
Cellular and Molecular Biological Alterations after Photon, Proton, and Carbon Ions Irradiation in Human Chondrosarcoma Cells Linked with High-Quality Physics Data
 , , and
, , and
Abstract
:1. Introduction
2. Results
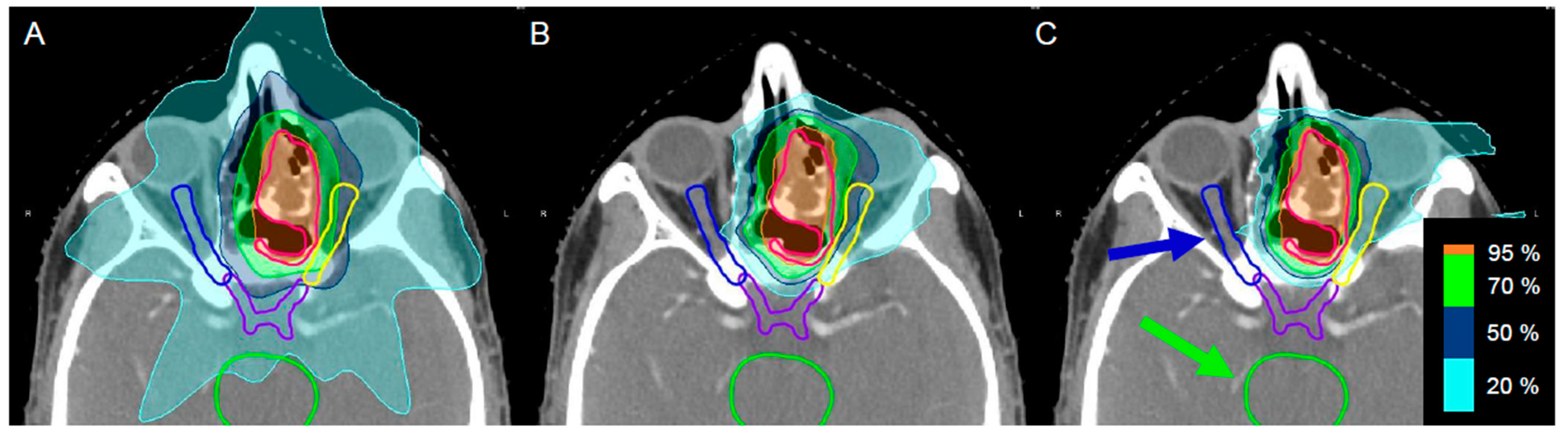
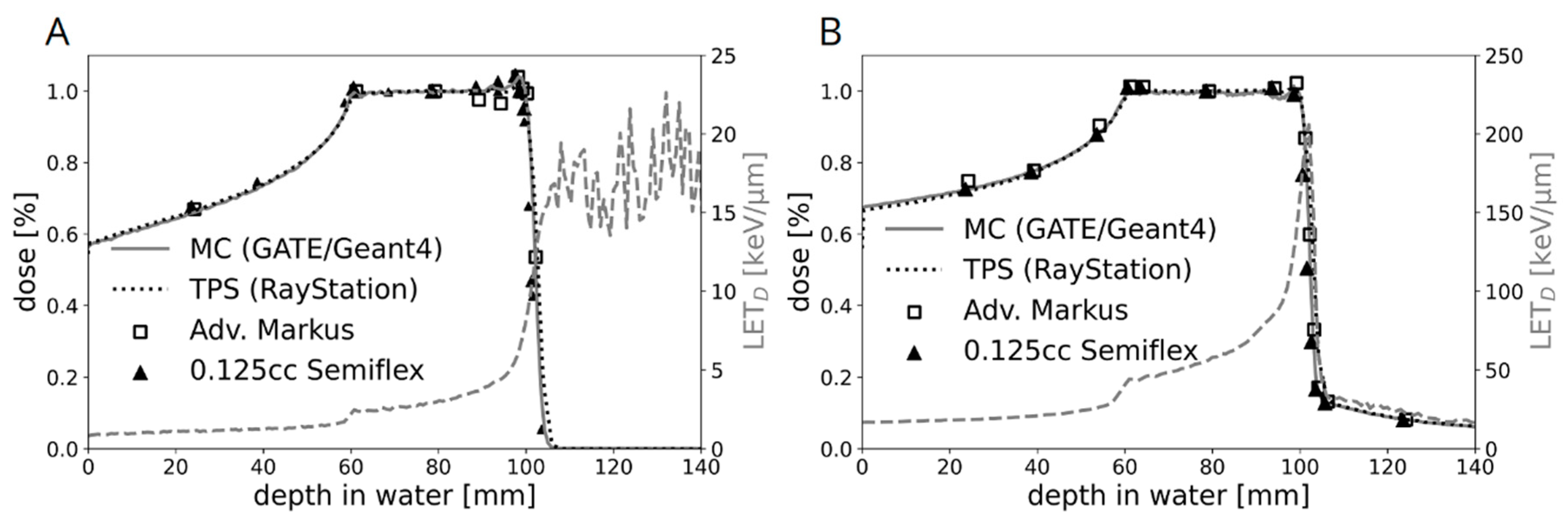
2.1. Dosimetric Characterization of the Novel Water Phantom Measurement Set-Up
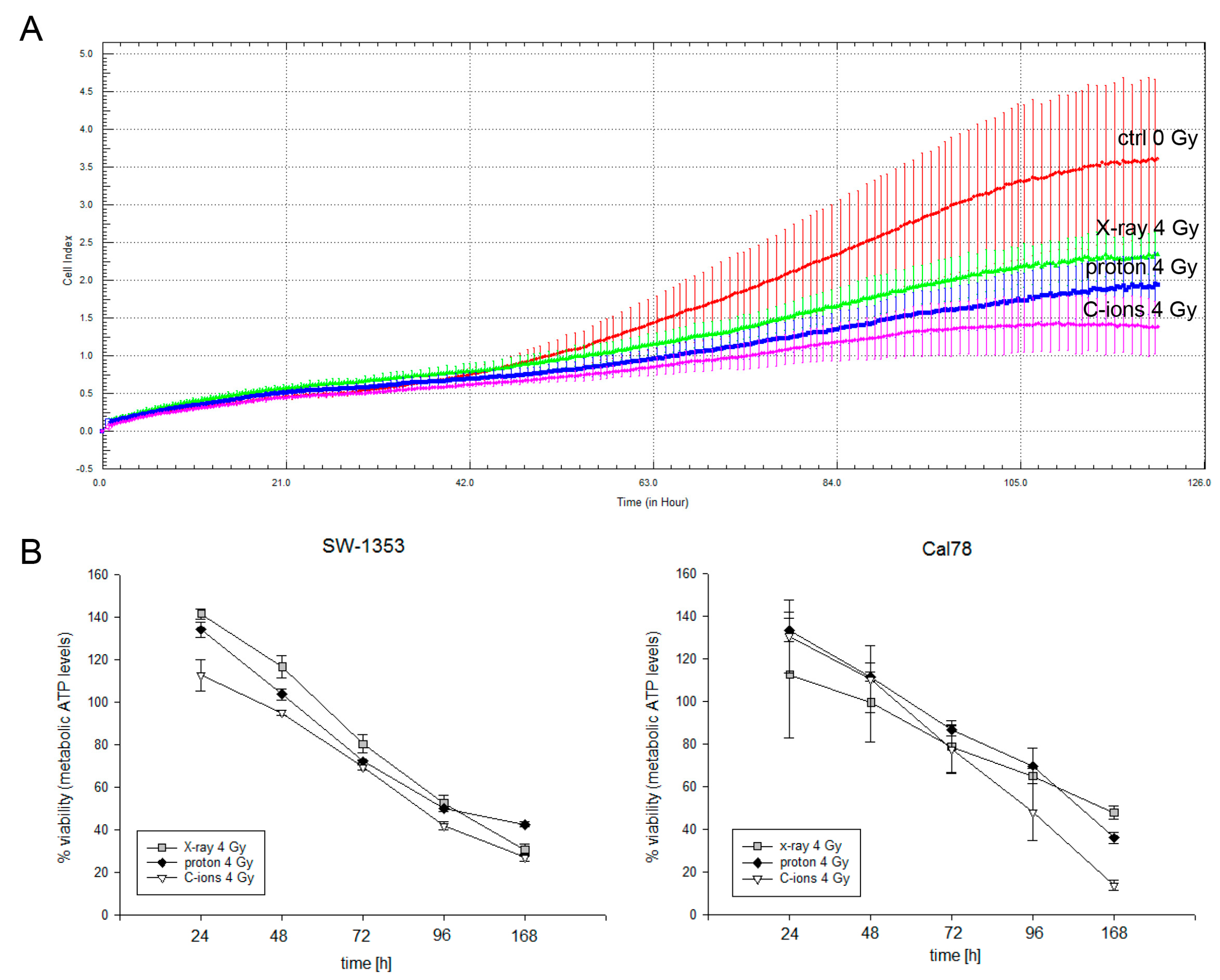
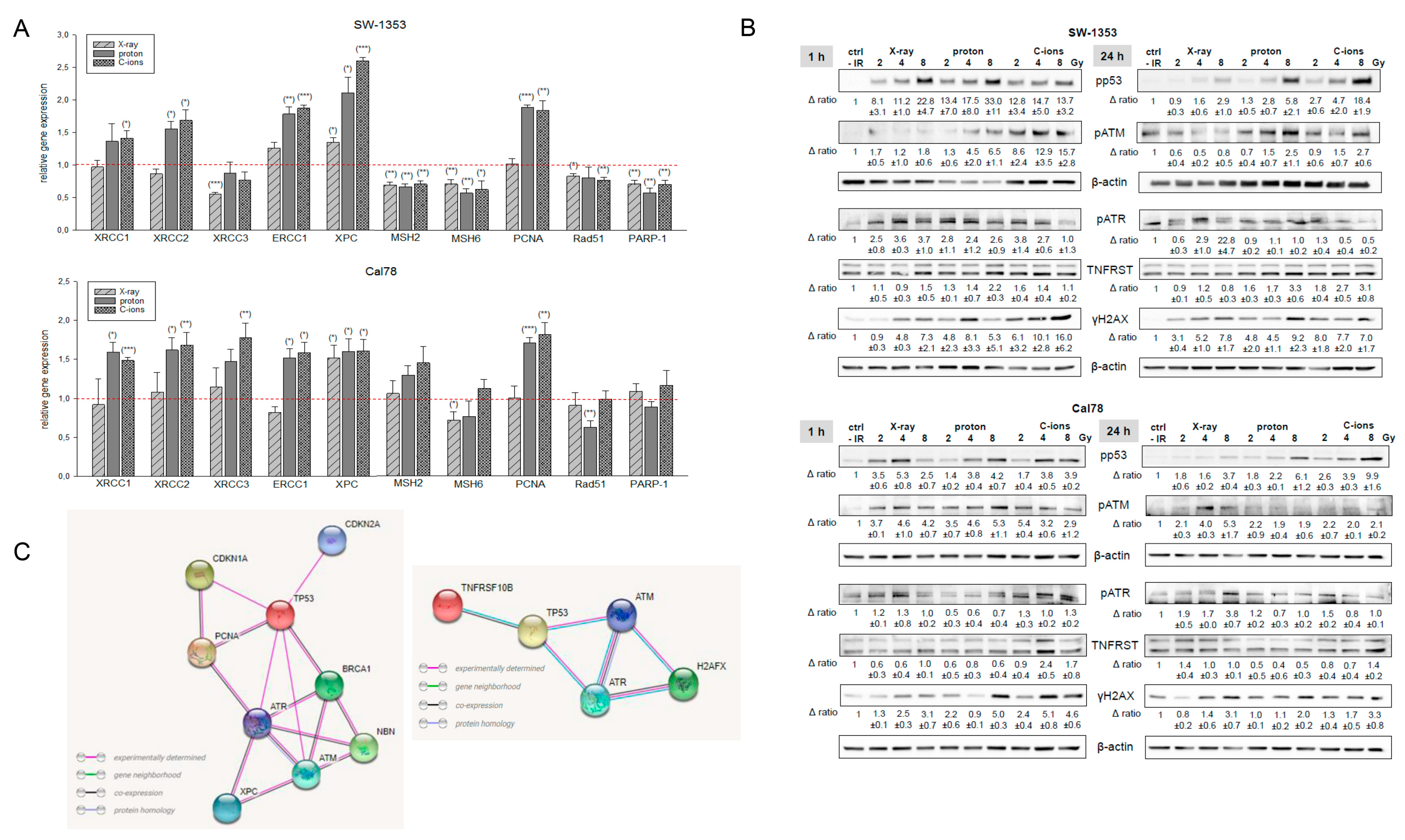
2.2. Cell Biological Alterations after Photon, Proton, and C-Ions IR in Human Chondrosarcoma Cells
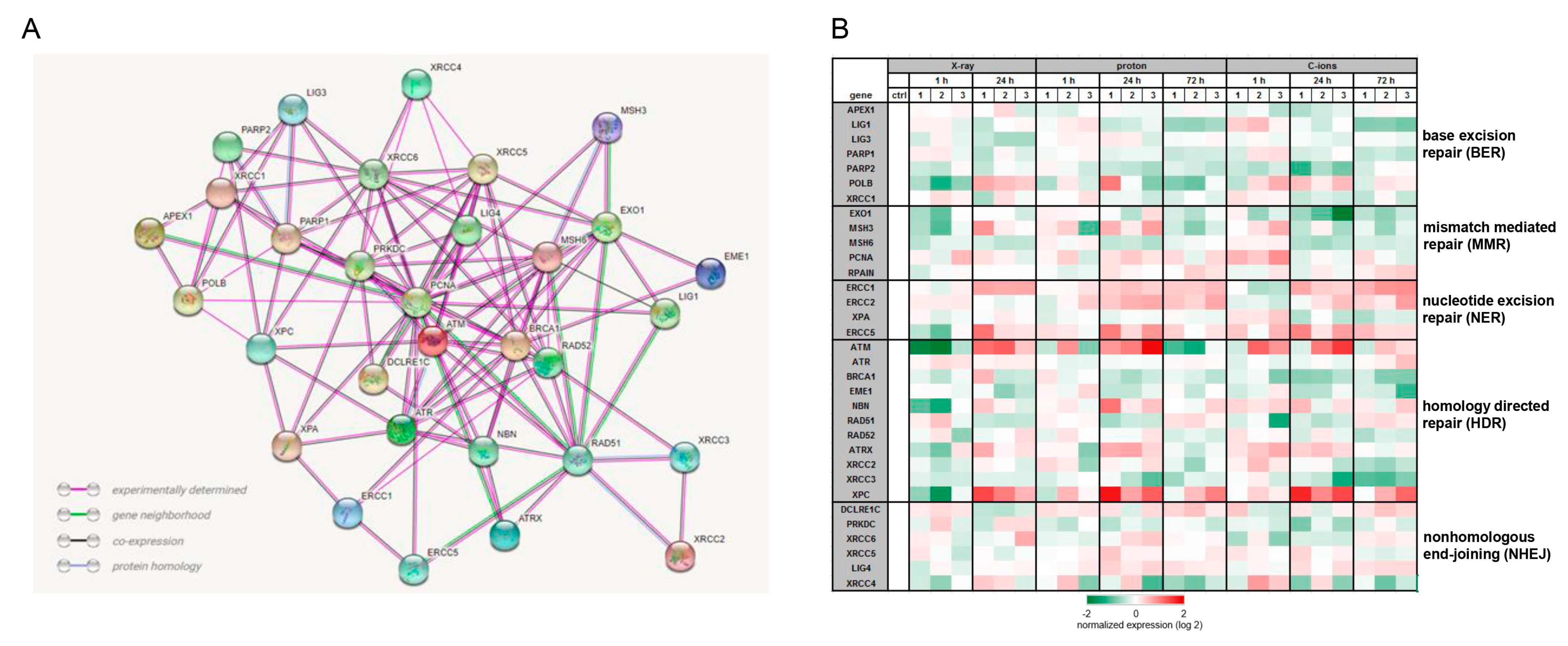
2.3. Activation of DNA Repair Mechanisms after IR
3. Discussion
4. Materials and Methods
4.1. Physical Parameters of Irradiation
4.2. Dosimetric Verification of the Experimental Setup
4.3. Monte Carlo Simulations
4.4. Cell Culture
4.5. Viability and Proliferation Analysis
4.6. Cell Cycle Analysis
4.7. Gene Expression Profiling (RNA Sequencing)
4.8. Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR)
4.9. Protein Expression Analysis
4.10. Study Limitation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, C.D.M.; Hogendoorn, P.C.W.; Mertens, F. Chondrosarcoma. In World Health Organization Classification of Tumours of Soft Tissue and Bone, 4th ed.; IARC Press: Lyon, France, 2013; Volume 5, pp. 264–274. [Google Scholar]
- Valery, P.C.; Laversanne, M.; Bray, F. Bone cancer incidence by morphological subtype: A global assessment. Cancer Causes Control. 2015, 26, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, M.K.; Parry, M.C.; Le Nail, L.R.; Wigley, C.H.; Stevenson, J.D.; Jeys, L.M. Locally recurrent chondrosarcoma of the pelvis and limbs can only be controlled by wide local excision. Bone Joint J. 2019, 101-B, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Zając, A.E.; Kopeć, S.; Szostakowski, B.; Spałek, M.J.; Fiedorowicz, M.; Bylina, E.; Filipowicz, P.; Szumera-Ciećkiewicz, A.; Tysarowski, A.; Czarnecka, A.M.; et al. Chondrosarcoma-from Molecular Pathology to Novel Therapies. Cancers 2021, 13, 2390. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Gao, J.; Hu, J.; Hu, W.; Yang, J.; Qiu, X.; Hu, C.; Kong, L.; Lu, J.J. The preliminary results of proton and carbon ion therapy for chordoma and chondrosarcoma of the skull base and cervical spine. Radiat. Oncol. 2019, 14, 206. [Google Scholar] [CrossRef]
- Minten, E.V.; Yu, D.S. DNA Repair: Translation to the Clinic. Clin Oncol. 2019, 31, 303–310. [Google Scholar] [CrossRef]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef]
- Cuccia, F.; Fiore, M.R.; Barcellini, A.; Iannalfi, A.; Vischioni, B.; Ronchi, S.; Bonora, M.; Riva, G.; Vai, A.; Facoetti, A.; et al. Outcome and Toxicity of Carbon Ion Radiotherapy for Axial Bone and Soft Tissue Sarcomas. Anticancer Res. 2020, 40, 2853–2859. [Google Scholar] [CrossRef]
- Riva, G.; Cavallo, I.; Gandini, S.; Ingargiola, R.; Pecorilla, M.; Imparato, S.; Rossi, E.; Mirandola, A.; Ciocca, M.; Orlandi, E.; et al. Particle Radiotherapy for Skull Base Chondrosarcoma: A Clinical Series from Italian National Center for Oncological Hadrontherapy. Cancers 2021, 13, 4423. [Google Scholar] [CrossRef]
- Barna, S.; Resch, A.F.; Puchalska, M.; Georg, D.; Palmans, H. Technical note: Experimental determination of the effective point of measurement of the PTW-31010 ionization chamber in proton and carbon ion beams. Med. Phys. 2022, 49, 675–681. [Google Scholar] [CrossRef]
- Durante, M.; Paganetti, H.; Pompos, A.; Kry, S.F.; Wu, X.; Grosshans, D.R. Report of a National Cancer Institute special panel: Characterization of the physical parameters of particle beams for biological research. Med. Phys. 2019, 46, e37–e52. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Kang, J.O. Basics of particle therapy I: Physics. Radiat. Oncol. J. 2011, 29, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Kuess, P.; Böhlen, T.T.; Lechner, W.; Elia, A.; Georg, D.; Palmans, H. Lateral response heterogeneity of Bragg peak ionization chambers for narrow-beam photon and proton dosimetry. Phys. Med. Biol. 2017, 62, 9189–9206. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Gales, S. Chapter I. Hadron therapy. In Nuclear Physics for Medicine; NuPECC Report; Nuclear Physics European Collaboration Committee: Strasbourg, France, 2014. [Google Scholar]
- Mizoe, J.E. Review of carbon ion radiotherapy for skull base tumors (especially chordomas). Rep. Pract. Oncol. Radiother. 2016, 21, 356–360. [Google Scholar] [CrossRef]
- Kamada, T.; Tsujii, H.; Blakely, E.A.; Debus, J.; De Neve, W.; Durante, M.; Jäkel, O.; Mayer, R.; Orecchia, R.; Pötter, R.; et al. Carbon ion radiotherapy in Japan: An assessment of 20 years of clinical experience. Lancet Oncol. 2015, 16, e93–e100. [Google Scholar] [CrossRef]
- Girard, N.; Lhuissier, E.; Aury-Landas, J.; Cauvard, O.; Lente, M.; Boittin, M.; Baugé, C.; Boumédiene, K. Heterogeneity of chondrosarcomas response to irradiations with X-rays and carbon ions: A comparative study on five cell lines. J. Bone Oncol. 2020, 22, 100283. [Google Scholar] [CrossRef] [PubMed]
- Lohberger, B.; Glänzer, D.; Eck, N.; Kerschbaum-Gruber, S.; Mara, E.; Deycmar, S.; Madl, T.; Kashofer, K.; Georg, P.; Leithner, A.; et al. Activation of efficient DNA repair mechanisms after photon and proton irradiation of human chondrosarcoma cells. Sci. Rep. 2021, 11, 24116. [Google Scholar] [CrossRef]
- Meerz, A.; Deville, S.S.; Müller, J.; Cordes, N. Comparative Therapeutic Exploitability of Acute Adaptation Mechanisms to Photon and Proton Irradiation in 3D Head and Neck Squamous Cell Carcinoma Cell Cultures. Cancers 2021, 13, 1190. [Google Scholar] [CrossRef]
- Görte, J.; Beyreuther, E.; Danen, E.H.J.; Cordes, N. Comparative Proton and Photon Irradiation Combined with Pharmacological Inhibitors in 3D Pancreatic Cancer Cultures. Cancers 2020, 12, 3216. [Google Scholar] [CrossRef]
- Keta, O.D.; Todorović, D.V.; Bulat, T.M.; Cirrone, P.G.; Romano, F.; Cuttone, G.; Petrović, I.M.; Ristić Fira, A.M. Comparison of human lung cancer cell radiosensitivity after irradiations with therapeutic protons and carbon ions. Exp. Biol. Med. 2017, 242, 1015–1024. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, Y.; Yang, Z.; Liu, Y.; Liu, R.; Geng, Y.; Luo, H.; Zhang, H.; Li, H.; Feng, S.; et al. Preliminary study on radiosensitivity to carbon ions in human breast cancer. J. Radiat. Res. 2020, 61, 399–409. [Google Scholar] [CrossRef]
- Maity, A.; McKenna, W.G.; Muschel, R.J. The molecular basis for cell cycle delays following ionizing radiation: A review. Radiother. Oncol. 1994, 31, 1–13. [Google Scholar] [CrossRef]
- Li, M.; You, L.; Xue, J.; Lu, Y. Ionizing Radiation-Induced Cellular Senescence in Normal, Non-transformed Cells and the Involved DNA Damage Response: A Mini Review. Front. Pharmacol. 2018, 9, 522. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, K.; Xia, Y.; Li, Y.; Hou, Y.; Wang, L.; Li, L.; Chang, L.; Li, W. Cellular senescence in ionizing radiation (Review). Oncol. Rep. 2019, 42, 883–894. [Google Scholar] [CrossRef] [PubMed]
- de Keizer, P.L.; Packer, L.M.; Szypowska, A.A.; Riedl-Polderman, P.E.; van den Broek, N.J.; de Bruin, A.; Dansen, T.B.; Marais, R.; Brenkman, A.B.; Burgering, B.M. Activation of forkhead box O transcription factors by oncogenic BRAF promotes p21cip1-dependent senescence. Cancer Res. 2010, 70, 8526–8536. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.L.; Liu, L.; Mitra, P.; van Wijnen, A.J.; Stein, G.S. Transcriptional activation of the histone nuclear factor P (HiNF-P) gene by HiNF-P and its cyclin E/CDK2 responsive co-factor p220NPAT defines a novel autoregulatory loop at the G1/S phase transition. Gene 2007, 402, 94–102. [Google Scholar] [CrossRef]
- Nickoloff, J.A.; Sharma, N.; Taylor, L. Clustered DNA Double-Strand Breaks: Biological Effects and Relevance to Cancer Radiotherapy. Genes 2020, 11, 99. [Google Scholar] [CrossRef]
- Toulany, M. Targeting DNA Double-Strand Break Repair Pathways to Improve Radiotherapy Response. Genes 2019, 10, 25. [Google Scholar] [CrossRef]
- Cleary, J.M.; Aguirre, A.J.; Shapiro, G.I.; D’Andrea, A.D. Biomarker-Guided Development of DNA Repair Inhibitors. Mol. Cell. 2020, 78, 1070–1085. [Google Scholar] [CrossRef]
- Gerelchuluun, A.; Manabe, E.; Ishikawa, T.; Sun, L.; Itoh, K.; Sakae, T.; Suzuki, K.; Hirayama, R.; Asaithamby, A.; Chen, D.J.; et al. The major DNA repair pathway after both proton and carbon-ion radiation is NHEJ, but the HR pathway is more relevant in carbon ions. Radiat. Res. 2015, 183, 345–356. [Google Scholar] [CrossRef]
- van de Kamp, G.; Heemskerk, T.; Kanaar, R.; Essers, J. DNA Double Strand Break Repair Pathways in Response to Different Types of Ionizing Radiation. Front. Genet. 2021, 12, 738230. [Google Scholar] [CrossRef]
- Menolfi, D.; Zha, S. ATM, ATR and DNA-PKcs kinases-the lessons from the mouse models: Inhibition ≠ deletion. Cell Biosci. 2020, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Canman, C.E.; Lim, D.S.; Cimprich, K.A.; Taya, Y.; Tamai, K.; Sakaguchi, K.; Appella, E.; Kastan, M.B.; Siliciano, J.D. Activation of the ATM kinase by ionizing radiation and phosphorylation of p53. Science 1998, 281, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Kuess, P.; Bozsaky, E.; Hopfgartner, J.; Seifritz, G.; Dörr, W.; Georg, D. Technical Note: Dosimetric challenges of small animal irradiation with a commercial X-ray unit. Med. Phys. 2014, 24, 363–372. [Google Scholar]
- Ruangchan, S.; Knäusl, B.; Fuchs, H.; Georg, D.; Clausen, M. Experimental benchmarking of RayStation proton dose calculation algorithms inside and outside the target region in heterogeneous phantom geometries. Med. Phys. 2020, 76, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Agostinelli, S.; Allison, J.; Amako, K.; Apostolakis, J.; Araujo, H.; Arce, P.; Asai, M.; Axen, D.; Banerjee, S.; Barrand, G. GEANT4-a simulation toolkit. In Nuclear Instruments and Methods in Physics Research Section A. Accelerators Spectrometers Detectors and Associated Equipment; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Elia, A.; Resch, A.F.; Carlino, A.; Böhlen, T.T.; Fuchs, H.; Palmans, H.; Letellier, V.; Dreindl, R.; Osorio, J.; Stock, M.; et al. A GATE/Geant4 beam model for the MedAustron non-isocentric proton treatment plans quality assurance. Phys. Med. 2020, 71, 115–123. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SW-1353 | Cal78 | |||||
|---|---|---|---|---|---|---|
| G1/G0 | S | G2/M | G1/G0 | S | G2/M | |
| ctrl 0 Gy | 56.9 ± 2.5 | 29.4 ± 1.1 | 13.7 ± 3.4 | 60.6 ± 2.6 | 27.3 ± 3.2 | 12.1 ± 1.2 |
| X-ray 4 Gy | 69.9 ± 3.6 ** | 14.8 ± 3.1 *** | 15.2 ± 0.8 n.s. | 42.7 ± 6.5 ** | 23.6 ± 2.6 n.s. | 33.6 ± 4.1 *** |
| X-ray 8 Gy | 59.1 ± 2.8 | 7.7 ± 1.1 *** | 33.2 ± 2.3 *** | 16.3 ± 1.8 *** | 29.6 ± 2.4 n.s. | 54.1 ± 1.5 *** |
| proton 4 Gy LET 2.9 keV/μm | 63.4 ± 1.1 ** | 18.8 ± 0.6 *** | 17.5 ± 1.0 n.s. | 39.9 ± 0.8 *** | 25.1 ± 0.7 n.s. | 35.3 ± 1.0 *** |
| proton 8 Gy LET 2.9 keV/μm | 58.1 ± 2.5 n.s. | 8.9 ± 1.3 *** | 33 ± 1.2 *** | 17.1 ± 0.9 *** | 29.8 ± 1.5 n.s. | 52.6 ± 0.9 *** |
| C-ions 4 Gy LET 55.2 keV/μm | 53.3 ± 4.0 | 7.8 ± 1.4 *** | 39.5 ± 4.0 *** | 19.8 ± 1.5 *** | 31.2 ± 2.9 | 48.9 ± 1.5 *** |
| C-ions 8 Gy LET 55.2 keV/μm | 28.7 ± 2.3 *** | 3.6 ± 0.3 *** | 67.7 ± 2.6 *** | 11.8 ± 0.8 *** | 36.7 ± 1.6 ** | 51.5 ± 1.3 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lohberger, B.; Barna, S.; Glänzer, D.; Eck, N.; Kerschbaum-Gruber, S.; Stasny, K.; Leithner, A.; Georg, D. Cellular and Molecular Biological Alterations after Photon, Proton, and Carbon Ions Irradiation in Human Chondrosarcoma Cells Linked with High-Quality Physics Data. Int. J. Mol. Sci. 2022, 23, 11464. https://doi.org/10.3390/ijms231911464
Lohberger B, Barna S, Glänzer D, Eck N, Kerschbaum-Gruber S, Stasny K, Leithner A, Georg D. Cellular and Molecular Biological Alterations after Photon, Proton, and Carbon Ions Irradiation in Human Chondrosarcoma Cells Linked with High-Quality Physics Data. International Journal of Molecular Sciences. 2022; 23(19):11464. https://doi.org/10.3390/ijms231911464
Chicago/Turabian StyleLohberger, Birgit, Sandra Barna, Dietmar Glänzer, Nicole Eck, Sylvia Kerschbaum-Gruber, Katharina Stasny, Andreas Leithner, and Dietmar Georg. 2022. "Cellular and Molecular Biological Alterations after Photon, Proton, and Carbon Ions Irradiation in Human Chondrosarcoma Cells Linked with High-Quality Physics Data" International Journal of Molecular Sciences 23, no. 19: 11464. https://doi.org/10.3390/ijms231911464