
Exploring the Expression of Pro-Inflammatory and Hypoxia-Related MicroRNA-20a, MicroRNA-30e, and MicroRNA-93 in Periodontitis and Gingival Mesenchymal Stem Cells under Hypoxia

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
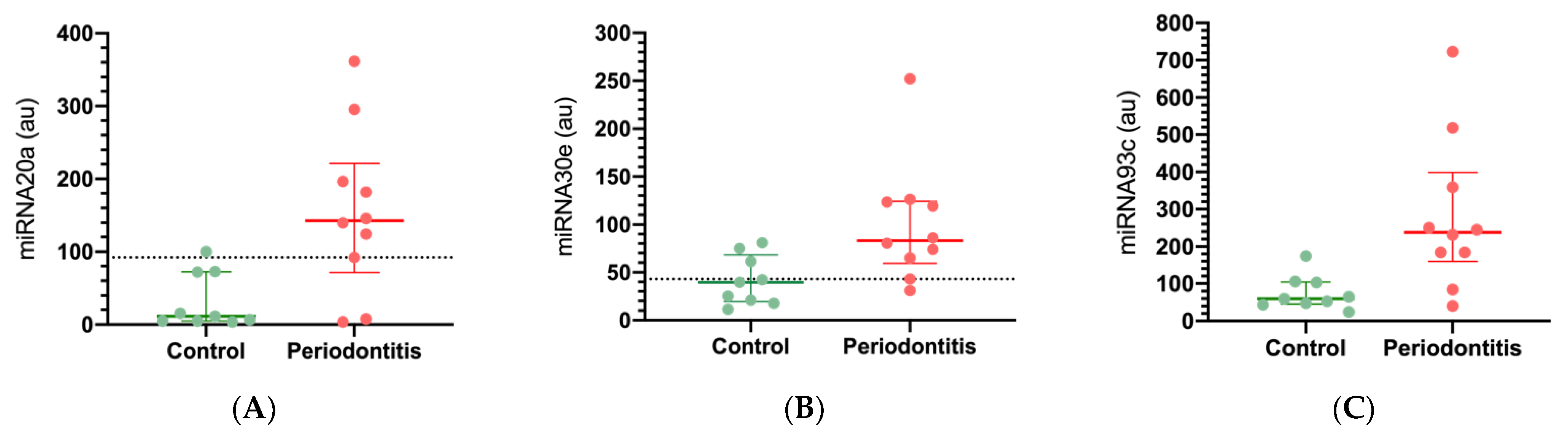
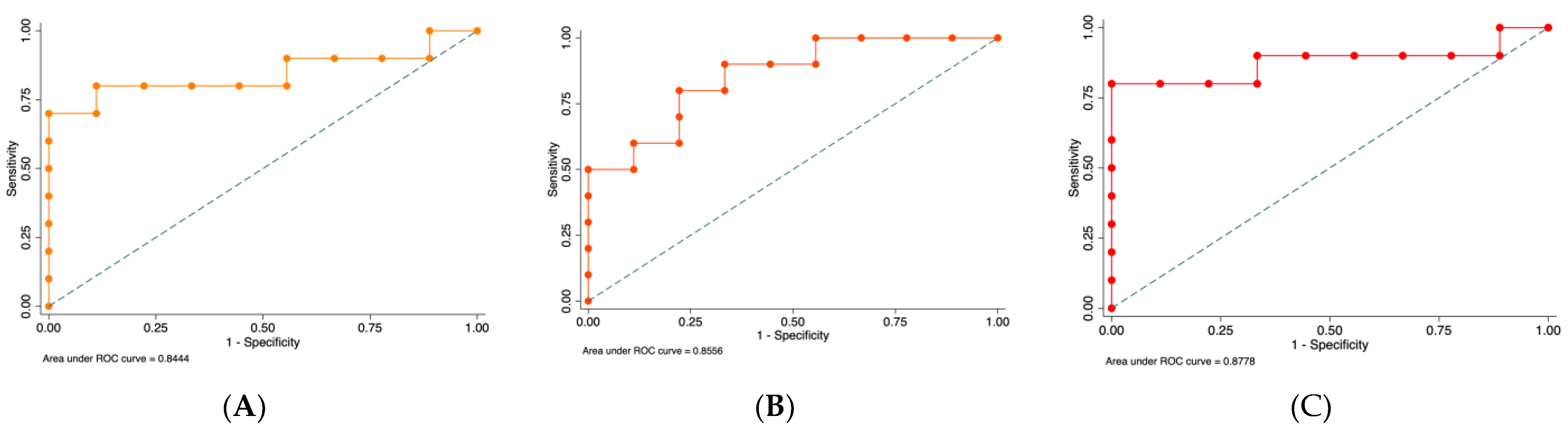
2.1. MiRNA Expression in Healthy and Periodontal Disease Biopsies
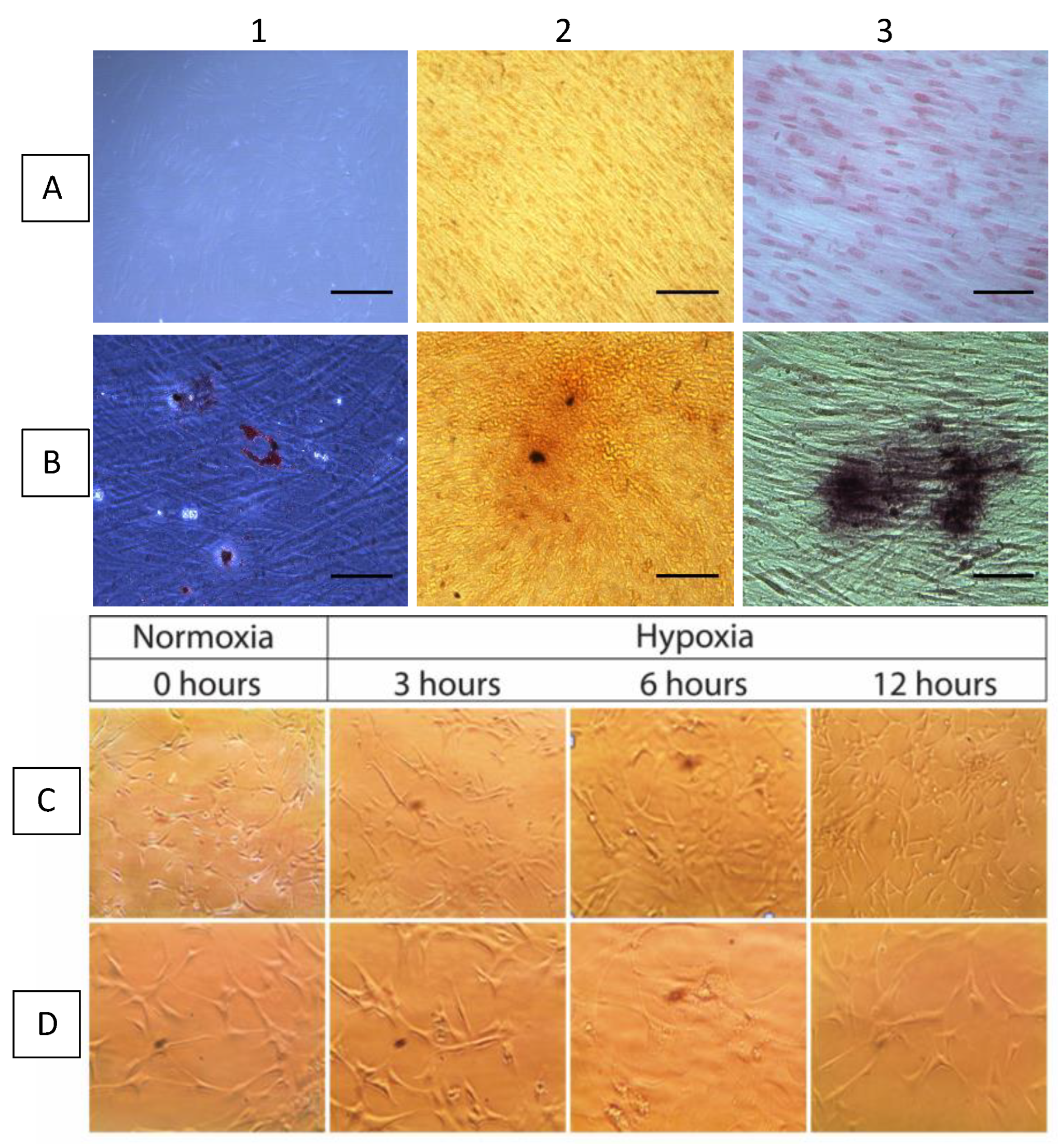
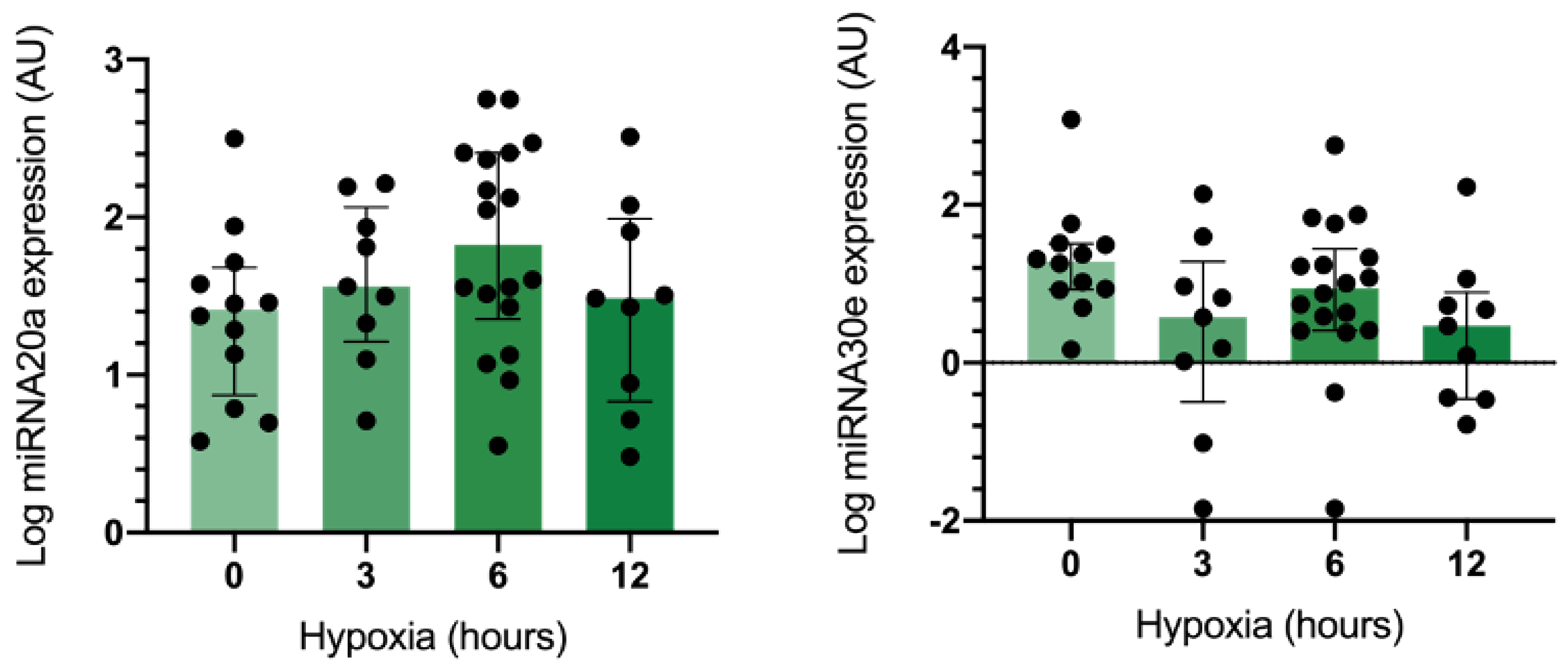
2.2. MiRNA Expression in Gingival Mesenchymal Stem Cells (GMSCs) under Hypoxic Conditions
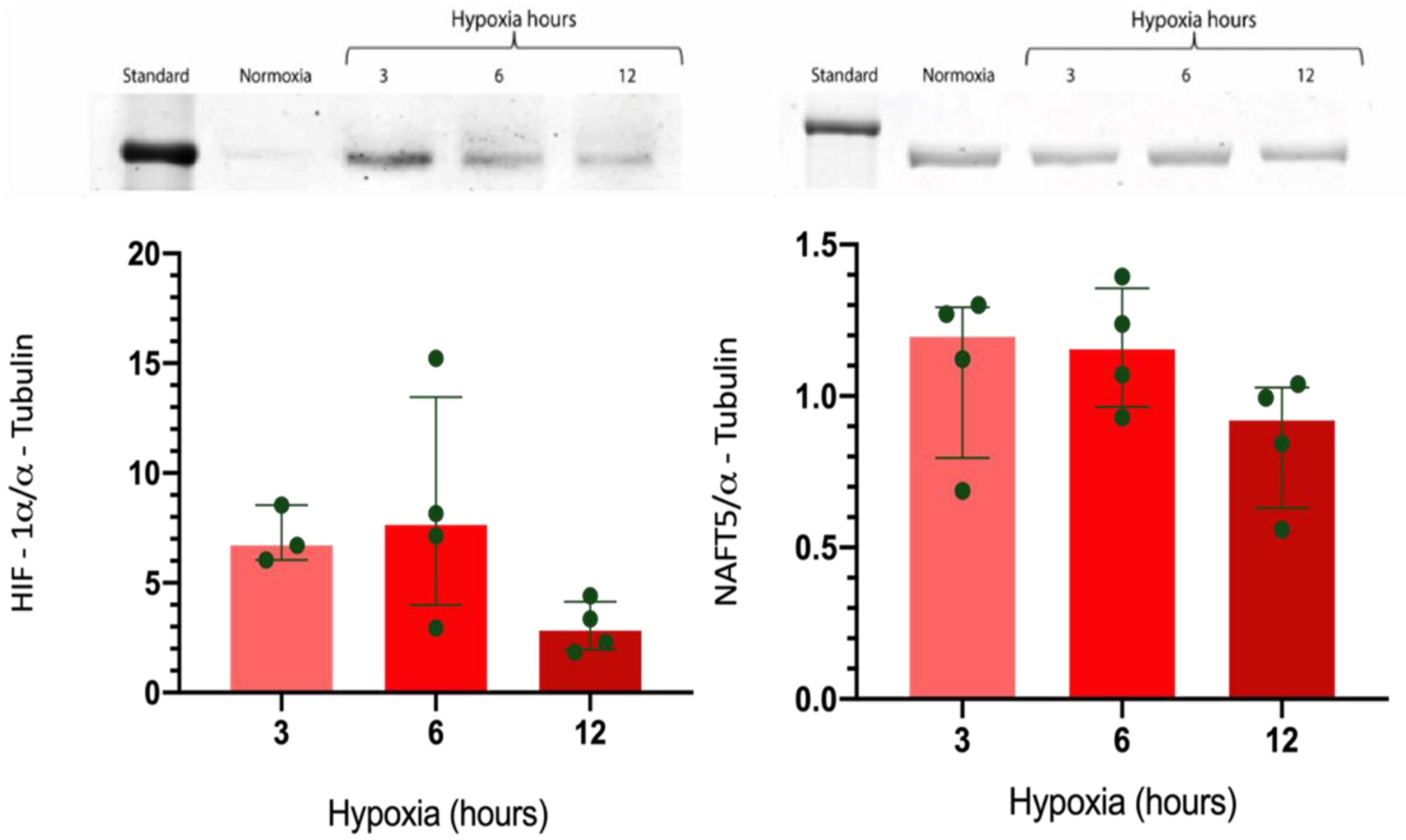
2.3. Effect of Hypoxia in the Expression of HIF-1α and NFAT5 in GMSC
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Sampling Collection
4.3. Cell Culture
4.4. RNA Extraction and qRT-PCR Analysis
4.5. HIF-1α and NFAT5 Expression
4.6. Statistics Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef]
- Barros, S.P.; Hefni, E.; Nepomuceno, R.; Offenbacher, S.; North, K. Targeting epigenetic mechanisms in periodontal diseases. Periodontology 2000 2018, 78, 174–184. [Google Scholar] [CrossRef]
- Kornman, K.S.; Crane, A.; Wang, H.Y.; Giovlne, F.S.; Newman, M.G.; Pirk, F.W.; Wilson, T.G.; Higginbottom, F.L.; Duff, G.W. The interleukin-1 genotype as a severity factor in adult periodontal disease. J. Clin. Periodontol. 1997, 24, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.; Cochran, D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Wood, N.; Serio, F. Interleukin-11 and IL17 and the pathogenesis of periodontal disease. J. Periodontol. 2004, 75, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Crivello, A.; Offenbacher, S.; Moretti, A.; Paquette, D.W.; Barros, S.P. Interferon-gamma promoter hypomethylation and increased expression in chronic periodontitis. J. Clin. Periodontol. 2010, 37, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Larsson, L.; Thorbert-Mros, S.; Rymo, L.; Berglundh, T. Influence of epigenetic modifications of the interleukin-10 promoter on IL10 gene expression. Eur. J. Oral Sci. 2012, 120, 14–20. [Google Scholar] [CrossRef]
- Ishida, K.; Kobayashi, T.; Ito, S.; Komatsu, Y.; Yokoyama, T.; Okada, M.; Abe, A.; Murasawa, A.; Yoshie, H. Interleukin-6 gene promoter methylation in rheumatoid arthritis and chronic periodontitis. J. Periodontol. 2012, 83, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, R.; Miccoli, G.; Reda, R.; Mazzoni, A.; Di Nardo, D.; Testarelli, L. Sulcus fluid volume, IL-6, and Il-1b concentrations in periodontal and peri-implant tissues comparing machined and laser-microtextured collar/abutment surfaces during 12 weeks of healing: A split-mouth RCT. Clin. Oral Implant. Res. 2022, 33, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.T.; Li, J.P.; Ng, K.M.; Tipoe, G.L.; Leung, W.K.; Fung, M.L. Expression of Hypoxia-Inducible Factor-1α in Human Periodontal Tissue. J. Periodontol. 2011, 82, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lu, F.; Zhang, Z.; Yang, X.; Chen, Y. The Role of Psychologic Stress-Induced Hypoxia-Inducible Factor-1α in Rat Experimental Periodontitis. J. Periodontol. 2011, 82, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Chen, Y.; Leung, W.K. Role of the Hypoxia-Inducible Factor in Periodontal Inflammation. In Hypoxia and Human Diseases; IntechOpen: London, UK, 2017; p. 285. [Google Scholar] [CrossRef]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Forster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef]
- Semenza, G.L. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu. Rev. Pathol. 2014, 9, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Blouin, C.C.; Page, E.L.; Soucy, G.M.; Richard, D.E. Hypoxic gene activation by lipopolysaccharide in macrophages: Implication of hypoxia-inducible factor 1 alpha. Blood 2004, 103, 1124–1130. [Google Scholar] [CrossRef]
- Palomäki, S.; Pietilä, M.; Laitinen, S.; Pesälä, J.; Sormunen, R.; Lehenkari, P.; Koivunen, P. HIF-1α is Upregulated in Human Mesenchymal Stem Cells. Stem Cells 2013, 31, 1902–1909. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Song, M.K.; Park, C.; Lee, Y.D.; Kim, H.; Kim, M.K.; Kwon, J.O.; Koo, J.H.; Joo, M.S.; Kim, S.G.; Kim, H.-H. Galpha12 regulates osteoclastogenesis by modulating NFATc1expression. J. Cell Mol. Med. Sci. 2018, 22, 849–860. [Google Scholar] [CrossRef]
- Gölz, L.; Memmert, S.; Rath-Deschner, B.; Jäger, A.; Appel, T.; Baumgarten, G.; Götz, W.; Frede, S. Hypoxia and P. gingivalis synergistically induce HIF-1 and NF-κB activation in PDL cells and periodontal diseases. Mediat. Inflamm. 2015, 2015, 438085. [Google Scholar] [CrossRef]
- Xiao, C.; Bai, G.; Du, Y.; Jiang, H.; Yu, X. Association of high HIF-1α levels in serous periodontitis with external root resorption by the NFATc1 pathway. J. Mol. Histol. 2020, 51, 649–658. [Google Scholar] [CrossRef]
- Razban, V.; Lotfi, A.S.; Soleimani, M.; Ahmadi, H.; Massumi, M.; Khajeh, S.; Ghaedi, M.; Arjmand, S.; Najavand, S.; Khoshdel, A. HIF-1α Overexpression Induces Angiogenesis in Mesenchymal Stem Cells. Bioresearch Open Access 2012, 1, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Arnett, T.R.; Gibbons, D.C.; Utting, J.C.; Orriss, I.R.; Hoebertz, A.; Rosendaal, M.; Meghji, S. Hypoxia is a major stimulator of osteoclast formation and bone resorption. J. Cell Physiol. 2003, 196, 2–8. [Google Scholar] [CrossRef]
- Muzylak, M.; Price, J.S.; Horton, M.A. Hypoxia induces giant osteoclast formation and extensive bone resorption in the cat. Calcif. Tissue Int. 2006, 79, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.G.; Croce, C.M.; Negrini, M.; et al. A microRNA signature of hypoxia. Mol. Cell Biol. 2007, 27, 1859–1867. [Google Scholar] [CrossRef]
- Villanueva, S.; Suazo, C.; Santapau, D.; Pérez, F.; Quiroz, M.; Carreño, J.E.; Illanes, S.; Lavandero, S.; Michea, L.; Irarrazabal, C.E. NFAT5 is activated by hypoxia: Role in ischemia and reperfusion in the rat kidney. PLoS ONE 2012, 7, e39665. [Google Scholar] [CrossRef]
- Serman, Y.; Fuentealba, R.A.; Pasten, C.; Rocco, J.; Ko, B.C.B.; Carrión, F.; Irarrázabal, C.E. Emerging new role of NFAT5 in inducible nitric oxide synthase in response to hypoxia in mouse embryonic fibroblast cells. Am. J. Physiol. Cell Physiol. 2019, 317, C31–C38. [Google Scholar] [CrossRef]
- Standart, N.; Jackson, R.J. MicroRNAs repress translation of m7Gppp-capped target mRNAs in vitro by inhibiting initiation and promoting deadenylation. Genes Dev. 2007, 21, 1975–1982. [Google Scholar] [CrossRef]
- Wang, B.; Love, T.M.; Call, M.E.; Doench, J.G.; Novina, C.D. Recapitulation of short RNA-directed translational gene silencing in vitro. Molecular Cell. 2006, 22, 553–560. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef]
- Hua, Z.; Lv, Q.; Ye, W.; Wong, C.K.; Cai, G.; Gu, D.; Ji, Y.; Zhao, C.; Wang, J.; Yang, B.B.; et al. MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 2006, 1, e116. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, C.E.; Lamont, R.J. Porphyromonas gingivalis Induction of MicroRNA-203 Expression Controls Suppressor of Cytokine Signaling 3 in Gingival Epithelial Cells. Infect. Immun. 2011, 79, 2632–2637. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, J.L.; Fang, Y.M.; Gong, S.T.; Li, M.Y.; Wu, M.H.; Lai, X.M.; Zeng, G.C.; Wang, Y.; Yang, K.; et al. microRNA-146a promotes mycobacterial survival in macrophages through suppressing nitric oxide production. Sci. Rep. 2016, 6, 23351. [Google Scholar] [CrossRef]
- Hulsmans, M.; De Keyzer, D.; Holvoet, P. MicroRNAs regulating oxidative stress and inflammation in relation to obesity and atherosclerosis. FASEB J. 2011, 25, 2515–2527. [Google Scholar] [CrossRef]
- Donker, R.B.; Mouillet, J.F.; Nelson, D.M.; Sadovsky, Y. The expression of Argonaute2 and related microRNA biogenesis proteins in normal and hypoxic trophoblasts. Mol. Hum. Reprod. 2007, 13, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Mouillet, J.F.; Chu, T.; Nelson, D.M.; Mishima, T.; Sadovsky, Y. MiR-205 silences MED1 in hypoxic primary human trophoblasts. FASEB J. 2010, 24, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.L.; Kesavalu, L. Polymicrobial Infection with Periodontal Pathogens Specifically Enhances MicroRNA miR-146a in ApoE-/- Mice during Experimental Periodontal Disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Na, H.S.; Jeong, S.Y.; Jeong, S.H.; Park, H.R.; Chung, J. Comparison of inflammatory microRNA expression in healthy and periodontitis tissues. Biocell 2011, 35, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.F.; Shu, R.; Jiang, S.Y.; Liu, D.L.; Zhang, X.L. Comparison of microRNA profiles of human periodontal disease and healthy gingival tissues. Int. J. Oral Sci. 2011, 3, 125–134. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, F.; Suvan, J. Obesity, inflammation, and oral infections: Are microRNAs the missing link? J. Dent. Res. 2012, 91, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Perri, R.; Nares, S.; Zhang, S.; Barros, S.P.; Offenbacher, S. Microrna modulation in obesity and periodontitis. J. Dent. Res. 2012, 91, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Stoecklin-Wasmer, C.; Guarnieri, P.; Celenti, R.; Demmer, R.T.; Kebschull, M.; Papapanou, P.N. MicroRNAs and Their Target Genes in Gingival Tissues. J. Dent. Res. 2012, 91, 934–940. [Google Scholar] [CrossRef]
- Kebschull, M.; Papapanou, P. Mini but Mighty-microRNAs in the Pathobiology of Periodontal Disease. Periodontology 2000 2015, 69, 201–220. [Google Scholar] [CrossRef]
- Mico-Martinez, P.; Garcia-Gimenez, J.L.; Seco-Cervera, M.; Lopez-Roldan, A.; Alminana-Pastor, P.J.; Alpiste-Illueca, F.; Pallardo, F.V. Mir-1226 detection in GCF as potential biomarker of chronic periodontitis: A pilot study. Med. Oral Patol. Oral Cir. Bucal. 2018, 23, e308–e314. [Google Scholar] [CrossRef] [PubMed]
- Luan, X.; Zhou, X.; Trombetta-eSilva, J.; Francis, M.; Gaharwar, A.K.; Atsawasuwan, P.; Diekwisch, T.G.H. MicroRNAs and Periodontal Homeostasis. J. Dent. Res. 2017, 96, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Pettiette, M.T.; Zhang, S.; Moretti, A.J.; Kim, S.J.; Naqvi, A.R.; Nares, S. Microrna expression profiles in external cervical resorption. J. Endod. 2019, 45, 1106–1113.e2. [Google Scholar] [CrossRef] [PubMed]
- Motedayyen, H.; Ghotloo, S.; Saffari, M.; Sattari, M.; Amid, R. Evaluation of microrna-146a and its targets in gingival tissues of patients with chronic periodontitis. J. Periodontol. 2015, 86, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Asaàd, F.; Garaicoa-Pazmiño, C.; Dahlin, C.; Larsson, L. Expression of MicroRNAs in Periodontal and Peri-Implant Diseases: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 4147. [Google Scholar] [CrossRef] [PubMed]
- Radović, N.; Nikolić Jakoba, N.; Petrović, N.; Milosavljević, A.; Brković, B.; Roganović, J. MicroRNA-146a and microRNA-155 as novel crevicular fluid biomarkers for periodontitis in non-diabetic and type 2 diabetic patients. J. Clin. Periodontol. 2018, 45, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Luan, X.; Zhou, X.; Naqvi, A.; Francis, M.; Foyle, D.; Nares, S.; Diekwisch, T.G.H. MicroRNAs and immunity in periodontal health and disease. Int. J. Oral Sci. 2018, 10, 24. [Google Scholar] [CrossRef]
- Yu, X.J.; Xiao, C.J.; Du, Y.M.; Liu, S.; Du, Y.; Li, S. Effect of hypoxia on the expression of RANKL/OPG in human periodontal ligament cells in vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 12929–12935. [Google Scholar] [PubMed]
- Pocock, R. Invited review: Decoding the microRNA response to hypoxia. Pflügers Arch.-Eur. J. Physiol. 2011, 461, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, T.; Tomofuji, T.; Ekuni, D.; Azuma, T.; Maruyama, T.; Fujimori, K.; Sugiura, Y.; Morita, M. Serum microRNAs and chronic periodontitis: A case-control study. Arc. Oral Biol. 2019, 101, 57–63. [Google Scholar] [CrossRef]
- Saito, A.; Horie, M.; Ejiri, K.; Aoki, A.; Katagiri, S.; Maekawa, S.; Suzuki, S.; Kong, S.; Yamauchi, T.; Yamaguchi, Y. MicroRNA profiling in gingival crevicular fluid of periodontitis-a pilot study. FEBS Open Bio. 2017, 7, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Su, L.; Duan, X.; Chen, X.; Hays, A.; Upadhyayula, S.; Shivde, J.; Wang, H.; Li, Y.; Huang, D.; et al. MicroRNA-21 down-regulates inflammation and inhibits periodontitis. Mol. Immunol. 2018, 101, 608–614. [Google Scholar] [CrossRef]
- Ning, Z.Q.; Lu, H.L.; Chen, C.; Wang, L.; Cai, W.; Li, Y.; Cao, T.H.; Zhu, J.; Shu, Y.Q.; Shen, H. MicroRNA-30e reduces cell growth and enhances drug sensitivity to gefitinib in lung carcinoma. Oncotarget 2017, 8, 4572–4581. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Ye, Z.; Sun, Y.; Yang, H.; Li, L. Relationship between circulating miRNA-30e and no-reflow phenomenon in STEMI patients undergoing primary coronary intervention. Scand. J. Clin. Lab. Investig. 2018, 78, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Bhattacharya, S.; Singh Rawat, B.; Kumar, A.; Kumar, A.; Niraj, K.; Chande, A.; Gandhi, P.; Khetan, D.; Aggarwal, A.; et al. MicroRNA-30e-5p has an Integrated Role in the Regulation of the Innate Immune Response during Virus Infection and Systemic Lupus Erythematosus. iScience 2020, 23, 101322. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, S.; Farber, C.R.; Dokun, A.O.; Pitsillides, A.N.; Wang, T.; Lye, R.J.; Annex, B.H. MicroRNA-93 controls perfusion recovery after hindlimb ischemia by modulating expression of multiple genes in the cell cycle pathway. Circulation 2013, 127, 1818–1828. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, Q.S.; Ding, W.B.; Zhang, L.L.; Wang, H.C.; Zhu, Y.J.; He, W.; Chai, Y.-N.; Liu, Y.-W. Increased microRNA-93-5p inhibits osteogenic differentiation by targeting bone morphogenetic protein-2. PLoS ONE 2017, 12, e0182678. [Google Scholar] [CrossRef] [PubMed]
- Imtiyaz, H.Z.; Simon, M.C. Hypoxia-Inducible Factors as Essential Regulators of Inflammation. Curr. Top. Microbiol. Immunol. 2010, 345, 105–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.P.; Li, F.Y.L.; Xu, A.; Cheng, B.; Tsao, S.W.; Fung, M.L.; Leung, W.K. Lipopolysaccharide and hypoxia-induced HIF-1 activation in human gingival fibroblasts. J. Periodontol. 2012, 83, 816–824. [Google Scholar] [CrossRef]
- Afacan, B.; Öztürk, V.Ö.; Paşalı, Ç.; Bozkurt, E.; Köse, T.; Emingil, G. Gingival crevicular fluid and salivary HIF-1α, VEGF, and TNF-α levels in periodontal health and disease. J. Periodontol. 2019, 90, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Neuhofer, W. Role of NFAT5 in inflammatory disorders associated with osmotic stress. Curr. Genomics 2010, 11, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Park, E.; Kim, H.J.; Na, H.S.; Chung, J. Porphyromonas gingivalis-induced miR-132 regulates TNFα expression in THP-1 derived macrophages. Springerplus 2016, 5, 761. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.Q.; Liu, H.; Sun, H.L.; Xiang, J.; Wang, X.; Jiang, C.; Ma, L.; Cao, Z. MiR-361-3p/Nfat5 Signaling Axis Controls Cementoblast Differentiation. J. Dent. Res. 2019, 98, 1131–1139. [Google Scholar] [CrossRef]
- Caton, J.G.; Armitage, G.; Berglundh, T.; Chapple, I.L.C.; Jepsen, S.; Kornman, K.S.; Mealey, B.L.; Papapanou, P.N.; Sanz, M.; Tonetti, M.S. A new classification scheme for periodontal and peri-implant diseases and conditions—Introduction and key changes from the 1999 classification. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S1–S8. [Google Scholar] [CrossRef] [PubMed]
- Mitrano, T.; Grob, M.S.; Carrión, F.; Nova-Lamperti, E.; Luz, P.A.; Fierro, F.; Quintero, A.; Chaparro, A.; Sanz, A. Culture and characterization of mesenchymal stem cells from human gingival tissue. J. Periodontol. 2010, 81, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.; Carrión, F.; Chaparro, A. Mesenchymal stem cells from the oral cavity and their potential value in tissue engineering. Periodontology 2000 2015, 67, 251–267. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaparro, A.; Lozano, M.; Gaedechens, D.; López, C.; Albers, D.; Hernández, M.; Pascual, A.; Nart, J.; Irarrazabal, C.E. Exploring the Expression of Pro-Inflammatory and Hypoxia-Related MicroRNA-20a, MicroRNA-30e, and MicroRNA-93 in Periodontitis and Gingival Mesenchymal Stem Cells under Hypoxia. Int. J. Mol. Sci. 2022, 23, 10310. https://doi.org/10.3390/ijms231810310
Chaparro A, Lozano M, Gaedechens D, López C, Albers D, Hernández M, Pascual A, Nart J, Irarrazabal CE. Exploring the Expression of Pro-Inflammatory and Hypoxia-Related MicroRNA-20a, MicroRNA-30e, and MicroRNA-93 in Periodontitis and Gingival Mesenchymal Stem Cells under Hypoxia. International Journal of Molecular Sciences. 2022; 23(18):10310. https://doi.org/10.3390/ijms231810310
Chicago/Turabian StyleChaparro, Alejandra, Mauricio Lozano, Dominique Gaedechens, Carolina López, Daniela Albers, Marcela Hernández, Andrés Pascual, José Nart, and Carlos E. Irarrazabal. 2022. "Exploring the Expression of Pro-Inflammatory and Hypoxia-Related MicroRNA-20a, MicroRNA-30e, and MicroRNA-93 in Periodontitis and Gingival Mesenchymal Stem Cells under Hypoxia" International Journal of Molecular Sciences 23, no. 18: 10310. https://doi.org/10.3390/ijms231810310