
Structural Characteristics of Polysaccharide GP2a in Gardenia jasminoides and Its Immunomodulatory Effect on Macrophages

 ,
,
Abstract
:1. Introduction
2. Results
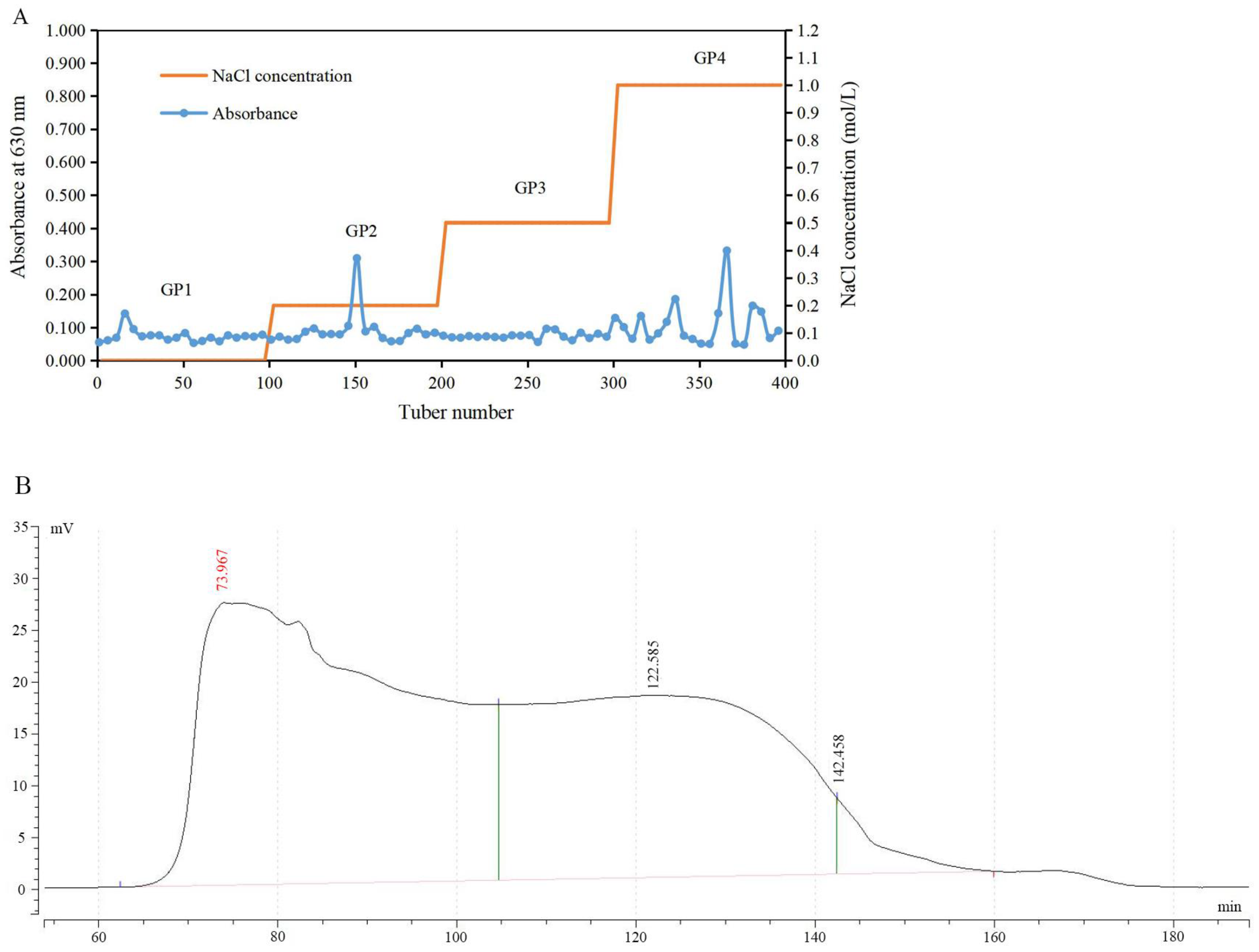
2.1. Polysaccharide Purification
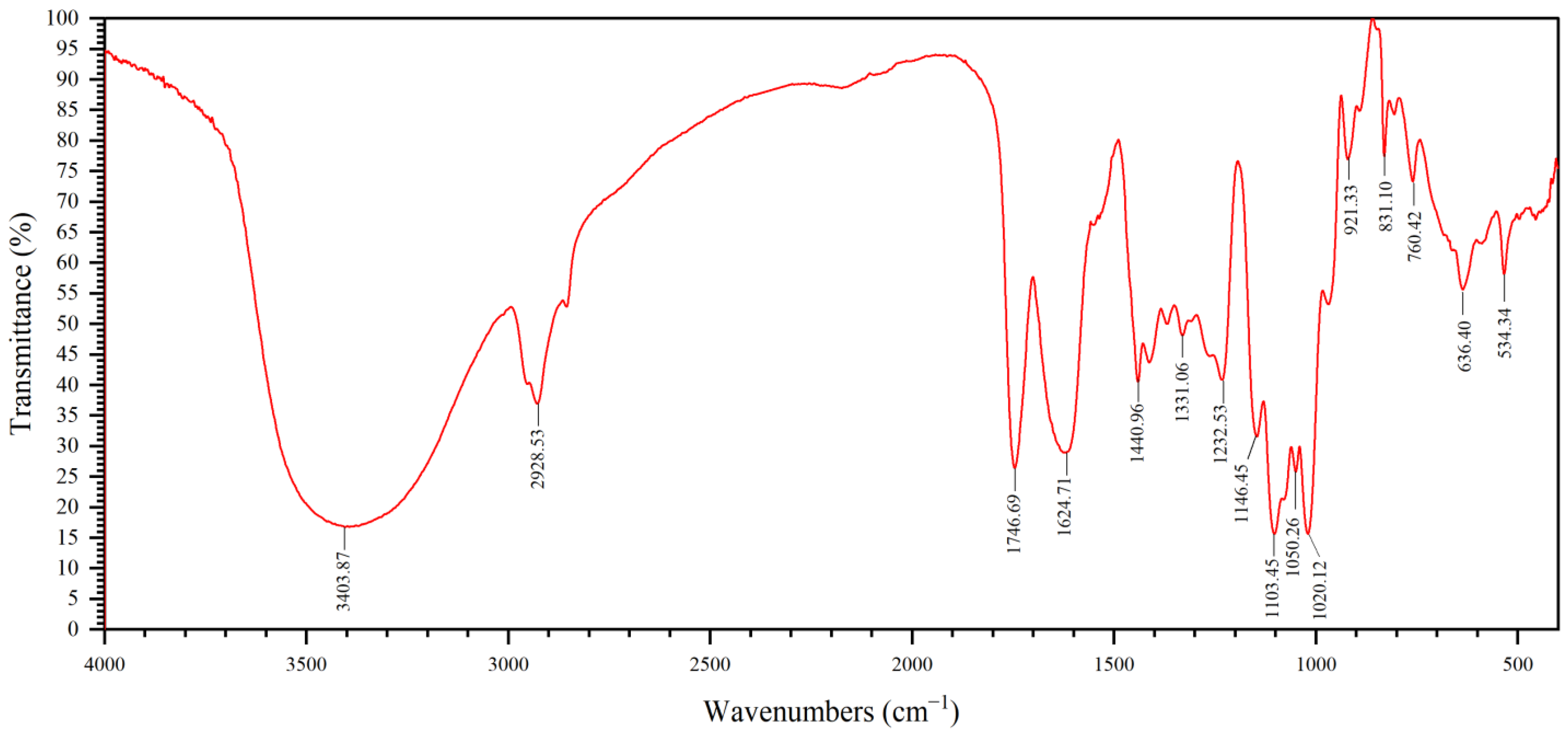
2.2. FTIR Analysis of GP2a
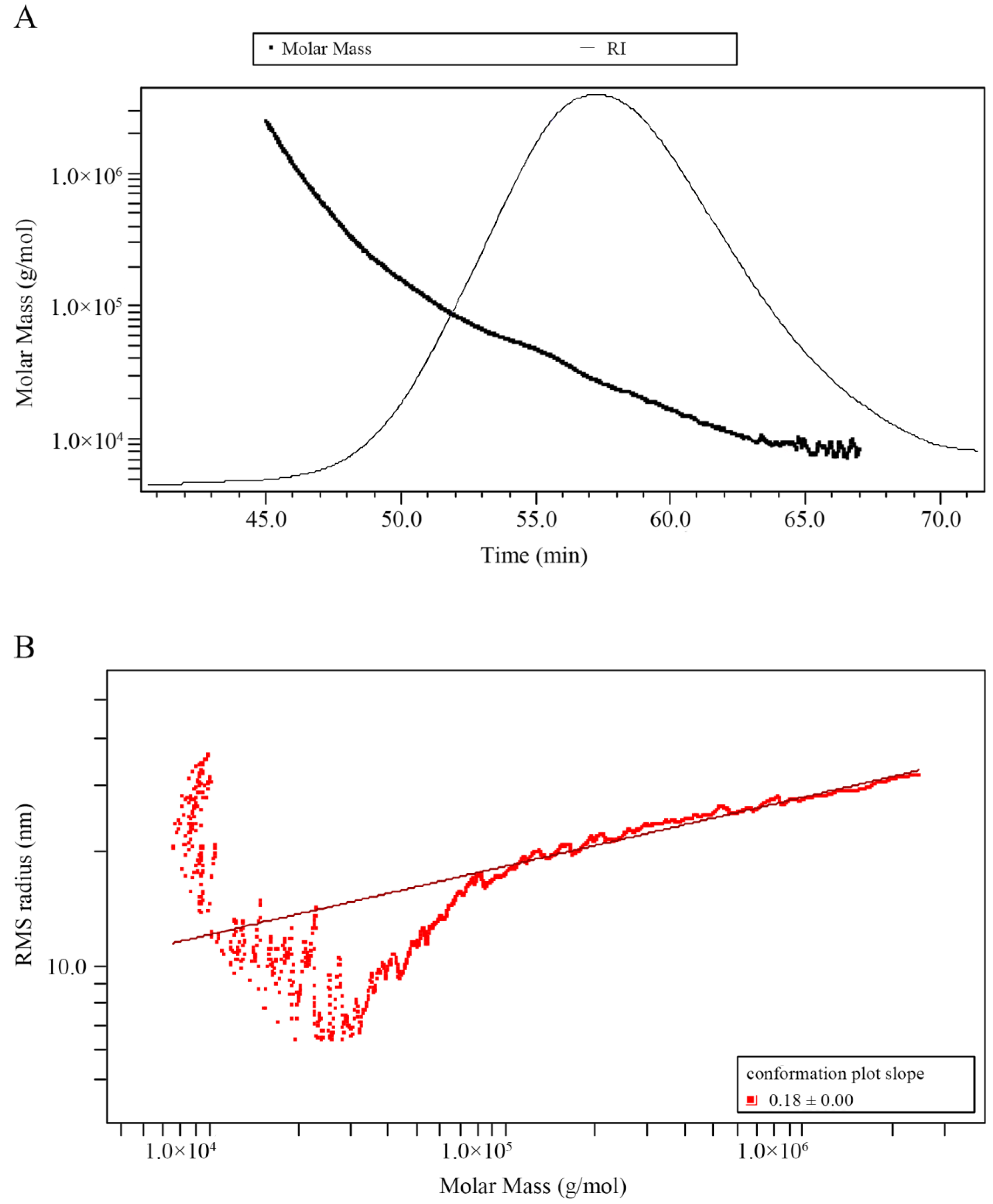
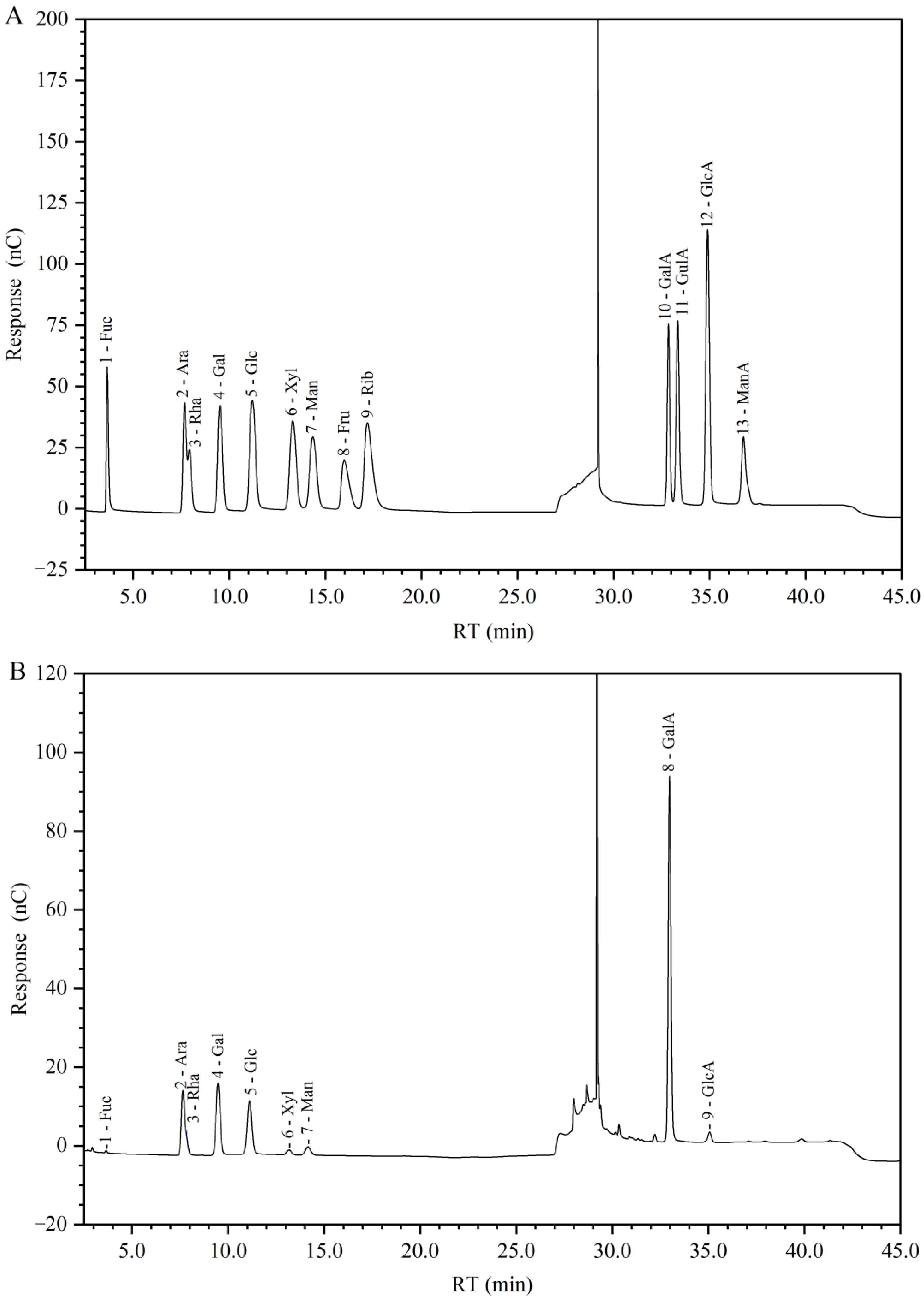
2.3. Molecular Weight and Monosaccharide Composition of GP2a
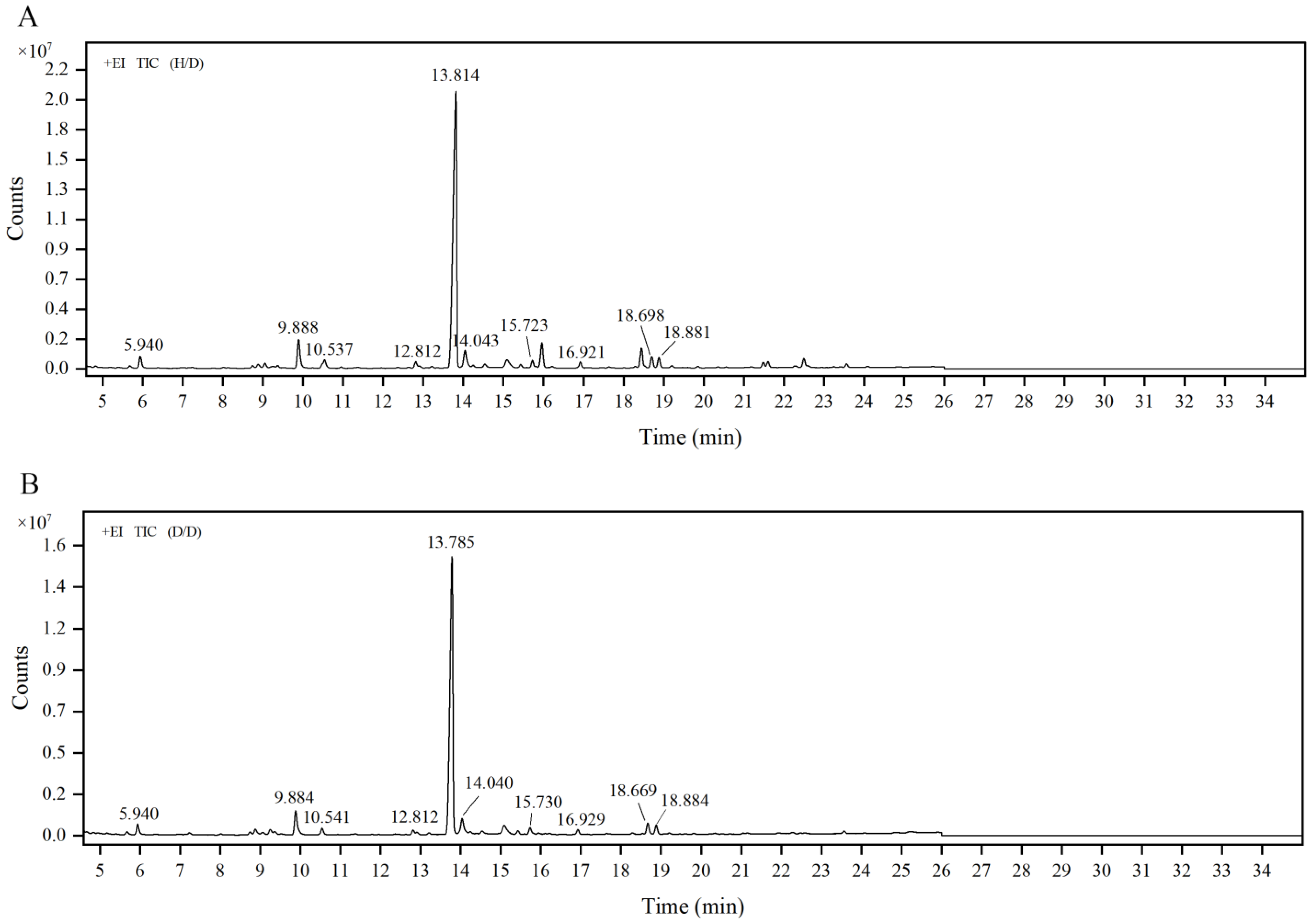
2.4. Linkage Composition of GP2a
2.5. Micromorphology Structure of GP2a
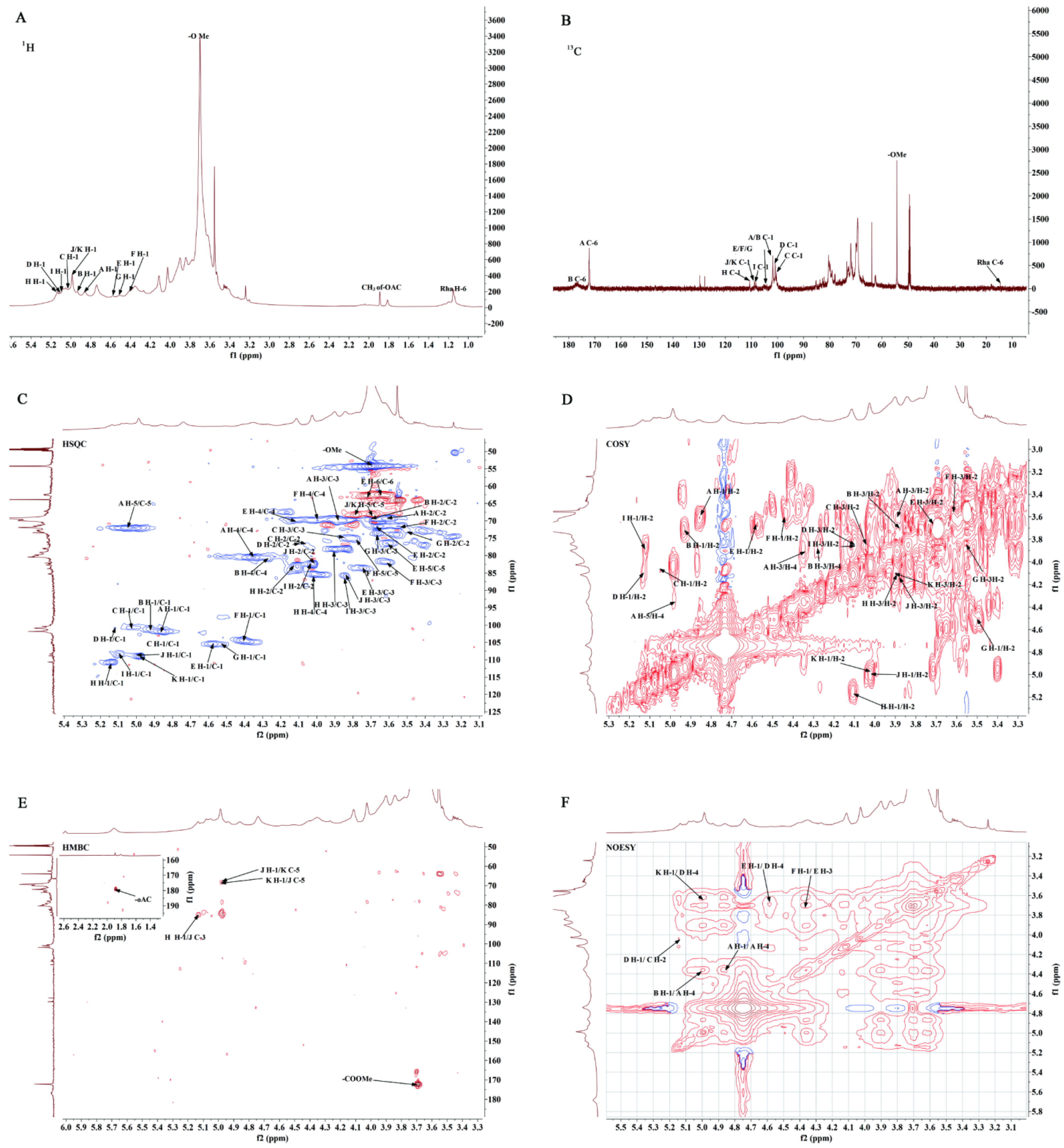
2.6. NMR Spectrum Analysis of GP2a
2.7. Effect of GP2a on RAW 264.7 Cell Viability
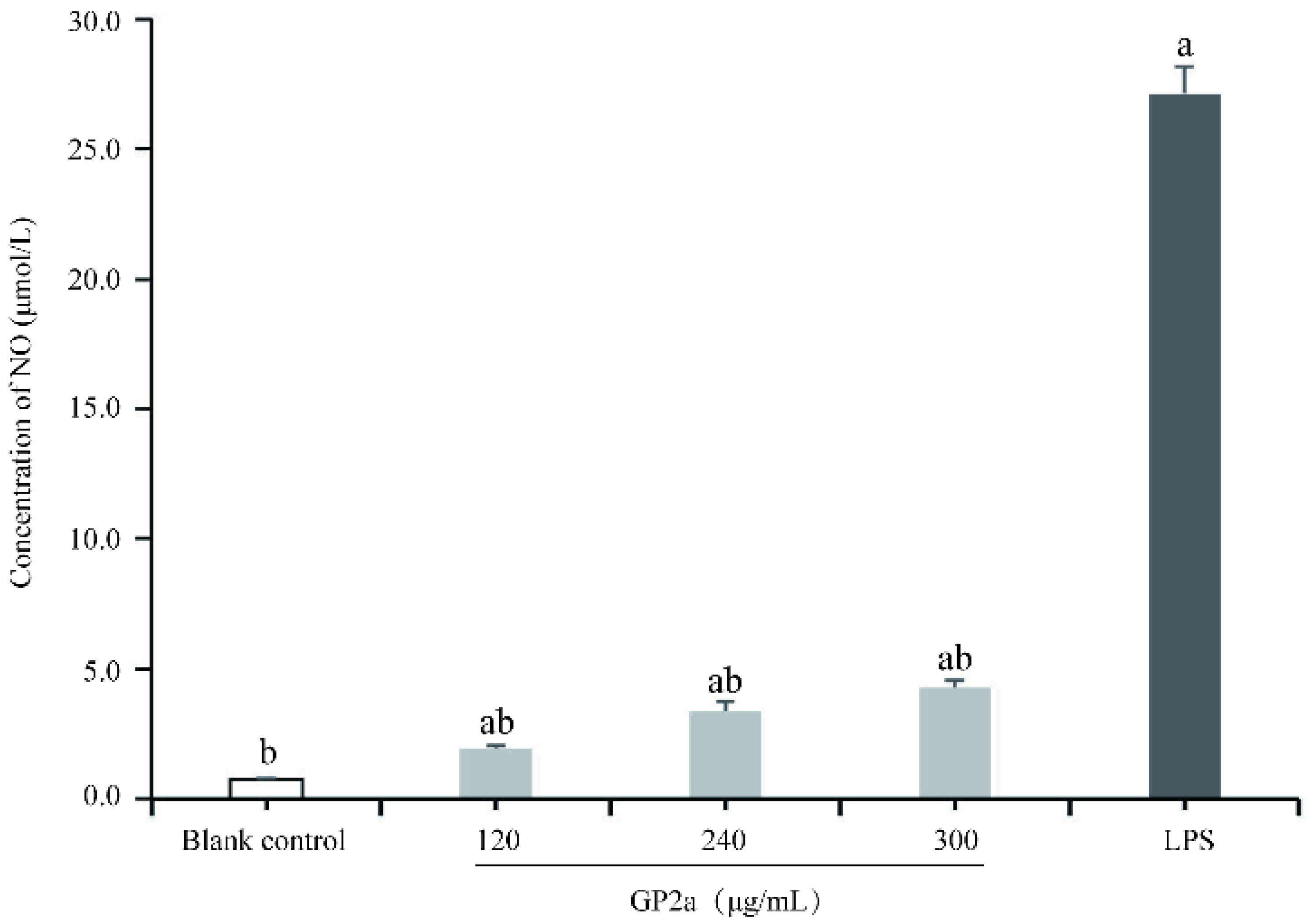
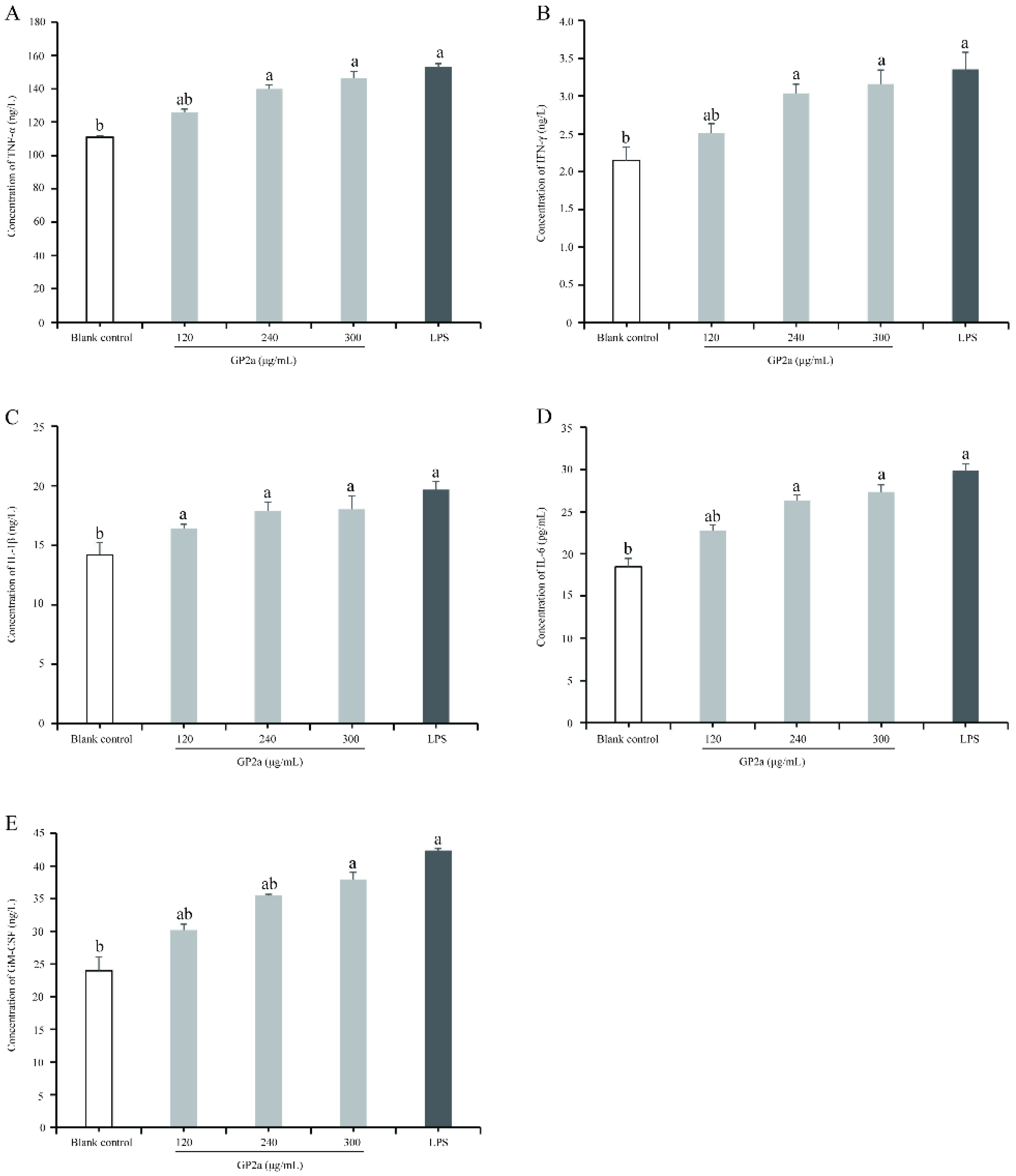
2.8. Effects of GP2a on NO and Cytokine Production in RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. GP2a Preparation
4.2. GP2a Identification
4.2.1. FTIR Scanning
4.2.2. Molecular Weight Determination
4.2.3. Monosaccharide Composition Analysis
4.2.4. Methylation Analysis
4.2.5. SEM Observation
4.2.6. NMR Spectroscopy
4.3. Immunomodulatory Effect of GP2a on RAW 264.7 Cells
4.3.1. Cell Culture
4.3.2. Cell Viability Assay
4.3.3. Determination of NO and Cytokine Production in RAW 264.7 Cells
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, S.C.; Tseng, T.Y.; Huang, C.M.; Tsai, T.H. Gardenia herbal active constituents: Applicable separation procedures. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 812, 193–202. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, F.; Ming, K.; Wang, D.; Hu, Y.; Liu, J. Polysaccharides from Traditional Chinese Medicines: Extraction, Purification, Modification, and Biological Activity. Molecules 2016, 21, 1705. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, R.B.Ö.; Özdemir, A.T.; Sarıboyacı, A.E.; Uysal, O.; Tuğlu, M.İ.; Kırmaz, C. The investigation of immunomodulatory effects of adipose tissue mesenchymal stem cell educated macrophages on the CD4 T cells. Immunobiology 2019, 224, 585–594. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Śliwiński, T.; Zajdel, R. Anti-Inflammatory Activity of Extracts and Pure Compounds Derived from Plants via Modulation of Signaling Pathways, Especially PI3K/AKT in Macrophages. Int. J. Mol. Sci. 2020, 21, 9605. [Google Scholar] [CrossRef]
- Cambeiro-Pérez, N.; González-Gómez, X.; González-Barreiro, C.; Pérez-Gregorio, M.R.; Fernandes, I.; Mateus, N.; de Freitas, V.; Sánchez, B.; Martínez-Carballo, E. Metabolomics Insights of the Immunomodulatory Activities of Phlorizin and Phloretin on Human THP-1 Macrophages. Molecules 2021, 26, 787. [Google Scholar] [CrossRef]
- Yin, M.; Zhang, Y.; Li, H. Advances in Research on Immunoregulation of Macrophages by Plant Polysaccharides. Front. Immunol. 2019, 10, 145. [Google Scholar] [CrossRef]
- Ji, X.; Yan, Y.; Hou, C.; Shi, M.; Liu, Y. Structural characterization of a galacturonic acid-rich polysaccharide from Ziziphus Jujuba cv. Muzao. Int. J. Biol. Macromol. 2020, 147, 844–852. [Google Scholar] [CrossRef]
- Manrique, G.D.; Lajolo, F.M. FT-IR spectroscopy as a tool for measuring degree of methyl esterification in pectins isolated from ripening papaya fruit. Postharvest. Biol. Technol. 2002, 25, 99–107. [Google Scholar] [CrossRef]
- Yang, J.S.; Mu, T.H.; Ma, M.M. Extraction, structure, and emulsifying properties of pectin from potato pulp. Food Chem. 2018, 244, 197–205. [Google Scholar] [CrossRef]
- Wyatt, P.J. Light scattering and the absolute characterization of macromolecules. Anal. Chim. Acta 1993, 272, 1–40. [Google Scholar] [CrossRef]
- Petersen, B.O.; Meier, S.; Duus, J.Ø.; Clausen, M.H. Structural characterization of homogalacturonan by NMR spectroscopy-assignment of reference compounds. Carbohydr. Res. 2008, 343, 2830–2833. [Google Scholar] [CrossRef]
- Yang, J.; Wen, L.; Zhao, Y.; Jiang, Y.; Tian, M.; Liu, H.; Liu, J.; Yang, B. Structure identification of an arabinogalacturonan in Citrus reticulata Blanco ‘Chachiensis’ peel. Food Hydrocolloids 2018, 84, 481–488. [Google Scholar] [CrossRef]
- Patova, O.A.; Smirnov, V.V.; Golovchenko, V.V.; Vityazev, F.V.; Shashkov, A.S.; Popov, S.V. Structural, rheological and antioxidant properties of pectins from Equisetum arvense L. and Equisetum sylvaticum L. Carbohydr. Polym. 2019, 209, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Perrone, P.; Hewage, C.M.; Thomson, A.R.; Bailey, K.; Sadler, I.H.; Fry, S.C. Patterns of methyl and O-acetyl esterification in spinach pectins: New complexity. Phytochemistry 2002, 60, 67–77. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Jarvis, M.C. Acetylation and methylation of homogalacturonans 1, optimisation of the reaction and characterisation of the products. Carbohydr. Polym. 1999, 39, 201–207. [Google Scholar] [CrossRef]
- Zeng, P.; Li, J.; Chen, Y.; Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 423–444. [Google Scholar] [PubMed]
- Han, B.; Baruah, K.; Cox, E.; Vanrompay, D.; Bossier, P. Structure-Functional Activity Relationship of β-Glucans From the Perspective of Immunomodulation: A Mini-Review. Front. Immunol. 2020, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti- Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Wu, N.; Wen, Z.; Xiang, X.; Huang, Y.; Gao, Y.; Qu, Y. Immunostimulative Activity of Low Molecular Weight Chitosans in RAW264.7 Macrophages. Mar. Drugs 2015, 13, 6210–6225. [Google Scholar] [CrossRef]
- Yoo, H.J.; You, D.J.; Lee, K.W. Characterization and Immunomodulatory Effects of High Molecular Weight Fucoidan Fraction from the Sporophyll of Undaria pinnatifida in Cyclophosphamide-Induced Immunosuppressed Mice. Mar. Drugs 2019, 17, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Yao, Y.; Gao, Y.; Hu, Y.; Shi, Z.; Ren, G. Suppressive Effects of Barley β-Glucans with Different Molecular Weight on 3T3-L1 Adipocyte Differentiation. J. Food Sci. 2016, 81, H786–H793. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; Torres, M.D.; González-Muñoz, M.J.; Domínguez, H. Potential of intensification techniques for the extraction and depolymerization of fucoidan. Algal. Res. 2018, 30, 128–148. [Google Scholar] [CrossRef]
- Vogt, L.M.; Sahasrabudhe, N.M.; Ramasamy, U.; Meyer, D.; Pullens, G.; Faas, M.M.; Venema, K.; Schols, H.A.; Vos, P. The impact of lemon pectin characteristics on TLR activation and T84 intestinal epithelial cell barrier function. J. Funct. Foods 2016, 22, 398–407. [Google Scholar] [CrossRef]
- Klosterhoff, R.R.; Bark, J.M.; Glänzel, N.M.; Iacomini, M.; Martinez, G.R.; Winnischofer, S.M.B.; Cordeiro, L.M.C. Structure and intracellular antioxidant activity of pectic polysaccharide from acerola (Malpighia emarginata). Int. J. Biol. Macromol. 2018, 106, 473–480. [Google Scholar] [CrossRef]
- Zou, Y.F.; Barsett, H.; Ho, G.T.; Inngjerdingen, K.T.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. Immunomodulating pectins from root bark, stem bark, and leaves of the Malian medicinal tree Terminalia macroptera, structure activity relations. Carbohydr. Res. 2015, 403, 167–173. [Google Scholar] [CrossRef]
- Ho, G.T.; Zou, Y.F.; Wangensteen, H.; Barsett, H. RG-I regions from elderflower pectins substituted on GalA are strong immunomodulators. Int. J. Biol. Macromol. 2016, 92, 731–738. [Google Scholar] [CrossRef]
- Van der Meide, P.H.; Schellekens, H. Cytokines and the immune response. Biotherapy 1996, 8, 243–249. [Google Scholar] [CrossRef]
- Cavaillon, J.M. Exotoxins and endotoxins: Inducers of inflammatory cytokines. Toxicon 2018, 149, 45–53. [Google Scholar] [CrossRef]
- Chakraborty, I.; Sen, I.K.; Mondal, S.; Rout, D.; Bhanja, S.K.; Maity, P.; Maity, G.N. Bioactive polysaccharides from natural sources: A review on the antitumor and immunomodulating activities. Biocatal. Agric. Biotechnol. 2019, 22, 101425. [Google Scholar] [CrossRef]
- Cao, J.; Tang, D.; Wang, Y.; Li, X.; Hong, L.; Sun, C. Characteristics and immune enhancing activity of pectic polysaccharides from sweet cherry (Prunus avium). Food Chem. 2018, 254, 47–54. [Google Scholar] [CrossRef]
- Wang, H.; Bi, H.; Gao, T.; Zhao, B.; Ni, W.; Liu, J. A homogalacturonan from Hippophae rhamnoides L. Berries enhance immunomodulatory activity through TLR4/MyD88 pathway mediated activation of macrophages. Int. J. Biol. Macromol. 2018, 107, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhao, J.; Wei, Y.; Yu, G.; Li, F.; Li, Q. Structural characterization and mechanisms of macrophage immunomodulatory activity of a pectic polysaccharide from Cucurbita moschata Duch. Carbohydr. Polym. 2021, 269, 118288. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, Y.N.; Paulsen, B.S.; Kiyohara, H.; Ciz, M.; Ognyanov, M.H.; Vasicek, O.; Rise, F.; Denev, P.N.; Yamada, H.; Lojek, A.; et al. The common lavender (Lavandula angustifolia Mill.) pectic polysaccharides modulate phagocytic leukocytes and intestinal Peyer’s patch cells. Carbohydr. Polym. 2017, 174, 948–959. [Google Scholar] [CrossRef]
- Zhang, B.; Leung, W.K.; Zou, Y.; Mabusela, W.; Johnson, Q.; Michaelsen, T.E.; Paulsen, B.S. Immunomodulating polysaccharides from Lessertia frutescens leaves: Isolation, characterization and structure activity relationship. J. Ethnopharmacol. 2014, 152, 340–348. [Google Scholar] [CrossRef]
- Patel, B.K.; Campanella, O.H.; Janaswamy, S. Impact of urea on the three-dimensional structure, viscoelastic and thermal behavior of iota-carrageenan. Carbohydr. Polym. 2013, 92, 1873–1879. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, Y.; Zhang, H.; Li, J.; Tao, W.; Linhardt, R.J.; Chen, S.; Ye, X. Physicochemical properties and conformations of water-soluble peach gums via different preparation methods. Food Hydrocol. 2019, 95, 571–579. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Y.; Sun, P.; Zhang, F.; Linhardt, R.J.; Zhang, A. Chemically modified polysaccharides: Synthesis, characterization, structure activity relationships of action. Int. J. Biol. Macromol. 2019, 132, 970–977. [Google Scholar] [CrossRef]
- Pettolino, F.A.; Walsh, C.; Fincher, G.B.; Bacic, A. Determining the polysaccharide composition of plant cell walls. Nat. Protoc. 2012, 7, 1590–1607. [Google Scholar] [CrossRef]
- Lategan, K.L.; Walters, C.R.; Pool, E.J. The effects of silver nanoparticles on RAW 264.7. Macrophages and human whole blood cell cultures. Front. Biosci. 2019, 24, 347–365. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharide | Relative Percentage * (%) |
|---|---|
| GalA | 63.1 ± 1.7 |
| Ara | 11.1 ± 0.6 |
| Gal | 10.9 ± 0.5 |
| Glc | 7.4 ± 0.3 |
| Rha | 2.7 ± 0.1 |
| Man | 2.2 ± 0.2 |
| GlcA | 1.2 ± 0.0 |
| Xyl | 0.8 ± 0.1 |
| Fuc | 0.2 ± 0.0 |
| Linkage Pattern | Methylated Sugar | Molecular Weight (Da) | Relative Molar Percentage (%) | RT (min) |
|---|---|---|---|---|
| t-Araf | 1,4-di-O-acetyl-2,3,5-tri-O-methyl arabinitol | 279 | 3.1 | 5.940 |
| 5-Araf | 1,4,5-tri-O-acetyl-2,3-di-O-methyl arabinitol | 307 | 3.0 | 10.537 |
| t-Galp | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl galactitol | 323 | 3.4 | 9.888 |
| 3-Galp | 1,3,5-tri-O-acetyl-2,4,6-tri-O-methyl galactitol | 351 | 1.4 | 12.812 |
| 3,4-Galp | 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl galactitol | 379 | 0.7 | 15.723 |
| 3,6-Galp | 1,3,5,6-tetra-O-acetyl-2,4-di-O-methyl galactitol | 379 | 1.9 | 18.881 |
| t-GalpA | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl galactitol (6,6′-dideuterio) | 325 | 3.4 | 9.888 |
| 4-GalpA | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl galactitol (6,6′-dideuterio) | 353 | 74.8 | 13.814 |
| 3,4-GalpA | 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl galactitol (6,6′-dideuterio) | 381 | 0.7 | 15.723 |
| 4,6-GalpA | 1,4,5,6-tetra-O-acetyl-2,3-di-O-methyl galactitol (6,6′-dideuterio) | 381 | 2.0 | 18.698 |
| 4-Glcp | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl glucitol | 351 | 4.3 | 14.043 |
| 2,4-GlcpA | 1,2,4,5-tetra-O-acetyl-3,6-di-O-methyl glucitol (6,6′-dideuterio) | 381 | 1.3 | 16.921 |
| Sugar Residue | Chemical Shift (ppm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6a | 6b | CH3 of OMe | |||
| A | →4)-α-D-GalpA-6-OMe-(1→ | H | 4.86 | 3.62 | 3.89 | 4.35 | 5.05/4.98 | 3.70 | ||
| C | 101.90 | 69.44 | 69.59 | 80.68 | 72.27 | 172.30 | 54.42 | |||
| B | →4)-α-D-GalpA-(1→ | H | 4.94 | 3.70 | 3.90 | 4.27 | n.d. | |||
| C | 101.74 | 69.59 | 69.80 | 80.68 | n.d. | 176.64 | ||||
| C | →2)-α-L-Rhap-(1→ | H | 5.03 | 4.06 | 3.84 | 3.66 | 3.90 | 1.15 | ||
| C | 100.78 | 77.30 | 74.29 | 74.13 | 69.78 | 15.64 | ||||
| D | →2,4)-α-L-Rhap-(1→ | H | 5.12 | 4.09 | 3.84 | 3.65 | 3.90 | 1.15 | ||
| C | 100.83 | 77.05 | 74.29 | 81.72 | 69.78 | 15.64 | ||||
| E | →3)-β-D-Galp-(1→ | H | 4.57 | 3.68 | 3.75 | 4.10 | 3.61 | 3.64 | 3.74 | |
| C | 105.59 | 71.81 | 83.57 | 69.75 | 76.08 | 62.30 | ||||
| F | →3,6)-β-D-Galp-(1→ | H | 4.41 | 3.56 | 3.61 | 4.02 | 3.82 | 3.80 | 3.94 | |
| C | 104.52 | 71.59 | 81.88 | 70.12 | 74.81 | 71.07 | ||||
| G | β-D-Galp-(1→ | H | 4.50 | 3.50 | 3.66 | 3.91 | 3.61 | 3.64 | 3.74 | |
| C | 105.84 | 72.54 | 73.65 | 69.96 | 76.66 | 62.30 | ||||
| H | α-L-Araf-(1→ | H | 5.13 | 4.12 | 3.89 | 4.02 | 3.69/3.74 | |||
| C | 110.85 | 83.57 | 78.14 | 85.26 | 64.89 | |||||
| K | →5)-α-L-Araf-(1→ | H | 4.98 | 4.03 | 3.90 | 4.01 | 3.69/3.78 | |||
| C | 109.38 | 82.46 | 78.14 | 85.47 | 68.43 | |||||
| Groups | Cell Viability (%) | |||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||
| Blank control | 100.0 ± 0.0 b | 100.00 ± 0.0 b | 100.0 ± 0.0 b | |
| Samples | GP2a 7.5 (µg/mL) | 99.4 ± 2.4 b | 103.6 ± 1.3 ab* | 104.8 ± 1.6 ab* |
| GP2a 15 (µg/mL) | 101.3 ± 1.4 b | 103.1 ± 1.5 b | 103.5 ± 2.4 b | |
| GP2a 30 (µg/mL) | 105.3 ± 1.5 ab | 102.2 ± 2.5 b | 104.0 ± 1.4 b | |
| GP2a 60 (µg/mL) | 106.2 ± 1.1 a | 104.9 ± 2.3 ab | 102.1 ± 2.4 b | |
| GP2a 120 (µg/mL) | 103.3 ± 1.5 b | 107.9 ± 2.2 ab* | 105.6 ± 2.4 ab | |
| GP2a 240 (µg/mL) | 102.5 ± 1.9 b | 110.2 ± 1.8 ab* | 110.3 ± 3.55 ab* | |
| GP2a 300 (µg/mL) | 104.3 ± 1.2 ab | 115.1 ± 2.8 ab* | 108.0 ± 3.0 ab | |
| Positive control (2 µg/mL LPS) | 109.8 ± 5.5 a | 118.6 ± 2.1 a* | 45.9 ± 3.2 a* | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, P.; Chen, L.; Huang, X.; Xiao, F.; Fu, L.; Jing, D.; Wang, J.; Zhang, H.; Sun, L.; Wu, Y. Structural Characteristics of Polysaccharide GP2a in Gardenia jasminoides and Its Immunomodulatory Effect on Macrophages. Int. J. Mol. Sci. 2022, 23, 11279. https://doi.org/10.3390/ijms231911279
Lin P, Chen L, Huang X, Xiao F, Fu L, Jing D, Wang J, Zhang H, Sun L, Wu Y. Structural Characteristics of Polysaccharide GP2a in Gardenia jasminoides and Its Immunomodulatory Effect on Macrophages. International Journal of Molecular Sciences. 2022; 23(19):11279. https://doi.org/10.3390/ijms231911279
Chicago/Turabian StyleLin, Pingdong, Lifei Chen, Xiaojing Huang, Fangnan Xiao, Lei Fu, Dingding Jing, Jingjing Wang, Hong Zhang, Lifang Sun, and Yunkun Wu. 2022. "Structural Characteristics of Polysaccharide GP2a in Gardenia jasminoides and Its Immunomodulatory Effect on Macrophages" International Journal of Molecular Sciences 23, no. 19: 11279. https://doi.org/10.3390/ijms231911279