1. Introduction
Osteogenesis imperfecta (OI) is a rare hereditary bone disease, with a frequency of 1 in 15,000 to 20,000 live births, characterized by phenotypic and genotypic heterogeneity [
1,
2]. The most common symptoms include bone fragility, skeletal deformities, reduced bone mineral density, short stature, blue sclera, dentinogenesis imperfecta, hearing impairment, joint hypermobility, skin fragility, muscle weakness, and cardio-respiratory defects [
1,
2,
3].
Despite recent discoveries of mutations in many genes, the most common are mutations in genes encoding type I collagen, which cause the majority (85%) of cases of OI [
1,
2,
3,
4,
5]. Mutations in the
COL1A1 or
COL1A2 genes encoding the α1(I) and α2(I) chains of type I collagen, respectively, are dominant and cause quantitative or structural disturbances in collagen. The classification of OI due to collagen mutations includes four types, the phenotype of which ranges from mild (type I) and moderate (type IV) to severe (type III) and perinatal lethal in OI type II [
6]. The molecular defect in OI type I is a null
COL1A1 allele due to premature stop codons, either directly or through frame shifts resulting in reduced synthesis of functional collagen type I [
4,
5,
7]. OI types II-IV are mainly caused by substitutions of glycine residues by another amino acid (80%), but also by splicing site mutations as well as small triplet deletion or duplication mutations, which shift the register of α-chains in the helix [
2,
4,
5,
8]. Most of them result in the synthesis of mutant misfolded collagen molecules. In recent years, many other causative genes associated with recessive and X-linked forms of the disease have been detected. Most of these genes code for type I collagen-related proteins that play an important role in folding and post-translational modifications, secretion as well as quality control of collagen synthesis. Mutations of proteins unrelated to collagen type I play an important role in osteoblast maturation and bone mineralization [
2,
3,
5,
8,
9,
10].
Synthesis of type I collagen is a complex process including the intracellular and extracellular steps preceding the formation of mature collagen fibrils. Two proα1 and one proα2 chain are synthesized in the endoplasmic reticulum (ER) and undergo important post-translational modifications prior to triple helix folding, including hydroxylation of proline (at C-4 and C-3) and lysine residues [
11,
12], which have fundamental importance for the stability of the helix. Proper folding and post-translational modifications in the ER determine the effectiveness of collagen secretion to the extracellular matrix (ECM) [
11,
12]. Glycine substitutions are responsible for the delay in the formation of the triple helix. Prolonged exposure of procollagen chains to post-translational modifying enzymes leads to increased hydroxylation of proline and lysine residues and glycosylation of hydroxylysine residues, causing the synthesis of collagen molecules with the abnormal structure [
2,
4,
5,
8,
13]. The mutated collagen is secreted into ECM, but it may be partially retained in the ER causing cellular stress, which may be related to the clinical outcome [
14,
15,
16,
17,
18]. It has been reported that some OI cells activate an unfolded protein response (UPR) to restore cell homeostasis [
14,
17,
18,
19,
20,
21]. The best-studied three ER membrane receptors of UPR include inositol requiring enzyme 1 (IRE1), PKR-like endoplasmic reticulum kinase (PERK), and activating transcription factor 6 (ATF6) [
19,
20,
22]. Under normal conditions, the chaperone—binding immunoglobulin protein (BiP) binds all three sensor proteins in their ER luminal domain and keeps them inactive, while under stressful conditions it binds preferentially to misfolded proteins leading to activation of UPR pathways. ATF6 moves to the Golgi apparatus, where it is cleaved by different proteases and then, as an active transcription factor, enters the nucleus and activates the promoter of its related target genes. IRE1 and PERK are activated by autophosphorylation and oligomerization. Activated IRE1 forms an alternative spliced variant of the X-Box binding protein 1 (XBP1s), which, as a transcription factor, increases the expression of various chaperones and proteins involved in the proteasomal of ER-associated degradation (ERAD). The activation of PERK inhibits global protein synthesis through phosphorylation of eukaryotic translation initiation factor (eIF2α) but favors the translation of some mRNAs, such as the activating transcription factor 4 (ATF4), which is involved in both cell survival and ER stress-dependent apoptosis. During chronic stress, ER promotes apoptosis by upregulating genes such as the homologous protein of the CCAAT enhancer binding protein (CHOP) [
20,
22,
23]. If conformation of mutated collagen is not improved by chaperones, it is destined for degradation most often via autophagy [
24].
Autophagy is a complex system regulated by more than 30 autophagy-related gene (ATG) proteins. The most studied markers associated with autophagy are beclin 1, a microtubule-associated protein 1 light chain 3 (LC3), and sequestosome 1 (SQSTM1/p62), later referred to as p62 [
25]. In some cases, especially the mutations occurring in C-propeptide, that most affect the trimer assembly, the retrotranslocation of misfolded procollagen chains into the cytosol may occur and result in their degradation by the proteasome [
26,
27].
So far, anti-catabolic bisphosphonates, denosumab as a synthetic parathyroid hormone and growth hormone have been used in the therapy of OI [
1,
3,
10]. Experimental OI therapy strategies such as genetically engineered stem cell transplantation methods, reprogramming of somatic cells into pluripotent stem cells as well as anti-transforming growth factor (TGF-β) therapy have been summarized by us recently [
28]. The major disadvantages of these therapies are their poor efficacy, or cytotoxic side effects. Research is still underway to find new and more effective drugs. According to the latest research, the chemical 4-phenylbutyrate chaperone (4-PBA) is of great interest, the molecular target of which is ER stress caused by intracellular retention of mutant collagen in osteoblasts and fibroblasts [
16,
17,
18,
29,
30].
Medicinal plants containing significant amounts of polyphenols are of great interest to researchers due to their high therapeutic potential. Polyphenols are widely studied as regulators of fundamental biological processes, including cell proliferation, apoptosis, and autophagy [
31,
32]. Rosemary (
Rosmarinus officinalis L., Lamiaceae) is a rich source of many bioactive polyphenol compounds with a wide range of biological activities, such as antioxidant, antimicrobial, anti-inflammatory, antidiabetic and anticancer [
33,
34]. In our previous study, we showed a beneficial effect of rosemary extract (RE) on the biosynthesis of type I collagen in OI type I fibroblasts with quantitative type I collagen defect [
35]. This time we examined whether RE could reduce the accumulation of mutant collagen in the fibroblasts of two patients with severe type III and two patients with lethal type II OI carrying mutations in α(I) chain. In addition, possible mechanisms involved in the action of RE were also investigated.
3. Discussion
OI is a genetically and phenotypically heterogeneous group of connective tissue disorders caused mainly by an autosomal dominant mutations in collagen type I, which is the most abundant protein in bone and skin ECM [
1,
2,
3,
4,
5,
6,
7,
8,
9,
10]. The discovery in the last two decades of new causative genes (around twenty) that are involved in the regulation and function of type I collagen, but also in other aspects of bone biology [
2,
3,
5,
8,
9,
10] confirms the complexity of the underlying mechanisms and complicates understanding the relationship between mutations and phenotype as well as finding an effective and a universal method of treating the disease. Recently, ER stress caused by intracellular retention of mutant collagen in osteoblasts and fibroblasts was found as an attractive target of OI therapy [
14,
15,
16,
17,
18,
19,
20,
21]. The use of the chemical chaperone 4-PBA, approved by
Food and Drug Administration, reduced ER stress and restored cell homeostasis in human fibroblasts of OI patients carrying dominant mutations in α1 and α2 collagen type I chains [
17] as well as recessive mutations in cartilage-associated protein
(CRTAP), prolyl-3-hydroxylase 1 (
P3H1) and cyclophilin B (
PPIB) impairing prolyl-3 hydroxylation of collagen type I [
18]. Moreover, it was found that 4-PBA normalizes the overproduction of type I collagen and improves the misfolding of the type I collagen helix in OI fibroblasts due to glycine substitution, and also improves the impaired mineralization of osteoblasts differentiated from OI induced pluripotent stem cells [
30]. Administration of this drug to the OI dominant zebrafish model, carrying typical glycine substitution G574D in
COL1A1, alleviated cellular stress and improved bone mineralization in larvae and skeletal deformity in adults. This was accompanied by the reduction of the ER cisternae size and promoting the secretion of collagen [
29]. Similarly, in osteoblasts of two murine OI models carrying G349C mutation in
COL1A1 (Brtl mouse) and G610C in
COL1A2 (Amish mouse), 4-PBA prevented collagen type I accumulation through increased its secretion and reduction of aggregates in mutant cells [
16]. In addition, increased collagen incorporation into the matrix and improved mineral deposition in osteoblasts was observed in both murine models, which convinces about the influence of ER stress on the phenotype. Therefore, as well as the discovery of the therapeutic potential of this chemical chaperone, finding of other safe compounds to reduce cellular stress and restore cell homeostasis may be a new strategy for treating this disease, or at least some OI cases.
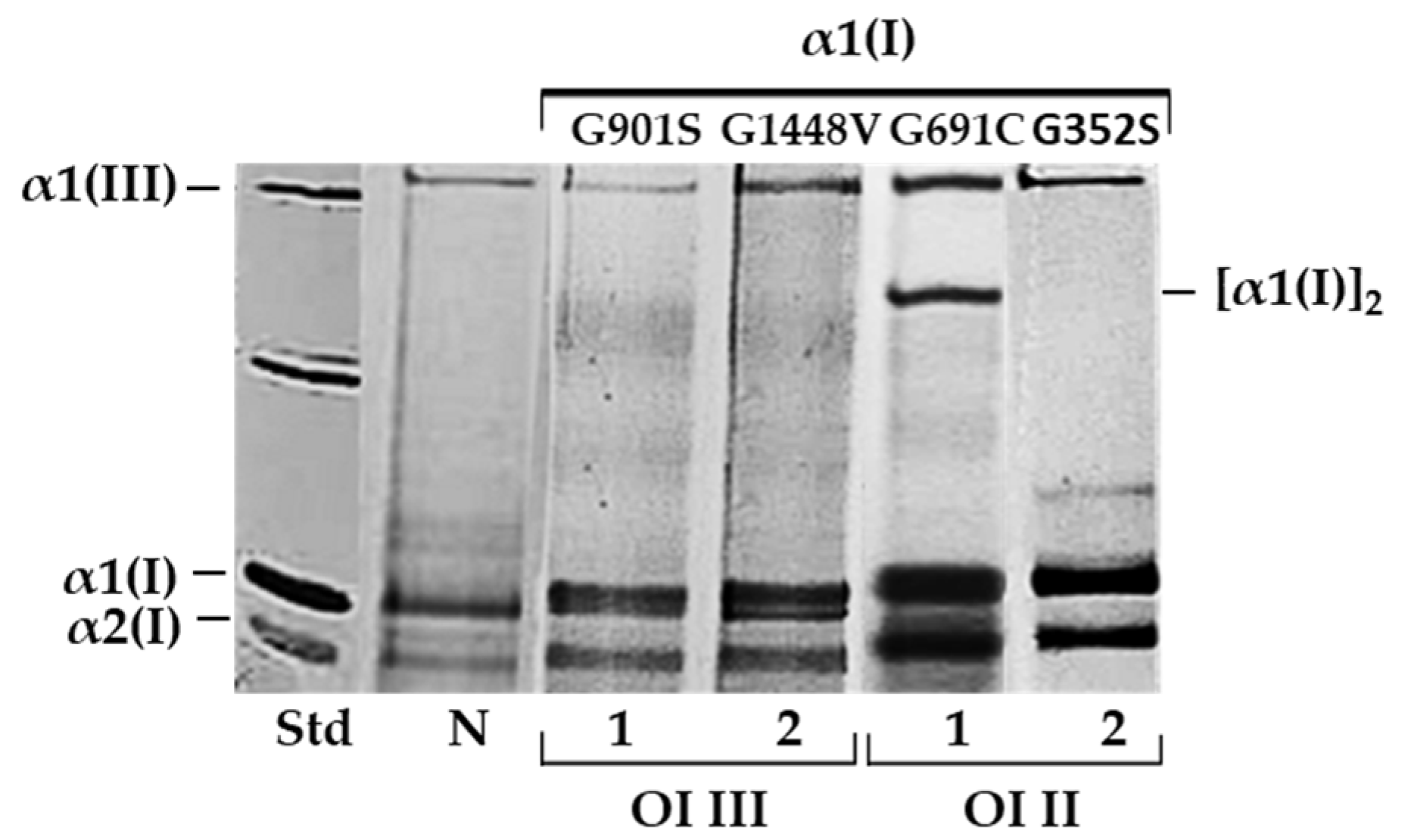
This study provides, for the first time, evidence of the beneficial effects of rosemary extract on fibroblasts with mutations in the collagen triple helix resulting in intracellular collagen accumulation. Moreover, the presented results explain the likely mechanisms of reducing cellular stress, mainly by enhancing the degradation of mutant collagen through autophagy. For our study, we chose glycine substitutions in the α1(I) chains of two patients with severe OI type III (G901S and G1448V), and two with lethal type II (G352S and G691C). As we demonstrated by SDS-urea-PAGE, all cells showed delayed migration of α1 and α2 collagen chains and intracellular accumulation of mutant collagen.
Collagen type I is characterized by a unique right-handed triple helical structure, and each of three left-handed polyproline-like helices contains a repeated sequence (Gly-X-Y) in which X and Y are often proline and hydroxyproline [
36]. The triple helical domain is flanked on both sides by N- and C-terminal propeptides. Procollagen folding takes place in the ER, after which the protein is transported to the Golgi apparatus and secreted, where the cleavage of propeptides takes place and mature collagen is formed. Folding is a very complex process involving many chaperones, such as BiP, PDI, prolyl 4-hydroxylase, various peptidyl-prolyl
cis-
trans isomerases, and heat shock protein 47, whose role is to provide stabilization of the structure and to protect aggregation of unfolded chains [
11,
12,
37].
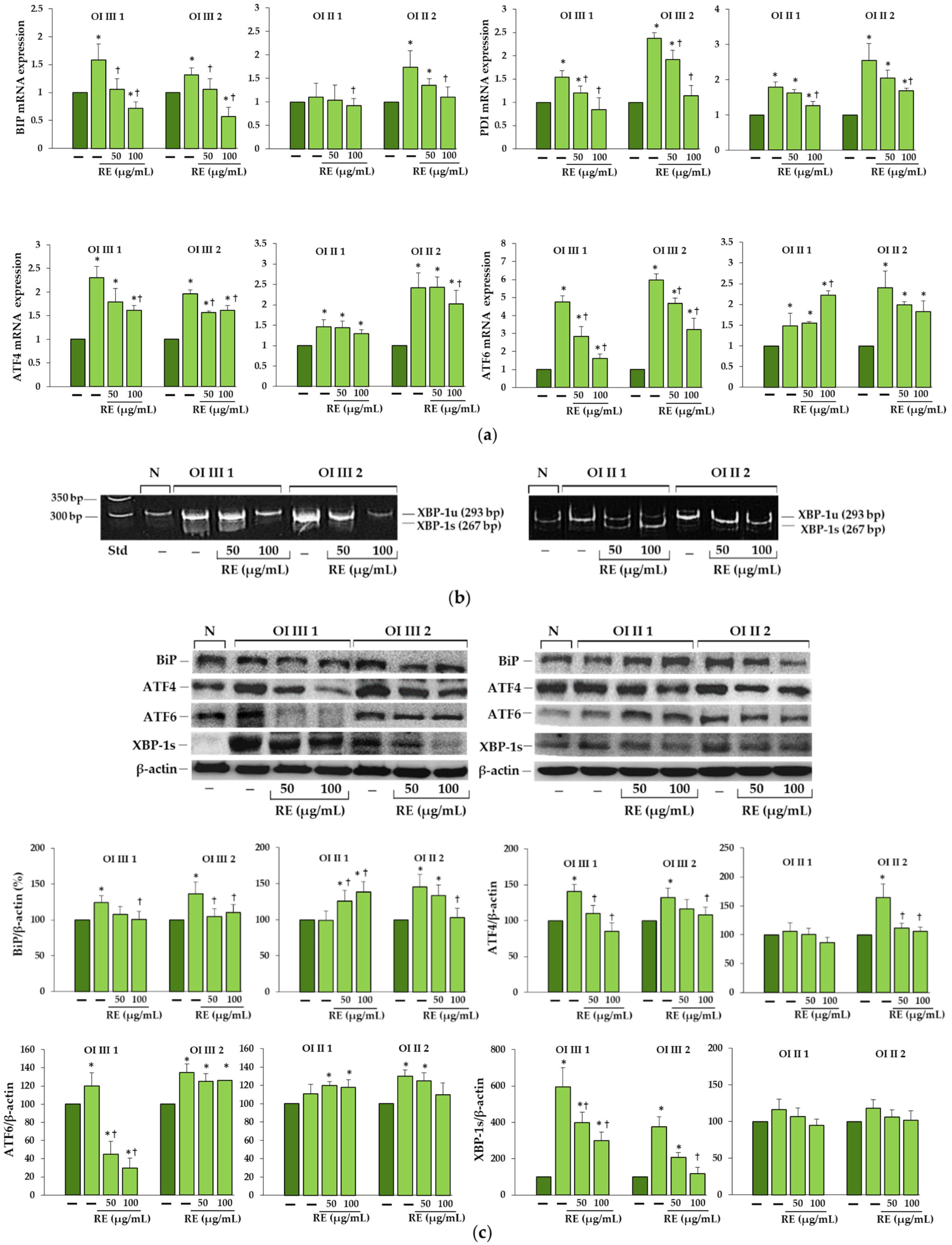
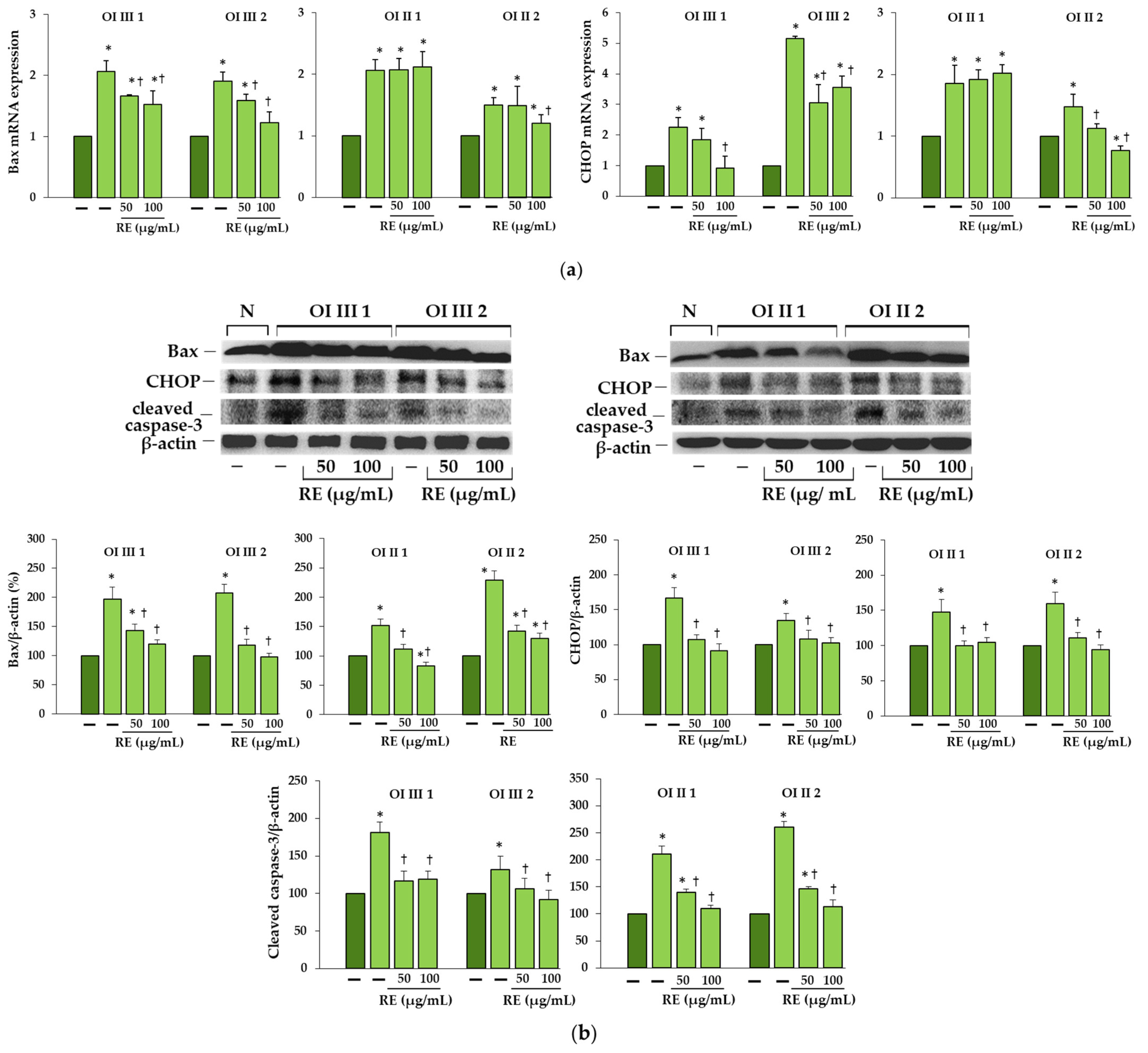
We found increased BiP expression in two OI type III (G901S and G1448V) and one OI type II (G352S) cells, and a consequent activation of UPR pathways, whereas in OI II 1 cells with G691C the expression of this chaperone remained unchanged compared to the normal cells. According to Besio et al. [
17] increased BiP was detected in three out of five tested fibroblasts with glycine substitution in α1 and α2 chains. Expression of PDI, which catalyzes the formation and isomerization of disulfide bonds and acts as a collagen chaperone by interacting with collagen α single chains, was upregulated in all α1(I) mutant cells, while in the study of Besio et al. [
17] in four out five cells with mutations in α1(I) and it remained unchanged in cells with mutations in α2(I). The same authors reported the activation of mainly the PERK pathway and increased ATF4 expression in cells with mutations in α1 and α2 as well as IRE1α pathway with a predominance in cells with mutations in α2, but there was no difference in the level of activated ATF6 [
17]. In contrast, in our study, along with the increase in BiP expression, activation of both transcription factors ATF4 (effector of PERK pathway) and ATF6 in cells with BiP upregulation was found, while expression of spliced forms of XBP1 (effector of IRE1α pathway) was predominant in OI type III as confirmed by real-time PCR and polyacrylamide gel electrophoresis of (reverse transcriptase) RT-PCR product. It should be added that in our studies, the lack of increase in BiP expression in cells with G691C mutation was consistently associated with the lack of activation of UPR proteins, while in the case of some collagen and non-collagen mutations, activation of some UPR pathways was also observed in the absence of BiP [
17,
18]. It is possible that other factors or regulatory mechanisms are involved in the activation of the UPR during ER stress. In the osteoblasts of the mouse OI model with the substitution of glycine by cysteine (G610C), closely located to the one we studied (G691C) but in α2(I), no conventional UPR was detected also, only enhanced autophagy [
15]. Moreover, as it turned out in our studies, the upregulation of genes of transcription factors ATF4 and ATF6 did not coincide with the increased amount of the protein, which means that their expression is regulated by post-transcriptional mechanisms and the expression of the genes themselves cannot be compared without determining the expression at the protein level.
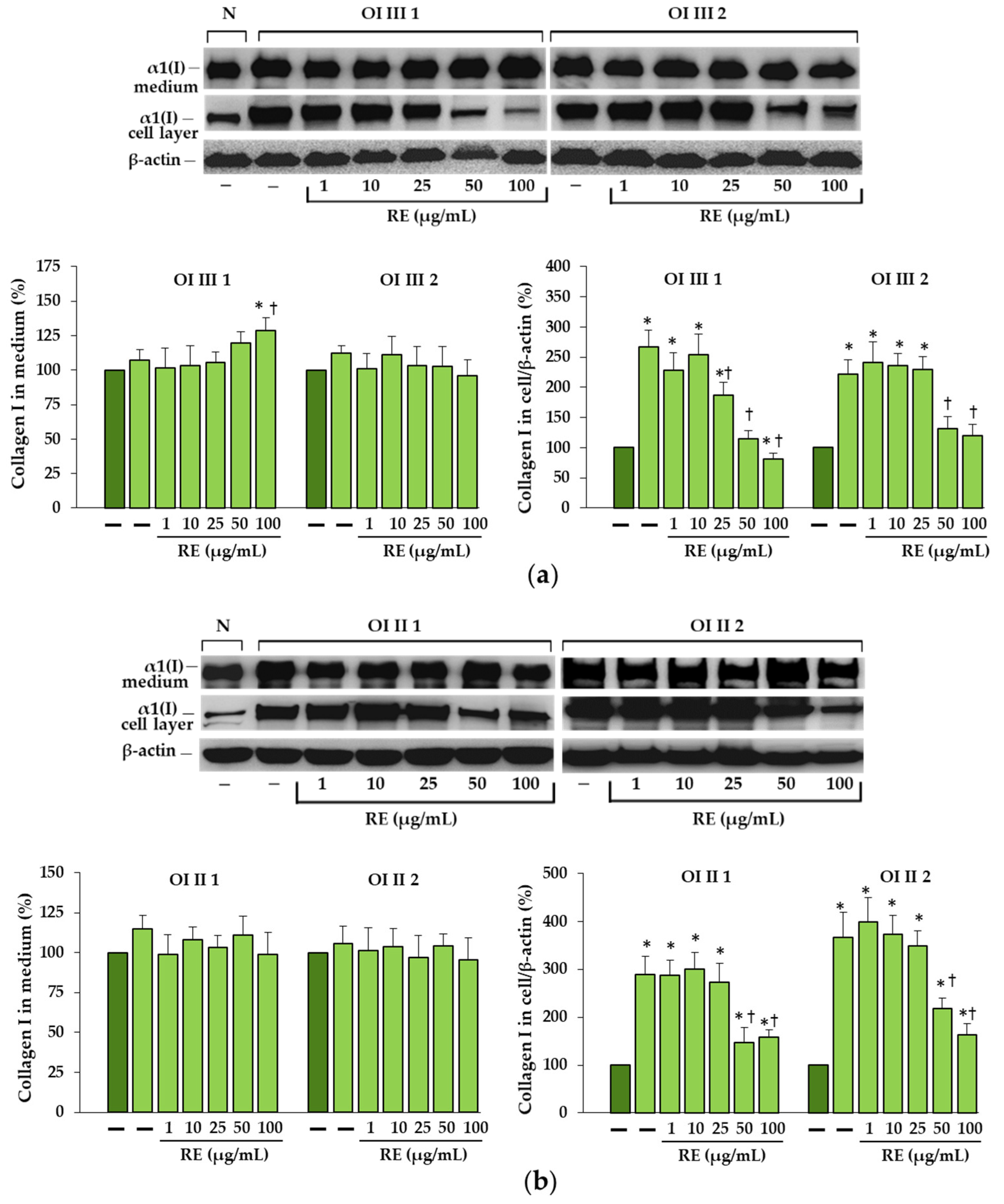
Based on the obtained results, we can say that RE, if not completely eliminated, largely reduced ER stress caused by the accumulation of mutant collagen. This was evidenced by the decreased expression of chaperone proteins BiP and PDI as well as effectors of UPR branches along with the decreased level of intracellular collagen. Interestingly, in OI II 1 cells where we did not detect increased BiP expression and activation of UPR pathways, upregulation of pro-survival factors (BiP, ATF6 and XBP-1s), but not ATF4, was found in RE-treated cells. Activation of the PERK pathway leads to inhibition of global protein translation by inhibition of eIF2α except for ATF4, which in the active form can upregulate both the survival (autophagy) and the apoptotic (CHOP) pathway genes. In turn, the active transcription factors ATF6 a and XBP1s enhance the expression of chaperones, and also ATF6 of genes for proteins involved in ERAD [
19,
20,
22]. While it is still unknown how activation of individual UPR pathways and their effectors directly affects collagen, one study found that forced XBP1s expression in cells with glycine substitution (G425S) in α1 (I) chain enhanced the folding/assembly and secretion of mutant type I collagen [
38].
Since the cell response to ER stress caused by intracellular accumulation of mutant collagen is most often autophagy or, less frequently, ERAD, we investigated the activation of these two degradation systems in untreated and RE exposed OI cells.
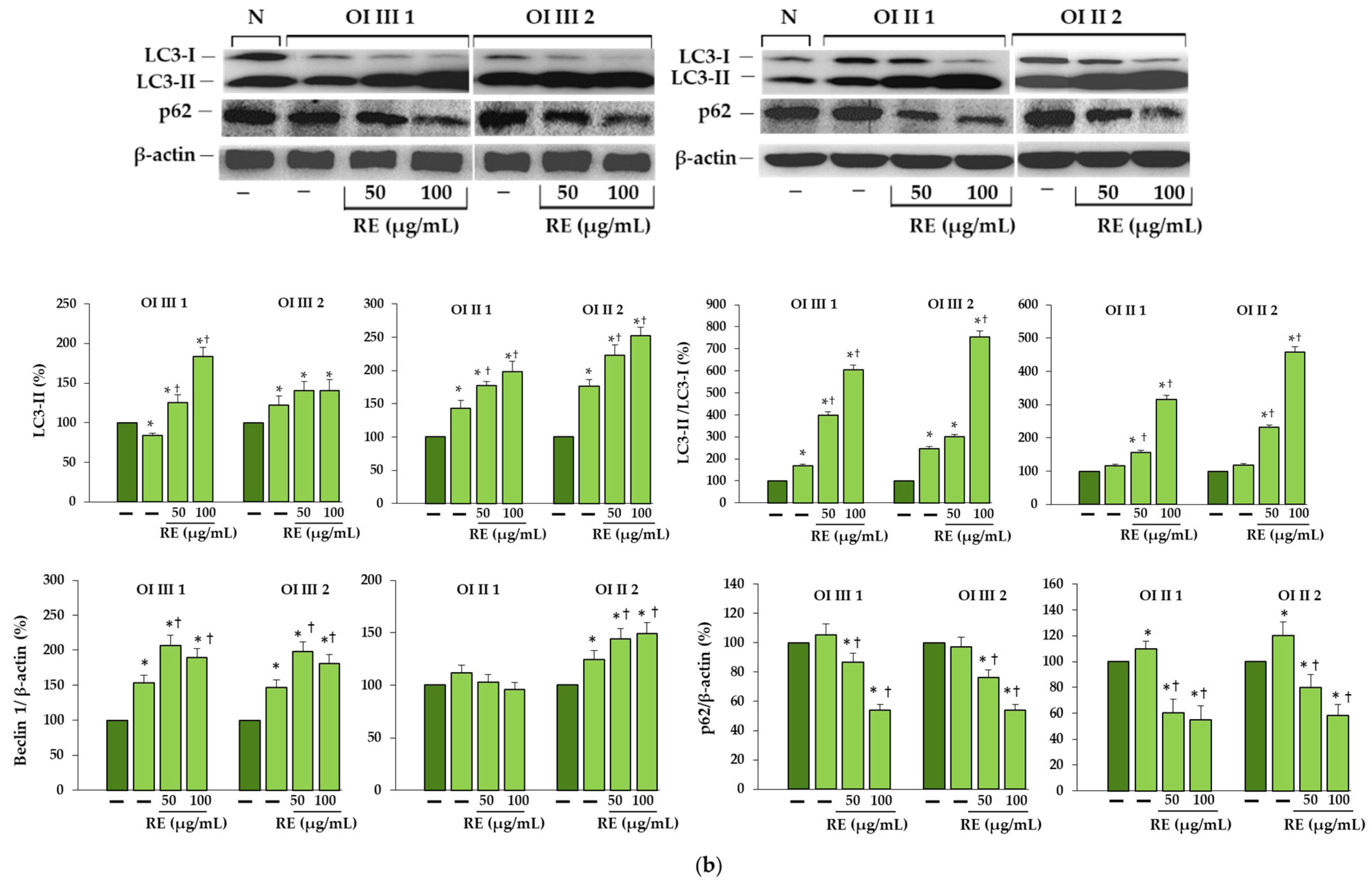
Autophagy, is a dynamic tightly regulated lysosomal pathway of degradation of intracellular components, including soluble proteins, aggregated proteins and damaged cell organelles. It is an evolutionarily conserved process, capable of responding to stress to limit cell damage. The autophagy process is regulated by several ATG core proteins, of which LC3 plays essential role in the formation and maturation of the autophagosome, [
25,
39]. Cytosolic form LC3-I is converted into an active membrane-bound form LC3-II during the formation of the autophagosomes, while the final degradation of the cargo takes place after fusion of autophagosome with lysosomes. It is strictly dependent on the p62, which apart from the ratio LC3-II/LC3I, is an important marker of effective autophagic flux [
40]. Even though the LC3-II level was increasing in lethal untreated cells as compared to the aged-matched control, the ratio LC3-II/LC3-I remained unchanged, which along with the p62 increase indicated a lack of activation of autophagy. OI III cells showed an increase in the LC3-II/LC3-I ratio, but the p62 level remained unchanged, which also did not suggest an increase in autophagic activity. p62 with LC3 recognition sequence binds to LC3-II and after the formation of the autophagosome and its fusion with the lysosome, is degraded inside the autophagolysosome, that is why the decrease in p62 expression may indicate an active process of autophagy.
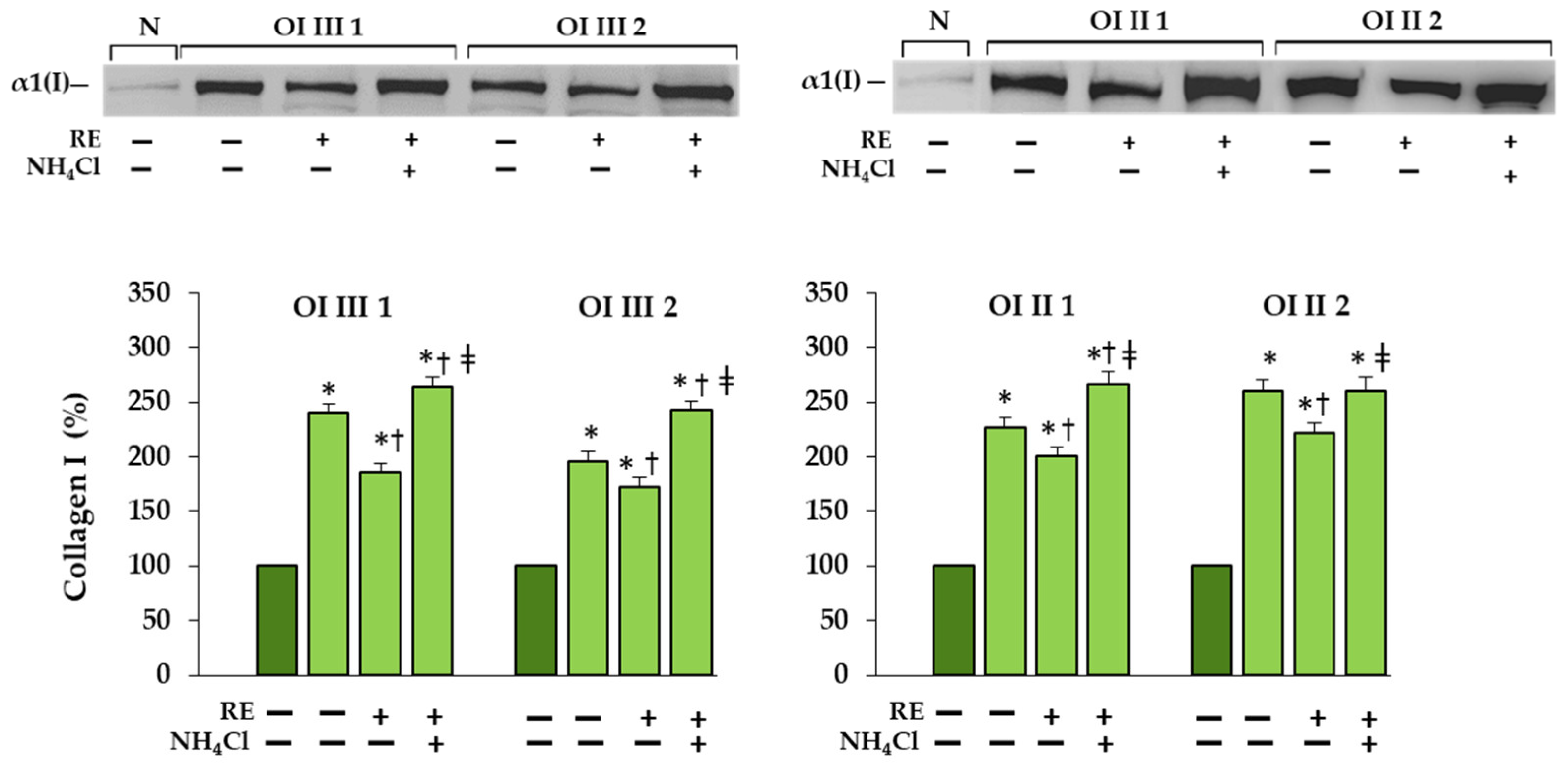
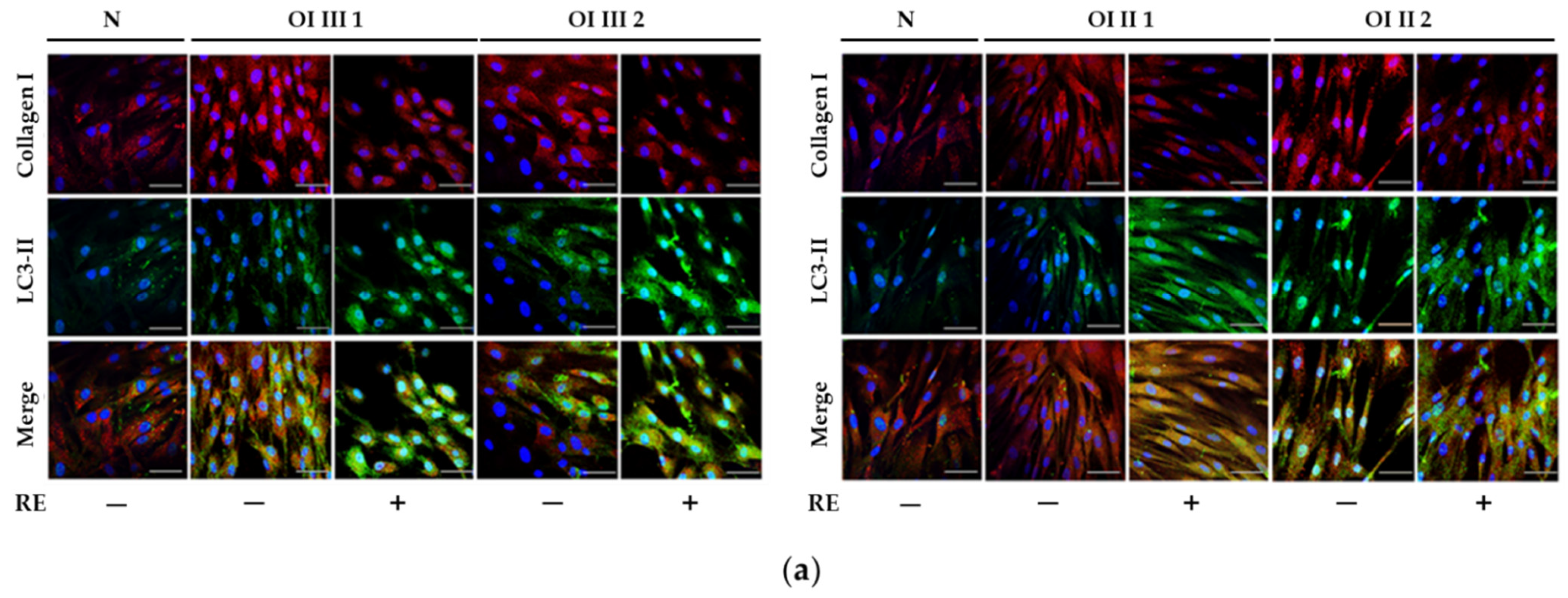
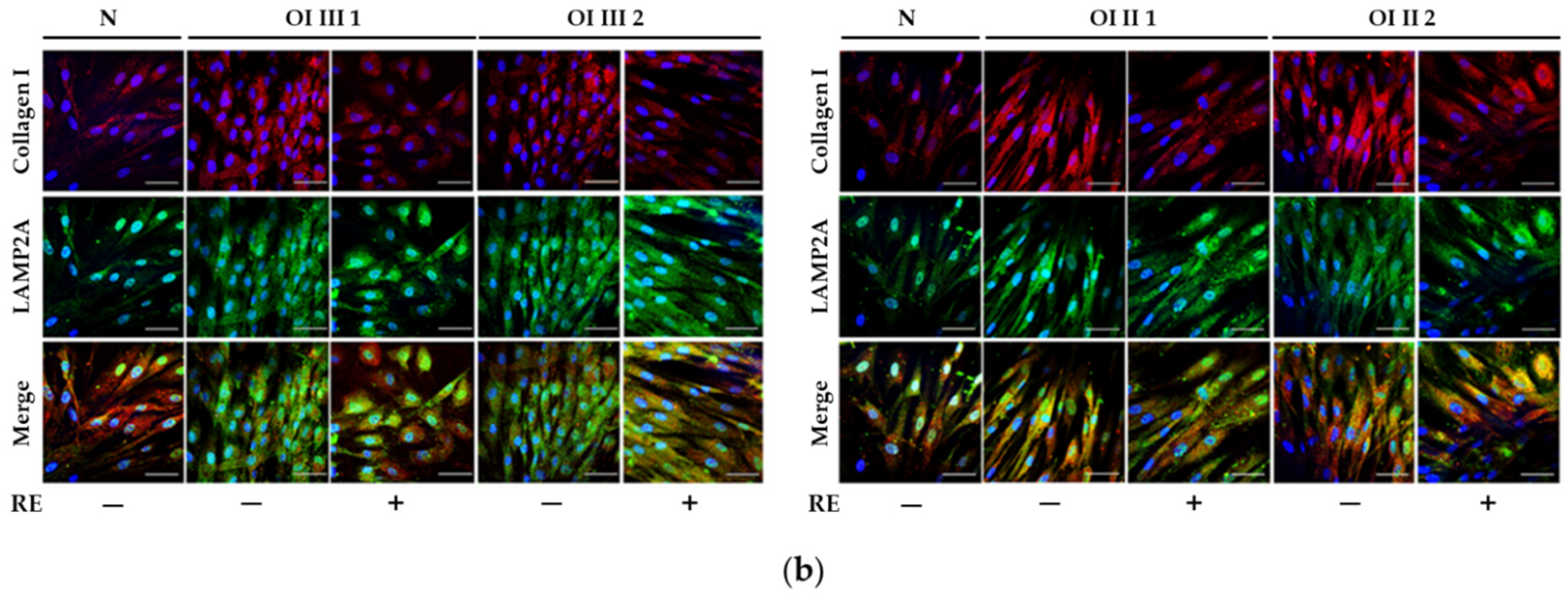
A markedly increased autophagic activity was observed in all OI cells after RE exposure, as shown by a dose-dependent increase in LC3-I to LC3-II conversion, along with accelerating p62 degradation. In addition, the stimulation of ATG5 mRNA and beclin 1 mRNA and protein, which initiate autophagosome formation, was demonstrated in RE-treated cells. Another evidence of the stimulating effect of RE on the degradation of mutant collagen with involvement of lysosomal pathway was the confirmation of the presence of collagen type I in the lysosomal fraction and the increase in its level in cells treated additionally with ammonium chloride, which raises the pH and thus inactivates lysosomes. The same results were obtained in the presence of chloroquine—another autophagy inhibitor (not shown). Moreover, the immunofluorescence microscopy studies have showed that collagen type I collocation increase in RE-treated OI cells with both the marker of autophagosome (LC3-II) and marker of lysosomes (LAMP2A). These results clearly indicate the RE-mediated degradation of mutated collagen type I in the autophagolysosomal process, although digestion of collagen by lysosomes regardless of autophagy cannot be ruled out. Omari et al. [
41] showed that in addition to autophagy, accumulated in OI collagen type I can be digested in a noncanonical autophagy process.
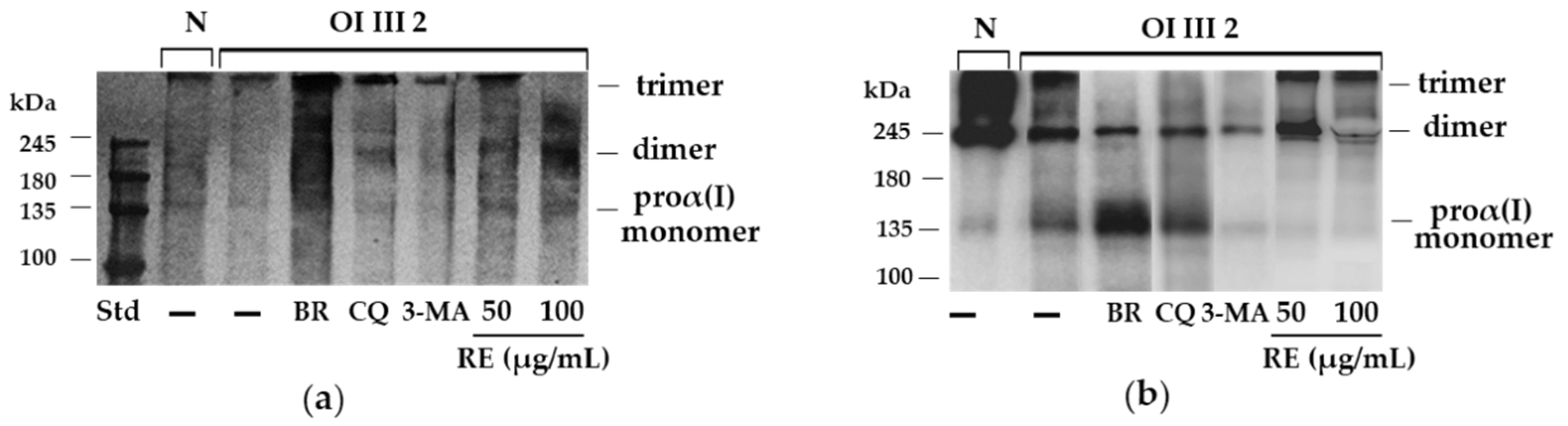
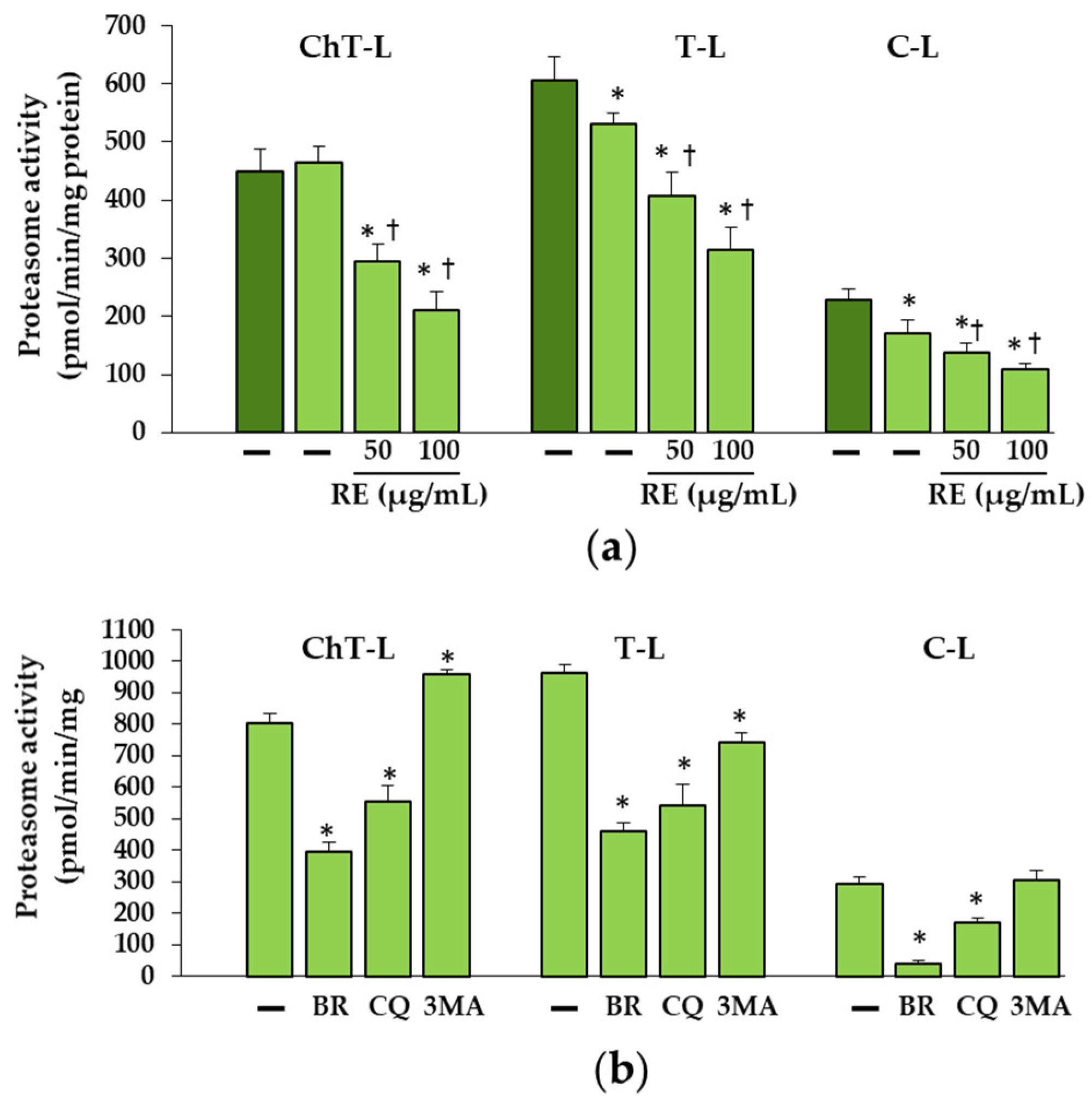
As reported earlier the proteasome may be involved in the removal of unfolded procollagen chains [
27]. Misfolded proteins or unfolded procollagen chains are retranslocated from the ER to the cytosol for degradation by the 26S proteasome after modification with polyubiquitin chains. As expected, polyubiquitination of unfolded proα1(I) chains was demonstrated in OI III 2 with a C-propeptide mutation (G1448V). The increase in the amount of unfolded procollagen chains in untreated OI cells can be explained by decreased C-L and T-L proteasome activities. The greater accumulation of these chains in the presence of BR, which inhibited to a much greater extent all three activities (ChT-L, T-L and C-L) of the proteasome confirms the proteasome’s contribution to the removal of these chains. However, while it turned out that RE also decreased ChT-L, T-L and C-L activities, this decrease did not coincide with the accumulation of unfolded chains. On the contrary, there were lower levels of them than in untreated cells. At this stage of the study, it is difficult to explain the mechanism of action of RE in these cells, but it is very likely that in the case of inhibition of the proteasome activity, unfolded chains may be partially degraded in the process of RE-activated autophagy or by the proteasome, as the activity of the proteasome was only partially inhibited. It was also noted that, despite the reduction in proteasome activity by CQ, which inhibits autophagy, there was no additional accumulation of unfolded chains. On the other hand, another inhibitor of autophagy 3-MA that block autophagy at the initiation and maturation stages by acting on phosphoinositol 3 phosphate kinase (PI3K), caused a decrease in the level of unfolded chains compared to untreated OI cells, perhaps due to its stimulating effect on ChT-L activity. Since the two protein degradation systems (proteasome and autophagy) appear to be mechanically linked, it is suspected that when the proteasome is inhibited, autophagy may be activated to remove polyubiquitinated/unfolded protein aggregates and promote cell survival [
42]. It is possible that a decrease in proteasome activity, also noted in other RE-treated OI cells used in this study (results not shown), triggers autophagy, protecting cells from the toxic long-term stress leading to cell apoptosis. The increase in the expression of pro-apoptotic proteins (Bax, CHOP and active caspase 3) in OI cells and their significant reduction or even normalization in the presence of RE, may unequivocally indicate that the accumulation of mutant type I collagen caused such stress and despite the activation of UPR (with the exception of OI II), the degradation processes mediated by autophagy and the proteasome were disturbed. The cell’s response to inhibited intracellular degradation processes could be a significant upregulation of extracellular MMP-1 and MMP-2 genes, lowered in the presence of RE, while the importance of these upregulation requires further study. Moreover, the observed increase in the level of extracellular type I collagen only in the presence of 100 µg/mL RE in OI III 1 can be explained by its strong inhibitory effect on the activity of MMP-2 and not by an increase in collagen secretion. Negative correlations between the activity of lysosomal enzymes and MMPs have been reported [
43] and are worth studying further, but our research focuses on the intracellular degradation processes.
It is also possible that, under the influence of RE, collagen folding is improved due to reduced over-modification of free procollagen chains as a RE-induced decrease in mRNA expression of one of the enzymes (β(1-O) galactosyltransferase (GLT25D1)) involved in these modifications was observed (data not shown).
Phytochemicals, due to their natural origin, low toxicity, as well as many valuable biological and pharmacological properties, are of wide interest among researchers as potential drugs with a high therapeutic and preventive potential for many diseases.
R. officinalis is a polyphenol-rich source constituents such as luteolin and apigenin derivatives, caffeic acid derivative (rosmarinic acid) and other such as diterpenes (rosmanol isomers), detailed qualitative analysis of which was presented earlier [
35].
In our previous studies, we have shown a stimulatory effect of rosemary and lemon balm extracts, RA as well as some flavonoids on collagen biosynthesis in OI type I [
35,
44] as well as normal human skin fibroblasts [
45,
46,
47]. In these studies, the use of RA alone in a wide range of concentrations did not bring the expected effects, which may suggest the participation of other components of the rosemary extract in stimulating autophagy or its other effects. It has been reported by other authors that luteolin 7-
O-glucoside (one of the identified RE components) protects against damage to the heart muscle induced by starvation by enhancing autophagy through inhibition of mechanistic target of rapamycin (mTOR) and extracellular signal-regulated kinase (ERK) signaling pathway [
48]. It has also been shown that apigenin increases the expression of LC3-II, the formation of autophagolysosomal vacuoles and triggers autophagic flow in hepatocellular carcinoma cells [
49]. It is widely accepted that a variety of plant extracts and dietary phytochemicals including resveratrol, curcumin, epigallocatechin-3-gallate, punicalagin, oleuropein, myricetin and rosmarinic, norhydroguaiaretic, and ferulic acids may stimulate autophagy [
31,
50]. These compounds remove protein aggregates, stimulate the antioxidant defense and ameliorate the ER stress, resulting in increased cell survival [
31,
32,
33,
34,
50]. Pierzynowska et al. [
51] reported on the removal of mutant huntingtin aggregates in the transfected HEK293 cells via genistein-induced autophagy, which may be the basis for the development of an effective therapy for this inherited neurodegenerative disease. Interestingly, like RE in OI cells, genistein showed significant inhibition of all protesome activities in the fibroblasts of patients with all types of mucopolysaccharidosis, which according to the authors, may lead to the stabilization of lysosomal enzymes and constitute a new approach in the treatment of this genetic disease [
52].
A synergistic effect of several different compounds present in the rosemary extract may also be likely. While the biological properties (e.g., antioxidant) of polyphenols were previously related mainly to the structure of these compounds, now a more convincing explanation is modulating the activity and/or expression of key proteins for signaling cascades by interacting with them or modulating epigenetic regulation of gene expression. Therefore, it is believed that pleiotropic mechanisms and specific polyphenol-protein interactions are involved in their beneficial effects [
53,
54].
Finally, the limitations of this study should also be mentioned. Firstly, experiments were conducted on fibroblasts, which is related to the availability of biological material, and the disease mainly affects the skeletal system. On the other hand, collagen type I is a major component of skin and bone and, with a few exceptions, is similarly expressed in fibroblasts and osteoblasts. It is also worth noting that the activity of the proteasome may vary with age and even tissues or may be different in OI patients; therefore, more detailed studies are needed to understand the molecular mechanisms of RE-induced changes in proteasome activity. Although at this stage of our research we did not focus on explaining the consequences of proteasome inhibition by RE, the lack of accumulation of non-collagen proteins was shown.
4. Materials and Methods
4.1. Chemicals
Dulbecco’s minimal essential medium (DMEM), fetal bovine serum (FBS) and phosphate-buffered saline (PBS) were obtained from Gibco (Thermo Fisher Scientific, Waltham, MA, USA); penicillin, streptomycin, and glutamine were purchased from Quality Biologicals Inc. (Gaithersburg, MD, USA). Radioimmunoprecipitation assay (RIPA) buffer, protease inhibitor cocktail (P8340), magnesium L-ascorbate, sodium dodecyl sulfate (SDS), dimethyl sulfoxide (DMSO), [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (MTT), bovine serum albumin (BSA), pepsin, gelatin, NH
4Cl, CQ, 3-MA, and MG132 were provided by Sigma-Aldrich Corp. (St. Louis, MO, USA). BR was a product of Selleck Chemicals (Houston, TX, USA). Proteasome substrates: N-Suc-LLVY-AMC (7-amido-4-methylcoumarin) was purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA), Bz-VGR-AMC and Z-LLE-AMC were obtained from Enzo Life Sciences, Inc. (Farmingdale, NY, USA). Rosemary extract (RE) was prepared and characterized using LC-MS technique according to procedure described in our previous study [
35]. RA was a product of BIOKOM (Warsaw, Poland).
4.2. Fibroblast Culture and Treatment
The study was performed on skin fibroblasts derived from two patients with severe OI type III and mutations in COL1A1: Gly901Ser (patient 1) and Gly1448Val (patient 2), and two patients with lethal OI type II and mutations in COL1A1: Gly691Cys (patient 1) and Gly352Ser (patient 2) as well as two age matched normal cells. The normal skin fibroblast line used as a control for OI type III was CRL-1474 obtained from American Type Culture Collection (Manassas, VA, USA), and as a control for OI type II the normal line was derived from the foreskin on the 7th day of life of the donor. Fibroblasts from skin biopsy of OI patients and healthy control were obtained after informed consent in accordance with the Declaration of Helsinki and was approved by Bioethical Committee of the Jagiellonian University in Kraków, Poland (KBET/108/B/2007).
Fibroblasts were cultured in DMEM supplemented with 10% FBS, 2 mM glutamine, penicillin (50 U/mL) and streptomycin (50 µg/mL) at 37 °C in a humidified incubator in atmosphere containing 5% CO2. For experiments, fibroblasts were grown to 90% confluence and the cultured medium was replaced with fresh DMEM without serum, supplemented with 25 µg/mL of magnesium ascorbate, before addition of compounds. Compounds were stored at 4 °C as the concentrated stock solutions in DMSO and were diluted in medium prior to addition to cell cultures. Fibroblasts were treated with RE at the concentration of 1–100 µg/mL and RA at the concentrations of 1–100 µM for 24 h. In addition, cells were treated with autophagy inhibitors: 50 μM CQ, 50 mM NH4Cl and 5 mM 3-MA or proteasome inhibitors: 50 nM BR and 2.5 µM MG132, all of which were dissolved in DMSO and appropriately diluted before adding to cell cultures. In all experiments the concentration of DMSO did not exceed 0.05% (v/v).
4.3. MTT Test to Determine Viability of Treated Cells
Fibroblasts (1 × 104 cells per well) were treated with RE (1–200 µg/mL) for 24 h. Then cells were washed three times with PBS and MTT solution (0.5 mg/mL) was added for 4 h. After removing MTT solution 1 mL of 0.1 M HCl in absolute isopropanol was added to dissolve formazan crystals by thoroughly shaking on a plate shaker (BioSan, Riga, Latvia), and the absorbance at 570 nm was measured using a microplate reader (TECAN, Männedorf, Switzerland).
4.4. Quantitative Real-Time PCR
Total RNA was isolated from cultured cells using a Total RNA Mini Plus concentrator (A&A Biotechnology, Gdynia, Poland) and the concentration of RNA was determined using NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The equal amounts (1 µg) of total RNA were used to the synthesis of complementary DNA (cDNA) with the use of cDNA Synthesis Kit (Bioline, London, UK). Quantitative Real-time PCR (qRT-PCR) analysis was performed in the CFX96 Real-Time System thermal cycler (Bio-Rad, Hercules, CA, USA) using the SensiFAST™ SYBR kit (Bioline, London, UK). The expression of desired gene was normalized to the level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and changes were calculated by the ΔΔCt method. The sequences of primers (Genomed, Warsaw, Poland) are shown in
Supplementary Table S1. The qRT-PCR parameters were as follows: 30 s at 95 °C followed by 40 cycles: 10 s at 95 °C, 10 s at 60–62 °C and 20 s at 72 °C. The reaction products were verified by analysis of their melting curves.
4.5. XBP1 Splicing Analysis
PCR mixture contained 1 µg of isolated RNA and primers (0.3 µM each): sense (5′-TCAG CTT TTA CGA GAG AAA ACT CAT GGC CT-3′) and antisense (5′-AGA ACA TGT GTG TCG TCC AAG TGT GTC GTC CAA GTG TG-3′) purchased in Genomed (Warsaw, Poland). Samples were incubated 30 min at 50 °C followed by 30 cycles at 94 °C, 60 °C, and 72 °C for 30 s each in the CFX96 Real-Time System thermal cycler (Bio-Rad, Hercules, CA, USA). Reaction products were analyzed by electrophoresis on 7% polyacrylamide gel and visualized with ethidium bromide.
4.6. Western Blot
Cell layers were harvested using RIPA buffer (Sigma-Aldrich Corp., St. Louis, MO, USA) and protease inhibitor cocktail (P8340) (Sigma-Aldrich Corp., St. Louis, MO, USA). The conditioned media were collected and concentrated 10 times with Centrifugal Filter Units (10K) (Merck Millipore Ltd., Carrigtwohill, County Cork, Ireland). The concentration of total protein in cell lysates and media was measured using BCA Protein Assay Kit (Pierce, Rockford, IL, USA) and Coomassie Plus—The Better Bradford Assay Reagent (ThermoFisher Scientific, Rockford, IL, USA), respectively. For Western blot an equal amount of protein (20 µg) was loaded on polyacrylamide gel (7.5%, 10% or 12% depending on the molecular mass of protein). Proteins were transferred from gels onto Immobilon-P Transfer membranes (Merck Millipore Ltd., Tullagreen, Carrigtwohill, County Cork, Ireland), which were blocked with 5% (w/v) non-fat dried milk diluted in 50 mM Tris-HCl, pH 7.5, 500 mM NaCl, 0.05% (v/v) Tween 20 (TBS-T) for 1 h at room temperature. Then, membranes were washed with TBS-T and incubated overnight at 4 °C with solutions of the following primary monoclonal antibodies: mouse anti-collagen type I (1:1000; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), rabbit anti-ATF4 (1:1000; Abcam, Cambridge, UK), rabbit anti-ATF6 (1.1000; Abcam, Cambridge, UK), rabbit Bax (1:1000; Cell Signaling Technology, Danvers, MA, USA), mouse Beclin-1 (1:1000; Cell Signaling Technology, Danvers, MA, USA), rabbit BiP (1:1000; Cell Signaling Technology, Danvers, MA, USA), mouse cleaved caspase-3 (1:1000; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), mouse CHOP (1:1000, Cell Signaling Technology, Danvers, MA, USA), rabbit anti-procollagen I (1:1000; Abcam, Cambridge, UK), rabbit LC3 (1:1000, Cell Signaling Technology, Danvers, MA, USA), mouse anti-p62 (1:1000; Abcam, Cambridge, UK), mouse XBP-1s (1:1000; Cell Signaling Technology, Danvers, MA, USA), mouse poly ubiquitinylated proteins, multi ubiquitin chains (1:500; Biomol Int., Plymouth Meeting, PA, USA), and rabbit anti- β-actin (1:1000; Sigma-Aldrich Corp., St. Louis, MO, USA) as a loading control. In the next step the appropriate horseradish peroxidase conjugated secondary antibody: anti-mouse immunoglobulin G (IgG) (whole molecule) (1:2000; Sigma-Aldrich Corp., St. Louis, MO, USA), anti-rabbit antibodies (1:2000; Cell Signaling Technology, Danvers, MA, USA), anti-rabbit immunoglobulin G (IgG), Fc, HRP conjugate antibodies (1:2000; EMD Millipore Corp., Temecula, CA, USA) or anti-mouse IgG (whole molecule)—alkaline phosphatase antibody (1:2000; Sigma-Aldrich Corp., St. Louis, MO, USA) was added for 1 h with gentle shaking. After washing with TBS-T membranes were subjected to Westar Supernova Chemiluminescent Substrate for Western Blotting (Cyanagen, Bologna, Italy) and analyzed by densitometry (G:BOX, Syngene, Cambridge, UK). The intensity of analyzed proteins were normalized to β-actin which was a loading control. The data were expressed as a percentage of the normal sample taken as 100%. Determination of polyubiquitinated proteins was performed using Sigma Fast BCIP/NBT Alkaline Phosphatase Substrate (Sigma-Aldrich, St. Louis, MO, USA) by colorimetric detection (Gel Doc XR and Gel Documentation System; Molecular Imager Gel Doc XR, Bio-Rad Laboratories Inc., Hercules, CA, USA).
4.7. Immunofluorescence
Fibroblasts grown on cover-slips were fixed in 4% paraformaldehyde in PBS for 10 min at room temperature. After fixation, the cells were permeabilized in PBS containing 0.2% Triton-X100 for 5 min and blocked in 5 % normal donkey serum (Sigma-Aldrich Corp., St. Louis, MO, USA) at room temperature for 60 min to block non-specific reactions. Then cells were incubated with mouse monoclonal anti-collagen type I antibody (1:250, Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) and rabbit monoclonal anti-LC3B antibody (1:2000, Cell Signaling Technology, Danvers, MA, USA) or rabbit polyclonal anti-LAMP2A antibody (1:100, Abcam) for 60 min at room temperature. After incubation, the cells were washed three times with PBS and incubated in donkey anti-mouse IgG conjugated with Alexa Fluor 543 (1:200, Molecular Probes) or donkey anti-rabbit IgG conjugated with Alexa Fluor 488 (1:200, Molecular Probes) at room temperature for 1 h. Then, the cells were washed three times in PBS and stained with 4′,6′-diamidino-2-phenylindole (DAPI, Sigma-Aldrich Corp., St. Louis, MO, USA) for 10 min to indicate cell nuclei. The samples were washed twice with PBS and embedded in fluorescent medium (Medium Coverquick, Hygeco, OH, USA), dried overnight and stored in the dark until assessment. The immune labeled cells were analyzed using Nikon Digital Sight DS-Fi1 camera and a fluorescence microscope Nikon ECLIPSE Ti/C1 Plus, equipped with three filters DAPI (blue), FITC (green), and TRITC (red) (excitation wavelength/emission filter: 405/450 nm, 488/515 nm, 543/605 nm, respectively). No fluorescence signal was detected when cells were incubated with secondary antibodies alone (data not shown). At least five pictures of different areas of each treatment group were taken, independently analyzed and one representative image for each study group was presented.
4.8. Steady-State Analysis of Type I Collagen
Procollagens was extracted from cell lysates by precipitation overnight at 4 °C with ammonium sulfate (176 mg/mL). To obtain collagen, procollagen was subjected to digestion with pepsin (50 μg/mL) for 4 h at 4 °C. For electrophoretic analysis of migration of collagen chains, SDS-urea-PAGE (5% polyacrylamide gel) and silver staining were used.
4.9. Subcellular Fractionation
Cells were suspended in buffer containing 40 mM KCl, 5 mM MgCl2, 2 mM EGTA, 10 mM HEPES, pH 7.5 for 30 min on ice. They were then homogenized by shearing 30 times through a 28.5-gauge needle and centrifuged at 1000× g for 10 min. The pellet was collected as the nuclear fraction, while the supernatant was subjected to centrifugation at 12,000× g for 10 min. The lysosome enriched pellet was washed using isotonic buffer (150 mM NaCl, 5 mM MgCl2, 2 mM EGTA, 10 mM HEPES pH 7.5) and dissolved in lysis buffer (1% Triton X-100, 150 mM NaCl, 50 mM Tris-HCl pH 7.5). The presence of collagen type I in this fraction was analyzed by Western blot.
4.10. Determination of Proteasome Activities
Cells were sonicated in lysis buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM EDTA,1 mM EGTA, 0.5% Triton X, and centrifuged at 12,000×
g for 15 min at 40 °C. The total protein concentration in supernatants was determined by the Bradford method in BioPhotometer (Eppendorf, Hamburg, Germany), using the Bio-Rad Protein Assay Dye Reagent Concentrate (Bio-Rad Laboratories Inc., Hercules, CA, USA) with BSA as a standard. The supernatants were diluted to concentration of 1.5 mg protein/mL in the lysis buffer. The reaction mixture (total volume of 50 μL) contained 30 μL of assay buffer (100 mM Tris/HCL, pH 7.5, 1 mM EDTA, 1 mM EGTA pH 7.5), 10 μL of cell lysate supernatant and 10 μL of fluorogenic peptide-AMC substrates: Suc-LLVY-AMC (Sigma Aldrich Corp., St. Louis, MO, USA) for chymotrypsin-like activity, Bz-VGR-AMC (Enzo Life Sciences, Inc., Farmingdale, NY, USA) for trypsin-like activity or Z-LLE-AMC (Enzo Life Sciences, Inc., Farmingdale, NY, USA) for caspase-like activity in a final concentration of 100 µM each [
55]. The 96-well black plates (Corning Inc., Corning, NY, USA) were used and the assays were performed at 37 °C in FLUOStar OPTIMA (BMG Labtech Gmbh, Offenburg, Germany) over 30 min with one reading every 2 min, at 355 nm for excitation and 460 nm for emission. One unit of the proteasome activity was expressed as the amount of AMC released from the substrate per minute (pmol/min). The activity was calculated for the amount of total protein (U/mg). All assays were performed in triplicates.
4.11. Zymography for the Determination of MMP Activity
The conditioned media were collected and subjected non-reducing SDS–PAGE. The substrate for MMP was gelatin (1 mg/mL) (Sigma-Aldrich Corp., St. Louis, MO, USA). In order to remove SDS, gels were incubated in 2.5% Triton X-100 solution for 30 min at room temperature. In the next step, gels were incubated overnight in 50 mM Tris-HCl, pH 8.0, 5 mM CaCl2, 5 µM ZnCl2 and 0.02% NaN3) at 37 °C with gentle shaking. After that, they were stained with Commassie blue R-250 solution and destained until the appearance of white stripes on a dark blue background. Images of the zymograms were analyzed by densitometry (G:BOX, Syngene, Cambridge, UK).
4.12. Statistical Analysis
The results were statistically analyzed using the Statistica 12 software (StatSoft, Tulsa, OK, USA) and presented as the mean ± standard deviation (SD). Statistical differences were estimated by the use of one-way ANOVA followed by Tukey’s test and values of p < 0.05 were considered as significant.



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










