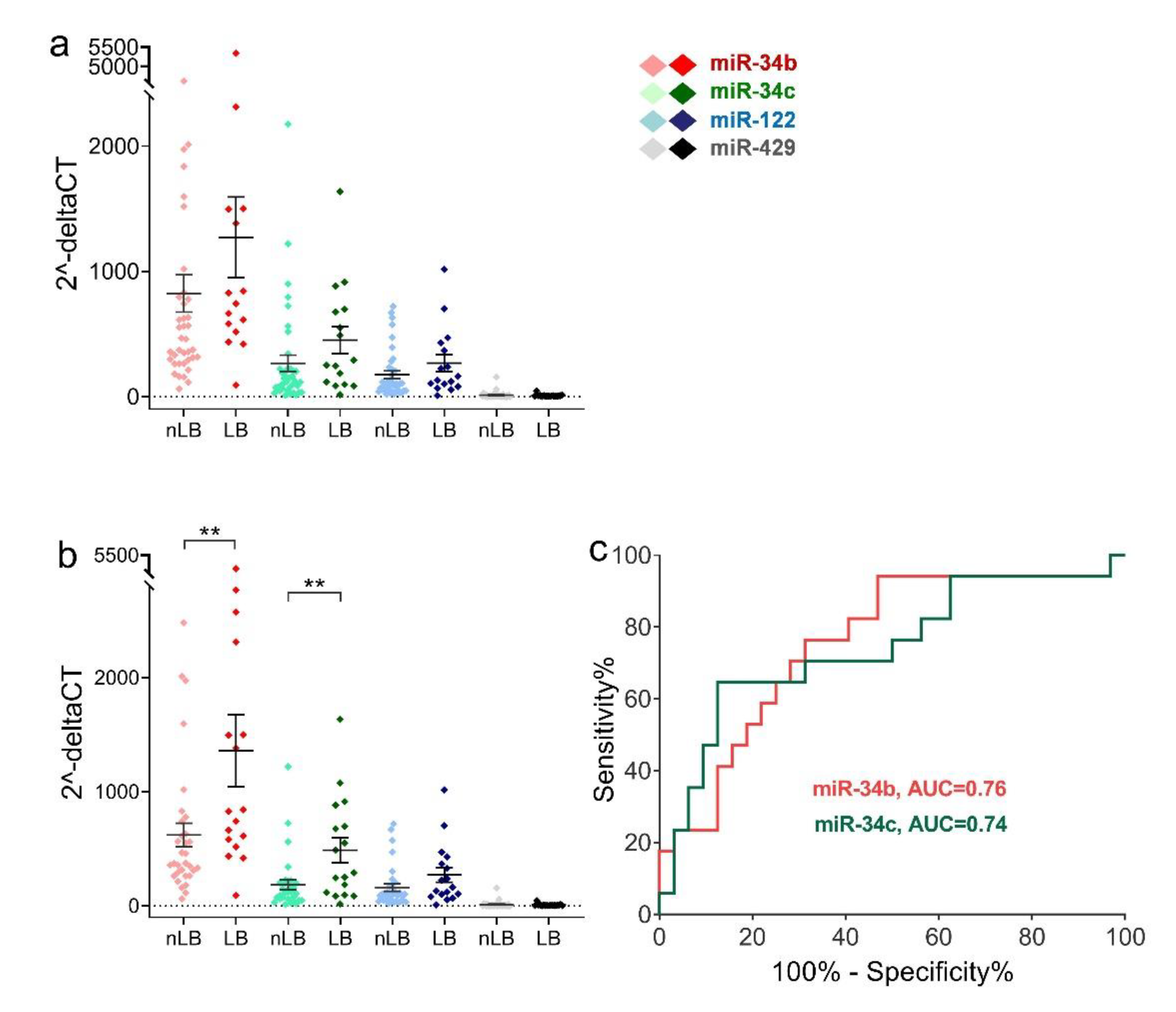
2.2. Sperm miRNA Levels in Men with Teratozoospermia or Asthenozoospermia
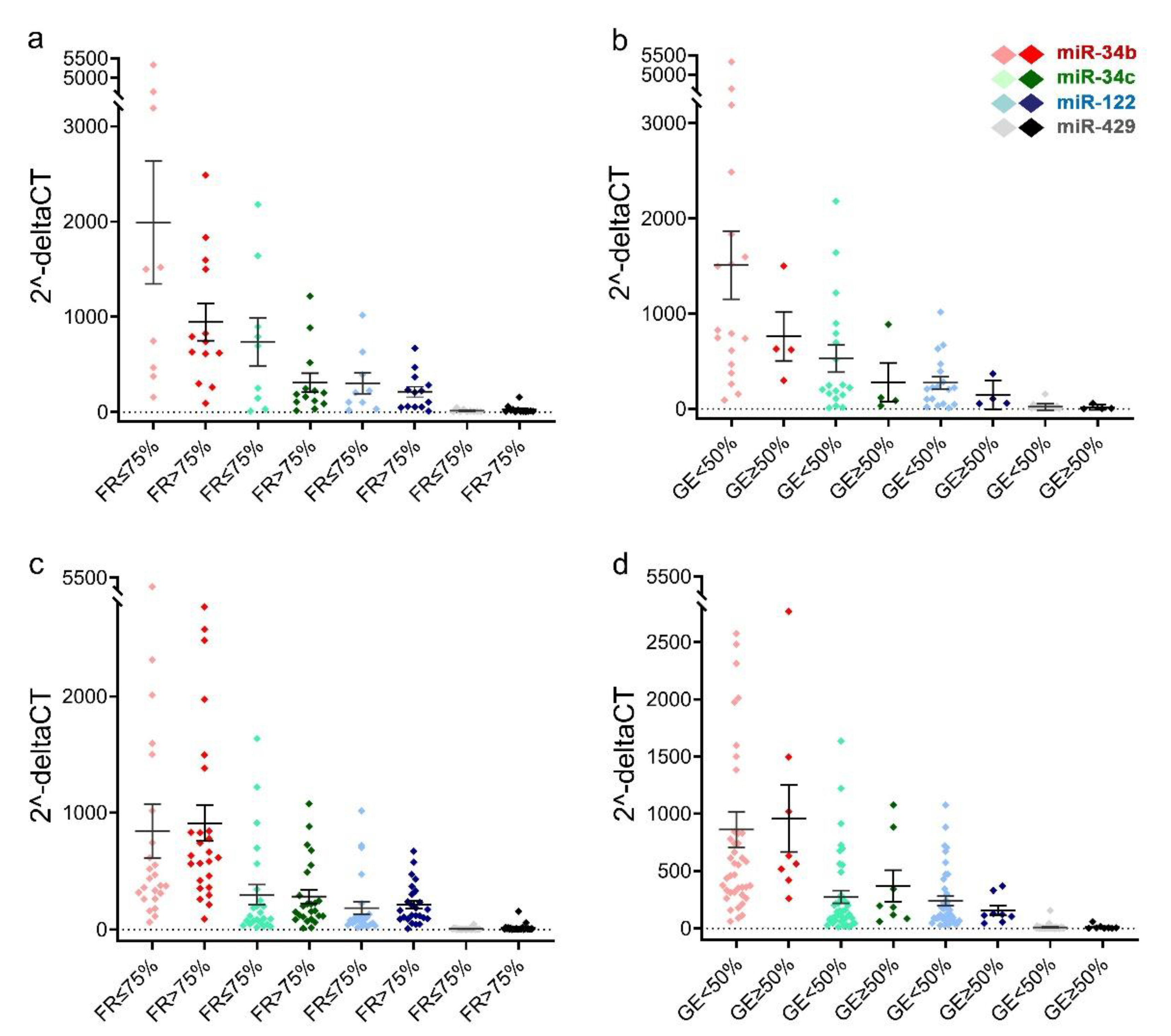
2.3. Comparison of Sperm miRNAs Levels in Different Groups of Fertilization Rate and Early Embryonic Development
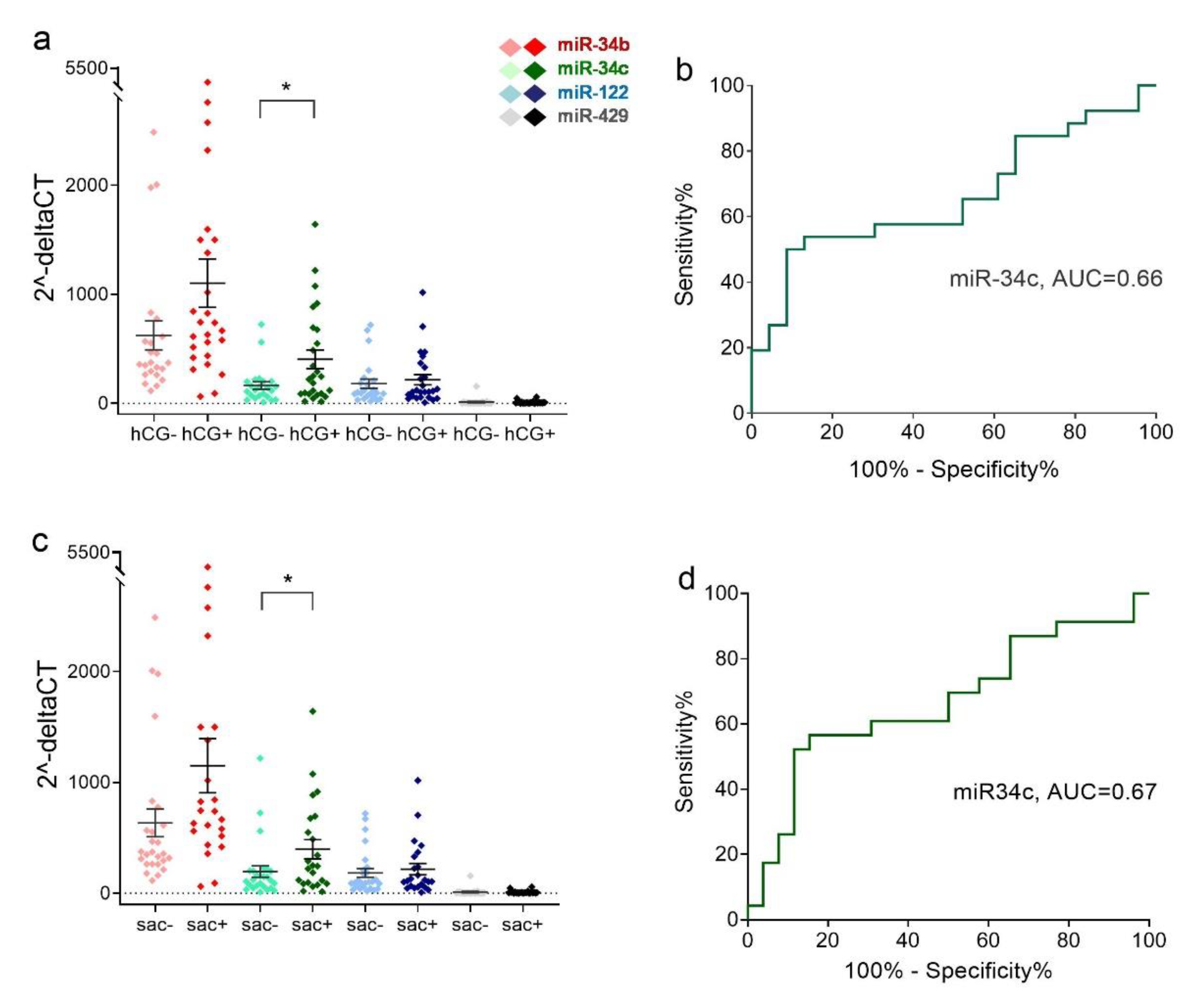
2.4. Relative Expression Levels of Sperm miRNAs in Different Clinical Outcome Groups after ICSI
3. Discussion
4. Materials and Methods
4.1. Sperm Sample Collection and Processing
4.2. Assisted Reproductive Technology
4.3. Fertilization and Determination of Clinical Outcomes
4.4. TaqMan MicroRNA Quantitative Polymerase Chain Reaction
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteller, M. Non-Coding RNAs in Human Disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Dadoune, J.-P. Spermatozoal RNAs: What about Their Functions? Microsc. Res. Tech. 2009, 72, 536–551. [Google Scholar] [CrossRef] [PubMed]
- McIver, S.C.; Roman, S.D.; Nixon, B.; McLaughlin, E.A. MiRNA and Mammalian Male Germ Cells. Hum. Reprod. Update 2012, 18, 44–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Halima, M.; Backes, C.; Leidinger, P.; Keller, A.; Lubbad, A.M.; Hammadeh, M.; Meese, E. MicroRNA Expression Profiles in Human Testicular Tissues of Infertile Men with Different Histopathologic Patterns. Fertil. Steril. 2014, 101, 78–86.e2. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Hammadeh, M.; Backes, C.; Fischer, U.; Leidinger, P.; Lubbad, A.M.; Keller, A.; Meese, E. Panel of Five MicroRNAs as Potential Biomarkers for the Diagnosis and Assessment of Male Infertility. Fertil. Steril. 2014, 102, 989–997.e1. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, C.; Chen, X.; Yao, B.; Yang, C.; Zhu, C.; Li, L.; Wang, J.; Li, X.; Shao, Y.; et al. Altered Profile of Seminal Plasma MicroRNAs in the Molecular Diagnosis of Male Infertility. Clin. Chem. 2011, 57, 1722–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannarella, R.; Barbagallo, F.; Crafa, A.; la Vignera, S.; Condorelli, R.A.; Calogero, A.E. Seminal Plasma Transcriptome and Proteome: Towards a Molecular Approach in the Diagnosis of Idiopathic Male Infertility. Int. J. Mol. Sci. 2020, 21, 7308. [Google Scholar] [CrossRef] [PubMed]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic Stem Cell-Specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Corney, D.C.; Flesken-Nikitin, A.; Godwin, A.K.; Wang, W.; Nikitin, A.Y. MicroRNA-34b and MicroRNA-34c Are Targets of P53 and Cooperate in Control of Cell Proliferation and Adhesion-Independent Growth. Cancer Res. 2007, 67, 8433–8438. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, K.; Spillare, E.A.; Fujita, K.; Horikawa, I.; Yamashita, T.; Appella, E.; Nagashima, M.; Takenoshita, S.; Yokota, J.; Harris, C.C. Nutlin-3a Activates P53 to Both down-Regulate Inhibitor of Growth 2 and up-Regulate Mir-34a, Mir-34b, and Mir-34c Expression, and Induce Senescence. Cancer Res. 2008, 68, 3193–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, M.; Munding, J.; Grüner, M.; Liffers, S.-T.; Verdoodt, B.; Hauk, J.; Steinstraesser, L.; Tannapfel, A.; Hermeking, H. Frequent Concomitant Inactivation of MiR-34a and MiR-34b/c by CpG Methylation in Colorectal, Pancreatic, Mammary, Ovarian, Urothelial, and Renal Cell Carcinomas and Soft Tissue Sarcomas. Virchows Arch. 2011, 458, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Li, D.; Wang, L.; Wu, J.; Hu, Y.; Wang, Z.; Chen, Y.; Cao, X.; Jiang, C.; Yan, W.; et al. MicroRNA-449 and MicroRNA-34b/c Function Redundantly in Murine Testes by Targeting E2F Transcription Factor-Retinoblastoma Protein (E2F-PRb) Pathway. J. Biol. Chem. 2012, 287, 21686–21698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, K.K.; Zhong, Y.; Liu, Y.-T.; Yamada, T.; Akatsuka, S.; Hu, Q.; Yoshihara, M.; Ohara, H.; Takehashi, M.; Shinohara, T.; et al. Association of MicroRNA-34a Overexpression with Proliferation Is Cell Type-Dependent. Cancer Sci. 2007, 98, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, S.; Novin, M.G.; Alizadeh, E.; Shahnazi, V.; Pashaei-Asl, F.; AsrBadr, Y.A.; Farzadi, L.; Ebrahimie, E.; Pashaiasl, M. New Insights into the Expression Profile of MicroRNA-34c and P53 in Infertile Men Spermatozoa and Testicular Tissue. Cell Mol. Biol. 2017, 63, 77–83. [Google Scholar] [CrossRef]
- Yan, N.; Lu, Y.; Sun, H.; Qiu, W.; Tao, D.; Liu, Y.; Chen, H.; Yang, Y.; Zhang, S.; Li, X.; et al. Microarray Profiling of MicroRNAs Expressed in Testis Tissues of Developing Primates. J. Assist. Reprod. Genet. 2009, 26, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Ye, L.; Liu, G.; Shao, G.; Zheng, R.; Ren, Z.; Zuo, B.; Xu, D.; Lei, M.; Jiang, S.; et al. Microarray-Based Approach Identifies Differentially Expressed MicroRNAs in Porcine Sexually Immature and Mature Testes. PLoS ONE 2010, 5, e11744. [Google Scholar] [CrossRef] [PubMed]
- Bouhallier, F.; Allioli, N.; Lavial, F.; Chalmel, F.; Perrard, M.-H.; Durand, P.; Samarut, J.; Pain, B.; Rouault, J.-P. Role of MiR-34c MicroRNA in the Late Steps of Spermatogenesis. RNA 2010, 16, 720–731. [Google Scholar] [CrossRef] [Green Version]
- Comazzetto, S.; di Giacomo, M.; Rasmussen, K.D.; Much, C.; Azzi, C.; Perlas, E.; Morgan, M.; O’Carroll, D. Oligoasthenoteratozoospermia and Infertility in Mice Deficient for MiR-34b/c and MiR-449 Loci. PLoS Genet. 2014, 10, e1004597. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-M.; Pang, R.T.K.; Chiu, P.C.N.; Wong, B.P.C.; Lao, K.; Lee, K.-F.; Yeung, W.S.B. Sperm-Borne MicroRNA-34c Is Required for the First Cleavage Division in Mouse. Proc. Natl. Acad. Sci. USA 2012, 109, 490–494. [Google Scholar] [CrossRef]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa Micro Ribonucleic Acid-34c Level Is Correlated with Intracytoplasmic Sperm Injection Outcomes. Fertil. Steril. 2015, 104, 312–317.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Shi, Q.; Sun, Y. The Effect of Sperm MiR-34c on Human Embryonic Development Kinetics and Clinical Outcomes. Life Sci. 2020, 256, 117895. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Raabe, T.; Hecht, N.B. MicroRNA Mirn122a Reduces Expression of the Posttranscriptionally Regulated Germ Cell Transition Protein 2 (Tnp2) Messenger RNA (MRNA) by MRNA Cleavage. Biol. Reprod. 2005, 73, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Huang, Y.; Liu, J.; Zhao, Y.; Jiang, L.; Huang, Q.; Cheng, W.; Guo, L. MicroRNA-122 Influences the Development of Sperm Abnormalities from Human Induced Pluripotent Stem Cells by Regulating TNP2 Expression. Stem Cells Dev. 2013, 22, 1839–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Cheng, W.; Gao, Y.; Wang, H.; Liu, Z. Microarray Analysis of MicroRNA Expression Patterns in the Semen of Infertile Men with Semen Abnormalities. Mol. Med. Rep. 2012, 6, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, S.; Pashaiasl, M.; Ezzati, M.; Ahmadi AsrBadr, Y.; Mohammadi-Dehcheshmeh, M.; Mohammadi, S.A.; Ghaffari Novin, M. MicroRNA-Based Regulatory Circuit Involved in Sperm Infertility. Andrologia 2020, 52, e13453. [Google Scholar] [CrossRef]
- Mokánszki, A.; Molnár, Z.; Tóthné, E.V.; Bodnár, B.; Jakab, A.; Bálint, B.L.; Balogh, I. Altered MicroRNAs Expression Levels of Sperm and Seminal Plasma in Patients with Infertile Ejaculates Compared with Normozoospermic Males. Hum. Fertil. 2020, 23, 246–255. [Google Scholar] [CrossRef]
- Zhu, M.; Fei, L.; Li, D.; Chen, D. Correlation Analysis of MiR-122-5p and Occludin with Sperm Density in Oligospermia Patients’ Sperm. Clin. Lab. 2019, 65, 156. [Google Scholar] [CrossRef]
- Capra, E.; Turri, F.; Lazzari, B.; Cremonesi, P.; Gliozzi, T.M.; Fojadelli, I.; Stella, A.; Pizzi, F. Small RNA Sequencing of Cryopreserved Semen from Single Bull Revealed Altered MiRNAs and PiRNAs Expression between High- and Low-Motile Sperm Populations. BMC Genom. 2017, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-Q.; He, L.-X.; Liu, S.-X. Expression of miR-429 and Its Target Gene HSPA4L in Sperms from Asthenospermia Patients. Sichuan Da Xue Xue Bao Yi Xue Ban 2016, 47, 869–873. [Google Scholar]
- Wu, W.; Qin, Y.; Li, Z.; Dong, J.; Dai, J.; Lu, C.; Guo, X.; Zhao, Y.; Zhu, Y.; Zhang, W.; et al. Genome-Wide MicroRNA Expression Profiling in Idiopathic Non-Obstructive Azoospermia: Significant up-Regulation of MiR-141, MiR-429 and MiR-7-1-3p. Hum. Reprod. 2013, 28, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Corral-Vazquez, C.; Salas-Huetos, A.; Blanco, J.; Vidal, F.; Sarrate, Z.; Anton, E. Sperm MicroRNA Pairs: New Perspectives in the Search for Male Fertility Biomarkers. Fertil. Steril. 2019, 112, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; Blanco, J.; Vidal, F.; Godo, A.; Grossmann, M.; Pons, M.C.; F-Fernández, S.; Garrido, N.; Anton, E. Spermatozoa from Patients with Seminal Alterations Exhibit a Differential Micro-Ribonucleic Acid Profile. Fertil. Steril. 2015, 104, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tscherner, A.; Gilchrist, G.; Smith, N.; Blondin, P.; Gillis, D.; Lamarre, J. MicroRNA-34 Family Expression in Bovine Gametes and Preimplantation Embryos. Reprod. Biol. Endocrinol. 2014, 12, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, M.; Yue, D.; Zhang, Y.; Xing, X.; Zhang, L.; Su, J.; Zheng, Y.; Zhang, Y. MicroRNA-34c Expression in Donor Cells Influences the Early Development of Somatic Cell Nuclear Transfer Bovine Embryos. Cell. Reprogram. 2014, 16, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conine, C.; Sun, F.; Song, L.; Rivera-Pérez, J.; Rando, O. Small RNAs Gained during Epididymal Transit of Sperm Are Essential for Embryonic Development in Mice. Dev. Cell 2018, 46, 470–480.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Tang, C.; Zhang, Y.; Wu, J.; Bao, J.; Zheng, H.; Xu, C.; Yan, W. Mir-34b/c and Mir-449a/b/c Are Required for Spermatogenesis, but Not for the First Cleavage Division in Mice. Biol. Open 2015, 4, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wang, X.; Wang, Z.; Li, J.; Xu, Z.; Miao, M.; Chen, G.; Lei, X.; Wu, J.; Shi, H.; et al. MicroRNA Expression Profile Analysis in Sperm Reveals Hsa-Mir-191 as an Auspicious Omen of in Vitro Fertilization. BMC Genom. 2020, 21, 165. [Google Scholar] [CrossRef] [Green Version]
- Fagerlind, M.; Stålhammar, H.; Olsson, B.; Klinga-Levan, K. Expression of MiRNAs in Bull Spermatozoa Correlates with Fertility Rates. Reprod. Domest. Anim. 2015, 50, 587–594. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Thomas, S.; Kenneth, D.; Livak, J. Analyzing Real-Time PCR Data by Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | All | nMF | MF | Terato. | Astheno. | p-Value |
|---|---|---|---|---|---|---|
| N | 81 | 24 | 57 | 49 | 22 | NS |
| Women’s age (y) | 36.94 ± 4 | 37.08 ± 4 | 36.88 ± 4 | 36.78 ± 4 | 37.14 ± 4 | NS |
| Men’s age (y) | 39.11 ± 6 | 38.38 ± 6 | 39.42 ± 5 | 39.31 ± 5 | 40.64 ± 5 | NS |
| Infertility year | 3.48 ± 3 | 3.42 ± 3 | 3.51 ± 3 | 3.61 ± 3 | 3.59 ± 2 | NS |
| Oocyte retrieved no. | 10.43 ± 6 | 9.96 ± 4 | 10.63 ± 6 | 11.00 ± 7 | 10.05 ± 5 | NS |
| Good embryo no. | 1.35 ± 2 | 1.04 ± 1 | 1.47 ± 6 | 1.43 ± 2 | 1.41 ± 2 | NS |
| Transferred embryo no. | 2.36 ± 1 | 2.50 ± 1 | 2.30 ± 1 | 2.31 ± 1 | 2.23 ± 1 | NS |
| E2 on hCG day (pmol/L) a | 1812 ± 991 | 1634 ± 863 | 1851 ± 1060 | 1875 ± 1113 | 1822 ± 938 | NS |
| P4 on hCG day (nmol/L) a | 0.59 ± 0.36 | 0.54 ± 0.23 | 0.61 ± 0.40 | 0.62 ± 0.41 | 0.69 ± 0.54 | NS |
| LH on hCG day (IU/L) a | 2.94 ± 6 | 4.13 ± 9 | 2.33 ± 2 | 2.14 ± 1.96 | 3.18 ± 2 | NS |
| ICSI Outcomes | miR-34b(−) 1 | miR-34b(+) 2 | p-Value |
|---|---|---|---|
| Cycle (n = 49) | 18 | 31 | |
| Normal fertilization rate (per MII, %) | 74.01% | 82.42% | 0.0696 |
| More than 50% good-quality embryo rate (%) | 5.56% | 22.58% | 0.2293 |
| Implantation rate (per embryo, %) | 5.50% | 43.52% | 0.0003 |
| hCG+ rate (per embryo transfer, %) | 27.78% | 67.74% | 0.0090 |
| Sac+ rate (per embryo transfer, %) | 16.67% | 64.52% | 0.0025 |
| FHB+ rate (per embryo transfer, %) | 16.67% | 58.06% | 0.0069 |
| Live birth rate (per embryo transfer, %) | 5.56% | 51.61% | 0.0014 |
| Miscarriages rate (%) | 40.00% | 14.28% | 0.2357 |
| ICSI Outcomes | miR-34c (−) 1 | miR-34c(+) 2 | p-Value |
|---|---|---|---|
| Cycle (n = 49) | 34 | 15 | |
| Normal fertilization rate (per MII, %) | 78.24% | 81.87% | 0.4582 |
| More than 50% good-quality embryo rate (%) | 14.71% | 20.00% | 0.6869 |
| Implantation rate (per embryo, %) | 20.56% | 49.93% | 0.0092 |
| hCG+ rate (per embryo transfer, %) | 38.24% | 86.67% | 0.0021 |
| Sac+ rate (per embryo transfer, %) | 32.35% | 80.00% | 0.0043 |
| FHB+ rate (per embryo transfer, %) | 26.47% | 80.00% | 0.0012 |
| Live birth rate (per embryo transfer, %) | 17.65% | 73.33% | 0.0003 |
| Miscarriages rate (%) | 36.35% | 8.33% | 0.1550 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, L.-Y.; Lee, R.K.-K.; Lin, M.-H.; Huang, C.-H.; Li, S.-H. Correlation between Sperm Micro Ribonucleic Acid-34b and -34c Levels and Clinical Outcomes of Intracytoplasmic Sperm Injection in Men with Male Factor Infertility. Int. J. Mol. Sci. 2022, 23, 12381. https://doi.org/10.3390/ijms232012381
Yeh L-Y, Lee RK-K, Lin M-H, Huang C-H, Li S-H. Correlation between Sperm Micro Ribonucleic Acid-34b and -34c Levels and Clinical Outcomes of Intracytoplasmic Sperm Injection in Men with Male Factor Infertility. International Journal of Molecular Sciences. 2022; 23(20):12381. https://doi.org/10.3390/ijms232012381
Chicago/Turabian StyleYeh, Ling-Yu, Robert Kuo-Kuang Lee, Ming-Huei Lin, Chih-Hung Huang, and Sheng-Hsiang Li. 2022. "Correlation between Sperm Micro Ribonucleic Acid-34b and -34c Levels and Clinical Outcomes of Intracytoplasmic Sperm Injection in Men with Male Factor Infertility" International Journal of Molecular Sciences 23, no. 20: 12381. https://doi.org/10.3390/ijms232012381