
Looking into the Eyes—In Vitro Models for Ocular Research

 , , , and
, , , and
Abstract
:1. Introduction
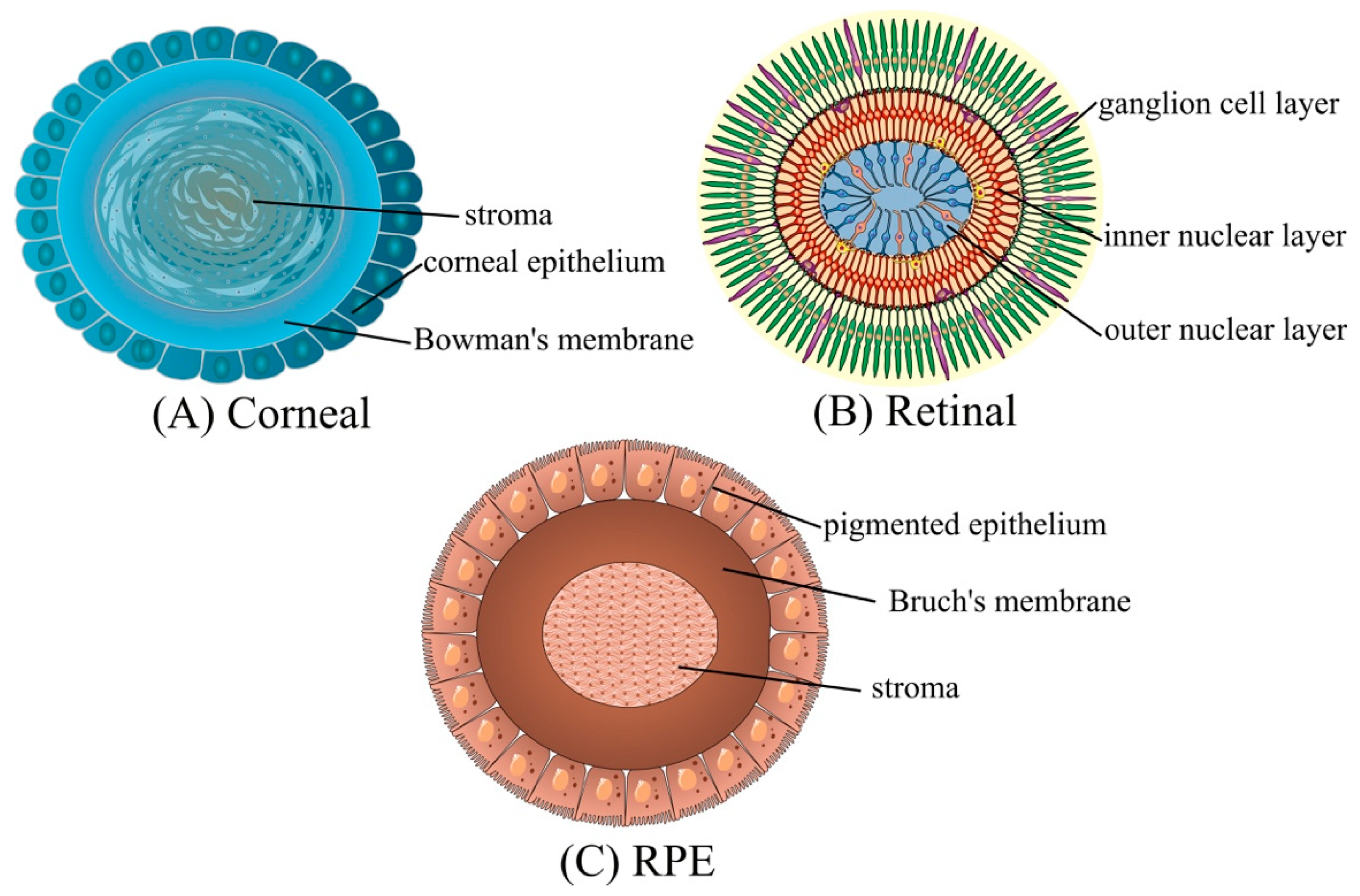
2. Eye Structure
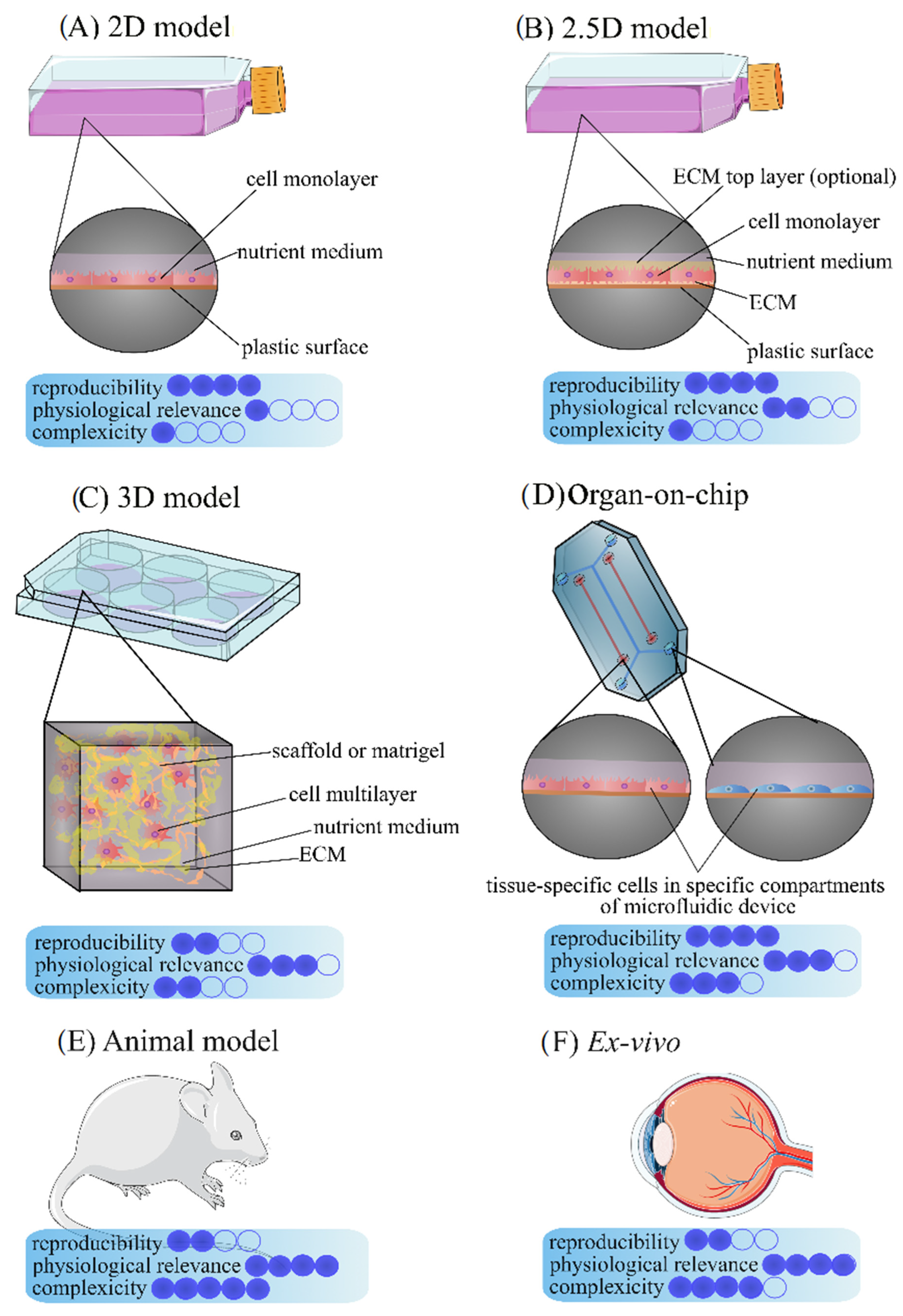
3. In Vitro Ocular Models
3.1. 2D Eye Models
3.1.1. Pigment Epithelium Cell Lines
3.1.2. Cornea Cells
3.1.3. Corneal Endothelial Cells
3.1.4. Conjunctival Cells
3.2. 3D Models
3.3. 2.5D Models
3.4. Ex Vivo Models
3.5. Spheroids, Organoids, and Organ-on-Chips
3.5.1. Spheroids
3.5.2. Organoids
3.5.3. Organ-on-Chips

{kind=link}
{kind=link}
| Spheroids | ||||
| Initial cells | Targeted cells | Culture techniques | Research possibilities | References |
| primary rabbit conjunctival epithelial cells (CECs) and lacrimal gland (LG) cell | cells that produce the aqueous and mucin layers of the tear film | an orbital shaker than Matrigel® matrix | dry eye disease | [88] |
| human RPE cells (hRPECs) obtained from donors | retinal pigment epithelium | culture in methylcellulose | Drusen-associated degeneration in the retina | [89,90] |
| human peripheral cornea | transplantable elements for limbal stem cell repopulation and limbal reconstruction | clear cornea component of the rim from a human donor | Regenerative medicine | [91,92] |
| human retoinoblastoma | human retoinoblastoma | different techniques | retinoblastoma | [93,94,95] |
| Organoids | ||||
| Initial cells | Targeted cells | Culture techniques | Research possibilities | References |
| mouse embryonic stem cell (ESCs) | mechanically rigid pigment epithelium, embryonic optic cup, stratified neural retinal tissue | G-MEM supplemented with knockout serum replacement, nonessential amino acids, pyruvate, mercaptoethanol. | development of eye, eye disorders, disease modeling, | [98] |
| human embryonic stem cells (ESCs) or human induced pluripotent stem cells (iPSCs) | after two weeks: retinal and corneal primordia, after six to eight weeks: primordia developed into minicorneas with specific morphological and marker similarities to the human cornea. | culture in a retinal differentiation medium supplemented with noggin. | neurodevelopmental disorders, disease modeling, | [99] |
| human induced pluripotent stem cells | cornea, harboring three distinct cell types with the expression of key epithelial, stromal and endothelial cell markers. | multistep procotol | investigating corneal developmental processes and their disruptions in diseased condition | [100] |
| human embryonic stem cells (ESCs), human induced pluripotent stem cells (iPSCs) | 3D ocular-like structures contains: retinal pigmented epithelium, neural retina, primitive lens and corneal-like structures. | differentiation: ventral neural induction media (VNIM) with IGF-1 | development of eye, eye disorders, disease modeling, | [102] |
| human induced pluripotent stem cells (iPSCs) | 3D multiocular organoids contans: retinal pigment epithelium, retina, and cornea. | multistep procotol | model the crosstalk between different cell types in eye development and disease | [104] |
| Organs on Chip | ||||
| Initial cells | Targeted cells | Culture techniques | Research possibilities | References |
| epithelium/stromal cells and keartnocytes isolated from rabbit eyes | microfluidic device consisting of a bilayer structure of corneal epithelial layer, a layer of stromal cells, and collagen vitrigel substrate | microfluidic devices containing collagen vitrigel (CV) | miniaturizing the standard transepithelial permeability (TEP) assay in order to measure the integrity of an array of corneal tissue micropatches. | [109] |
| immortalized human corneal epithelial cells | microengineered corneal epithelium-on-a-chip | porous membrane embedded microfluidic platform separated a chip into an apical and basal side | preclinical evaluations of potential therapeutic drugs and to mimic the environment of the human cornea. | [110] |
| human cells derived from the cornea and conjunctiva | mimic spontaneous eye blinking in humans | dome-shaped three-dimensional (3D) scaffolds in in vivo-like spatial arrangements | used for disease modeling and drug testing | [111] |
| human corneal epithelial (HCE–T) cells | dynamic cell cultivation and dynamic drug absorption testing on physiological barriers | DynaMiTES | improvement of common in vitro drug testing procedures | [112] |
3.6. In Silico Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levkovitch-Verbin, H. Animal models of optic nerve diseases. Eye 2004, 18, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Pennesi, M.E.; Neuringer, M.; Courtney, R.J. Animal models of age related macular degeneration. Mol. Asp. Med. 2012, 33, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmus, K.R. The Draize Eye Test. Surv. Ophthalmol. 2001, 45, 493–515. [Google Scholar] [CrossRef]
- Liebsch, M.; Grune, B.; Seiler, A.; Butzke, D.; Oelgeschläger, M.; Pirow, R.; Adler, S.; Riebeling, C.; Luch, A. Alternatives to animal testing: Current status and future perspectives. Arch. Toxicol. 2011, 85, 841–858. [Google Scholar] [CrossRef]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef]
- Danilova, I.; Medvedeva, S.; Shmakova, S.; Chereshneva, M.; Sarapultsev, A.; Sarapultsev, P. Pathological changes in the cellular structures of retina and choroidea in the early stages of alloxan-induced diabetes. World J. Diabetes 2018, 9, 239–251. [Google Scholar] [CrossRef]
- Pfeffer, B.A.; Philp, N.J. Cell culture of retinal pigment epithelium: Special Issue. Exp. Eye Res. 2014, 126, 1–4. [Google Scholar] [CrossRef]
- Strauss, O. The Retinal Pigment Epithelium in Visual Function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef]
- Salazar, J.J.; Ramírez, A.I.; De Hoz, R.; Salobrar-Garcia, E.; Rojas, P.; Fernández-Albarral, J.A.; López-Cuenca, I.; Rojas, B.; Triviño, A.; Ramírez, J.M. Anatomy of the Human Optic Nerve: Structure and Function; Intech: London, UK, 2019. [Google Scholar] [CrossRef]
- Komai, Y.; Ushiki, T. The three-dimensional organization of collagen fibrils in the human cornea and sclera. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2244–2258. [Google Scholar]
- Nabi, I.R.; Mathews, A.P.; Cohen-Gould, L.; Gundersen, D.; Rodriguez-Boulan, E. Immortalization of polarized rat retinal pigment epithelium. J. Cell Sci. 1993, 104, 37–49. [Google Scholar] [CrossRef]
- Badyal, D.K.; Desai, C. Animal use in pharmacology education and research: The changing scenario. Indian J. Pharmacol. 2014, 46, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R.C.; Davis, A.A. Altered expression of keratin and vimentin in human retinal pigment epithelial cells in vivo and in vitro. J. Cell. Physiol. 1990, 145, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.A.; Bernstein, P.S.; Bok, D.; Turner, J.; Nachtigal, M.; Hunt, R.C. A human retinal pigment epithelial cell line that retains epithelial characteristics after prolonged culture. Investig. Ophthalmol. Vis. Sci. 1995, 36, 955–964. [Google Scholar]
- Catanuto, P.; Espinosa-Heidmann, D.; Pereira-Simon, S.; Sanchez, P.; Salas, P.; Hernandez, E.; Cousins, S.W.; Elliot, S.J. Mouse retinal pigmented epithelial cell lines retain their phenotypic characteristics after transfection with human papilloma virus: A new tool to further the study of RPE biology. Exp. Eye Res. 2009, 88, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Zech, J.C.; Pouvreau, I.; Cotinet, A.; Goureau, O.; Le Varlet, B.; De Kozak, Y. Effect of cytokines and nitric oxide on tight junctions in cultured rat retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1600–1608. [Google Scholar]
- Chang, C.W.; Ye, L.; Defoe, D.M.; Caldwell, R.B. Serum inhibits tight junction formation in cultured pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1082–1093. [Google Scholar]
- Israel, P.; Masterson, E.; Goldman, A.I.; Wiggert, B.; Chader, G.J. Retinal pigment epithelial cell differentiation in vitro. Influence of culture medium. Investig. Ophthalmol. Vis. Sci. 1980, 19, 720–727. [Google Scholar]
- Oka, M.S.; Landers, R.A.; Bridges, C.D. A serum-free defined medium for retinal pigment epithelial cells. Exp. Cell Res. 1984, 154, 537–547. [Google Scholar] [CrossRef]
- Defoe, D.M.; Ahmad, A.; Chen, W.; Hughes, B.A. Membrane Polarity of the Na+-K+ Pump in Primary Cultures of Xenopus Retinal Pigment Epithelium. Exp. Eye Res. 1994, 59, 587–596. [Google Scholar] [CrossRef]
- Engelmann, K.; Valtink, M. RPE cell cultivation. Graefe’s Arch. Clin. Exp. Ophthalmol. 2004, 242, 65–67. [Google Scholar] [CrossRef]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, A Human Retinal Pigment Epithelial Cell Line with Differentiated Properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, G.M.; Van Rossem, M.; de Vos, A.F.; Willekens, B.; Peek, R.; Kijlstra, A. Polarized secretion of IL-6 and IL-8 by human retinal pigment epithelial cells. Clin. Exp. Immunol. 1998, 112, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, H.; Handa, J.T.; Aotaki-Keen, A.; Sherwood, S.W.; West, M.D.; Hjelmeland, L.M. Beta-galactosidase histochemistry and telomere loss in senescent retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 197–202. [Google Scholar]
- Rambhatla, L.; Chiu, C.-P.; Glickman, R.D.; Rowe-Rendleman, C. In vitro differentiation capacity of telomerase immortalized human RPE cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1622–1630. [Google Scholar]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef]
- Kuznetsova, A.V.; Kurinov, A.M.; Aleksandrova, M.A. Cell Models to Study Regulation of Cell Transformation in Pathologies of Retinal Pigment Epithelium. J. Ophthalmol. 2014, 2014, e801787. [Google Scholar] [CrossRef]
- Chadwick, B.P. Variation in Xi chromatin organization and correlation of the H3K27me3 chromatin territories to transcribed sequences by microarray analysis. Chromosoma 2007, 116, 147–157. [Google Scholar] [CrossRef]
- Culver-Cochran, A.E.; Chadwick, B.P. The WSTF-ISWI Chromatin Remodeling Complex Transiently Associates with the Human Inactive X Chromosome during Late S-Phase Prior to BRCA1 and γ-H2AX. PLoS ONE 2012, 7, e50023. [Google Scholar] [CrossRef]
- Kawazu, K.; Yamada, K.; Nakamura, M.; Ota, A. Characterization of cyclosporin A transport in cultured rabbit corneal epithelial cells: P-glycoprotein transport activity and binding to cyclophilin. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1738–1744. [Google Scholar]
- Kawazu, K.; Shiono, H.; Tanioka, H.; Ota, A.; Ikuse, T.; Takashina, H.; Kawashima, Y. Beta adrenergic antagonist permeation across cultured rabbit corneal epithelial cells grown on permeable supports. Curr. Eye Res. 1998, 17, 125–131. [Google Scholar] [CrossRef]
- Seigel, G.M. Review: R28 retinal precursor cells: The first 20 years. Mol. Vis. 2014, 20, 301–306. [Google Scholar] [PubMed]
- Nakanishi, Y.; Nakamura, M.; Mukuno, H.; Kanamori, A.; Seigel, G.M.; Negi, A. Latanoprost rescues retinal neuro-glial cells from apoptosis by inhibiting caspase-3, which is mediated by p44/p42 mitogen-activated protein kinase. Exp. Eye Res. 2006, 83, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Adamus, G.; Machnicki, M.; Seigel, G.M. Apoptotic retinal cell death induced by antirecoverin autoantibodies of cancer-associated retinopathy. Investig. Ophthalmol. Vis. Sci. 1997, 38, 283–291. [Google Scholar]
- Adamus, G.; Amundson, D.; Seigel, G.M.; Machnicki, M. Anti-Enolase-α Autoantibodies in Cancer-Associated Retinopathy: Epitope Mapping and Cytotoxicity on Retinal Cells. J. Autoimmun. 1998, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Luthra, S.; Narayanan, R.; Marques, L.E.A.; Chwa, M.; Kim, D.W.; Dong, J.; Seigel, G.M.; Neekhra, A.; Gramajo, A.L.; Brown, D.J.; et al. Evaluation of in vitro effects of bevacizumab (avastin) on retinal pigment epithelial, neurosensory retinal, and microvascular endothelial cells. Retina 2006, 26, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; Kenney, M.C.; Kamjoo, S.; Trinh, T.-H.T.; Seigel, G.M.; Resende, G.P.; Kuppermann, B.D. Toxicity of Indocyanine Green (ICG) in Combination with Light on Retinal Pigment Epithelial Cells and Neurosensory Retinal Cells. Curr. Eye Res. 2005, 30, 471–478. [Google Scholar] [CrossRef]
- Wu, X.; Reiter, C.E.N.; Antonetti, D.A.; Kimball, S.R.; Jefferson, L.S.; Gardner, T.W. Insulin Promotes Rat Retinal Neuronal Cell Survival in a p70S6K-dependent Manner. J. Biol. Chem. 2004, 279, 9167–9175. [Google Scholar] [CrossRef]
- Shen, J.; Wu, Y.; Xu, J.-Y.; Zhang, J.; Sinclair, S.H.; Yanoff, M.; Xu, G.; Li, W.; Xu, G.-T. ERK- and Akt-Dependent Neuroprotection by Erythropoietin (EPO) against Glyoxal-AGEs via Modulation of Bcl-xL, Bax, and BAD. Investig. Opthalmol. Vis. Sci. 2010, 51, 35–46. [Google Scholar] [CrossRef]
- Mukuno, H.; Nakamura, M.; Kanamori, A.; Nagai, A.; Negi, A.; Seigel, G.M. Unoprostone isopropyl rescues retinal progenitor cells from apoptosis in Vitro. Curr. Eye Res. 2004, 29, 457–464. [Google Scholar] [CrossRef]
- Van Bergen, N.J.; Wood, J.P.M.; Chidlow, G.; Trounce, I.A.; Casson, R.J.; Ju, W.-K.; Weinreb, R.N.; Crowston, J.G. Recharacterization of the RCG-5 Retinal Ganglion Cell Line. Investig. Opthalmol. Vis. Sci. 2009, 50, 4267–4272. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.R.; Clark, A.F.; Daudt, D.; Vishwanatha, J.K.; Yorio, T. A Forensic Path to RGC-5 Cell Line Identification: Lessons Learned. Investig. Opthalmol. Vis. Sci. 2013, 54, 5712–5719. [Google Scholar] [CrossRef] [PubMed]
- Wheway, G.; Nazlamova, L.; Turner, D.; Cross, S. 661W Photoreceptor Cell Line as a Cell Model for Studying Retinal Ciliopathies. Front. Genet. 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Sayyad, Z.; Sirohi, K.; Radha, V.; Swarup, G. 661W is a retinal ganglion precursor-like cell line in which glaucoma-associated optineurin mutants induce cell death selectively. Sci. Rep. 2017, 7, 16855. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, S.; Jung, Y.; Jung, E.; Kwon, H.J.; Kim, J. Eupatilin rescues ciliary transition zone defects to ameliorate ciliopathy-related phenotypes. J. Clin. Investig. 2018, 128, 3642–3648. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, M.; Mohamed, Y.H.; Onizuka, N.; Ueki, R.; Inoue, D.; Fujikawa, A.; Kitaoka, T. A novel in vivo corneal trans-epithelial electrical resistance measurement device. J. Pharmacol. Toxicol. Methods 2015, 76, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Ramaesh, K.; Dhillon, B. Ex Vivo Expansion of Corneal Limbal Epithelial/Stem Cells for Corneal Surface Reconstruction. Eur. J. Ophthalmol. 2003, 13, 515–524. [Google Scholar] [CrossRef]
- Ward, S.L.; Walker, T.L.; Dimitrijevich, S.D. Evaluation of chemically induced toxicity using an in vitro model of human corneal epithelium. Toxicol. In Vitro 1997, 11, 121–139. [Google Scholar] [CrossRef]
- Sie, N.M.; Yam, G.H.-F.; Soh, Y.Q.; Lovatt, M.; Dhaliwal, D.; Kocaba, V.; Mehta, J.S. Regenerative capacity of the corneal transition zone for endothelial cell therapy. Stem Cell Res. Ther. 2020, 11, 523. [Google Scholar] [CrossRef]
- Pistsov, M.Y.; Sadovnikova, E.Y.; Danilov, S.M. Human corneal endothelial cells: Isolation, characterization and long-term cultivation. Exp. Eye Res. 1988, 47, 403–414. [Google Scholar] [CrossRef]
- Parekh, M.; Romano, V.; Hassanin, K.; Testa, V.; Wongvisavavit, R.; Ferrari, S.; Haneef, A.; Willoughby, C.; Ponzin, D.; Jhanji, V.; et al. Biomaterials for corneal endothelial cell culture and tissue engineering. J. Tissue Eng. 2021, 12, 2041731421990536. [Google Scholar] [CrossRef]
- Götze, T.; Valtink, M.; Nitschke, M.; Gramm, S.; Hanke, T.; Engelmann, K.; Werner, C. Cultivation of an immortalized human corneal endothelial cell population and two distinct clonal subpopulations on thermo-responsive carriers. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Frausto, R.F.; Le, D.J.; Aldave, A.J. Transcriptomic Analysis of Cultured Corneal Endothelial Cells as a Validation for Their Use in Cell Replacement Therapy. Cell Transplant. 2016, 25, 1159–1176. [Google Scholar] [CrossRef] [PubMed]
- Valtink, M.; Gruschwitz, R.; Funk, R.H.W.; Engelmann, K. Two Clonal Cell Lines of Immortalized Human Corneal Endothelial Cells Show either Differentiated or Precursor Cell Characteristics. Cells Tissues Organs 2008, 187, 286–294. [Google Scholar] [CrossRef] [PubMed]
- McCabe, K.L.; Kunzevitzky, N.J.; Chiswell, B.P.; Xia, X.; Goldberg, J.L.; Lanza, R. Efficient Generation of Human Embryonic Stem Cell-Derived Corneal Endothelial Cells by Directed Differentiation. PLoS ONE 2015, 10, e0145266. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Khan, S.Y.; Gottsch, J.D.; Hutchinson, E.K.; Khan, A.; Riazuddin, S.A. Pluripotent stem cell–derived corneal endothelial cells as an alternative to donor corneal endothelium in keratoplasty. Stem Cell Rep. 2021, 16, 2320–2335. [Google Scholar] [CrossRef]
- Saha, P. A Primary Culture Model of Rabbit Conjunctival Epithelial Cells Exhibiting Tight Barrier Properties. Curr. Eye Res. 1996, 15, 1163–1169. [Google Scholar] [CrossRef]
- Yang, J.J.; Ueda, H.; Kim, K.-J.; Lee, V.H. Meeting future challenges in topical ocular drug delivery:: Development of an air-interfaced primary culture of rabbit conjunctival epithelial cells on a permeable support for drug transport studies. J. Control. Release 2000, 65, 1–11. [Google Scholar] [CrossRef]
- Civiale, C.; Paladino, G.; Marino, C.; Trombetta, F.; Pulvirenti, T.; Enea, V. Multilayer Primary Epithelial Cell Culture from Bovine Conjunctiva as a Model for in vitro Toxicity Tests. Ophthalmic Res. 2003, 35, 126–136. [Google Scholar] [CrossRef]
- Reichl, S.; Müller-Goymann, C.C. The use of a porcine organotypic cornea construct for permeation studies from formulations containing befunolol hydrochloride. Int. J. Pharm. 2003, 250, 191–201. [Google Scholar] [CrossRef]
- Scuderi, N.; Alfano, C.; Paolini, G.; Marchese, C.; Scuderi, G. Transplantation of autologous cultivated conjunctival epithelium for the restoration of defects in the ocular surface. Scand. J. Plast. Reconstr. Surg. Hand Surg. 2002, 36, 340–348. [Google Scholar] [CrossRef]
- Gipson, I.K.; Spurr-Michaud, S.; Argueso, P.; Tisdale, A.; Ng, T.F.; Russo, C.L. Mucin Gene Expression in Immortalized Human Corneal–Limbal and Conjunctival Epithelial Cell Lines. Investig. Opthalmol. Vis. Sci. 2003, 44, 2496–2506. [Google Scholar] [CrossRef] [PubMed]
- Diebold, Y.; Calonge, M.; Enríquez de Salamanca, A.; de Salamanca, A.E.; Callejo, S.; Corrales, R.M.; Sa’ez, V.; Siemasko, K.F.; Stern, M.E. Characterization of a spontaneously immortalized cell line (IOBA-NHC) from normal human conjunctiva. Investig. Opthalmol. Vis. Sci. 2003, 44, 4263–4274. [Google Scholar] [CrossRef] [PubMed]
- Barile, F.A. Validating and troubleshooting ocular invitro toxicology tests. J. Pharmacol. Toxicol. Methods 2010, 61, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Shafaie, S.; Hutter, V.; Cook, M.T.; Brown, M.B.; Chau, D.Y.S. In Vitro Cell Models for Ophthalmic Drug Development Applications. BioRes. Open Access 2016, 5, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, S.; Papantoniou, I.; Müller-Goymann, C.C. Reconstruction of an in vitro cornea and its use for drug permeation studies from different formulations containing pilocarpine hydrochloride. Eur. J. Pharm. Biopharm. 2001, 51, 119–125. [Google Scholar] [CrossRef]
- Jung, K.-M.; Lee, S.-H.; Ryu, Y.-H.; Jang, W.-H.; Jung, H.-S.; Han, J.-H.; Seok, S.-H.; Park, J.-H.; Son, Y.; Park, Y.-H.; et al. A new 3D reconstituted human corneal epithelium model as an alternative method for the eye irritation test. Toxicol. In Vitro 2011, 25, 403–410. [Google Scholar] [CrossRef]
- Kulkarni, A.; Chang, W.W.; Shen, J.; Welty, D. Use of Clonetics® Human Corneal Epithelial Cell Model for Evaluating Corneal Penetration and Hydrolysis of Ophthalmic Drug Candidates. Investig. Opthalmol. Vis. Sci. 2011, 52, 3259. [Google Scholar]
- Cotovio, J.; Grandidier, M.-H.; Lelièvre, D.; Bremond, C.; Amsellem, C.; Maloug, S.; Ovigne, J.-M.; Loisel-Joubert, S.; Van Der Lee, A.; Minondo, A.-M.; et al. In vitro assessment of eye irritancy using the Reconstructed Human Corneal Epithelial SkinEthic™ HCE model: Application to 435 substances from consumer products industry. Toxicol. In Vitro 2010, 24, 523–537. [Google Scholar] [CrossRef]
- Xiang, C.D.; Batugo, M.; Gale, D.C.; Zhang, T.; Ye, J.; Li, C.; Zhou, S.; Wu, E.Y.; Zhang, E.Y. Characterization of Human Corneal Epithelial Cell Model As a Surrogate for Corneal Permeability Assessment: Metabolism and Transport. Drug Metab. Dispos. 2009, 37, 992–998. [Google Scholar] [CrossRef]
- van Goethem, F.; Adriaens, E.; Alépée, N.; Straube, F.; De Wever, B.; Cappadoro, M.; Catoire, S.; Hansen, E.; Wolf, A.; Vanparys, P. Prevalidation of a new in vitro reconstituted human cornea model to assess the eye irritating potential of chemicals. Toxicol. In Vitro 2006, 20, 1–17. [Google Scholar] [CrossRef]
- de Brugerolle, A. SkinEthic Laboratories, a Company Devoted to Develop and Produce In Vitro Alternative Methods to Animal Use. ALTEX-Altern. Anim. Exp. 2007, 24, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Hamajima, F.; Ogasawara, T.; Hata, K.-I. Establishment of a new in vitro test method for evaluation of eye irritancy using a reconstructed human corneal epithelial model, LabCyte CORNEA-MODEL. Toxicol. In Vitro 2013, 27, 2184–2192. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Uemura, N.; Hamajima, F.; Ogasawara, T.; Hata, K. ichiro Morphological Characterization of a Reconstructed Human Corneal Epithelial Model (LabCyte CORNEA-MODEL) as an Alternative to the Draize Eye Test for the Assessment of Eye Irritation. Altern. Anim. Test. Exp. 2012, 17, 2–8. [Google Scholar] [CrossRef]
- Li, X.; Lu, Y.; Deng, Z. Recombinant Cornea Model for Ocular Surface Irritation Evaluation and Preparation Method Thereof. Patent No: CN110066847A, 2019. Available online: https://worldwide.espacenet.com/patent/search/family/067364436/publication/CN110066847A?q=%22Recombinant%20Cornea%20Model%20for%20Ocular%20Surface%20Irritation%20Evaluation%20and%20Preparation%22 (accessed on 5 April 2022).
- Zuguo, L.; Hui, L.; Wei, L.; Yangluowa, Q. Construction Method for Dry Eye Model In Vitro. Patent No: CN101508974B, 2009. Available online: https://worldwide.espacenet.com/patent/search/family/041001507/publication/CN101508974B?q=%22Construction%20Method%20for%20Dry%20Eye%20Model%20in%20Vitro%22 (accessed on 5 April 2022).
- Minami, Y.; Sugihara, H.; Oono, S. Reconstruction of cornea in three-dimensional collagen gel matrix culture. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2316–2324. [Google Scholar]
- Schneider, A.I.; Maier-Reif, K.; Graeve, T. Constructing an in vitro cornea from cultures of the three specific corneal cell types. In Vitro Cell. Dev. Biol. Anim. 1999, 35, 515–526. [Google Scholar] [CrossRef]
- Zieske, J.D.; Mason, V.S.; Wasson, M.E.; Meunier, S.F.; Nolte, C.J.; Fukai, N.; Olsen, B.R.; Parenteau, N.L. Basement Membrane Assembly and Differentiation of Cultured Corneal Cells: Importance of Culture Environment and Endothelial Cell Interaction. Exp. Cell Res. 1994, 214, 621–633. [Google Scholar] [CrossRef]
- Shamir, E.R.; Ewald, A.J. Three-dimensional organotypic culture: Experimental models of mammalian biology and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 647–664. [Google Scholar] [CrossRef]
- Tchao, R. Trans-Epithelial Permeability of Fluorescein in Vitro as an Assay to Determine Eye Irritants. Altern. Methods Toxicol. 1988, 6, 271–283. [Google Scholar]
- van den Berghe, C.; Guillet, M.C.; Compan, D. Performance of porcine corneal opacity and permeability assay to predict eye irritation for water-soluble cosmetic ingredients. Toxicol. In Vitro 2005, 19, 823–830. [Google Scholar] [CrossRef]
- Huh, D.; Hamilton, G.A.; Ingber, D.E. From 3D cell culture to organs-on-chips. Trends Cell Biol. 2011, 21, 745–754. [Google Scholar] [CrossRef]
- Yu, F.-S.X. Corneal Organ Culture System for Chemical Toxicity Tests of Consumer Products. Patent No: WO 99/51763, 1999. Available online: https://patentimages.storage.googleapis.com/c8/4a/0d/d3b3a03378b3e4/WO1999051763A1.pdf (accessed on 17 May 2022).
- Ryu, N.-E.; Lee, S.-H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef] [PubMed]
- Matak, D.; Brodaczewska, K.K.; Lipiec, M.; Szymanski, Ł.; Szczylik, C.; Czarnecka, A.M. Colony, hanging drop, and methylcellulose three dimensional hypoxic growth optimization of renal cell carcinoma cell lines. Cytotechnology 2017, 69, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, S.; Cohen-Harazi, R.; Maizels, Y.; Koman, I. Patient-derived tumor spheroid cultures as a promising tool to assist personalized therapeutic decisions in breast cancer. Transl. Cancer Res. 2022, 11, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Yin, H.; Grant, M.P.; Elisseeff, J.H. An In Vitro Model for the Ocular Surface and Tear Film System. Sci. Rep. 2017, 7, 6163. [Google Scholar] [CrossRef]
- Sato, R.; Yasukawa, T.; Kacza, J.; Eichler, W.; Nishiwaki, A.; Iandiev, I.; Ohbayashi, M.; Kato, A.; Yafai, Y.; Bringmann, A.; et al. Three-Dimensional Spheroidal Culture Visualization of Membranogenesis of Bruch’s Membrane and Basolateral Functions of the Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2013, 54, 1740–1749. [Google Scholar] [CrossRef]
- Usui, H.; Nishiwaki, A.; Landiev, L.; Kacza, J.; Eichler, W.; Wako, R.; Kato, A.; Takase, N.; Kuwayama, S.; Ohashi, K.; et al. In vitro drusen model: Three-dimensional spheroid culture of retinal pigment epithelial cells. J. Cell Sci. 2018, 132, jcs.215798. [Google Scholar] [CrossRef]
- Yoon, J.J.; Wang, E.F.; Ismail, S.; McGhee, J.J.; Sherwin, T. Sphere-forming cells from peripheral cornea demonstrate polarity and directed cell migration. Cell Biol. Int. 2013, 37, 949–960. [Google Scholar] [CrossRef]
- Mathan, J.J.; Ismail, S.; McGhee, J.J.; McGhee, C.N.J.; Sherwin, T. Sphere-forming cells from peripheral cornea demonstrate the ability to repopulate the ocular surface. Stem Cell Res. Ther. 2016, 7, 81. [Google Scholar] [CrossRef]
- Winter, U.; Aschero, R.; Fuentes, F.; Buontempo, F.; Zugbi, S.; Sgroi, M.; Sampor, C.; Abramson, D.H.; Carcaboso, A.M.; Schaiquevich, P. Tridimensional Retinoblastoma Cultures as Vitreous Seeds Models for Live-Cell Imaging of Chemotherapy Penetration. Int. J. Mol. Sci. 2019, 20, 1077. [Google Scholar] [CrossRef]
- Bond, W.S.; Akinfenwa, P.Y.; Perlaky, L.; Hurwitz, M.Y.; Hurwitz, R.L.; Chévez-Barrios, P. Tumorspheres but Not Adherent Cells Derived from Retinoblastoma Tumors Are of Malignant Origin. PLoS ONE 2013, 8, e63519. [Google Scholar] [CrossRef]
- Tang, Z.; Ma, H.; Mao, Y.; Ai, S.; Zhang, P.; Nie, C.; Gao, Y.; Lu, R. Identification of stemness in primary retinoblastoma cells by analysis of stem-cell phenotypes and tumorigenicity with culture and xenograft models. Exp. Cell Res. 2019, 379, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Manafi, N.; Shokri, F.; Achberger, K.; Hirayama, M.; Mohammadi, M.H.; Noorizadeh, F.; Hong, J.; Liebau, S.; Tsuji, T.; Quinn, P.M.J.; et al. Organoids and organ chips in ophthalmology. Ocul. Surf. 2020, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Eiraku, M.; Takata, N.; Ishibashi, H.; Kawada, M.; Sakakura, E.; Okuda, S.; Sekiguchi, K.; Adachi, T.; Sasai, Y. Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 2011, 472, 51–56. [Google Scholar] [CrossRef]
- Susaimanickam, P.J.; Kiral, F.R.; Park, I.-H. Region Specific Brain Organoids to Study Neurodevelopmental Disorders. Int. J. Stem Cells 2022, 15, 26–40. [Google Scholar] [CrossRef]
- Foster, J.W.; Wahlin, K.; Adams, S.M.; Birk, D.E.; Zack, D.J.; Chakravarti, S. Cornea organoids from human induced pluripotent stem cells. Sci. Rep. 2017, 7, 41286. [Google Scholar] [CrossRef]
- Mellough, C.B.; Sernagor, E.; Moreno-Gimeno, I.; Steel, D.H.W.; Lako, M. Efficient Stage-Specific Differentiation of Human Pluripotent Stem Cells Toward Retinal Photoreceptor Cells. Stem Cells 2012, 30, 673–686. [Google Scholar] [CrossRef]
- Mellough, C.B.; Collin, J.; Khazim, M.; White, K.; Sernagor, E.; Steel, D.H.W.; Lako, M. IGF-1 Signaling Plays an Important Role in the Formation of Three-Dimensional Laminated Neural Retina and Other Ocular Structures From Human Embryonic Stem Cells. Stem Cells 2015, 33, 2416–2430. [Google Scholar] [CrossRef]
- Mellough, C.B.; Collin, J.; Queen, R.; Hilgen, G.; Dorgau, B.; Zerti, D.; Felemban, M.; White, K.; Sernagor, E.; Lako, M. Systematic Comparison of Retinal Organoid Differentiation from Human Pluripotent Stem Cells Reveals Stage Specific, Cell Line, and Methodological Differences. Stem Cells Transl. Med. 2019, 8, 694–706. [Google Scholar] [CrossRef]
- Isla-Magrané, H.; Veiga, A.; García-Arumí, J.; Duarri, A. Multiocular organoids from human induced pluripotent stem cells displayed retinal, corneal, and retinal pigment epithelium lineages. Stem Cell Res. Ther. 2021, 12, 581. [Google Scholar] [CrossRef]
- Mazerik, J.N.; Becker, S.; Sieving, P.A. 3-D retina organoids. Cell Med. 2018, 10, 2155179018773758. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, M.; Chichagova, V.; Hilgen, G.; Dorgau, B.; Sernagor, E.; Armstrong, L.; Lako, M. Room temperature shipment does not affect the biological activity of pluripotent stem cell-derived retinal organoids. PLoS ONE 2020, 15, e0233860. [Google Scholar] [CrossRef] [PubMed]
- Chichagova, V.; Hallam, D.; Nicholds, M.; Thomas, R.; Lako, M.; Armstrong, L. Using Statistical Models to Identify Key Variavles Critical for Generation of Retinal Organoids from IPSC. Investig. Opthalmol. Vis. Sci. 2018, 59, 565. [Google Scholar]
- National Centre for Replacement Refinement & Reduction of Animals in Research Retinal 3D. Available online: https://nc3rs.org.uk/crackit/retinal-3d (accessed on 21 May 2022).
- Puleo, C.M.; Ambrose, W.M.; Takezawa, T.; Elisseeff, J.; Wang, T.-H. Integration and application of vitrified collagen in multilayered microfluidic devices for corneal microtissue culture. Lab Chip 2009, 9, 3221–3227. [Google Scholar] [CrossRef] [PubMed]
- Bennet, D.; Estlack, Z.; Reid, T.; Kim, J. A microengineered human corneal epithelium-on-a-chip for eye drops mass transport evaluation. Lab Chip 2018, 18, 1539–1551. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Byun, W.Y.; Frank, A.; Massaro-Giordano, M.; Lee, V.; Bunya, V.Y.; Huh, D. A Human Blinking “Eye-on-a-Chip”. Investig. Opthalmol. Vis. Sci. 2016, 57, 3872. [Google Scholar]
- Seo, J.; Byun, W.Y.; Alisafaei, F.; Georgescu, A.; Yi, Y.-S.; Massaro-Giordano, M.; Shenoy, V.B.; Lee, V.; Bunya, V.Y.; Huh, D. Multiscale reverse engineering of the human ocular surface. Nat. Med. 2019, 25, 1310–1318. [Google Scholar] [CrossRef]
- Beiβner, N.; Mattern, K.; Dietzel, A.; Reichl, S. DynaMiTES—A dynamic cell culture platform for in vitro drug testing PART 2 —Ocular DynaMiTES for drug absorption studies of the anterior eye. Eur. J. Pharm. Biopharm. 2018, 126, 166–176. [Google Scholar] [CrossRef] [PubMed]
- In Silico Cancer Research towards 3R-PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5897933/ (accessed on 22 April 2022).
- Vincze, A.; Dargó, G.; Rácz, A.; Balogh, G.T. A corneal-PAMPA-based in silico model for predicting corneal permeability. J. Pharm. Biomed. Anal. 2021, 203, 114218. [Google Scholar] [CrossRef]
- Trone, M.-C.; Campolmi, N.; Gauthier, A.-S.; Nefzaoui, C.; Bernard, A.; Naigeon, N.; Nangoum-Fosso, T.; Piselli, S.; Garraud, O.; Acquart, S.; et al. Conception and optimization of a corneal bioreactor. Acta Ophthalmol. 2013, 91. [Google Scholar] [CrossRef]
- Martin, I.; Wendt, D.; Heberer, M. The role of bioreactors in tissue engineering. Trends Biotechnol. 2004, 22, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Alfonsetti, M.; Castelli, V.; D’Angelo, M.; Benedetti, E.; Allegretti, M.; Barboni, B.; Cimini, A. Looking for In Vitro Models for Retinal Diseases. Int. J. Mol. Sci. 2021, 22, 10334. [Google Scholar] [CrossRef] [PubMed]
- Hornof, M.; Toropainen, E.; Urtti, A. Cell culture models of the ocular barriers. Eur. J. Pharm. Biopharm. 2005, 60, 207–225. [Google Scholar] [CrossRef]
- Achberger, K.; Probst, C.; Haderspeck, J.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging organoid and organ-on-a-chip technology to generate complex multi-layer tissue models in a human retina-on-a-chip platform. eLife 2019, 8, e46188. [Google Scholar] [CrossRef] [PubMed]
- O’Hara-Wright, M.; Gonzalez-Cordero, A. Retinal organoids: A window into human retinal development. Development 2020, 147, dev189746. [Google Scholar] [CrossRef]


| Membrane | Part of Membrane | Structure and Function |
|---|---|---|
| Fibrous tunic | Sclera |
|
| Cornea |
| |
| Vascular anteriorly | Iris |
|
| Ciliary body |
| |
| Choroid |
| |
| Nervous | Retina |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lieto, K.; Skopek, R.; Lewicka, A.; Stelmasiak, M.; Klimaszewska, E.; Zelent, A.; Szymański, Ł.; Lewicki, S. Looking into the Eyes—In Vitro Models for Ocular Research. Int. J. Mol. Sci. 2022, 23, 9158. https://doi.org/10.3390/ijms23169158
Lieto K, Skopek R, Lewicka A, Stelmasiak M, Klimaszewska E, Zelent A, Szymański Ł, Lewicki S. Looking into the Eyes—In Vitro Models for Ocular Research. International Journal of Molecular Sciences. 2022; 23(16):9158. https://doi.org/10.3390/ijms23169158
Chicago/Turabian StyleLieto, Krystyna, Rafał Skopek, Aneta Lewicka, Marta Stelmasiak, Emilia Klimaszewska, Arthur Zelent, Łukasz Szymański, and Sławomir Lewicki. 2022. "Looking into the Eyes—In Vitro Models for Ocular Research" International Journal of Molecular Sciences 23, no. 16: 9158. https://doi.org/10.3390/ijms23169158