
Cellular Redox Metabolism Is Modulated by the Distinct Localization of Cyclic Nucleotide Phosphodiesterase 5A Isoforms

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
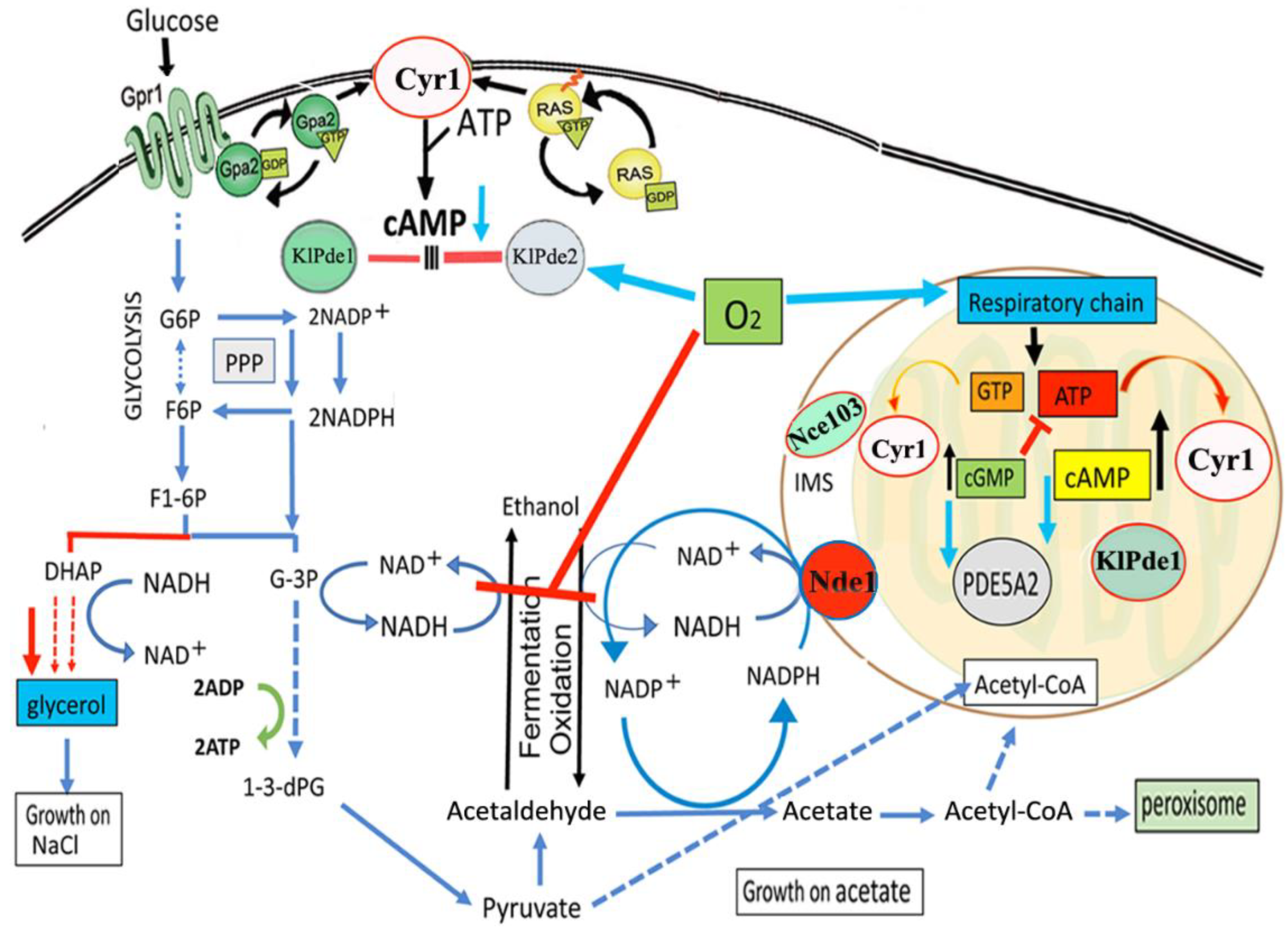
2. Results
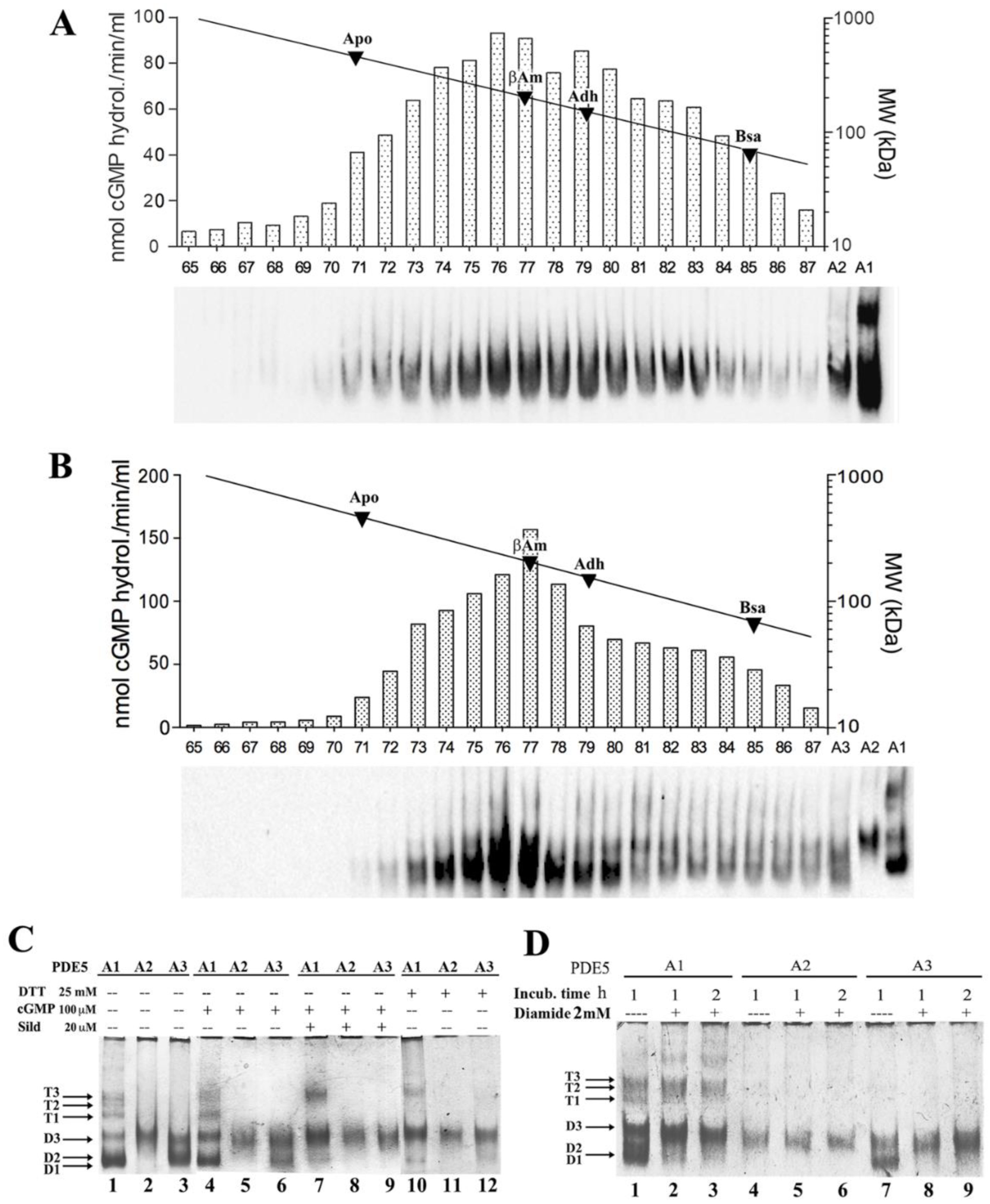
2.1. Quaternary Structure of Recombinant MmPDE5A Isoforms
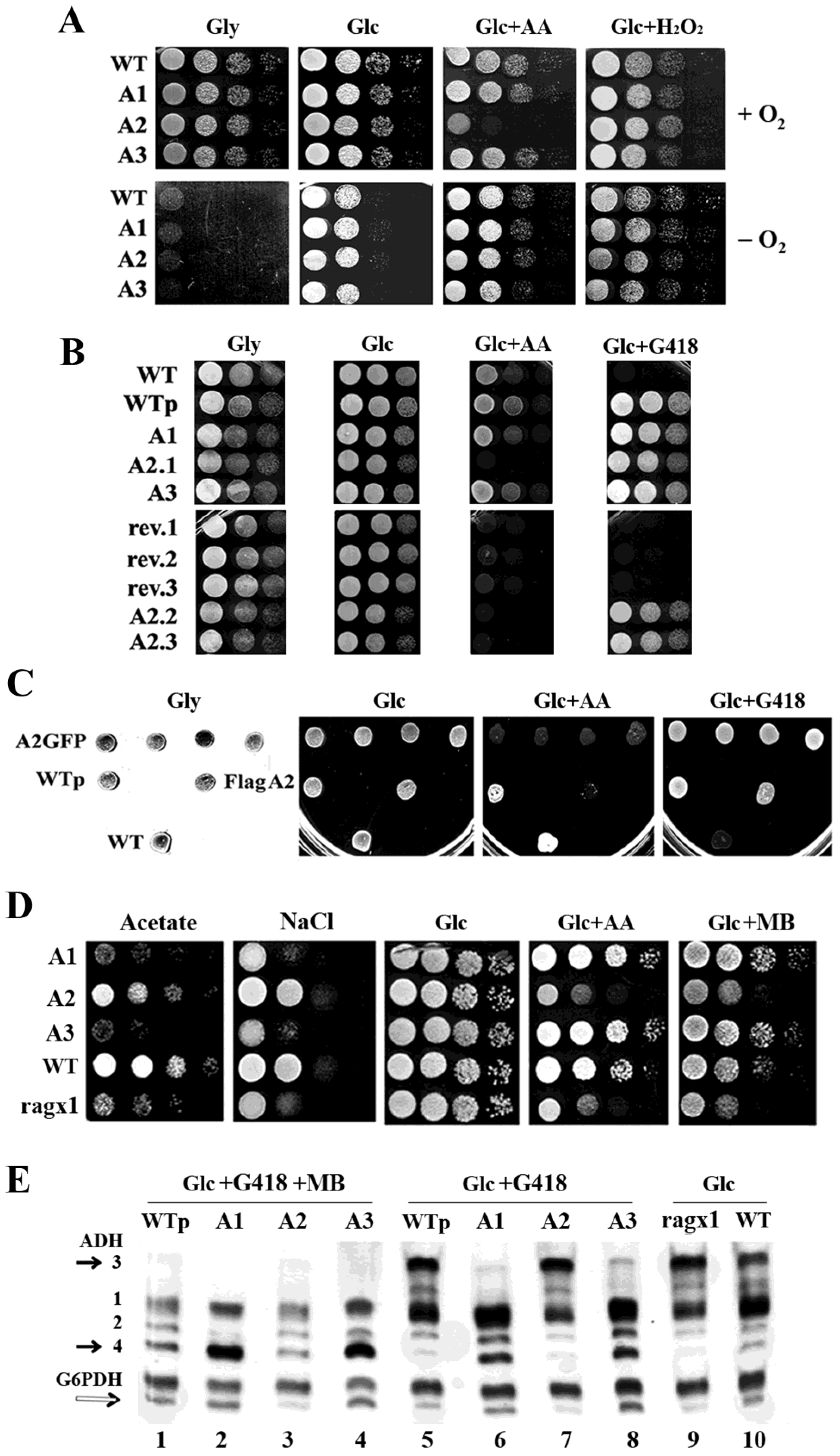
2.2. Overexpression of MmPde5a2 Induces a Mutation That Affects Glucose Oxidation in K. lactis
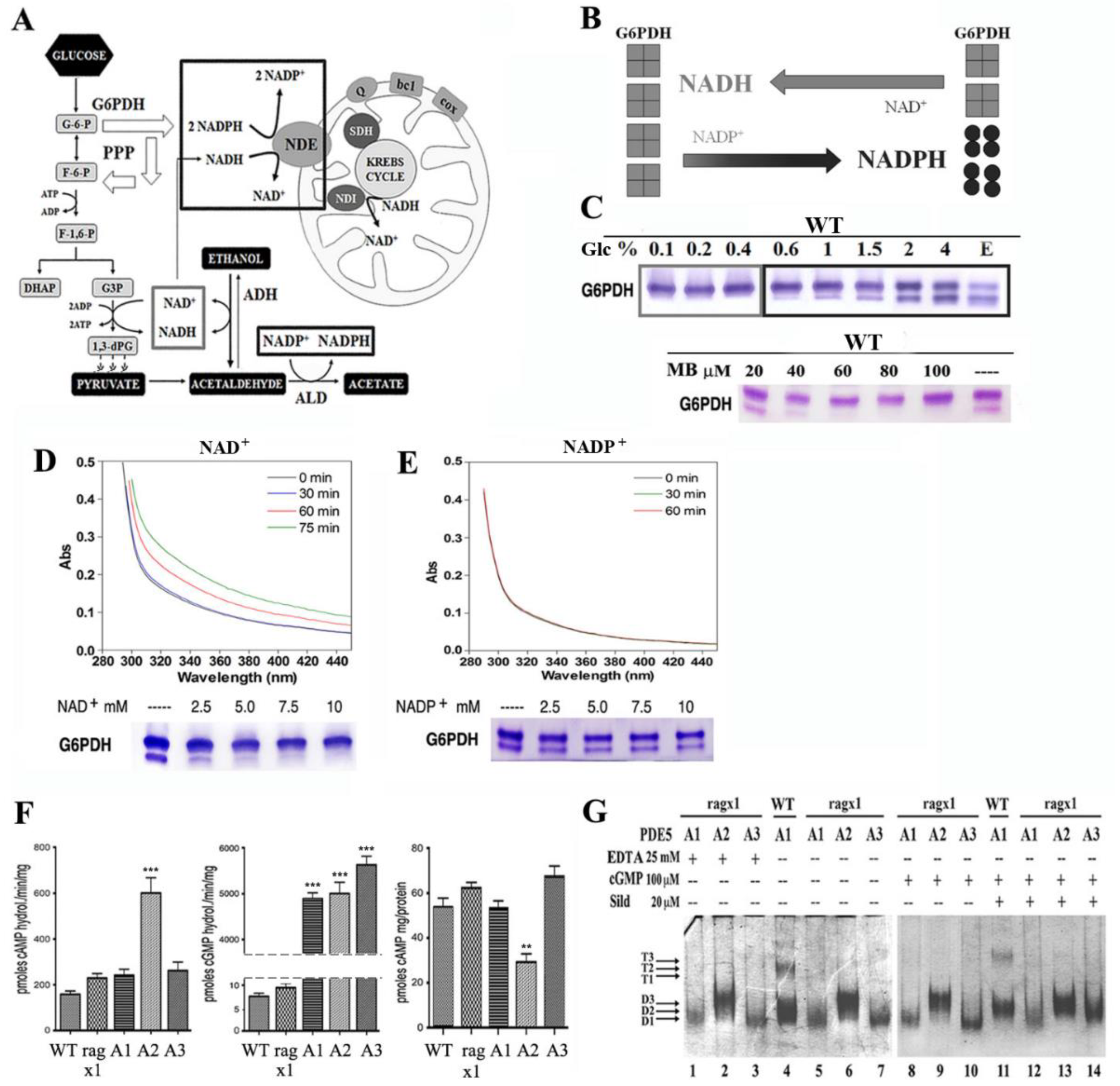
2.3. Overexpression of PDE5A Isoforms Affects the Cytosolic NAD+/NADH–NADP+/NADPH Redox Balance
2.4. Over-Expression of PDE5A2 Affects Both cAMP Content and cAMP-Hydrolyzing Activity
2.5. The ragx1 Mutant Context Modifies the Quaternary Structure of MmPDE5A Isoforms
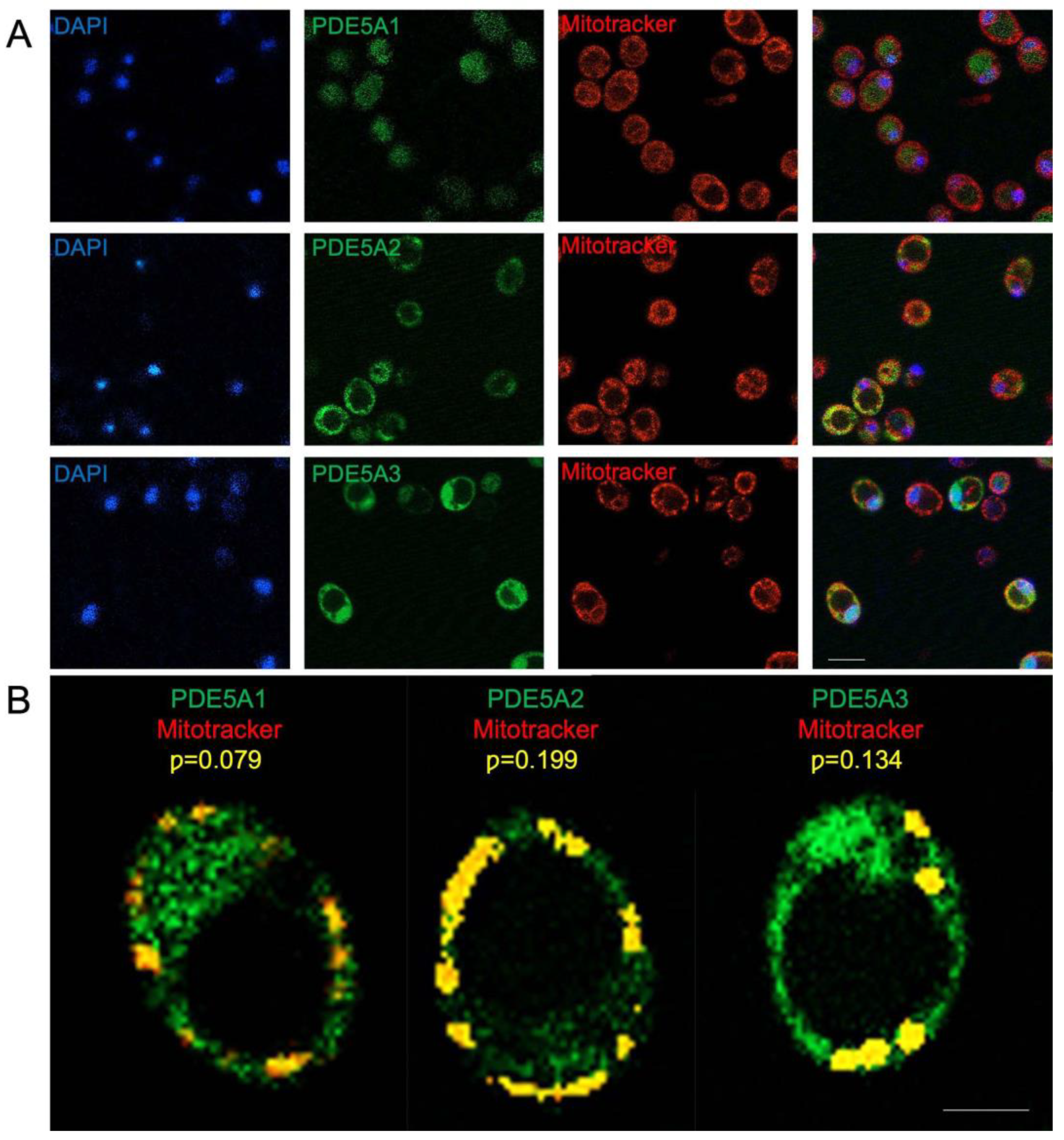
2.6. MmPDE5A2 Is Localized in the Mitochondria
3. Discussion
3.1. Quaternary Assembly and Cellular Localization of MmPDE5 Isoforms
3.2. Metabolic Role of MmPDE5A2 in K. lactis
4. Materials and Methods
4.1. Strains, Media, Culture Conditions, and Vectors
4.2. Size Exclusion Chromatography (SEC)
4.3. PDE Enzymatic Assays
4.4. Native Gel Electrophoresis Analysis
4.5. Native Western Blot Analysis
4.6. Alcohol Dehydrogenase (ADH) and Glucose 6-Phosphate Dehydrogenase (G6PDH) Native Assays
4.7. Determination of cAMP Content
4.8. Cellular Localization by Fluorescence Microscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Beavo, J.A.; Conti, M.; Heaslip, R.J. Multiple cyclic nucleotide phosphodiesterases. Mol. Pharmacol. 1994, 46, 399–405. [Google Scholar] [PubMed]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 3, 488–520. [Google Scholar] [CrossRef]
- Keravis, T.; Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) isozymes as targets of the intracellular signalling network: Benefits of PDE inhibitors in various diseases and perspectives for future therapeutic development. Br. J. Pharmacol. 2012, 165, 1288–1305. [Google Scholar] [CrossRef] [Green Version]
- Maurice, D.H.; Ke, H.; Ahmad, F.; Wang, Y.; Chung, J.; Manganiello, V.C. Advances in targeting cyclic nucleotide phosphodiesterases. Nat. Rev. Drug Discov. 2014, 13, 290–314. [Google Scholar] [CrossRef] [Green Version]
- Omori, K.; Kotera, J. Overview of PDEs and their regulation. Circ. Res. 2007, 100, 309–327. [Google Scholar] [CrossRef]
- Pandit, J.; Forman, M.D.; Fennell, K.F.; Dillman, K.S.; Menniti, F.S. Mechanism for the allosteric regulation of phosphodiesterase 2A deduced from the X-ray structure of a near full-length construct. Proc. Natl. Acad. Sci. USA 2009, 106, 18225–18230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.S.; Lin, G.; Xin, Z.C.; Lue, T.F. Expression, distribution and regulation of phosphodiesterase 5. Curr. Pharm. Des. 2006, 12, 3439–3457. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.F.; Faucz, F.R.; Bimpaki, E.; Horvath, A.; Levy, I.; de Alexandre, R.B.; Ahmad, F.; Manganiello, V.; Stratakis, C.A. Clinical and molecular genetics of the phosphodiesterases (PDEs). Endocr. Rev. 2014, 35, 19–23. [Google Scholar] [CrossRef]
- Lugnier, C.; Schoeffter, P.; le Bec, A.; Strouthou, E.; Stoclet, J.C. Selective inhibition of cyclic nucleotide phosphodiesterase of human, bovine and rat aorta. Biochem. Pharmacol. 1986, 35, 1743–1751. [Google Scholar] [CrossRef]
- Rybalkin, S.D.; Rybalkina, I.G.; Feil, R.; Hofmann, F.; Beavo, J.A. Regulation of cGMP-specific phosphodiesterase (PDE5) phosphorylation in smooth muscle cells. J. Biol. Chem. 2002, 277, 3310–3317. [Google Scholar] [CrossRef] [Green Version]
- Ioakeimidis, N.; Kostos, J.B. Pharmacologic therapy for erectile dysfunction and its interaction with the cardiovascular system. J. Cardiovasc. Pharmacol. Ther. 2014, 19, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; de Stefano, M.E.; Citro, G.; Modica, A.; Giorgi, M. Expression of cGMP-binding cGMP-specific phosphodiesterase (PDE5) in mouse tissues and cell lines using an antibody against the enzyme amino-terminal domain. Biochim. Biophys. Acta 2001, 1539, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Prickaerts, J.; Sik, A.; van Staveren, W.C.; Koopmans, G.; Steinbusch, H.W.; van der Staay, F.J.; de Vente, J.; Blokland, A. Phosphodiesterase type 5 inhibition improves early memory consolidation of object information. Neurochem. Int. 2004, 45, 915–928. [Google Scholar] [CrossRef]
- Tsai, E.J.; Kass, D.A. Cyclic GMP signaling in cardiovascular pathophysiology and therapeutics. Pharmacol. Ther. 2009, 122, 216–238. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, M.; Pompili, A.; Cardarelli, S.; Castelli, V.; Biagioni, S.; Sancesario, G.; Gasbarri, A. Zaprinast impairs spatial memory by increasing PDE5 expression in the rat hippocampus. Behav. Brain. Res. 2015, 278, 129–136. [Google Scholar] [CrossRef]
- Sponziello, M.; Verrienti, A.; Rosignolo, F.; de Rose, R.F.; Pecce, V.; Maggisano, V.; Durante, C.; Bullotta, S.; Damante, G.; Giacomelli, L.; et al. PDE5 expression in human thyroid tumors and effects of PDE5 inhibitors on growth and migration of cancer cells. Endocrine 2015, 50, 434–441. [Google Scholar] [CrossRef]
- Giannetta, E.; Isidori, A.; Galea, N.; Carbone, I.; Mandosi, E.; Vizza, C.D.; Naro, F.; Morano, S.; Fedele, F.; Lenzi, A. Chronic inhibition of cGMP phosphodiesterase5A improves diabetic cardiomyopathy. A randomized, controlled clinical trial using magnetic resonance imaging with myocardial tagging. Circulation 2012, 125, 2323–2333. [Google Scholar] [CrossRef] [Green Version]
- Campolo, F.; Pofi, R.; Venneri, M.A.; Isidori, A.M. Priming metabolism with the type 5 phosphodiesterase: The role of cGMP-hydrolyzing enzymes. Curr. Opin. Pharmacol. 2021, 60, 298–305. [Google Scholar] [CrossRef]
- Lin, C.S.; Lau, A.; Tu, R.; Lue, T.F. Expression of three isoforms of cGMP-binding cGMP-specific phosphodiesterase (PDE5) in human penile cavernosum. Biochem. Biophys. Res. Commun. 2000, 268, 628–635. [Google Scholar] [CrossRef]
- Campolo, F.; Zevini, A.; Cardarelli, S.; Monaco, L.; Barbagallo, F.; Pellegrini, M.; Cornacchione, M.; di Grazia, A.; de Arcangelis, V.; Gianfrilli, D.; et al. Identification of murine phosphodiesterase 5A isoforms and their functional characterization in HL-1 cardiac cell line. J. Cell. Physiol. 2018, 233, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Cardarelli, S.; Giorgi, M.; Naro, F.; Malatesta, F.; Biagioni, S.; Saliola, M. Use of the KlADH3 promoter for the quantitative production of the murine PDE5A isoforms in the yeast Kluyveromyces lactis. Microb. Cell Fact. 2017, 16, 159. [Google Scholar] [CrossRef] [Green Version]
- Cardarelli, S.; Miele, A.E.; Zamparelli, C.; Biagioni, S.; Naro, F.; Malatesta, F.; Giorgi, M.; Saliola, M. The oligomeric assembly of the phosphodiesterase-5 is a mixture of dimers and tetramers: A putative role in the regulation of function. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2183–2190. [Google Scholar] [CrossRef] [PubMed]
- Cardarelli, S.; Giorgi, M.; Poiana, G.; Biagioni, S.; Saliola, M. Metabolic role of cGMP in S. cerevisiae: The murine phosphodiesterase-5 activity affects yeast cell proliferation by altering the cAMP/cGMP equilibrium. FEMS Yeast Res. 2019, 19, foz016. [Google Scholar] [CrossRef]
- Giorgi, M.; Cardarelli, S.; Ragusa, F.; Saliola, M.; Biagioni, S.; Poiana, G.; Naro, F.; Massimi, M. Phosphodiesterase Inhibitors: Could They Be Beneficial for the Treatment of COVID-19? Int. J. Mol. Sci. 2020, 21, 5338. [Google Scholar] [CrossRef]
- Corbin, J.D.; Zoraghi, R.; Francis, S.H. Allosteric-site and catalytic-site ligand effect on PDE5 functions are associated with distinct changes in physical form of the enzyme. Cell Signal. 2009, 21, 1768–1774. [Google Scholar] [CrossRef] [Green Version]
- Rodicio, R.; Heinisch, J.J. Yeast on the milky way: Genetics, physiology and biotechnology of Kluyveromyces lactis. Yeast 2013, 30, 165–177. [Google Scholar] [CrossRef] [PubMed]
- De Deken, R.H. The Crabtree effect: A regulatory system in yeast. J. Gen. Microbiol. 1966, 44, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Goffrini, P.; Algeri, A.A.; Donnini, C.; Wésolowski-Louvel, M.; Ferrero, I. RAG1 and RAG2: Nuclear genes involved in the dependence/independence on mitochondrial respiratory function for growth on sugar. Yeast 1981, 5, 99–105. [Google Scholar] [CrossRef]
- Wésolowski–Louvel, M.; Prior, C.; Bornecque, D.; Fukuhara, H. Rag2 mutations involved in glucose metabolism in yeast: Isolation and genetic characterization. Yeast 1992, 8, 711–719. [Google Scholar] [CrossRef]
- Neil, H.; Lemaire, M.; Wésolowski-Louvel, M. Regulation of glycolysis in Kluyveromyces lactis: Role of KlGCR1 and KlGCR2 in glucose uptake and catabolism. Curr. Genet. 2004, 45, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Cialfi, S.; Uccelletti, D.; Carducci, A.; Wésolowski-Louvel, M.; Mancini, P.; Heipieper, H.J.; Saliola, M. KlHsl1 is a component of glycerol response pathways in the milk yeast Kluyveromyces lactis. Microbiology 2011, 157, 1509–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, R.H.; Adler, H.; Pickhardt, M.; Mandelkow, E. Lest we forget you—Methylene blue… Neurobiol. Aging 2011, 32, 2325.e7–2325.e16. [Google Scholar]
- Mayer, B.; Brunner, F.; Schmidt, K. Inhibition of nitric oxide synthesis by methylene blue. Biochem. Pharmacol. 1993, 45, 367–374. [Google Scholar] [CrossRef]
- Santangelo, G.M. Glucose signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 7, 253–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliola, M.; Falcone, C. Two mitochondrial alcohol dehydrogenase activities of Kluyveromyces lactis are differentially expressed during respiration and fermentation. Mol. Gen. Genet. 1995, 249, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Saliola, M.; Tramonti, A.; Lanini, C.; Cialfi, S.; de Biase, D.; Falcone, C. Intracellular NADPH levels affect the oligomeric state of the glucose 6-phosphate dehydrogenase Eukaryot. Cell 2012, 11, 1503–1511. [Google Scholar]
- Tramonti, A.; Saliola, M. Glucose 6-phosphate and alcohol dehydrogenase activities are components of dynamic macromolecular depots structures. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Siso, M.I.; Freire-Picos, M.A.; Ramil, E.; Gonzalez-Domınguez, M.; Torres, A.R.; Cerdan, M.E. Respirofermentative metabolism in Kluyveromyces lactis: Insights and perspectives. Enzym. Microb. Technol. 2000, 26, 699–705. [Google Scholar] [CrossRef]
- Tarrío, N.; Becerra, M.; Cerdán, M.E.; Siso, M.I.G. Reoxidation of cytosolic NADPH in Kluyveromyces lactis. FEMS Yeast Res. 2006, 6, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Saliola, M.; D’Amici, S.; Sponziello, M.; Mancini, P.; Tassone, P.; Falcone, C. The transdehydrogenase genes KlNDE1 and KlNDI1 regulate the expression of KlGUT2 in the yeast Kluyveromyces lactis. FEMS Yeast Res. 2010, 10, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Overkamp, K.M.; Bakker, B.M.; Steensma, H.Y.; van Dijken, J.P.; Pronk, J.T. Two mechanisms for oxidation of cytosolic NADPH by Kluyveromyces lactis mitochondria. Yeast 2002, 19, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Acin-Perez, R.; Russwurm, M.; Guennewig, K.; Gertz, M.; Zoidi, G.; Ramos, L.; Buck, J.; Levin, L.R.; Rassow, J.; Manfredi, G.; et al. A phosphodiesterase 2A isoform localized to mitochondria regulates respiration. J. Biol. Chem. 2011, 286, 30423–30432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claros, M.G.; Vincens, P. Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur. J. Biochem. 1996, 241, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Siso, M.I.G.; Garcìa-Leiro, A.; Tarrio, N.; Becerra, M.; Cerdan, M.E. Sugar metabolism redox balance and oxidative stress response in the respiratory yeast Kluyveromyces lactis. Microb. Cell Fact. 2009, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Belotti, F.; Tisi, R.; Paiardi, C.; Rigamonti, M.; Groppi, S.; Martegani, E. Localization of Ras signaling complex in budding yeast. Biochim. Biophys. Acta 2012, 1823, 1208–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Cui, W.; Huang, M.; Robinson, H.; Wan, Y.; Wang, Y.; Ke, H. Dual specificity and novel structural folding of yeast phosphodiesterase-1 for hydrolysis of second messengers cyclic adenosine and guanosine 3′,5′-monophosphate. Biochemistry 2014, 53, 4938–4945. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.J.; Appleman, M.M. Multiple cyclic nucleotide phosphodiesterase activities from rat brain. Biochemistry 1971, 10, 311–316. [Google Scholar] [PubMed]
- Falcone, C.; Mazzoni, C.; Saliola, M. Alcohol dehydrogenase (ADH) isozymes in Kluyveromyces lactis: Detection by activity. In Springer Lab Manual Non-Conventional Yeasts in Genetics, Biochemistry and Biotechnology; Wolf, K., Breunig, K., Barth, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 195–201. [Google Scholar]
- Saliola, M.; Scappucci, G.; de Maria, I.; Lodi, T.; Mancini, P.; Falcone, C. Deletion of the glucose-6-phosphate dehydrogenase gene KlZWF1 affects both fermentative and respiratory metabolism in Kluyveromyces lactis. Eukaryot. Cell 2007, 6, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-Phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Zippin, J.H.; Chen, Y.; Nahirney, P.; Kamenetsky, M.; Wuttke, M.S.; Fischman, D.A.; Levin, L.R.; Buck, J. Compartmentalization of bicarbonate-sensitive adenylyl cyclase in distinct signaling microdomains. FASEB J. 2003, 17, 82–84. [Google Scholar] [CrossRef]
- Takimoto, E.; Koitabashi, N.; Hsu, S.; Ketner, E.A.; Zhang, M.; Nagayama, T.; Bedja, D.; Gabrielson, K.L.; Blanton, R.; Siderovski, D.P.; et al. Regulator of G protein signaling 2 mediates cardiac compensation to pressure overload and antihypertrophic effects of PDE5 inhibition in mice. J. Clin. Investig. 2009, 119, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevi, F.; Campolo, F.; Naro, F.; Zolla, L. The cardioprotective effect of sildenafil is mediated by the activation of malate dehydrogenase and an increase in the malate-aspartate shuttle in cardiomyocytes. Biochem. Pharmacol. 2017, 127, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Siregar, S.; Romdam, T.; Stanzah, H.; Firmansyah, I. The effect of long-acting PDE-5 inhibitor on renal ischemia/reperfusion injury in Wistar rats; a study on serum cystatin C and renal histopathological findings. J. Nephropathol. 2019, 8, e07. [Google Scholar] [CrossRef]
- Zhu, G.; Ueda, K.; Hashimoto, M.; Zhang, M.; Sasaki, M.; Kariya, T.; Sasaki, H.; Kaludercic, N.; Lee, D.I.; Bedja, D.; et al. The mitochondrial regulator PGC1α is induced by cGMP-PKG signaling and mediates the protective effects of phosphodiesterase 5 inhibition in heart failure. FEBS Lett. 2022, 596, 17–28. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MmPDE5A1 | MERAGPNSVRSQQQRDPDWVEAWLDDHRDFTFSYFIRATRDMVNAWFSE… |
| MmPDE5A2 | MLPFGDKTRDMVNAWFSE… |
| MmPDE5A3 | MVNAWFSE… |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardarelli, S.; Miele, A.E.; Campolo, F.; Massimi, M.; Mancini, P.; Biagioni, S.; Naro, F.; Giorgi, M.; Saliola, M. Cellular Redox Metabolism Is Modulated by the Distinct Localization of Cyclic Nucleotide Phosphodiesterase 5A Isoforms. Int. J. Mol. Sci. 2022, 23, 8587. https://doi.org/10.3390/ijms23158587
Cardarelli S, Miele AE, Campolo F, Massimi M, Mancini P, Biagioni S, Naro F, Giorgi M, Saliola M. Cellular Redox Metabolism Is Modulated by the Distinct Localization of Cyclic Nucleotide Phosphodiesterase 5A Isoforms. International Journal of Molecular Sciences. 2022; 23(15):8587. https://doi.org/10.3390/ijms23158587
Chicago/Turabian StyleCardarelli, Silvia, Adriana Erica Miele, Federica Campolo, Mara Massimi, Patrizia Mancini, Stefano Biagioni, Fabio Naro, Mauro Giorgi, and Michele Saliola. 2022. "Cellular Redox Metabolism Is Modulated by the Distinct Localization of Cyclic Nucleotide Phosphodiesterase 5A Isoforms" International Journal of Molecular Sciences 23, no. 15: 8587. https://doi.org/10.3390/ijms23158587