
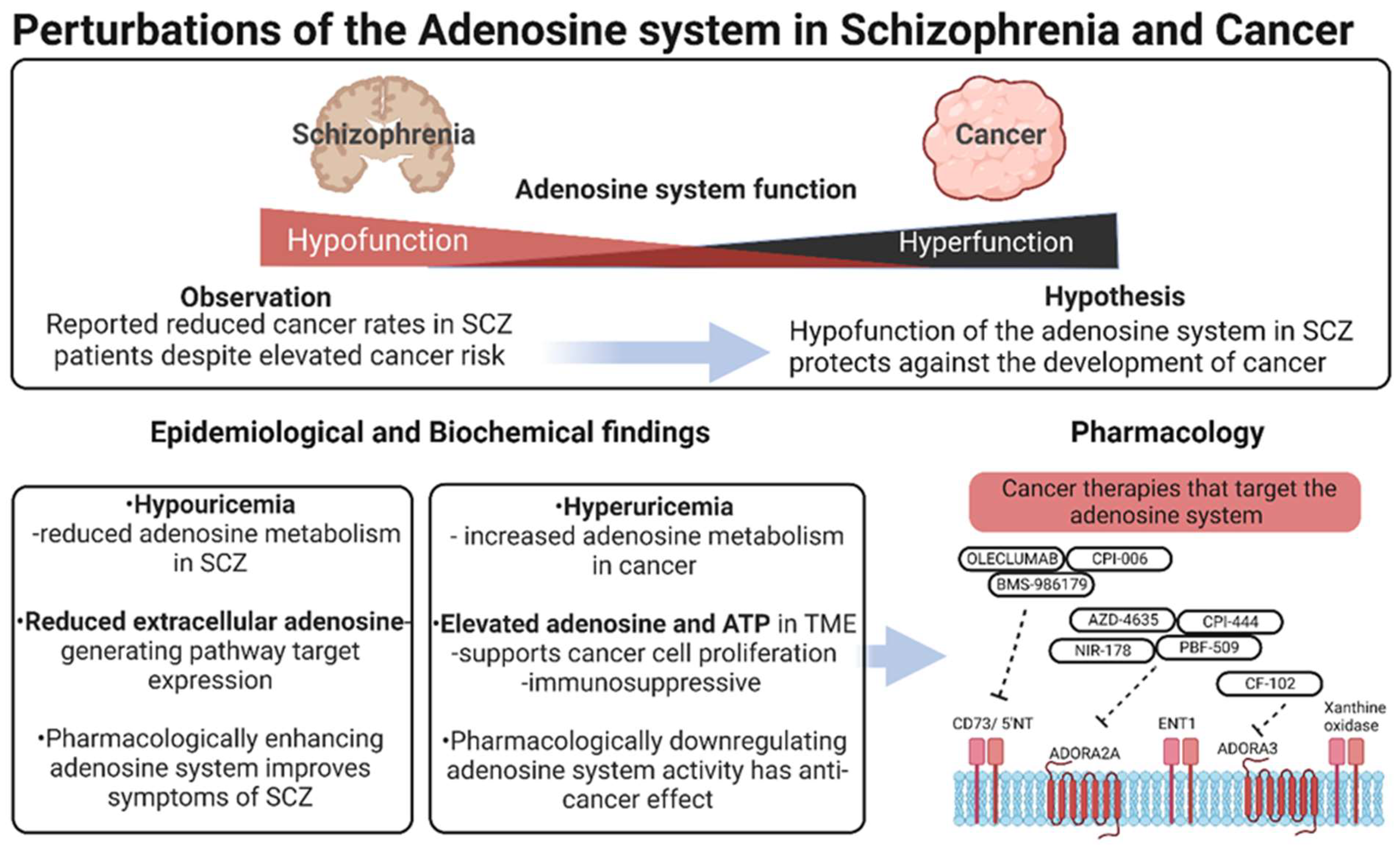
Adenosine, Schizophrenia and Cancer: Does the Purinergic System Offer a Pathway to Treatment?
 , ,
, ,
Abstract
:1. Introduction
2. Cancer Incidence in Schizophrenia
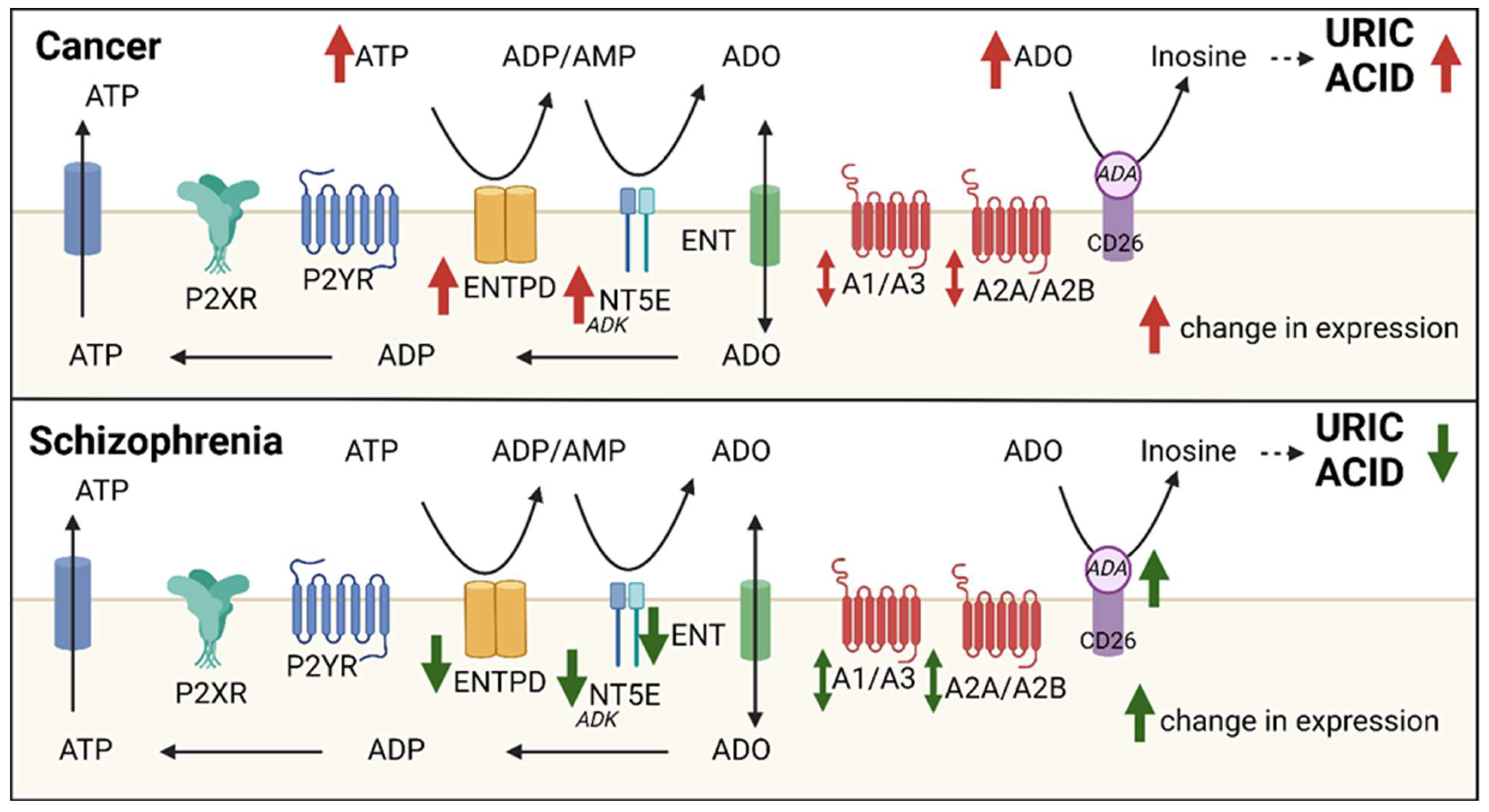
3. The Adenosine System: Extracellular Adenosine Generating Pathways
4. The Adenosine System: Purinergic Receptors
5. The Adenosine System: Perturbed Adenosine Metabolism in Disease: Uric Acid
6. Schizophrenia: Evidence for Adenosine System Perturbation
7. Schizophrenia: The Adenosine System as a Therapeutic Target
8. Cancer: Extracellular Adenosine Generation and Cancer
9. Cancer: Adenosine System as a Therapeutic Target in Cancer
10. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- HMSO; The Board of Control of the Commissioners in Lunacy. Sixty-Fourth Report of the Commissioners in Lunacy to the Lord Chancellor; His Majety’s Stationery Office: London, UK, 1910.
- Berchuck, J.E.; Meyer, C.S.; Zhang, N.; Berchuck, C.M.; Trivedi, N.N.; Cohen, B.; Wang, S. Association of Mental Health Treatment with Outcomes for US Veterans Diagnosed with Non–Small Cell Lung Cancer. JAMA Oncol. 2020, 6, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Bushe, C.J.; Hodgson, R. Schizophrenia and cancer: In 2010 do we understand the connection? Can. J. Psychiatry 2010, 55, 761–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catts, V.S.; Catts, S.V.; O’Toole, B.I.; Frost, A.D. Cancer incidence in patients with schizophrenia and their first-degree relatives—A meta-analysis. Acta Psychiatr. Scand. 2008, 117, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Yu, X.; Zheng, H.; Sun, X.; Lu, Y.; Zhang, Y.; Li, C.; Bi, X. The incidence rate of cancer in patients with schizophrenia: A meta-analysis of cohort studies. Schizophr. Res. 2018, 195, 519–528. [Google Scholar] [CrossRef]
- Mortensen, P.B. The occurrence of cancer in first admitted schizophrenic patients. Schizophr. Res. 1994, 12, 185–194. [Google Scholar] [CrossRef]
- Mortensen, P.B. The incidence of cancer in schizophrenic patients. J. Epidemiol. Community Health 1989, 43, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Goldacre, M.J.; Kurina, L.M.; Wotton, C.J.; Yeates, D.; Seagroat, V. Schizophrenia and cancer: An epidemiological study. Br. J. Psychiatry 2005, 187, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Lichtermann, D.; Ekelund, J.; Pukkala, E.; Tanskanen, A.; Lonnqvist, J. Incidence of cancer among persons with schizophrenia and their relatives. Arch. Gen. Psychiatry 2001, 58, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Chen, G.; Kong, L.; Zhang, W.; Hu, L.; Chen, C.; Li, J.; Zhuo, C. Lower risk of liver cancer in patients with schizophrenia: A systematic review and meta-analysis of cohort studies. Oncotarget 2017, 8, 102328–102335. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, C.; Zhuang, H.; Gao, X.; Triplett, P.T. Lung cancer incidence in patients with schizophrenia: Meta-analysis. Br. J. Psychiatry 2019, 215, 704–711. [Google Scholar] [CrossRef]
- Hippisley-Cox, J.; Vinogradova, Y.; Coupland, C.; Parker, C. Risk of Malignancy in Patients with Schizophrenia or Bipolar Disorder: Nested Case-Control Study. Arch. Gen. Psychiatry 2007, 64, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Osborn, D.P.; Limburg, H.; Walters, K.; Petersen, I.; King, M.; Green, J.; Watson, J.; Nazareth, I. Relative incidence of common cancers in people with severe mental illness. Cohort study in the United Kingdom THIN primary care database. Schizophr. Res. 2013, 143, 44–49. [Google Scholar] [CrossRef]
- Lasser, K.; Boyd, J.W.; Woolhandler, S.; Himmelstein, D.U.; McCormick, D.; Bor, D.H. Smoking and mental illness: A population-based prevalence study. J. Am. Med. Assoc. 2000, 284, 2606–2610. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Nakao, M.; Takeuchi, T.; Yano, E. Smoking rates among schizophrenia patients in Japan. Psychiatry Res. 2011, 186, 165–169. [Google Scholar] [CrossRef]
- Campo-Arias, A.; Diaz-Martinez, L.A.; Rueda-Jaimes, G.E.; Rueda-Sanchez, M.; Farelo-Palacin, D.; Diaz, F.J.; de Leon, J. Smoking is associated with schizophrenia, but not with mood disorders, within a population with low smoking rates: A matched case-control study in Bucaramanga, Colombia. Schizophr. Res. 2006, 83, 269–276. [Google Scholar] [CrossRef]
- Murphy, K.A.; Stone, E.M.; Presskreischer, R.; McGinty, E.E.; Daumit, G.L.; Pollack, C.E. Cancer Screening Among Adults with and without Serious Mental Illness: A Mixed Methods Study. Med. Care 2021, 59, 327–333. [Google Scholar] [CrossRef]
- Hodgson, R.; Wildgust, H.J.; Bushe, C.J. Cancer and schizophrenia: Is there a paradox? J. Psychopharmacol. 2010, 24 (Suppl. 4), 51–60. [Google Scholar] [CrossRef]
- Hjorthoj, C.; Sturup, A.E.; McGrath, J.J.; Nordentoft, M. Years of potential life lost and life expectancy in schizophrenia: A systematic review and meta-analysis. Lancet Psychiatry 2017, 4, 295–301. [Google Scholar] [CrossRef]
- Kisely, S.; Crowe, E.; Lawrence, D. Cancer-related mortality in people with mental illness. JAMA Psychiatry 2013, 70, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Ni, L.; Wu, J.; Long, Y.; Tao, J.; Xu, J.; Yuan, X.; Yu, N.; Wu, R.; Zhang, Y. Mortality of site-specific cancer in patients with schizophrenia: A systematic review and meta-analysis. BMC Psychiatry 2019, 19, 323. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Xiping, Z.; Shuai, Z.; Feijiang, Y.; Bo, C.; Shifeng, Y.; Qihui, C. Meta-analysis of the Correlation Between Schizophrenia and Breast Cancer. Clin. Breast Cancer 2019, 19, e172–e185. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Song, J.; Lu, Y.; Fall, K.; Chen, X.; Fang, F.; Landen, M.; Hultman, C.M.; Czene, K.; Sullivan, P.; et al. A shared genetic contribution to breast cancer and schizophrenia. Nat. Commun. 2020, 11, 4637. [Google Scholar] [CrossRef]
- Yuan, K.; Song, W.; Liu, Z.; Lin, G.N.; Yu, S. Mendelian Randomization and GWAS Meta Analysis Revealed the Risk-Increasing Effect of Schizophrenia on Cancers. Biology 2022, 11, 1345. [Google Scholar] [CrossRef]
- Wootten, J.C.; Wiener, J.C.; Blanchette, P.S.; Anderson, K.K. Cancer incidence and stage at diagnosis among people with psychotic disorders: Systematic review and meta-analysis. Cancer Epidemiol. 2022, 80, 102233. [Google Scholar] [CrossRef]
- Ge, F.; Huo, Z.; Liu, Y.; Du, X.; Wang, R.; Lin, W.; Wang, R.; Chen, J.; Lu, Y.; Wen, Y.; et al. Association between schizophrenia and prostate cancer risk: Results from a pool of cohort studies and Mendelian randomization analysis. Compr. Psychiatry 2022, 115, 152308. [Google Scholar] [CrossRef]
- Durand, N.; Simsir, M.; Signetti, L.; Labbal, F.; Ballotti, R.; Mus-Veteau, I. Methiothepin Increases Chemotherapy Efficacy against Resistant Melanoma Cells. Molecules 2021, 26, 1867. [Google Scholar] [CrossRef]
- Matteoni, S.; Matarrese, P.; Ascione, B.; Buccarelli, M.; Ricci-Vitiani, L.; Pallini, R.; Villani, V.; Pace, A.; Paggi, M.G.; Abbruzzese, C. Anticancer Properties of the Antipsychotic Drug Chlorpromazine and Its Synergism with Temozolomide in Restraining Human Glioblastoma Proliferation In Vitro. Front. Oncol. 2021, 11, 635472. [Google Scholar] [CrossRef]
- Weissenrieder, J.S.; Reed, J.L.; Moldovan, G.L.; Johnson, M.T.; Trebak, M.; Neighbors, J.D.; Mailman, R.B.; Hohl, R.J. Antipsychotic drugs elicit cytotoxicity in glioblastoma multiforme in a calcium-dependent, non-D2 receptor-dependent, manner. Pharmacol. Res. Perspect. 2021, 9, e00689. [Google Scholar] [CrossRef]
- Zhuo, C.; Xun, Z.; Hou, W.; Ji, F.; Lin, X.; Tian, H.; Zheng, W.; Chen, M.; Liu, C.; Wang, W.; et al. Surprising Anticancer Activities of Psychiatric Medications: Old Drugs Offer New Hope for Patients with Brain Cancer. Front. Pharmacol. 2019, 10, 1262. [Google Scholar] [CrossRef]
- Ji, J.; Sundquist, K.; Ning, Y.; Kendler, K.S.; Sundquist, J.; Chen, X. Incidence of cancer in patients with schizophrenia and their first-degree relatives: A population-based study in Sweden. Schizophr. Bull. 2013, 39, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Nollen, E.A.A.; Rohrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Chakroborty, D.; Basu, S. Neurotransmitters as regulators of tumor angiogenesis and immunity: The role of catecholamines. J. Neuroimmune Pharmacol. 2013, 8, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, D.R.; Souza, D.O. Schizophrenia: A purinergic hypothesis. Med. Hypotheses 2000, 54, 157–166. [Google Scholar] [CrossRef]
- Vigano, S.; Alatzoglou, D.; Irving, M.; Menetrier-Caux, C.; Caux, C.; Romero, P.; Coukos, G. Targeting Adenosine in Cancer Immunotherapy to Enhance T-Cell Function. Front. Immunol. 2019, 10, 925. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.Y.; Singer, P.; Lytle, N.; Wei, C.J.; Lan, J.Q.; Williams-Karnesky, R.L.; Chen, J.F.; Yee, B.K.; Boison, D. Adenosine augmentation ameliorates psychotic and cognitive endophenotypes of schizophrenia. J. Clin. Investig. 2012, 122, 2567–2577. [Google Scholar] [CrossRef] [Green Version]
- Dunwiddie, T.V.; Masino, S.A. The role and regulation of adenosine in the central nervous system. Annu. Rev. Neurosci. 2001, 24, 31–55. [Google Scholar] [CrossRef] [Green Version]
- Boison, D.; Singer, P.; Shen, H.Y.; Feldon, J.; Yee, B.K. Adenosine hypothesis of schizophrenia—Opportunities for pharmacotherapy. Neuropharmacology 2012, 62, 1527–1543. [Google Scholar] [CrossRef] [Green Version]
- Blay, J.; White, T.D.; Hoskin, D.W. The extracellular fluid of solid carcinomas contains immunosuppressive concentrations of adenosine. Cancer Res. 1997, 57, 2602–2605. [Google Scholar]
- Cekic, C.; Day, Y.J.; Sag, D.; Linden, J. Myeloid expression of adenosine A2A receptor suppresses T and NK cell responses in the solid tumor microenvironment. Cancer Res. 2014, 74, 7250–7259. [Google Scholar] [CrossRef] [Green Version]
- Gessi, S.; Bencivenni, S.; Battistello, E.; Vincenzi, F.; Colotta, V.; Catarzi, D.; Varano, F.; Merighi, S.; Borea, P.A.; Varani, K. Inhibition of A2A Adenosine Receptor Signaling in Cancer Cells Proliferation by the Novel Antagonist TP455. Front. Pharmacol. 2017, 8, 888. [Google Scholar] [CrossRef]
- Huang, Y.; Gu, Z.; Fan, Y.; Zhai, G.; Zhao, X.; Sun, Q.; Shi, Y.; Lin, G. Inhibition of the adenosinergic pathway: The indispensable part of oncological therapy in the future. Purinergic Signal. 2019, 15, 53–67. [Google Scholar] [CrossRef]
- Boswell-Casteel, R.C.; Hays, F.A. Equilibrative nucleoside transporters—A review. Nucleosides Nucleotides Nucleic Acids 2017, 36, 7–30. [Google Scholar] [CrossRef]
- Cristalli, G.; Costanzi, S.; Lambertucci, C.; Lupidi, G.; Vittori, S.; Volpini, R.; Camaioni, E. Adenosine deaminase: Functional implications and different classes of inhibitors. Med. Res. Rev. 2001, 21, 105–128. [Google Scholar] [CrossRef]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [Green Version]
- Li, A. Mechanisms of ATP release, the enabling step in purinergic dynamics. Cell. Physiol. Biochem. 2011, 28, 1135–1144. [Google Scholar] [CrossRef]
- Lohman, A.W. Mechanisms of ATP release and signaling in the blood vessel wall. Cardiovasc. Res. 2012, 95, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Lazarowski, E.R. Molecular mechanisms of purine and pyrimidine nucleotide release. Adv. Pharmacol. 2011, 61, 2211–2261. [Google Scholar]
- Chavan, V.; Willis, J.; Walker, S.K.; Clark, H.R.; Liu, X.; Fox, M.A.; Srivastava, S.; Mukherjee, K. Correction: Central Presynaptic Terminals Are Enriched in ATP but the Majority Lack Mitochondria. PLoS ONE 2017, 12, e0181140. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, D.; Shan, D.; Ayers-Ringler, J.; Oliveros, A.; Benitez, J.; Prieto, M.; McCullumsmith, R.; Choi, D.S. Purinergic signaling and energy homeostasis in psychiatric disorders. Curr. Mol. Med. 2015, 15, 275–295. [Google Scholar] [CrossRef] [Green Version]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sévigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell-associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, M.G.; Pesi, R.; Allegrini, S. On the physiological role of cytosolic 5′-nucleotidase II (cN-II): Pathological and therapeutical implications. Curr. Med. Chem. 2013, 34, 4285–4291. [Google Scholar] [CrossRef] [PubMed]
- Romio, M.; Reinbeck, B.; Bongardt, S.; Hüls, S.; Burghoff, S.; Schrader, J. Extracellular purine metabolism and signaling of CD73-derived adenosine in murine Treg and Teff cells. Am. J. Physiol. Cell Physiol. 2011, 301, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, S. The E-NTPDase family of ectonucleotidases: Structure function relationships and pathophysiological significance. Purinergic Signal. 2006, 2, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Cekic, G. Purinergic regulation of the immune system. Nature Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef]
- Friedman, D.J.; Kunzli, B.M.; Yi, A.R.; Sevigny, J.; Berberat, P.O.; Enjyoji, K.; Csizmadia, E.; Friess, H.; Robson, S.C. From the Cover: CD39 deletion exacerbates experimental murine colitis and human polymorphisms increase susceptibility to inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2009, 106, 16788–16793. [Google Scholar] [CrossRef] [Green Version]
- Louis, N.A.; Robinson, A.M.; MacManus, C.F.; Karhausen, J.; Scully, M.; Colgan, S.P. Control of IFN-alphaA by CD73: Implications for mucosal inflammation. J. Immunol. 2008, 180, 4246–4255. [Google Scholar] [CrossRef] [Green Version]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta—Mol. Cell Res. 2008, 1783, 673–694. [Google Scholar] [CrossRef] [Green Version]
- Kopp, R.; Krautloher, A.; Ramirez-Fernandez, A.; Nicke, A. P2 × 7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Samways, D.S.; Li, Z.; Egan, T.M. Principles and properties of ion flow in P2X receptors. Front. Cell. Neurosci. 2014, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Ormond, S.J.; Barrera, N.P.; Qureshi, O.S.; Henderson, R.M.; Edwardson, J.M.; Murrell-Lagnado, R.D. An uncharged region within the N terminus of the P2 × 6 receptor inhibits its assembly and exit from the endoplasmic reticulum. Mol. Pharmacol. 2006, 69, 1692–1700. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.H.; Walsh, M.C.; Yu, J.; Shen, H.; Wherry, E.J.; Choi, Y. Mice Lacking the Purinergic Receptor P2 × 5 Exhibit Defective Inflammasome Activation and Early Susceptibility to Listeria monocytogenes. J. Immunol. 2020, 205, 760–766. [Google Scholar] [CrossRef]
- Stokes, L.; Layhadi, J.A.; Bibic, L.; Dhuna, K.; Fountain, S.J. P2 × 4 Receptor Function in the Nervous System and Current Breakthroughs in Pharmacology. Front. Pharmacol. 2017, 8, 291. [Google Scholar] [CrossRef] [Green Version]
- Fabbretti, E. ATP P2 × 3 receptors and neuronal sensitization. Front. Cell. Neurosci. 2013, 7, 236. [Google Scholar] [CrossRef] [Green Version]
- Richler, E.; Shigetomi, E.; Khakh, B.S. Neuronal P2 × 2 receptors are mobile ATP sensors that explore the plasma membrane when activated. J. Neurosci. 2011, 31, 16716–16730. [Google Scholar] [CrossRef] [Green Version]
- Mahaut-Smith, M.P.; Jones, S.; Evans, R.J. The P2 × 1 receptor and platelet function. Purinergic Signal. 2011, 7, 341–356. [Google Scholar] [CrossRef] [Green Version]
- Erb, L.; Weisman, G.A. Coupling of P2Y receptors to G proteins and other signaling pathways. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Olah, M.E. Adenosine receptor subtypes: Characterization and therapeutic regulation. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 581–606. [Google Scholar] [CrossRef]
- Ralevic, V.; Burnstock, G. Receptors for purines and pyrimidines. Pharmacol. Rev. 1998, 50, 413–492. [Google Scholar]
- Klotz, K.N.; Hessling, J.; Hegler, J.; Owman, C.; Kull, B.; Fredholm, B.B.; Lohse, M.J. Comparative pharmacology of human adenosine receptor subtypes—Characterization of stably transfected receptors in CHO cells. Naunyn-Schmiedeberg Arch. Pharmacol. 1998, 357, 1–9. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Irenius, E.; Kull, B.; Schulte, G. Comparison of the potency of adenosine as an agonist at human adenosine receptors expressed in Chinese hamster ovary cells. Biochem. Pharmacol. 2001, 61, 443–448. [Google Scholar] [CrossRef]
- Schulte, G.; Fredholm, B.B. Human adenosine A(1), A(2A), A(2B), and A(3) receptors expressed in Chinese hamster ovary cells all mediate the phosphorylation of extracellular-regulated kinase 1/2. Mol. Pharmacol. 2000, 58, 477–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskó, G. Adenosine receptor signaling in the brain immune system. Trends Pharmacol. Sci. 2005, 26, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Augusto, E.; Matos, M.; Sevigny, J.; El-Tayeb, A.; Bynoe, M.S.; Muller, C.E.; Cunha, R.A.; Chen, J.F. Ecto-5’-nucleotidase (CD73)-mediated formation of adenosine is critical for the striatal adenosine A2A receptor functions. J. Neurosci. 2013, 33, 11390–11399. [Google Scholar] [CrossRef] [Green Version]
- Rebola, N.; Lujan, R.; Cunha, R.A.; Mulle, C. Adenosine A2A receptors are essential for long-term potentiation of NMDA-EPSCs at hippocampal mossy fiber synapses. Neuron 2008, 57, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Cunha, R.A.; Correia-de-Sa, P.; Sebastiao, A.M.; Ribeiro, J.A. Preferential activation of excitatory adenosine receptors at rat hippocampal and neuromuscular synapses by adenosine formed from released adenine nucleotides. Br. J. Pharmacol. 1996, 119, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Cunha, R.A. How does adenosine control neuronal dysfunction and neurodegeneration? J. Neurochem. 2016, 139, 1019–1055. [Google Scholar] [CrossRef]
- Cunha, R.A. Neuroprotection by adenosine in the brain: From A(1) receptor activation to A (2A) receptor blockade. Purinergic Signal. 2005, 1, 111–134. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.R.; Lourenco, V.S.; Tome, A.R.; Cunha, R.A.; Canas, P.M. Use of knockout mice to explore CNS effects of adenosine. Biochem. Pharmacol. 2021, 187, 114367. [Google Scholar] [CrossRef]
- Agnati, L.F.; Ferre, S.; Lluis, C.; Franco, R.; Fuxe, K. Molecular mechanisms and therapeutical implications of intramembrane receptor/receptor interactions among heptahelical receptors with examples from the striatopallidal GABA neurons. Pharmacol. Rev. 2003, 55, 509–550. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; von Euler, G.; Johansson, B.; Fredholm, B.B.; Fuxe, K. Stimulation of high-affinity adenosine A2 receptors decreases the affinity of dopamine D2 receptors in rat striatal membranes. Proc. Natl. Acad. Sci. USA 1991, 88, 7238–7241. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; Fredholm, B.B.; Morelli, M.; Popoli, P.; Fuxe, K. Adenosine-dopamine receptor-receptor interactions as an integrative mechanism in the basal ganglia. Trends Neurosci. 1997, 20, 482–487. [Google Scholar] [CrossRef]
- Cervetto, C.; Venturini, A.; Passalacqua, M.; Guidolin, D.; Genedani, S.; Fuxe, K.; Borroto-Esquela, D.O.; Cortelli, P.; Woods, A.; Maura, G.; et al. A2A-D2 receptor-receptor interaction modulates gliotransmitter release from striatal astrocyte processes. J. Neurochem. 2017, 140, 268–279. [Google Scholar] [CrossRef]
- Hasko, G.; Cronstein, B.N. Adenosine: An endogenous regulator of innate immunity. Trends Immunol. 2004, 25, 33–39. [Google Scholar] [CrossRef]
- Panther, E.; Corinti, S.; Idzko, M.; Herouy, Y.; Napp, M.; La Sala, A.; Girolomoni, G.; Norgauer, J. Adenosine affects expression of membrane molecules, cytokine and chemokine release, and the T-cell stimulatory capacity of human dendritic cells. Blood 2003, 101, 3985–3990. [Google Scholar] [CrossRef] [Green Version]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Krysko, D.V.; Garg, A.D.; Kaczmarek, A.; Krysko, O.; Agostinis, P.; Vandenabeele, P. Immunogenic cell death and DAMPs in cancer therapy. Nat. Rev. Cancer 2012, 12, 860–875. [Google Scholar] [CrossRef]
- Dosch, M. Mechanisms of ATP Release by Inflammatory Cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef] [Green Version]
- Borea, P.A.; Gessi, S.; Merighi, S.; Varani, K. Adenosine as a multi-signalling guardian angel in human diseases: When, where and how does it exert its protective effects? Trends Pharmacol. Sci. 2016, 37, 419–434. [Google Scholar] [CrossRef]
- Ernst, P.B.; Garrison, J.C.; Thompson, L.F. Much ado about adenosine: Adenosine synthesis and function in regulatory T cell biology. J. Immunol. 2010, 185, 1993–1998. [Google Scholar] [CrossRef] [Green Version]
- Di Virgilio, F.; Vuerich, M. Purinergic signaling in the immune system. Auton. Neurosci. 2015, 191, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qin, W.; Xu, X.; Xiong, Y.; Zhang, Y.; Zhang, H.; Sun, B. Endotoxin-induced autocrine ATP signaling inhibits neutrophil chemotaxis through enhancing myosin light chain phosphorylation. Proc. Natl. Acad. Sci. USA 2017, 114, 4483–4488. [Google Scholar] [CrossRef] [Green Version]
- Koshiba, M.; Kojima, H.; Huang, S.; Apasov, S.; Sitkovsky, M.V. Memory of extracellular adenosine A2A purinergic receptor-mediated signalling in murine T cells. J. Biol. Chem. 1997, 272, 25881–25889. [Google Scholar] [CrossRef]
- Ernens, I. Adenosine up-regulates vascular endothelial growth factor in human macrophages. Biochem. Biophys. Res. Commun. 2010, 392, 351–356. [Google Scholar] [CrossRef]
- Novitskiy, S.V. Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 2008, 112, 1822–1831. [Google Scholar] [CrossRef]
- Abbas, M.; Verville, J.; Mroue, K. Resolution of bronchoconstriction with positive airway pressure after intravenous adenosine. Am. J. Emerg. Med. 2021, 44, e481–e482. [Google Scholar] [CrossRef]
- Shams, S.; Martinez, J.M.; Dawson, J.R.D.; Flores, J.; Gabriel, M.; Garcia, G.; Guevara, A.; Murray, K.; Pacifici, N.; Vargas, M.V.; et al. The Therapeutic Landscape of Rheumatoid Arthritis: Current State and Future Directions. Front. Pharmacol. 2021, 12, 680043. [Google Scholar] [CrossRef]
- Yao, J.K.; Reddy, R.; van Kammen, D.P. Reduced level of plasma antioxidant uric acid in schizophrenia. Psychiatry Res. 1998, 80, 29–39. [Google Scholar] [CrossRef]
- Reddy, R.; Keshavan, M.; Yao, J.K. Reduced plasma antioxidants in first-episode patients with schizophrenia. Schizophr. Res. 2003, 62, 205–212. [Google Scholar] [CrossRef]
- Nagamine, T. Abnormal laboratory values during the acute and recovery phases in schizophrenic patients: A retrospective study. Neuropsychiatr. Dis. Treat. 2010, 6, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Gültekin, B.K.; Kesebir, S.; Kabak, S.G.; Ergün, F.F.; Yaylaci, E.T. Are Uric Acid Levels Different from Healthy Subjects in Bipolar Affective Disorder and Schizophrenia? Relationship Between Clinical Improvement and Episode Severity in Male Patients. Nöro Psikiyatr. Arşivi 2014, 51, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; You, Y.; Yu, L.; Yao, L.; Lu, H.; Zhou, X.; Wu, S.; Chen, L.; Chen, Y.; Zhao, X. Uric acid levels in subjects with schizophrenia: A systematic review and meta-analysis. Psychiatry Res. 2020, 292, 113305. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.H.; Ha, S.K. Uric Acid Puzzle: Dual Role as Anti-oxidantand Pro-oxidant. Electrolyte Blood Press. 2014, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Cathcart, R.; Schwiers, E.; Hochstein, P. Uric acid provides an antioxidant defense in humans against oxidant- and radical-caused aging and cancer: A hypothesis. Proc. Natl. Acad. Sci. USA 1981, 78, 6858–6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godin, O.; Leboyer, M.; Gaman, A.; Aouizerate, B.; Berna, F.; Brunel, L.; Capdevielle, D.; Chereau, I.; Dorey, J.M.; Dubertret, C.; et al. Metabolic syndrome, abdominal obesity and hyperuricemia in schizophrenia: Results from the FACE-SZ cohort. Schizophr. Res. 2015, 168, 388–394. [Google Scholar] [CrossRef]
- Rajan, S.; Zalpuri, I.; Harrington, A.; Cimpeanu, C.; Song, X.; Fan, X. Relationship between serum uric acid level and cardiometabolic risks in nondiabetic patients with schizophrenia. Int. Clin. Psychopharmacol. 2016, 31, 51–56. [Google Scholar] [CrossRef]
- Flatow, J.; Buckley, P.; Miller, B.J. Meta-analysis of oxidative stress in schizophrenia. Biol. Psychiatry 2013, 74, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Ermakov, E.A.; Dmitrieva, E.M.; Parshukova, D.A.; Kazantseva, D.V.; Vasilieva, A.R.; Smirnova, L.P. Oxidative Stress-Related Mechanisms in Schizophrenia Pathogenesis and New Treatment Perspectives. Oxid. Med. Cell. Longev. 2021, 2021, 8881770. [Google Scholar] [CrossRef]
- Arvindakshan, M.; Ghate, M.; Ranjekar, P.K.; Evans, D.R.; Mahadik, S.P. Supplementation with a combination of omega-3 fatty acids and antioxidants (vitamins E and C) improves the outcome of schizophrenia. Schizophr. Res. 2003, 62, 195–204. [Google Scholar] [CrossRef]
- Mahadik, S.P.; Pillai, A.; Joshi, S.; Foster, A. Prevention of oxidative stress-mediated neuropathology and improved clinical outcome by adjunctive use of a combination of antioxidants and omega-3 fatty acids in schizophrenia. Int. Rev. Psychiatry 2006, 18, 119–131. [Google Scholar] [CrossRef]
- Ng, F.; Berk, M.; Dean, O.; Bush, A.I. Oxidative stress in psychiatric disorders: Evidence base and therapeutic implications. Int. J. Neuropsychopharmacol. 2008, 11, 851–876. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.K.; Condray, R.; Dougherty, G.G., Jr.; Keshavan, M.S.; Montrose, D.M.; Matson, W.R.; McEvoy, J.; Kaddurah-Daouk, R.; Reddy, R.D. Associations between purine metabolites and clinical symptoms in schizophrenia. PLoS ONE 2012, 7, e42165. [Google Scholar] [CrossRef] [Green Version]
- Ben Salem, C.; Slim, R.; Fathallah, N.; Hmouda, H. Drug-induced hyperuricaemia and gout. Rheumatology 2017, 56, 679–688. [Google Scholar] [CrossRef]
- Mi, S.; Gong, L.; Sui, Z. Friend or Foe? An Unrecognized Role of Uric Acid in Cancer Development and the Potential Anticancer Effects of Uric Acid-lowering Drugs. J. Cancer 2020, 11, 5236–5244. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Zhao, Q.; Zhang, X.; Wang, X.; Qin, X.; Zhang, R.; Shen, L.; Jiang, X.; Jiang, H.; et al. Association of serum uric acid with increased risk of cancer among hypertensive Chinese. Int. J. Cancer 2017, 141, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Guo, C.; Zheng, S. Elevated urinary 8-oxo-7,8-dihydro-2’-deoxyguanosine and serum uric acid are associated with progression and are prognostic factors of colorectal cancer. Onco Targets Ther. 2018, 11, 5895–5902. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; Li, Q.; Chen, D.T.; Pan, J.H.; Chen, Y.H.; Wen, Z.S.; Zeng, W.A. Prognostic value of pre-operative serum uric acid levels in esophageal squamous cell carcinoma patients who undergo R0 esophagectomy. Cancer Biomark. 2016, 17, 89–96. [Google Scholar] [CrossRef]
- Yuan, C.; Xu, X.H.; Wang, X.L.; Xu, L.; Chen, Z.; Li, Y.Q. Relationship between serum uric acid and metastatic and nonmetastatic rectal cancer patients with undergoing no chemotherapy. Medicine 2016, 95, e5463. [Google Scholar] [CrossRef]
- Stotz, M.; Szkandera, J.; Seidel, J.; Stojakovic, T.; Samonigg, H.; Reitz, D.; Gary, T.; Kornprat, P.; Schaberl-Moser, R.; Hoefler, G.; et al. Evaluation of uric acid as a prognostic blood-based marker in a large cohort of pancreatic cancer patients. PLoS ONE 2014, 9, e104730. [Google Scholar] [CrossRef]
- Stevenson, W.S.; Hyland, C.D.; Zhang, J.G.; Morgan, P.O.; Willson, T.A.; Gill, A.; Hilton, A.A.; Viney, E.M.; Bahlo, M.; Masters, S.L.; et al. Deficiency of 5-hydroxyisourate hydrolase causes hepatomegaly and hepatocellular carcinoma in mice. Proc. Natl. Acad. Sci. USA 2010, 107, 16625–16630. [Google Scholar] [CrossRef] [Green Version]
- Serra, S.; Horenstein, A.L.; Vaisitti, T.; Brusa, D.; Rossi, D.; Laurenti, L.; D’Arena, G.; Coscia, M.; Tripodo, C.; Inghirami, G.; et al. CD73-generated extracellular adenosine in chronic lymphocytic leukemia creates local conditions counteracting drug-induced cell death. Blood 2011, 118, 6141–6152. [Google Scholar] [CrossRef]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.L.; Roberts, O.T. Plasma adenosine deaminase isoform 2 in cancer patients undergoing chemotherapy. Br. J. Biomed. Sci. 2012, 69, 11–13. [Google Scholar] [CrossRef]
- Aghaei, M.; Karami-Tehrani, F.; Salami, S.; Atri, M. Adenosine deaminase activity in the serum and malignant tumors of breast cancer: The assessment of isoenzyme ADA1 and ADA2 activities. Clin. Biochem. 2005, 38, 887–891. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.; Saha, S.; Welham, J.; El Saadi, O.; MacCauley, C.; Chant, D. A systematic review of the incidence of schizophrenia: The distribution of rates and the influence of sex, urbanicity, migrant status and methodology. BMC Med. 2004, 2, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. A systematic review of the prevalence of schizophrenia. PLoS Med. 2005, 2, e141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krugel, U. Purinergic receptors in psychiatric disorders. Neuropharmacology 2016, 104, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Haroutunian, V.; Meador-Woodruff, J.H.; McCullumsmith, R.E. Expression of equilibrative nucleoside transporter type 1 protein in elderly patients with schizophrenia. Neuroreport 2012, 23, 224–227. [Google Scholar] [CrossRef] [Green Version]
- Aliagas, E.; Villar-Menendez, I.; Sevigny, J.; Roca, M.; Romeu, M.; Ferrer, I.; Martin-Satue, M.; Barrachina, M. Reduced striatal ecto-nucleotidase activity in schizophrenia patients supports the “adenosine hypothesis”. Purinergic Signal. 2013, 9, 599–608. [Google Scholar] [CrossRef] [Green Version]
- O’Donovan, S.M.; Sullivan, C.; Koene, R.; Devine, E.; Hasselfeld, K.; Moody, C.L.; McCullumsmith, R.E. Cell-subtype-specific changes in adenosine pathways in schizophrenia. Neuropsychopharmacology 2018, 43, 1667–1674. [Google Scholar] [CrossRef] [Green Version]
- Boison, D. Adenosine kinase: Exploitation for therapeutic gain. Pharmacol. Rev. 2013, 65, 906–943. [Google Scholar] [CrossRef] [Green Version]
- Villar-Menendez, I.; Diaz-Sanchez, S.; Blanch, M.; Albasanz, J.L.; Pereira-Veiga, T.; Monje, A.; Planchat, L.M.; Ferrer, I.; Martin, M.; Barrachina, M. Reduced striatal adenosine A2A receptor levels define a molecular subgroup in schizophrenia. J. Psychiatr. Res. 2014, 51, 49–59. [Google Scholar] [CrossRef]
- Villar-Menendez, I.; Porta, S.; Buira, S.P.; Pereira-Veiga, T.; Diaz-Sanchez, S.; Albasanz, J.L.; Ferrer, I.; Martin, M.; Barrachina, M. Increased striatal adenosine A2A receptor levels is an early event in Parkinson’s disease-related pathology and it is potentially regulated by miR-34b. Neurobiol. Dis. 2014, 69, 206–214. [Google Scholar] [CrossRef]
- Deckert, J.; Brenner, M.; Durany, N.; Zochling, R.; Paulus, W.; Ransmayr, G.; Tatschner, T.; Danielczyk, W.; Jellinger, K.; Riederer, P. Up-regulation of striatal adenosine A(2A) receptors in schizophrenia. Neuroreport 2003, 14, 313–316. [Google Scholar] [CrossRef]
- Valle-Leon, M.; Callado, L.F.; Aso, E.; Cajiao-Manrique, M.M.; Sahlholm, K.; Lopez-Cano, M.; Soler, C.; Altafaj, X.; Watanabe, M.; Ferre, S.; et al. Decreased striatal adenosine A2A-dopamine D2 receptor heteromerization in schizophrenia. Neuropsychopharmacology 2021, 46, 665–672. [Google Scholar] [CrossRef]
- Brunstein, M.G.; Ghisolfi, E.S.; Ramos, F.L.; Lara, D.R. A clinical trial of adjuvant allopurinol therapy for moderately refractory schizophrenia. J. Clin. Psychiatry 2005, 66, 213–219. [Google Scholar] [CrossRef]
- Dickerson, F.B.; Stallings, C.R.; Origoni, A.E.; Sullens, A.; Khushalani, S.; Sandson, N.; Yolken, R.H. A double-blind trial of adjunctive allopurinol for schizophrenia. Schizophr. Res. 2009, 109, 66–69. [Google Scholar] [CrossRef]
- Akhondzadeh, S.; Shasavand, E.; Jamilian, H.; Shabestari, O.; Kamalipour, A. Dipyridamole in the treatment of schizophrenia: Adenosine-dopamine receptor interactions. J. Clin. Pharm. Ther. 2000, 25, 131–137. [Google Scholar] [CrossRef]
- Akhondzadeh, S.; Safarcherati, A.; Amini, H. Beneficial antipsychotic effects of allopurinol as add-on therapy for schizophrenia: A double blind, randomized and placebo controlled trial. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 253–259. [Google Scholar] [CrossRef]
- Wonodi, I.; Gopinath, H.V.; Liu, J.; Adami, H.; Hong, L.E.; Allen-Emerson, R.; McMahon, R.P.; Thaker, G.K. Dipyridamole monotherapy in schizophrenia: Pilot of a novel treatment approach by modulation of purinergic signaling. Psychopharmacology 2011, 218, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Weiser, M.; Gershon, A.A.; Rubinstein, K.; Petcu, C.; Ladea, M.; Sima, D.; Podea, D.; Keefe, R.S.; Davis, J.M. A randomized controlled trial of allopurinol vs. placebo added on to antipsychotics in patients with schizophrenia or schizoaffective disorder. Schizophr. Res. 2012, 138, 35–38. [Google Scholar] [CrossRef]
- Lintunen, J.; Lahteenvuo, M.; Tiihonen, J.; Tanskanen, A.; Taipale, H. Adenosine modulators and calcium channel blockers as add-on treatment for schizophrenia. NPJ Schizophr. 2021, 7, 1. [Google Scholar] [CrossRef]
- Huang, H.; Zheng, S.; Chen, M.; Xie, L.; Li, Z.; Guo, M.; Wang, J.; Lu, M.; Zhu, X. The potential of the P2X7 receptor as a therapeutic target in a sub-chronic PCP-induced rodent model of schizophrenia. J. Chem. Neuroanat. 2021, 116, 101993. [Google Scholar] [CrossRef]
- Wardas, J. Potential role of adenosine A2A receptors in the treatment of schizophrenia. Front. Biosci.-Landmark 2008, 13, 4071–4096. [Google Scholar] [CrossRef] [Green Version]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [Green Version]
- Lawson, D.A.; Kessenbrock, K.; Davis, R.T.; Pervolarakis, N.; Werb, Z. Tumour heterogeneity and metastasis at single-cell resolution. Nat. Cell Biol. 2018, 20, 1349–1360. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Takai, K.; Le, A.; Weaver, V.M.; Werb, Z. Targeting the cancer-associated fibroblasts as a treatment in triple-negative breast cancer. Oncotarget 2016, 7, 82889–82901. [Google Scholar] [CrossRef] [Green Version]
- Di Virgilio, F.; Adinolfi, E. Extracellular purines, purinergic receptors and tumor growth. Oncogene 2017, 36, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Allard, B.; Longhi, M.S.; Robson, S.C.; Stagg, J. The ectonucleotidases CD39 and CD73: Novel checkpoint inhibitor targets. Immunol. Rev. 2017, 276, 121–144. [Google Scholar] [CrossRef] [Green Version]
- Ohta, A.; Sitkovsky, M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage. Nature 2001, 414, 916–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchio, E.A.; Tan, C.Y.; Gregory, K.J.; Christopoulos, A.; White, P.J.; May, L.T. Ligand-Independent Adenosine A2B Receptor Constitutive Activity as a Promoter of Prostate Cancer Cell Proliferation. J. Pharmacol. Exp. Ther. 2016, 357, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannoni, D.; Bernini, A.; Carlucci, F.; Civitelli, S.; Di Pietro, M.C.; Leoncini, R.; Rosi, F.; Tabucchi, A.; Tanzini, G.; Marinello, E. Enzyme activities controlling adenosine levels in normal and neoplastic tissues. Med. Oncol. 2004, 21, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, M.; Tiwari, N.; Sharma, R.; Kaur, S.; Singh, N. Oxidative stress and its relationship with adenosine deaminase activity in various stages of breast cancer. Indian J. Clin. Biochem. 2013, 28, 51–54. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, B.; Amini, S.; Maroofi, F.; Jalali, C.; Javanmardi, M.; Roshani, D.; Abdi, M. Adenosine Deaminase Activity in Chronic Lymphocytic Leukemia and Healthy Subjects. Iran. J. Cancer Prev. 2016, 9, e5069. [Google Scholar] [CrossRef] [Green Version]
- Biri, H.; Ozturk, H.S.; Kacmaz, M.; Karaca, K.; Tokucoglu, H.; Durak, I. Activities of DNA turnover and free radical metabolizing enzymes in cancerous human prostate tissue. Cancer Investig. 1999, 17, 314–319. [Google Scholar] [CrossRef]
- Namiot, Z.; Stasiewicz, J.; Namiot, A.; Kemona, A.; Kralisz, M.; Gorski, J. Adenosine deaminase activity in patients with the intestinal type of gastric carcinoma. Cancer Lett. 1996, 109, 199–202. [Google Scholar] [CrossRef]
- Murray, J.L.; Perez-Soler, R.; Bywaters, D.; Hersh, E.M. Decreased adenosine deaminase (ADA) and 5’nucleotidase (5NT) activity in peripheral blood T cells in Hodgkin disease. Am. J. Hematol. 1986, 21, 57–66. [Google Scholar] [CrossRef]
- Turcotte, M.; Spring, K.; Pommey, S.; Chouinard, G.; Cousineau, I.; George, J.; Chen, G.M.; Gendoo, D.M.; Haibe-Kains, B.; Karn, T.; et al. CD73 is associated with poor prognosis in high-grade serous ovarian cancer. Cancer Res. 2015, 75, 4494–4503. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.Y.; Wang, X.F.; Li, J.; Dong, J.N.; Liu, J.Q.; Li, N.P.; Yun, B.; Xia, R.L. Overexpression of CD39 and high tumoral CD39(+)/CD8(+) ratio are associated with adverse prognosis in resectable gastric cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 14757–14764. [Google Scholar]
- Inoue, Y.; Yoshimura, K.; Kurabe, N.; Kahyo, T.; Kawase, A.; Tanahashi, M.; Ogawa, H.; Inui, N.; Funai, K.; Shinmura, K.; et al. Prognostic impact of CD73 and A2A adenosine receptor expression in non-small-cell lung cancer. Oncotarget 2017, 8, 8738–8751. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, B.G.; Charlebois, R.; Chouinard, G.; Allard, B.; Pommey, S.; Saad, F.; Stagg, J. CD73 Expression Is an Independent Prognostic Factor in Prostate Cancer. Clin. Cancer Res. 2016, 22, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Tondell, A.; Wahl, S.G.F.; Sponaas, A.M.; Sorhaug, S.; Borset, M.; Haug, M. Ectonucleotidase CD39 and Checkpoint Signalling Receptor Programmed Death 1 are Highly Elevated in Intratumoral Immune Cells in Non-small-cell Lung Cancer. Transl. Oncol. 2020, 13, 17–24. [Google Scholar] [CrossRef]
- Nagate, Y.; Ezoe, S.; Fujita, J.; Okuzakis, D.; Motooka, D.; Ishibashi, T.; Ichii, M.; Tanimura, A.; Kurashige, M.; Morii, E.; et al. Ectonucleotidase CD39 is highly expressed on ATLL cells and is responsible for their immunosuppressive function. Leukemia 2020, 35, 107–118. [Google Scholar] [CrossRef]
- Mandapathil, M.; Boduc, M.; Roessler, M.; Guldner, C.; Walliczek-Dworschak, U.; Mandic, R. Ectonucleotidase CD39 expression in regional metastases in head and neck cancer. Acta Otolaryngol. 2018, 138, 428–432. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Chen, X.; Li, L.; Li, Y.; Ping, Y.; Huang, L.; Yue, D.; Zhang, Z.; Wang, F.; et al. CD39/CD73 upregulation on myeloid-derived suppressor cells via TGF-beta-mTOR-HIF-1 signaling in patients with non-small cell lung cancer. Oncoimmunology 2017, 6, e1320011. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Cheng, B.; Li, F.S.; Ding, J.H.; Feng, Y.Y.; Zhuo, G.Z.; Wei, H.F.; Zhao, K. High expression of CD39/ENTPD1 in malignant epithelial cells of human rectal adenocarcinoma. Tumour Biol. 2015, 36, 9411–9419. [Google Scholar] [CrossRef]
- Quezada, C.; Garrido, W.; Oyarzun, C.; Fernandez, K.; Segura, R.; Melo, R.; Casanello, P.; Sobrevia, L.; San Martin, R. 5′-ectonucleotidase mediates multiple-drug resistance in glioblastoma multiforme cells. J. Cell. Physiol. 2013, 228, 602–608. [Google Scholar] [CrossRef]
- Loi, S.; Pommey, S.; Haibe-Kains, B.; Beavis, P.A.; Darcy, P.K.; Smyth, M.J.; Stagg, J. CD73 promotes anthracycline resistance and poor prognosis in triple negative breast cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 11091–11096. [Google Scholar] [CrossRef] [Green Version]
- Cappellari, A.R.; Rockenbach, L.; Dietrich, F.; Clarimundo, V.; Glaser, T.; Braganhol, E.; Abujamra, A.L.; Roesler, R.; Ulrich, H.; Battastini, A.M. Characterization of ectonucleotidases in human medulloblastoma cell lines: Ecto-5’NT/CD73 in metastasis as potential prognostic factor. PLoS ONE 2012, 7, e47468. [Google Scholar] [CrossRef]
- Hilchey, S.P.; Kobie, J.J.; Cochran, M.R.; Secor-Socha, S.; Wang, J.C.; Hyrien, O.; Burack, W.R.; Mosmann, T.R.; Quataert, S.A.; Bernstein, S.H. Human follicular lymphoma CD39+-infiltrating T cells contribute to adenosine-mediated T cell hyporesponsiveness. J. Immunol. 2009, 183, 6157–6166. [Google Scholar] [CrossRef] [Green Version]
- Azambuja, J.H.; Schuh, R.S.; Michels, L.R.; Iser, I.C.; Beckenkamp, L.R.; Roliano, G.G.; Lenz, G.S.; Scholl, J.N.; Sevigny, J.; Wink, M.R.; et al. Blockade of CD73 delays glioblastoma growth by modulating the immune environment. Cancer Immunol. Immunother. 2020, 69, 1801–1812. [Google Scholar] [CrossRef]
- Kashyap, A.S.; Thelemann, T.; Klar, R.; Kallert, S.M.; Festag, J.; Buchi, M.; Hinterwimmer, L.; Schell, M.; Michel, S.; Jaschinski, F.; et al. Antisense oligonucleotide targeting CD39 improves anti-tumor T cell immunity. J. Immunother. Cancer 2019, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, Y.; Lin, X.; Gao, Y.; Zhu, Y. Prognositic value of CD73-adenosinergic pathway in solid tumor: A meta-analysis and systematic review. Oncotarget 2017, 8, 57327–57336. [Google Scholar] [CrossRef]
- Waickman, A.T.; Alme, A.; Senaldi, L.; Zarek, P.E.; Horton, M.; Powell, J.D. Enhancement of tumor immunotherapy by deletion of the A2A adenosine receptor. Cancer Immunol. Immunother. 2012, 61, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Koroskenyi, K.; Kiss, B.; Szondy, Z. Adenosine A2A receptor signaling attenuates LPS-induced pro-inflammatory cytokine formation of mouse macrophages by inducing the expression of DUSP1. Biochim. Biophys. Acta 2016, 1863 Pt A, 1461–1471. [Google Scholar] [CrossRef]
- Cekic, C.; Linden, J. Adenosine A2A receptors intrinsically regulate CD8+ T cells in the tumor microenvironment. Cancer Res. 2014, 74, 7239–7249. [Google Scholar] [CrossRef] [Green Version]
- Hasko, G.; Pacher, P.; Deitch, E.A.; Vizi, E.S. Shaping of monocyte and macrophage function by adenosine receptors. Pharmacol. Ther. 2007, 113, 264–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehbi, V.L.; Tasken, K. Molecular Mechanisms for cAMP-Mediated Immunoregulation in T cells—Role of Anchored Protein Kinase A Signaling Units. Front. Immunol. 2016, 7, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desrosiers, M.D.; Cembrola, K.M.; Fakir, M.J.; Stephens, L.A.; Jama, F.M.; Shameli, A.; Mehal, W.Z.; Santamaria, P.; Shi, Y. Adenosine deamination sustains dendritic cell activation in inflammation. J. Immunol. 2007, 179, 1884–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panther, E.; Idzko, M.; Herouy, Y.; Rheinen, H.; Gebicke-Haerter, P.J.; Mrowietz, U.; Dichmann, S.; Norgauer, J. Expression and function of adenosine receptors in human dendritic cells. FASEB J. 2001, 15, 1963–1970. [Google Scholar] [CrossRef]
- Mirza, A.; Basso, A.; Black, S.; Malkowski, M.; Kwee, L.; Pachter, J.A.; Lachowicz, J.E.; Wang, Y.; Liu, S. RNA interference targeting of A1 receptor-overexpressing breast carcinoma cells leads to diminished rates of cell proliferation and induction of apoptosis. Cancer Biol. Ther. 2005, 4, 1355–1360. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Yin, P.; Reierstad, S.; O’Halloran, M.; Coon, V.J.; Pearson, E.K.; Mutlu, G.M.; Bulun, S.E. Adenosine A1 receptor, a target and regulator of estrogen receptoralpha action, mediates the proliferative effects of estradiol in breast cancer. Oncogene 2010, 29, 1114–1122. [Google Scholar] [CrossRef] [Green Version]
- Merighi, S.; Benini, A.; Mirandola, P.; Gessi, S.; Varani, K.; Leung, E.; Maclennan, S.; Borea, P.A. A3 adenosine receptor activation inhibits cell proliferation via phosphatidylinositol 3-kinase/Akt-dependent inhibition of the extracellular signal-regulated kinase 1/2 phosphorylation in A375 human melanoma cells. J. Biol. Chem. 2005, 280, 19516–19526. [Google Scholar] [CrossRef]
- Merighi, S.; Mirandola, P.; Milani, D.; Varani, K.; Gessi, S.; Klotz, K.N.; Leung, E.; Baraldi, P.G.; Borea, P.A. Adenosine receptors as mediators of both cell proliferation and cell death of cultured human melanoma cells. J. Investig. Dermatol. 2002, 119, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Kasama, H.; Sakamoto, Y.; Kasamatsu, A.; Okamoto, A.; Koyama, T.; Minakawa, Y.; Ogawara, K.; Yokoe, H.; Shiiba, M.; Tanzawa, H.; et al. Adenosine A2b receptor promotes progression of human oral cancer. BMC Cancer 2015, 15, 563. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chu, X.; Deng, F.; Tong, L.; Tong, G.; Yi, Y.; Liu, J.; Tang, J.; Tang, Y.; Xia, Y.; et al. The adenosine A2b receptor promotes tumor progression of bladder urothelial carcinoma by enhancing MAPK signaling pathway. Oncotarget 2017, 8, 48755–48768. [Google Scholar] [CrossRef] [Green Version]
- Gessi, S.; Merighi, S.; Varani, K.; Cattabriga, E.; Benini, A.; Mirandola, P.; Leung, E.; Mac Lennan, S.; Feo, C.; Baraldi, S.; et al. Adenosine receptors in colon carcinoma tissues and colon tumoral cell lines: Focus on the A(3) adenosine subtype. J. Cell. Physiol. 2007, 211, 826–836. [Google Scholar] [CrossRef]
- Koszalka, P.; Golunska, M.; Urban, A.; Stasilojc, G.; Stanislawowski, M.; Majewski, M.; Skladanowski, A.C.; Bigda, J. Specific Activation of A3, A2A and A1 Adenosine Receptors in CD73-Knockout Mice Affects B16F10 Melanoma Growth, Neovascularization, Angiogenesis and Macrophage Infiltration. PLoS ONE 2016, 11, e0151420. [Google Scholar] [CrossRef] [Green Version]
- Allen-Gipson, D.S.; Wong, J.; Spurzem, J.R.; Sisson, J.H.; Wyatt, T.A. Adenosine A2A receptors promote adenosine-stimulated wound healing in bronchial epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 290, L849–L855. [Google Scholar] [CrossRef]
- Ghiringhelli, F.; Apetoh, L.; Tesniere, A.; Aymeric, L.; Ma, Y.; Ortiz, C.; Vermaelen, K.; Panaretakis, T.; Mignot, G.; Ullrich, E.; et al. Activation of the NLRP3 inflammasome in dendritic cells induces IL-1beta-dependent adaptive immunity against tumors. Nat. Med. 2009, 15, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.K.; Williams, D.; Drummond, K.; O’Brien, T.; Monif, M. The role of microglia and P2X7 receptors in gliomas. J. Neuroimmunol. 2019, 332, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, E.; Capece, M.; Franceschini, A.; Falzoni, S.; Giuliani, A.L.; Rotondo, A.; Sarti, A.C.; Bonora, M.; Syberg, S.; Corigliano, D.; et al. Accelerated tumor progression in mice lacking the ATP receptor P2X7. Cancer Res. 2015, 75, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marchi, E.; Orioli, E.; Pegoraro, A.; Sangaletti, S.; Portararo, P.; Curti, A.; Colombo, M.P.; Di Virgilio, F.; Adinolfi, E. The P2X7 receptor modulates immune cells infiltration, ectonucleotidases expression and extracellular ATP levels in the tumor microenvironment. Oncogene 2019, 38, 3636–3650. [Google Scholar] [CrossRef]
- Takai, E.; Tsukimoto, M.; Harada, H.; Sawada, K.; Moriyama, Y.; Kojima, S. Autocrine regulation of TGF-beta1-induced cell migration by exocytosis of ATP and activation of P2 receptors in human lung cancer cells. J. Cell Sci. 2012, 125, 5051–5060. [Google Scholar] [CrossRef] [Green Version]
- Leone, R.D.; Emens, L.A. Targeting adenosine for cancer immunotherapy. J. Immunother. Cancer 2018, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Levy, R. SD-101 and BMS-986178 in Treating Patients with Advanced or Metastatic Solid Malignancies. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT03831295 (accessed on 9 August 2022).
- Terp, M.G.; Olesen, K.A.; Arnspang, E.C.; Lund, R.R.; Lagerholm, B.C.; Ditzel, H.J.; Leth-Larsen, R. Anti-human CD73 monoclonal antibody inhibits metastasis formation in human breast cancer by inducing clustering and internalization of CD73 expressed on the surface of cancer cells. J. Immunol. 2013, 191, 4165–4173. [Google Scholar] [CrossRef] [Green Version]
- Yegutkin, G.G.; Marttila-Ichihara, F.; Karikoski, M.; Niemela, J.; Laurila, J.P.; Elima, K.; Jalkanen, S.; Salmi, M. Altered purinergic signaling in CD73-deficient mice inhibits tumor progression. Eur. J. Immunol. 2011, 41, 1231–1241. [Google Scholar] [CrossRef]
- Stagg, J.; Divisekera, U.; Duret, H.; Sparwasser, T.; Teng, M.W.; Darcy, P.K.; Smyth, M.J. CD73-deficient mice have increased antitumor immunity and are resistant to experimental metastasis. Cancer Res. 2011, 71, 2892–2900. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, X.; Zhou, T.; Ma, D.; Chen, S.; Zhi, X.; Yin, L.; Shao, Z.; Ou, Z.; Zhou, P. Ecto-5’-nucleotidase promotes invasion, migration and adhesion of human breast cancer cells. J. Cancer Res. Clin. Oncol. 2008, 134, 365–372. [Google Scholar] [CrossRef]
- Desmet, C.J.; Gallenne, T.; Prieur, A.; Reyal, F.; Visser, N.L.; Wittner, B.S.; Smit, M.A.; Geiger, T.R.; Laoukili, J.; Iskit, S.; et al. Identification of a pharmacologically tractable Fra-1/ADORA2B axis promoting breast cancer metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 5139–5144. [Google Scholar] [CrossRef] [Green Version]
- Zuber, V.; Jonsson, E.G.; Frei, O.; Witoelar, A.; Thompson, W.K.; Schork, A.J.; Bettella, F.; Wang, Y.; Djurovic, S.; Smeland, O.B.; et al. Identification of shared genetic variants between schizophrenia and lung cancer. Sci. Rep. 2018, 8, 674. [Google Scholar] [CrossRef] [Green Version]
- Sargazi, S.; Nia, M.H.; Mirinejad, S.; Moudi, M.; Shahroudi, M.J.; Saravani, R.; Valian-Borojeni, S. Association of a Novel KIF26B Gene Polymorphism with Susceptibility to Schizophrenia and Breast Cancer: A Case-Control Study. Iran. J. Public Health 2021, 50, 397–406. [Google Scholar] [CrossRef]
- Ibanez, K.; Boullosa, C.; Tabares-Seisdedos, R.; Baudot, A.; Valencia, A. Molecular evidence for the inverse comorbidity between central nervous system disorders and cancers detected by transcriptomic meta-analyses. PLoS Genet. 2014, 10, e1004173. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source | Sex | Participants (N) | Cancers | Major Findings |
|---|---|---|---|---|
| [5] | M/F | 480,356 SCZ patients | All sites |
|
| [4] | M/F | 279,938 patients across all 21 analyses | All sites |
|
| [26] | M/F | 31 studies with a median of 33,372 psychotic patients | All sites |
|
| [27] | M | 218,076 men across 13 studies | Prostate |
|
| [11] | M/F | 496,265 SCZ patients across 12 studies | Lung |
|
| [10] | M/F | 312,834 SCZ patients across seven studies | Liver |
|
| [23] | F | 466,244 patients across 15 studies | Breast |
|
| Gene Name | Protein Name | Additional Names |
|---|---|---|
| ENTPD1 | NTPDase1 | CD39, ATPase, ecto-apyrase |
| ENTPD2 | NTPDase2 | CD39L1, ecto-ATPase |
| ENTPD3 | NTPDase3 | CD39L3 |
| ENTPD8 | NTPDase8 | liver canalicular ecto-ATPase |
| NT5E | Ecto-5′nucleotidase | 5′NT, CD73, NT5E |
| Target | Drug Name | Phase | Indication | Case Number | Formulas | Combination Therapy |
|---|---|---|---|---|---|---|
| NT5E | BMS-986179 | I, II | Advanced solid tumors | NCT02754141 | Monocolonal Ab | Nivolumab, rHuPH20 |
| CPI-006 | I, Ib | Advanced solid tumors | NCT03454451 | Monocolonal Ab | Ciforadenant, Pembrolizumab | |
| MEDI-9447 (OLECLUMAB) | I, II | Advanced solid tumors | NCT03611556 | Monocolonal Ab | durvalumab, gemcitabine, nab-paclitaxel, oxaliplatin, leucovorin, 5-FU | |
| I, II | Advanced solid tumors | NCT03381274 | Osimertinib, AZD4635 | |||
| I, II | Advanced solid tumors | NCT03616886 | Durvalumab, Carboplatin, Paclitaxel | |||
| II | Advanced solid tumors | NCT03267589 | Durbalumab, Tremelilumab, MEDI0562 | |||
| II | Breast cancer | NCT03875573 | Durvalumab, Oleclumab | |||
| I, II | TNBC | NCT03742102 | Durvalumab, Capivasertib, Oleclumab, Paclitaxel, Trastuzumab deruxtecan | |||
| II | NCSLC | NCT03334617 | Durvalumab, AZD9150, AZD6738, Vistusertib, Olaparib, Oleclumab, Trastuzumab deruxtecan, cediranib | |||
| A2AR | AZD-4635 | II | Prostate, mCRPC | NCT04089553 | C15H11ClFN5 | Oleclumab, Durvalumab |
| NCSLC | NCT03381274 | Osimertinib, MEDI9447 | ||||
| CPI-444 | I, II | NSCLC | NCT03337698 | C20H21N7O3 | Atezolizumab, Cobimetinib, RO6958688, Pemetrexed, Carboplatin, Linagliptin, Tocilizumab, Ipatasertib, Idasanutlin | |
| NIR-178 | I, II | Solid tumors | NCT03207867 | C10H8BrN7 | PDR001 | |
| I, II | NCSLC | NCT02403193 | ||||
| Caffeine | N/A | Schizophrenia | NCT02832401 | C8H10N4O2 | N/A | |
| Pentoxifylline | I | Schizophrenia | NCT04094207 | C13H18N4O3 | N/A | |
| A3R | CF-102 | Complete | Hepatocellular Carcinoma | NCT00790218 | C18H18ClIN6O4 | Cl-IB-MECA |
| II | NASH | NCT02927314 | Placebo | |||
| I, II | Hepatitis C | NCT00790673 | Placebo | |||
| II | Hepatocellular Carcinoma | NCT02128958 | Placebo |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamoud, A.-R.; Bach, K.; Kakrecha, O.; Henkel, N.; Wu, X.; McCullumsmith, R.E.; O’Donovan, S.M. Adenosine, Schizophrenia and Cancer: Does the Purinergic System Offer a Pathway to Treatment? Int. J. Mol. Sci. 2022, 23, 11835. https://doi.org/10.3390/ijms231911835
Hamoud A-R, Bach K, Kakrecha O, Henkel N, Wu X, McCullumsmith RE, O’Donovan SM. Adenosine, Schizophrenia and Cancer: Does the Purinergic System Offer a Pathway to Treatment? International Journal of Molecular Sciences. 2022; 23(19):11835. https://doi.org/10.3390/ijms231911835
Chicago/Turabian StyleHamoud, Abdul-Rizaq, Karen Bach, Ojal Kakrecha, Nicholas Henkel, Xiaojun Wu, Robert E. McCullumsmith, and Sinead M. O’Donovan. 2022. "Adenosine, Schizophrenia and Cancer: Does the Purinergic System Offer a Pathway to Treatment?" International Journal of Molecular Sciences 23, no. 19: 11835. https://doi.org/10.3390/ijms231911835