
Bmal1 Regulates Prostate Growth via Cell-Cycle Modulation

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
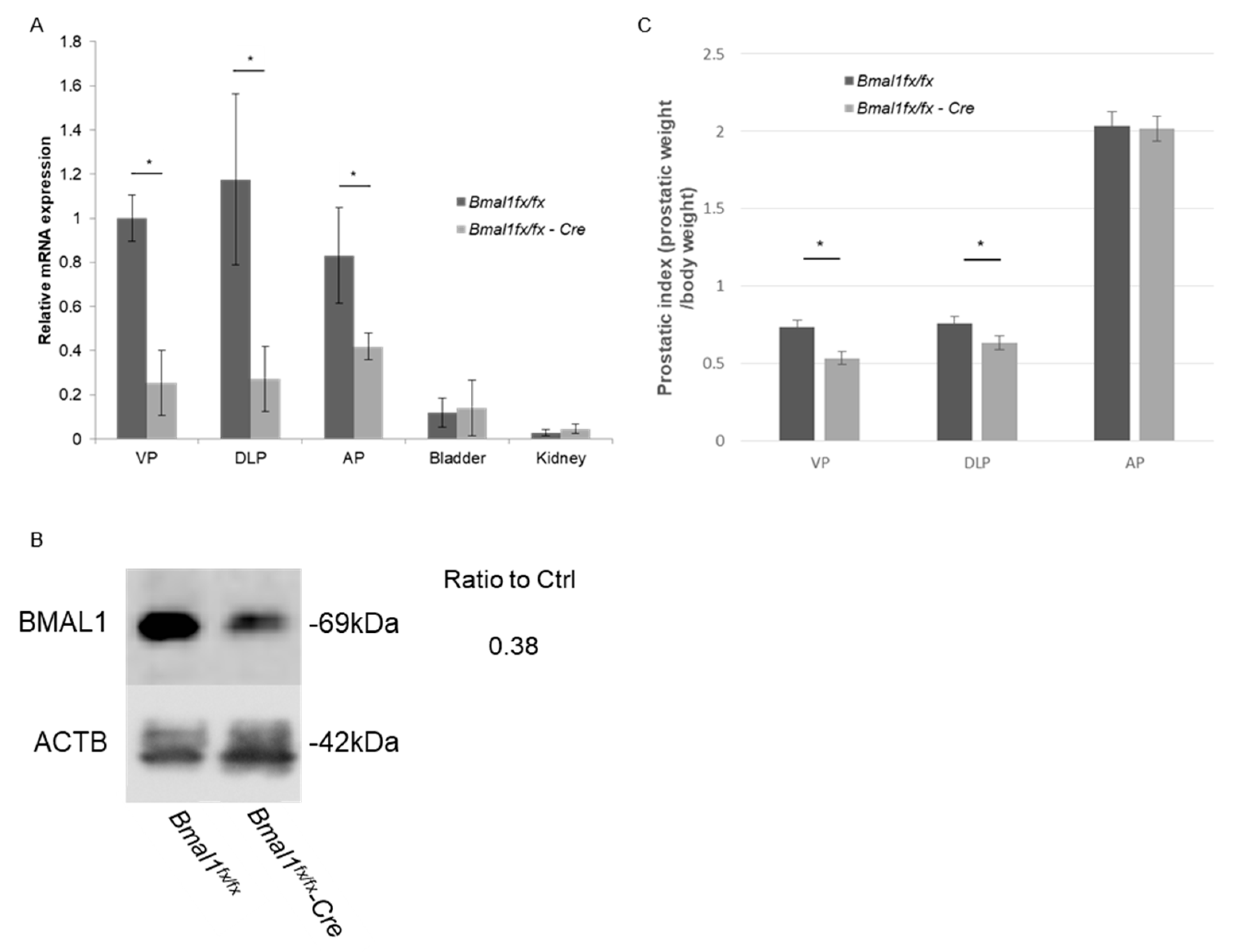
2.1. Prostate-Specific Deletion of Bmal1
2.2. Evaluation of Prostatic Weight in pBmal1 KO Mice
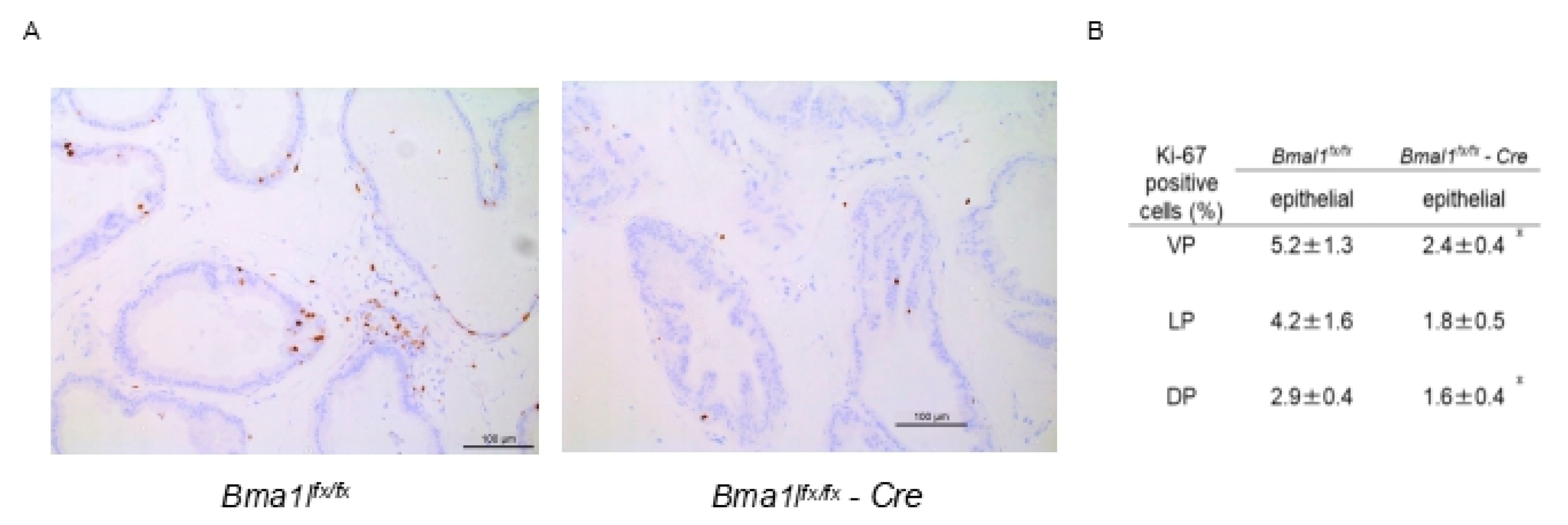
2.3. Immunostaining of pBmal1 KO Mouse Prostates with Ki-67 Antibody
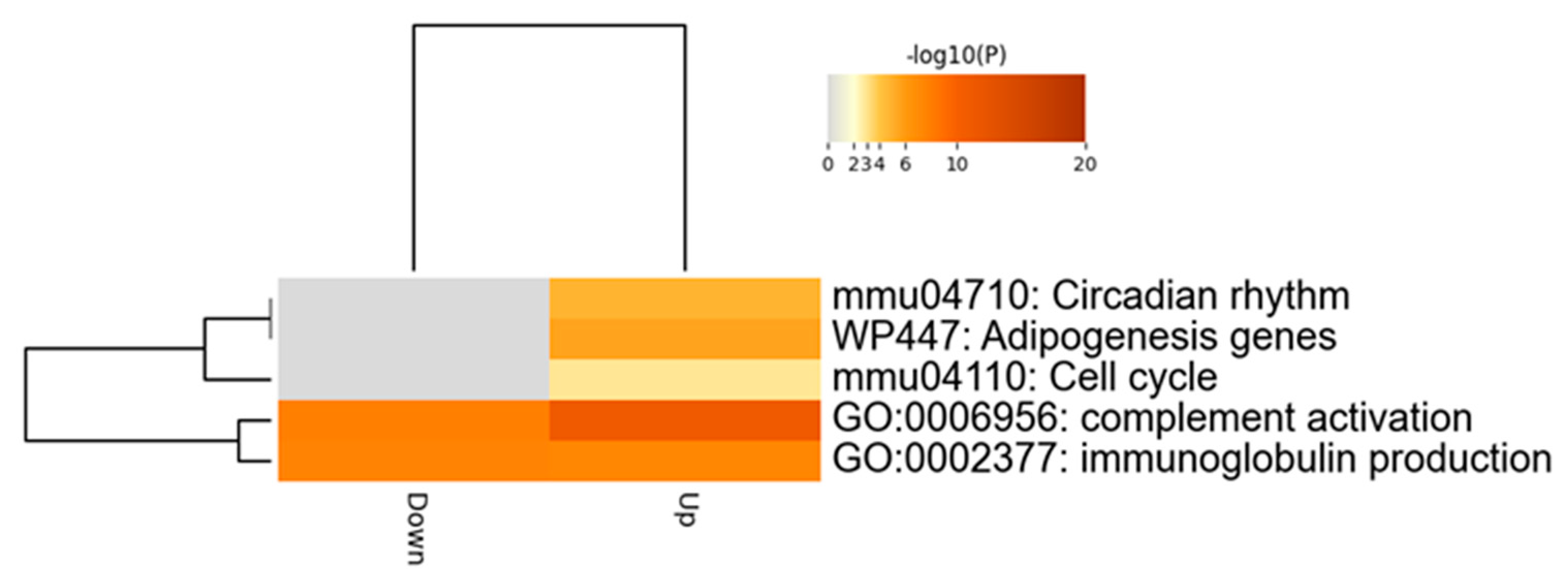
2.4. Comprehensive Analysis of Differently Expressed Genes in pBmal1 KO Mouse Prostates
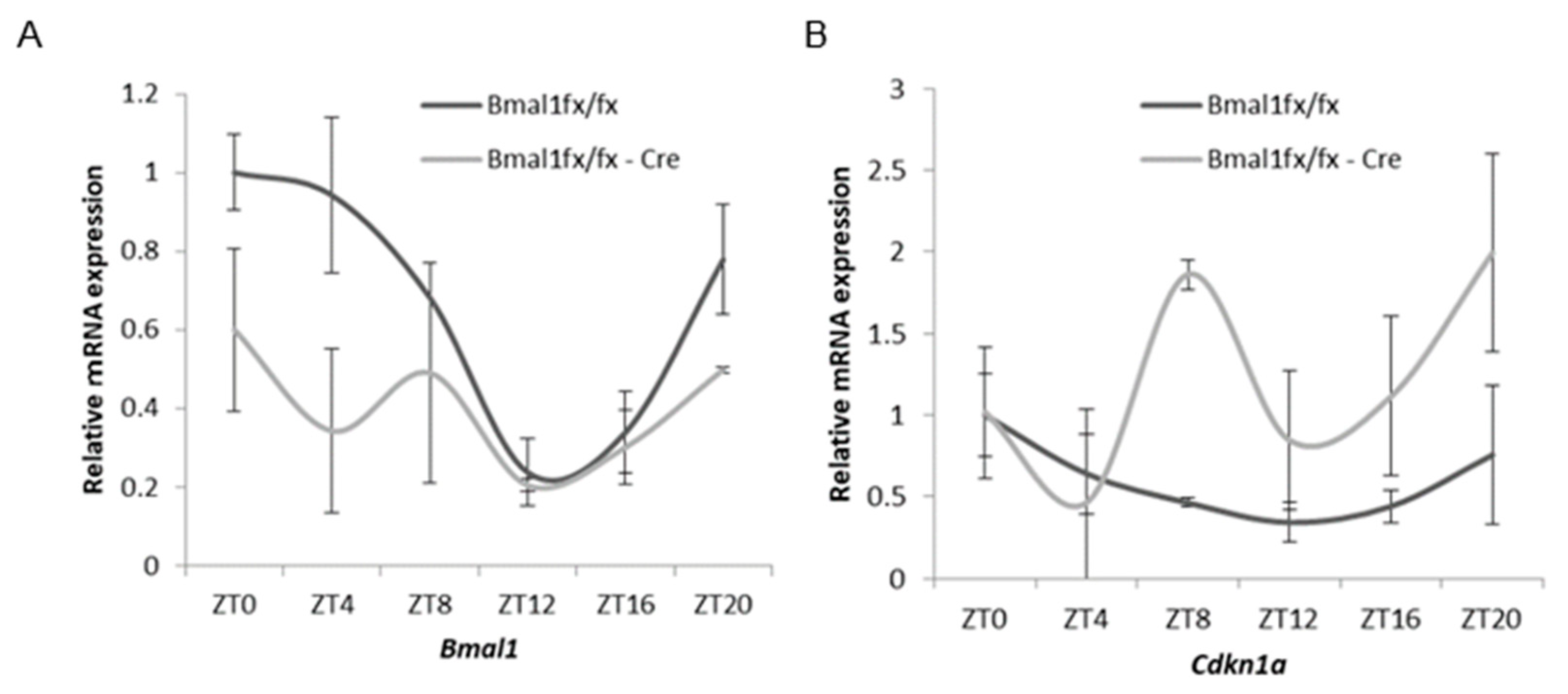
2.5. Oscillation of Cdkn1a and Clock Gene Expression in Mouse Prostates
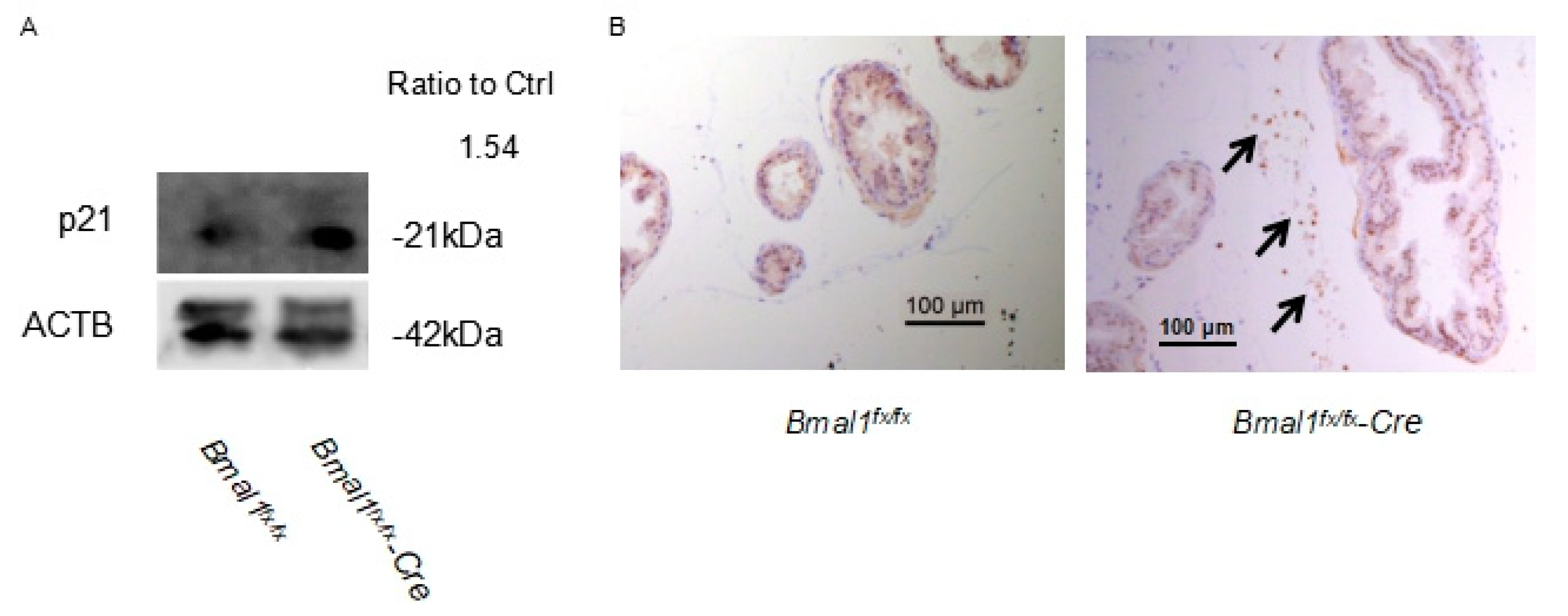
2.6. Expression of p21 in pBmal1 KO Mice
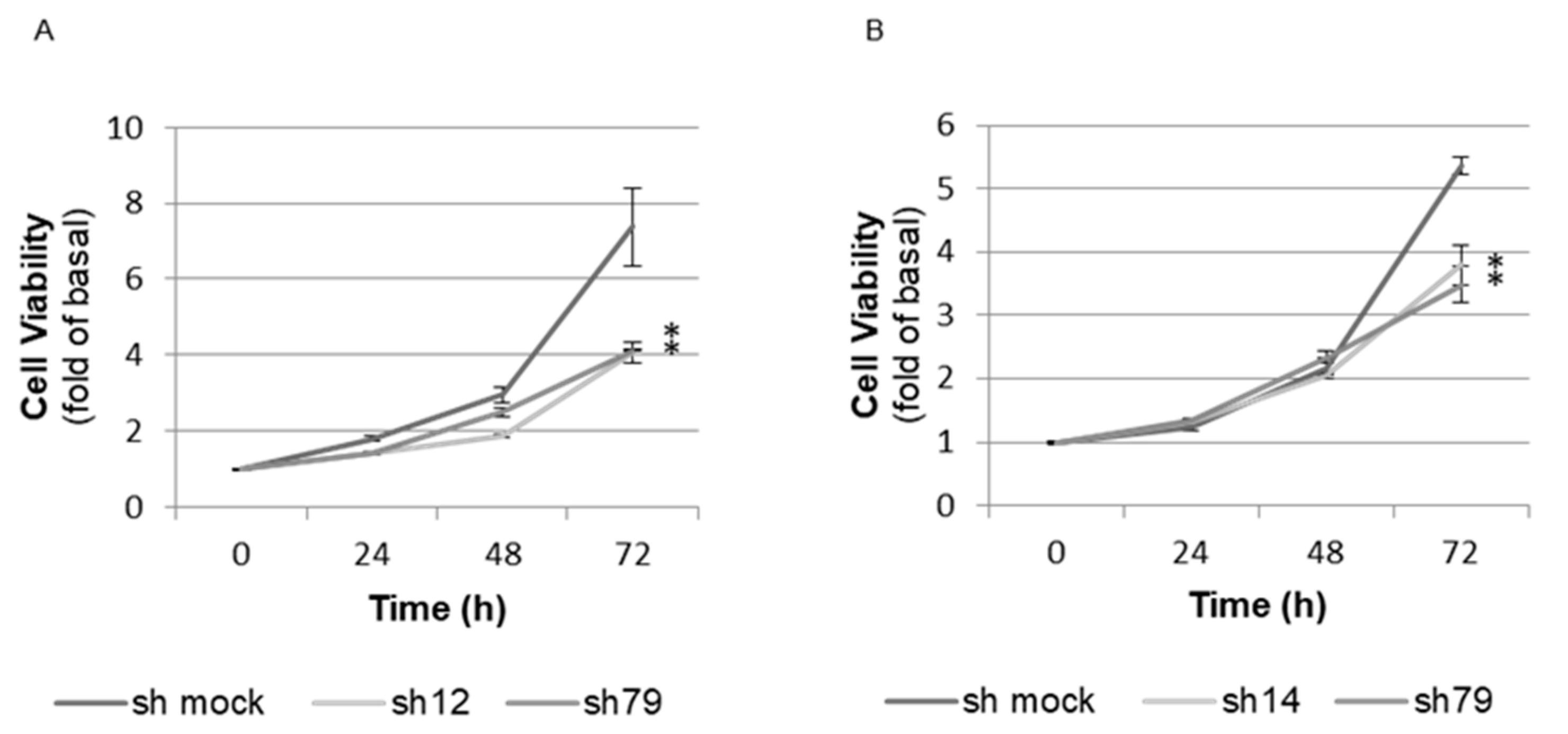
2.7. Proliferation Assay (WST-8 Assay) in BMAL1-Knockdown Immortalized Human Prostate Cells
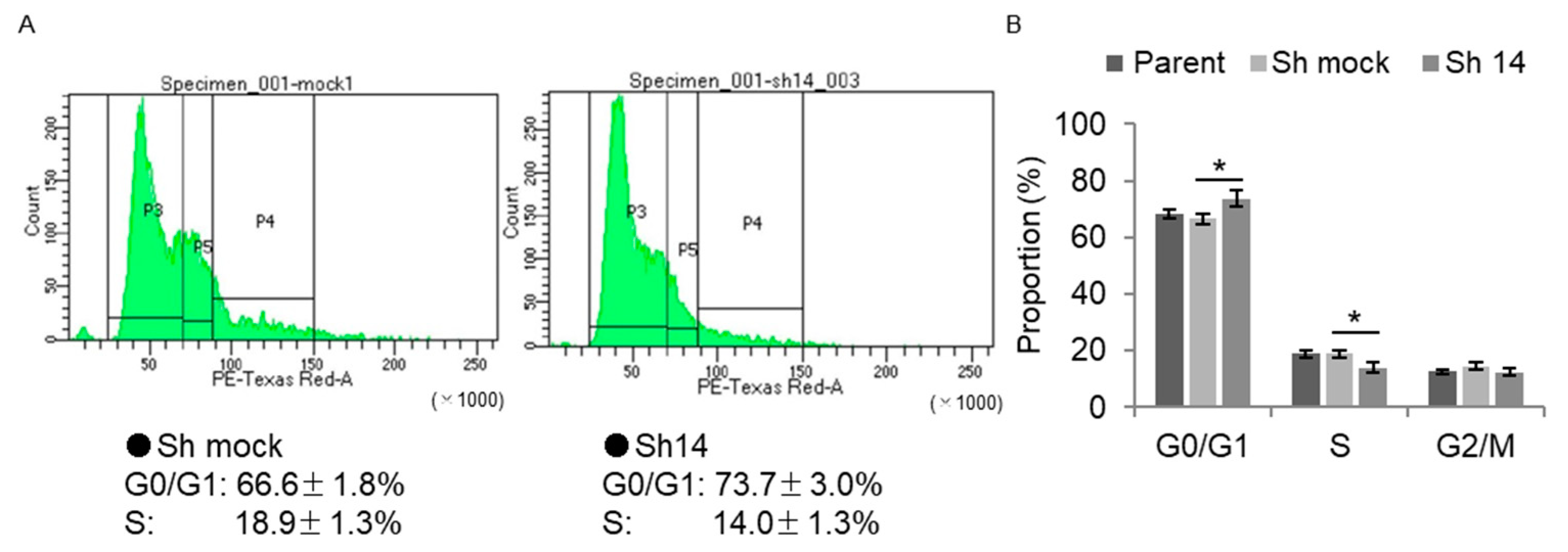
2.8. Cell-Cycle Analysis via Flow Cytometry in BMAL1-Knockdown Immortalized Human Prostate Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Dissection and Weight Measurement of Mouse Prostate Glands
4.3. Histology and Immunohistochemistry of Mouse Prostate Glands
4.4. Cap Analysis of Gene Expression (CAGE)
4.5. Real-Time Quantitative RT-PCR Analysis
4.6. Immunoblotting
4.7. Cell Culture
4.8. Cell Transduction
4.9. Cellular Proliferation Assay
4.10. Flow Cytometry
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Dai, M.; Wang, X.; Jiang, S.H.; Hu, L.P.; Zhang, X.L.; Zang, Z.G. Signaling entrains the peripheral circadian clock. Cell Signal. 2020, 69, 109443. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, S.; Xu, W.; Ying, J.; Qu, Y.; Jiang, X.; Zhang, A.; Yue, Y.; Zhou, R.; Ruan, T.; et al. Critical Roles of the Circadian Transcription Factor BMAL1 in Reproductive Endocrinology and Fertility. Front. Endocrinol. 2022, 13, 818272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; Qian, H.N.; Zhao, Y.; Sun, K.; Wang, H.Q.; Liang, G.Q.; Li, F.H.; Li, Z. Relationship between age and prostate size. Asian J. Androl. 2013, 15, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Elkahwaji, J.E. The role of inflammatory mediators in the development of prostatic hyperplasia and prostate cancer. Res. Rep. Urol. 2012, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Vital, P.; Castro, P.; Ittmann, M. Oxidative stress promotes benign prostatic hyperplasia. Prostate 2016, 76, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Olumi, A.F. Diabetes, growth hormone-insulin-like growth factor pathways and association to benign prostatic hyperplasia. Differentiation 2011, 82, 261–271. [Google Scholar] [CrossRef]
- Alvarez, J.D.; Hansen, A.; Ord, T.; Bebas, P.; Chappell, P.E.; Giebultowicz, J.M.; Williams, C.; Moss, S.; Sehgal, A. The circadian clock protein BMAL1 is necessary for fertility and proper testosterone production in mice. J. Biol. Rhythm. 2008, 23, 26–36. [Google Scholar] [CrossRef]
- Yang, L.; Ma, T.; Zhao, L.; Jiang, H.; Zhang, J.; Liu, D.; Zhang, L.; Wang, X.; Pan, T.; Zhang, H.; et al. Circadian regulation of apolipoprotein gene expression affects testosterone production in mouse testis. Theriogenology 2021, 174, 9–19. [Google Scholar] [CrossRef]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Bebas, P.; Goodall, C.P.; Majewska, M.; Neumann, A.; Giebultowicz, J.M.; Chappell, P.E. Circadian clock and output genes are rhythmically expressed in extratesticular ducts and accessory organs of mice. FASEB J. 2009, 23, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Birbach, A. Use of PB-Cre4 mice for mosaic gene deletion. PLoS ONE 2013, 8, e53501. [Google Scholar] [CrossRef] [PubMed]
- Shimba, S.; Ogawa, T.; Hitosugi, S.; Ichihashi, Y.; Nakadaira, Y.; Kobayashi, M.; Tezuka, M.; Kosuge, Y.; Ishige, K.; Ito, Y.; et al. Deficient of a clock gene, brain and muscle Arnt-like protein-1 (BMAL1), induces dyslipidemia and ectopic fat formation. PLoS ONE 2011, 6, e25231. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Sonoshita, M.; Kakizaki, F.; Aoyama, N.; Itatani, Y.; Uegaki, M.; Sakamoto, H.; Kobayashi, T.; Inoue, T.; Kamba, T.; et al. Amino-terminal enhancer of split gene AES encodes a tumor and metastasis suppressor of prostate cancer. Cancer Sci. 2017, 108, 744–752. [Google Scholar] [CrossRef]
- Gréchez-Cassiau, A.; Rayet, B.; Guillaumond, F.; Teboul, M.; Delaunay, F. The circadian clock component BMAL1 is a critical regulator of p21WAF1/CIP1 expression and hepatocyte proliferation. J. Biol. Chem. 2008, 283, 4535–4542. [Google Scholar] [CrossRef]
- Harper, J.W.; Adami, G.R.; Keyomarsi, N.W.K.; Elledge, S.J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- Shidaifat, F.; Canatan, H.; Kulp, S.K.; Sugimoto, Y.; Zhang, Y.; Brueggemeier, R.W.; Somers, W.J.; Chang, W.Y.; Wang, H.C.; Lin, Y.C. Gossypol arrests human benign prostatic hyperplastic cell growth at G0/G1 phase of the cell cycle. Anticancer Res. 1997, 17, 1003–1009. [Google Scholar]
- Pannek, J.; Berges, R.R.; Sauvageot, J.; Lecksell, K.L.; Epstein, J.I.; Partin, A.W. Cell turnover in human seminal vesicles and the prostate: An immunohistochemical study. Prostate Cancer Prostatic Dis. 1999, 2, 200–203. [Google Scholar] [CrossRef]
- Cao, Q.; Gery, S.; Dashti, A.; Yin, D.; Zhou, Y.; Gu, J.; Koeffler, H.P. A role for the clock gene per1 in prostate cancer. Cancer Res. 2009, 69, 7619–7625. [Google Scholar] [CrossRef]
- Kaur, P.; Mohamed, N.E.; Archer, M.; Figueiro, M.G.; Kyprianou, N. Impact of Circadian Rhythms on the Development and Clinical Management of Genitourinary Cancers. Front. Oncol. 2022, 12, 759153. [Google Scholar] [CrossRef]
- Jin, C.; McKeehan, K.; Wang, F. Transgenic mouse with high Cre recombinase activity in all prostate lobes, seminal vesicle, and ductus deferens. Prostate 2003, 57, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Ohmiya, H.; Vitezic, M.; Frith, M.C.; Itoh, M.; Carninci, P.; Forrest, A.R.; Hayashizaki, Y.; Lassmann, T.; FANTOM Consortium. RECLU: A pipeline to discover reproducible transcriptional start sites and their alternative regulation using capped analysis of gene expression (CAGE). BMC Genom. 2014, 15, 269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulation | Down-Regulation | ||||
|---|---|---|---|---|---|
| Gm16971 | Ighv10-3 | Cyp2b10 | Cfd | Mpz | Igkv8-18 |
| Igkv1-135 | Gm16698 | Gm14017 | Cdkn1a | Fam195b | Gm16710 |
| Gm13253 | Npas2 | Gm16948 | Cry1 | Gbp8 | Gm16700 |
| Gm11755 | Abpb | Iglv2 | Rorc | AC125484.1 | Gm16829 |
| Gm16708 | Igkv15-103 | Igkv12-89 | Iglv1 | Gm16717 | Igkv4-61 |
| Gm16792 | Iglc2 | Thrsp | Gm14326 | Cyp2e1 | Rpl9-ps4 |
| Ccne2 | Ephx2 | Igkv16-104 | Ighv1-62-2 | Igkv4-78 | |
| Retn | Gm16842 | Ifi27l2a | Igkv8-27 | Gm16949 | |
| SNORA19 | Plin1 | Fabp4 | Igkv5-39 | Hp | |
| Gm5417 | Car3 | Pttg1 | Igkv1-110 | Abpz | |
| Igkv1-117 | Adipoq | Prss28 | Zfp959 | Psca | |
| Igkv6-13 | Gm4167 | Gm6644 | Gm10243 | Gm6793 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, M.; Kono, J.; Sengiku, A.; Nagumo, Y.; Mathis, B.J.; Shimba, S.; Taketo, M.M.; Kobayashi, T.; Ogawa, O.; Negoro, H. Bmal1 Regulates Prostate Growth via Cell-Cycle Modulation. Int. J. Mol. Sci. 2022, 23, 11272. https://doi.org/10.3390/ijms231911272
Ueda M, Kono J, Sengiku A, Nagumo Y, Mathis BJ, Shimba S, Taketo MM, Kobayashi T, Ogawa O, Negoro H. Bmal1 Regulates Prostate Growth via Cell-Cycle Modulation. International Journal of Molecular Sciences. 2022; 23(19):11272. https://doi.org/10.3390/ijms231911272
Chicago/Turabian StyleUeda, Masakatsu, Jin Kono, Atsushi Sengiku, Yoshiyuki Nagumo, Bryan J. Mathis, Shigeki Shimba, Makoto Mark Taketo, Takashi Kobayashi, Osamu Ogawa, and Hiromitsu Negoro. 2022. "Bmal1 Regulates Prostate Growth via Cell-Cycle Modulation" International Journal of Molecular Sciences 23, no. 19: 11272. https://doi.org/10.3390/ijms231911272