
Potentiality and Inflammatory Marker Expression Are Maintained in Dental Pulp Cell Cultures from Carious Teeth

 , and
, and
Abstract
:1. Introduction
2. Results
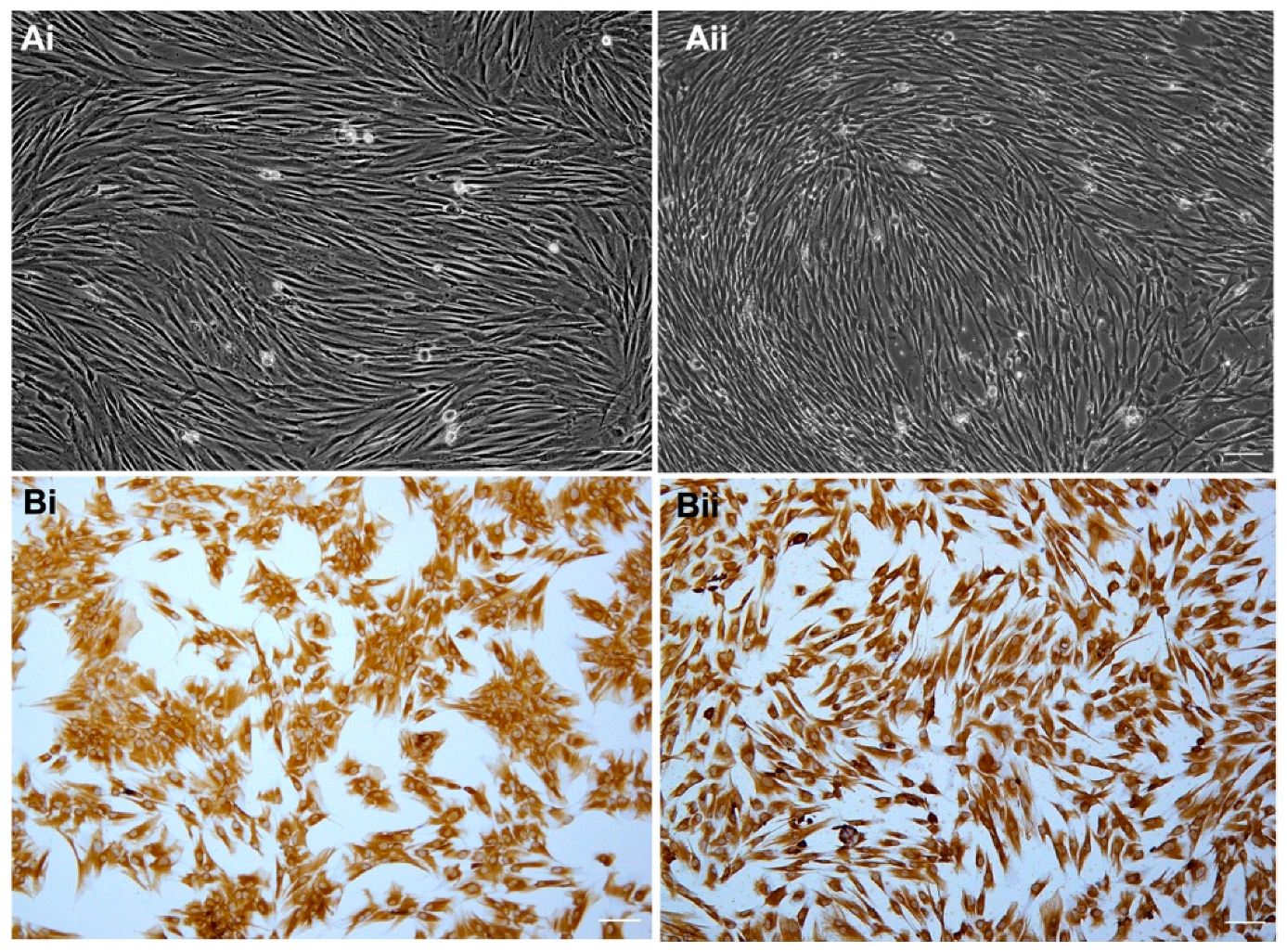
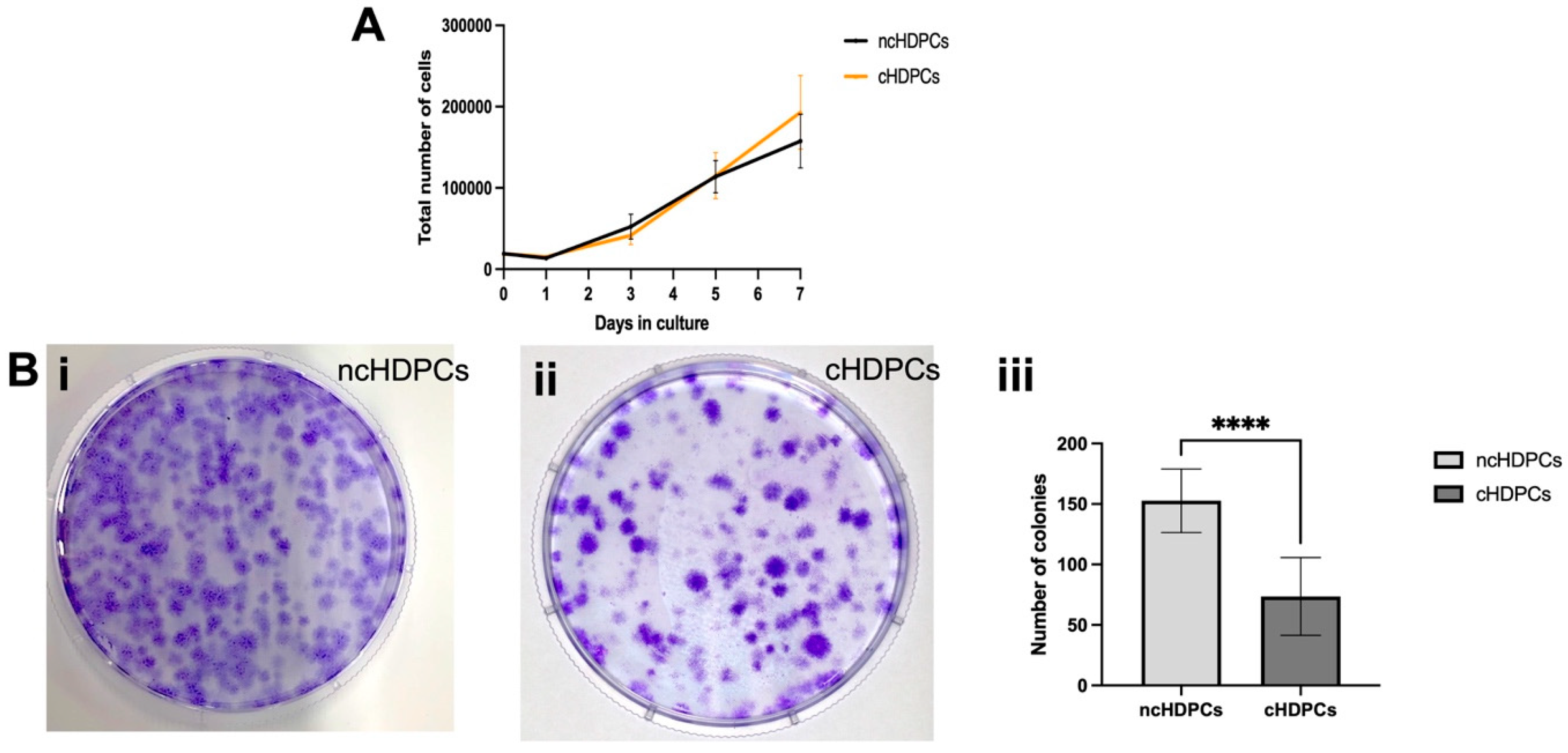
2.1. Establishment and Characterization of Primary Cell Cultures
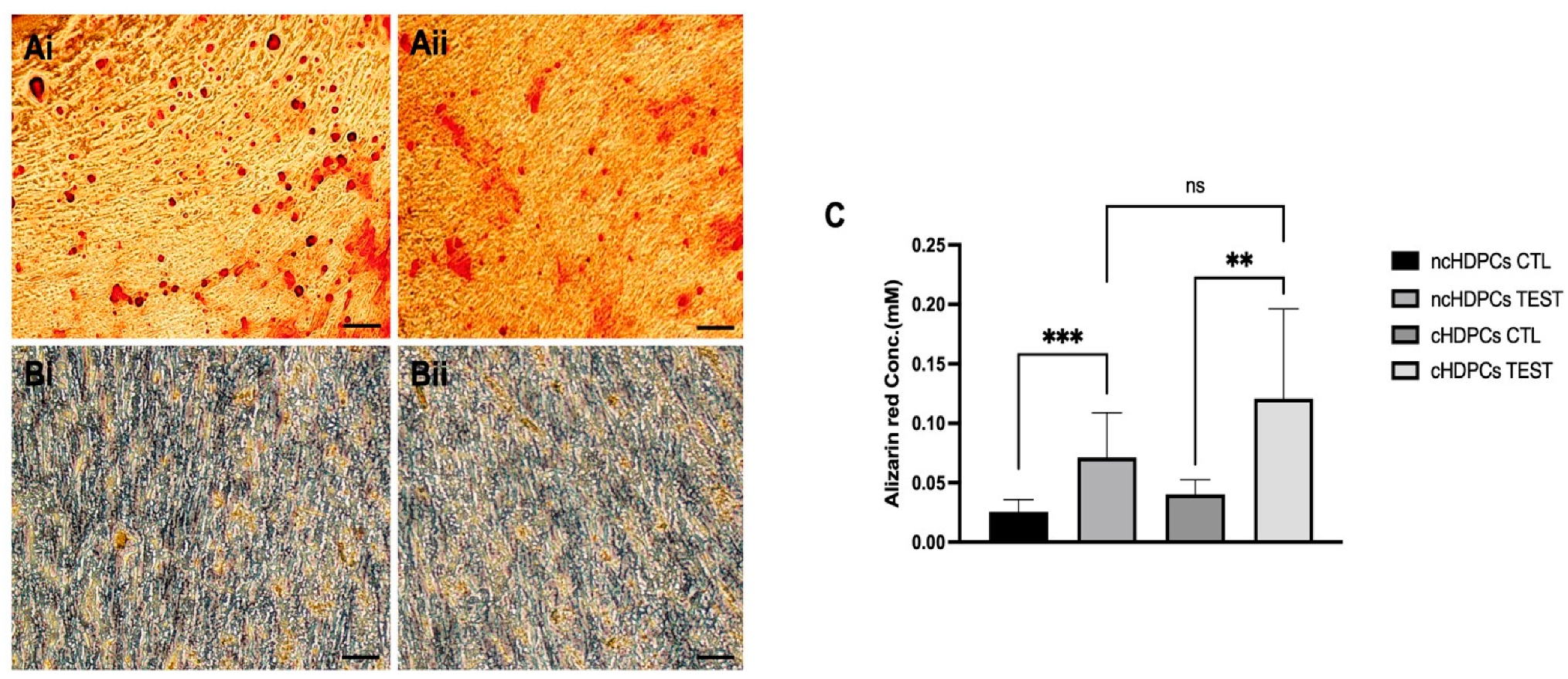
2.2. Differentiation Potential of Human Dental Pulp Cell Cultures

2.3. Gene Expression Analysis in Cultures of Non-Carious and Carious Human Dental Pulp Cells
3. Discussion
4. Methods and Materials
4.1. Cell Isolation and Culture
4.2. Generation of Growth Curves
4.3. Colony Forming Capacity
4.4. Immunocytochemistry
4.5. Differentiation Potential of Human Dental Pulp Cell Cultures
4.6. Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruch, J.V. Odontoblast commitment and differentiation. Biochem. Cell Biol. 1998, 76, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.E. Pulp Anatomy and Characterization of Pulp Cells. In The Dental Pulp, 1st ed.; Goldberg, M.E., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 13–34. [Google Scholar]
- Manivasagam, G.; Reddy, A.; Sen, D.; Nayak, S.; Mathew, M.T.; Rajamanikam, A. Dentistry: Restorative and Regenerative Approaches. In Encyclopedia of Biomedical Engineering; Narayan, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 332–347. [Google Scholar]
- Arora, S.; Cooper, P.R.; Ratnayake, J.T.; Friedlander, L.T.; Rizwan, S.B.; Seo, B.; Hussaini, H.M. A critical review of in vitro research methodologies used to study mineralization in human dental pulp cell cultures. Int. Endod. J. 2022, 55, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Min. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.E.; Garcia-Godoy, F.; Hargreaves, K.M. Regenerative endodontics: A review of current status and a call for action. J. Endod. 2007, 33, 377–390. [Google Scholar] [CrossRef]
- Bergenholtz, G. Inflammatory response of the dental pulp to bacterial irritation. J. Endod. 1981, 7, 100–104. [Google Scholar] [CrossRef]
- Cooper, P.R.; McLachlan, J.L.; Simon, S.; Graham, L.W.; Smith, A.J. Mediators of inflammation and regeneration. Adv. Dent. Res. 2011, 23, 290–295. [Google Scholar] [CrossRef]
- Huang, G.T.; Potente, A.P.; Kim, J.W.; Chugal, N.; Zhang, X. Increased interleukin-8 expression in inflamed human dental pulps. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1999, 88, 214–220. [Google Scholar] [CrossRef]
- Barkhordar, R.A.; Hayashi, C.; Hussain, M.Z. Detection of interleukin-6 in human dental pulp and periapical lesions. Endod. Dent. Traumatol. 1999, 15, 26–27. [Google Scholar] [CrossRef]
- Pezelj-Ribaric, S.; Anic, I.; Brekalo, I.; Miletic, I.; Hasan, M.; Simunovic-Soskic, M. Detection of tumor necrosis factor alpha in normal and inflamed human dental pulps. Arch. Med. Res. 2002, 33, 482–484. [Google Scholar] [CrossRef]
- Xiong, H.; Wei, L.; Peng, B. IL-17 stimulates the production of the inflammatory chemokines IL-6 and IL-8 in human dental pulp fibroblasts. Int. Endod. J. 2015, 48, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, M.; Xiong, H.; Peng, B. Up-regulation of IL-23 expression in human dental pulp fibroblasts by IL-17 via activation of the NF-kappaB and MAPK pathways. Int. Endod. J. 2018, 51, 622–631. [Google Scholar] [CrossRef]
- Cooper, P.R.; Holder, M.J.; Smith, A.J. Inflammation and regeneration in the dentin-pulp complex: A double-edged sword. J. Endod. 2014, 40, S46–S51. [Google Scholar] [CrossRef] [PubMed]
- Alkharobi, H.; Beattie, J.; Meade, J.; Devine, D.; El-Gendy, R. Dental Pulp Cells Isolated from Teeth with Superficial Caries Retain an Inflammatory Phenotype and Display an Enhanced Matrix Mineralization Potential. Front. Physiol. 2017, 8, 244. [Google Scholar] [CrossRef]
- Ma, D.; Gao, J.; Yue, J.; Yan, W.; Fang, F.; Wu, B. Changes in proliferation and osteogenic differentiation of stem cells from deep caries in vitro. J. Endod. 2012, 38, 796–802. [Google Scholar] [CrossRef]
- Louvrier, A.; Euvrard, E.; Nicod, L.; Rolin, G.; Gindraux, F.; Pazart, L.; Houdayer, C.; Risold, P.Y.; Meyer, F.; Meyer, C. Odontoblastic differentiation of dental pulp stem cells from healthy and carious teeth on an original PCL-based 3D scaffold. Int. Endod. J. 2018, 5, e252–e263. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, X.; Wu, J.; Lu, W.; Xu, W.; Wu, B. Dental pulp stem cells from human teeth with deep caries displayed an enhanced angiogenesis potential in vitro. J. Dent. Sci. 2021, 16, 318–326. [Google Scholar] [CrossRef]
- Pereira, L.O.; Rubini, M.R.; Silva, J.R.; Oliveira, D.M.; Silva, I.C.R.; Poças-Fonseca, M.J.; Azevedo, R.B. Comparison of stem cell properties of cells isolated from normal and inflamed dental pulps. Int. Endod. J. 2012, 45, 1080–1090. [Google Scholar] [CrossRef]
- Fanibunda, K.B. A method of measuring the volume of human dental pulp cavities. Int. Endod. J. 1986, 19, 194–197. [Google Scholar] [CrossRef]
- Martens, W.; Wolfs, E.; Struys, T.; Politis, C.; Bronckaers, A.; Lambrichts, I. Expression pattern of basal markers in human dental pulp stem cells and tissue. Cells Tissues Organs 2012, 196, 490–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harumi Miyagi, S.P.; Kerkis, I.; da Costa Maranduba, C.M.; Gomes, C.M.; Martins, M.D.; Marques, M.M. Expression of extracellular matrix proteins in human dental pulp stem cells depends on the donor tooth conditions. J. Endod. 2010, 36, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Alt, E.; Yan, Y.; Gehmert, S.; Song, Y.H.; Altman, A.; Gehmert, S.; Vykoukal, D.; Bai, X. Fibroblasts share mesenchymal phenotypes with stem cells, but lack their differentiation and colony-forming potential. Biol. Cell 2011, 103, 197–208. [Google Scholar] [CrossRef]
- Wang, Z.; Pan, J.; Wright, J.T.; Bencharit, S.; Zhang, S.; Everett, E.T.; Teixeira, F.B.; Preisser, J.S. Putative stem cells in human dental pulp with irreversible pulpitis: An exploratory study. J. Endod. 2010, 36, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alongi, D.J.; Yamaza, T.; Song, Y.; Fouad, A.F.; Romberg, E.E.; Shi, S.; Tuan, R.S.; Huang, G.T. Stem/progenitor cells from inflamed human dental pulp retain tissue regeneration potential. Regen. Med. 2010, 5, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.W.; Ling, J.Q.; Gong, Q.M. The expression of stromal cell-derived factor 1 (SDF-1) in inflamed human dental pulp. J. Endod. 2008, 34, 1351–1354. [Google Scholar] [CrossRef]
- Cooper, P.R.; Smith, A.J. Molecular mediators of pulp inflammation and regeneration. Endod. Top. 2013, 28, 90–105. [Google Scholar] [CrossRef]
- Farges, J.C.; Carrouel, F.; Keller, J.F.; Baudouin, C.; Msika, P.; Bleicher, F.; Staquet, M.J. Cytokine production by human odontoblast-like cells upon Toll-like receptor-2 engagement. Immunobiology 2011, 216, 513–517. [Google Scholar] [CrossRef]
- Staquet, M.J.; Durand, S.H.; Colomb, E.; Romeas, A.; Vincent, C.; Bleicher, F.; Lebecque, S.; Farges, J.C. Different Roles of Odontoblasts and Fibroblasts in Immunity. J. Dent. Res. 2008, 87, 256–261. [Google Scholar] [CrossRef]
- Mutoh, N.; Tani-Ishii, N.; Tsukinoki, K.; Chieda, K.; Watanabe, K. Expression of toll-like receptor 2 and 4 in dental pulp. J. Endod. 2007, 33, 1183–1186. [Google Scholar] [CrossRef]
- Fawzy El-Sayed, K.M.; Klingebiel, P.; Dörfer, C.E. Toll-like Receptor Expression Profile of Human Dental Pulp Stem/Progenitor Cells. J. Endod. 2016, 42, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, Q.L.; Huang, P.; Yu, Q.; Wang, Z.H.; Cooper, P.R.; Smith, A.J.; He, W. CpG ODN-induced matrix metalloproteinase-13 expression is mediated via activation of the ERK and NF-κB signalling pathways in odontoblast cells. Int. Endod. J. 2013, 46, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Smith, A.J.; Berdal, A.; Lumley, P.J.; Cooper, P.R. The MAP kinase pathway is involved in odontoblast stimulation via p38 phosphorylation. J. Endod. 2010, 36, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Zhang, C.; Tani-Ishii, N.; Shi, S.; Wang, C.Y. NF-kappaB activation in human dental pulp stem cells by TNF and LPS. J. Dent. Res. 2005, 84, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Takahashi, Y.; Graham, L.W.; Simon, S.; Imazato, S.; Smith, A.J. Inflammation-regeneration interplay in the dentine-pulp complex. J. Dent. 2010, 38, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.; Farges, J.C.; Lacerda-Pinheiro, S.; Six, N.; Jegat, N.; Decup, F.; Septier, D.; Carrouel, F.; Durand, S.; Chaussain-Miller, C.; et al. Inflammatory and immunological aspects of dental pulp repair. Pharmacol. Res. 2008, 58, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, M.; Delaleu, N.; Du, Y.; Bickel, M. Cytokine gene expression-part of host defence in pulpitis. Cytokine 2003, 22, 84–88. [Google Scholar] [CrossRef]
- Silva, A.C.; Faria, M.R.; Fontes, A.; Campos, M.S.; Cavalcanti, B.N. Interleukin-1 beta and interleukin-8 in healthy and inflamed dental pulps. J. Appl. Oral Sci. 2009, 17, 527–532. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, J.L.; Sloan, A.J.; Smith, A.J.; Landini, G.; Cooper, P.R. S100 and cytokine expression in caries. Infect. Immun. 2004, 72, 4102–4108. [Google Scholar] [CrossRef] [Green Version]
- Matsumura-Kawashima, M.; Ogata, K.; Moriyama, M.; Murakami, Y.; Kawado, T.; Nakamura, S. Secreted factors from dental pulp stem cells improve Sjögren’s syndrome via regulatory T cell-mediated immunosuppression. Stem Cell Res. Ther. 2021, 12, 182. [Google Scholar] [CrossRef]
- Hung, S.C.; Pochampally, R.R.; Chen, S.C.; Hsu, S.C.; Prockop, D.J. Angiogenic effects of human multipotent stromal cell conditioned medium activate the PI3K-Akt pathway in hypoxic endothelial cells to inhibit apoptosis, increase survival, and stimulate angiogenesis. Stem Cells 2007, 25, 2363–2370. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Cooper, P.R.; Friedlander, L.T.; Rizwan, S.; Seo, B.; Rich, A.M.; Hussaini, H.M. Potential application of immunotherapy for modulation of pulp inflammation: Opportunities for vital pulp treatment. Int. Endod. J. 2021, 54, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.I.; Pitts, N.B.; Tellez, M. The International Caries Classification and Management System (ICCMS™) An Example of a Caries Management Pathway. BMC Oral Health 2015, 15, S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.; Seo, B.; Friedlander, L.T.; Hussaini, H.M. A cell culture method for the isolation and study of primary human dental pulp cells. In Oral Biology: Molecular Techniques and Applications, 3rd ed.; Seymour, G.J., Cullinan, M., Heng, N., Cooper, P.R., Eds.; Springer Nature: New York, NY, USA, 2022; in press. [Google Scholar]
- Hilkens, P.; Gervois, P.; Fanton, Y.; Vanormelingen, J.; Martens, W.; Struys, T.; Politis, C.; Lambrichts, I.; Bronckaers, A. Effect of isolation methodology on stem cell properties and multilineage differentiation potential of human dental pulp stem cells. Cell Tissue Res. 2013, 353, 65–78. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbols | Gene Name | Gene Alias | Assay ID | |
|---|---|---|---|---|
| Dentinogenic markers | DMP1 | Dentine matrix acidic phosphoprotein 1 | ARHP | Hs01009392_m1 |
| DSPP | Dentine sialo phosphoprotein | DFNA39 | Hs00171962_m1 | |
| Inflammatory markers | TLR-2 | Toll like receptor 2 | TIL4 | Hs00610101_m1 |
| TLR-4 | Toll like receptor 4 | ARMD10 | Hs00152939_m1 | |
| TLR-9 | Toll like receptor 9 | CD289 | Hs00370913_s1 | |
| IL1-β | Interleukin 1 beta | IL1F2 | Hs01555410_m1 | |
| TNF-α | Tumour necrosis factor | TNFSF2 | Hs00174128_m1 | |
| IL-6 | Interleukin 6 | BSF-2 | Hs00174131_m1 | |
| IL-8 | C-X-C motif chemokine ligand 8 | IL8 | Hs00174103_m1 | |
| IL-17RA | Interleukin 17 receptor | IL17R | Hs01056316_m1 | |
| IL-17A | Interleukin 17A | IL17 | Hs00174383_m1 | |
| IL-23A | Interleukin 23 subunit alpha | IL-23 | Hs00372324_m1 | |
| Transcription markers | NFκB1 | Nuclear factor kappa B subunit 1 | NF-kappa-B | Hs00765730_m1 |
| MAPK1 | Mitogen-activated protein kinase 1 | ERK, p38, | Hs01046830_m1 | |
| Proliferative marker | MKi67 | Marker of proliferation Ki-67 | MIB-1 | Hs00606991_m1 |
| Stemness marker | SOX2 | Sex determining region Y-box 2 | ANOP3 | Hs04234836_s1 |
| Reference genes | GAPDH | Glyceraldehyde 3-phosphate dehydrogenase | G3PD | Hs02786624_g1 |
| ACTR1B | ARP1 actin-related protein 1 homolog B | ARP1B | Hs00194899_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arora, S.; Cooper, P.R.; Friedlander, L.T.; Seo, B.; Rizwan, S.B.; Rich, A.M.; Hussaini, H.M. Potentiality and Inflammatory Marker Expression Are Maintained in Dental Pulp Cell Cultures from Carious Teeth. Int. J. Mol. Sci. 2022, 23, 9425. https://doi.org/10.3390/ijms23169425
Arora S, Cooper PR, Friedlander LT, Seo B, Rizwan SB, Rich AM, Hussaini HM. Potentiality and Inflammatory Marker Expression Are Maintained in Dental Pulp Cell Cultures from Carious Teeth. International Journal of Molecular Sciences. 2022; 23(16):9425. https://doi.org/10.3390/ijms23169425
Chicago/Turabian StyleArora, Shelly, Paul R. Cooper, Lara T. Friedlander, Benedict Seo, Shakila B. Rizwan, Alison M. Rich, and Haizal Mohd Hussaini. 2022. "Potentiality and Inflammatory Marker Expression Are Maintained in Dental Pulp Cell Cultures from Carious Teeth" International Journal of Molecular Sciences 23, no. 16: 9425. https://doi.org/10.3390/ijms23169425