
MicroRNA-7a2 Contributes to Estrogen Synthesis and Is Modulated by FSH via the JNK Signaling Pathway in Ovarian Granulosa Cells

Abstract
:1. Introduction
2. Results
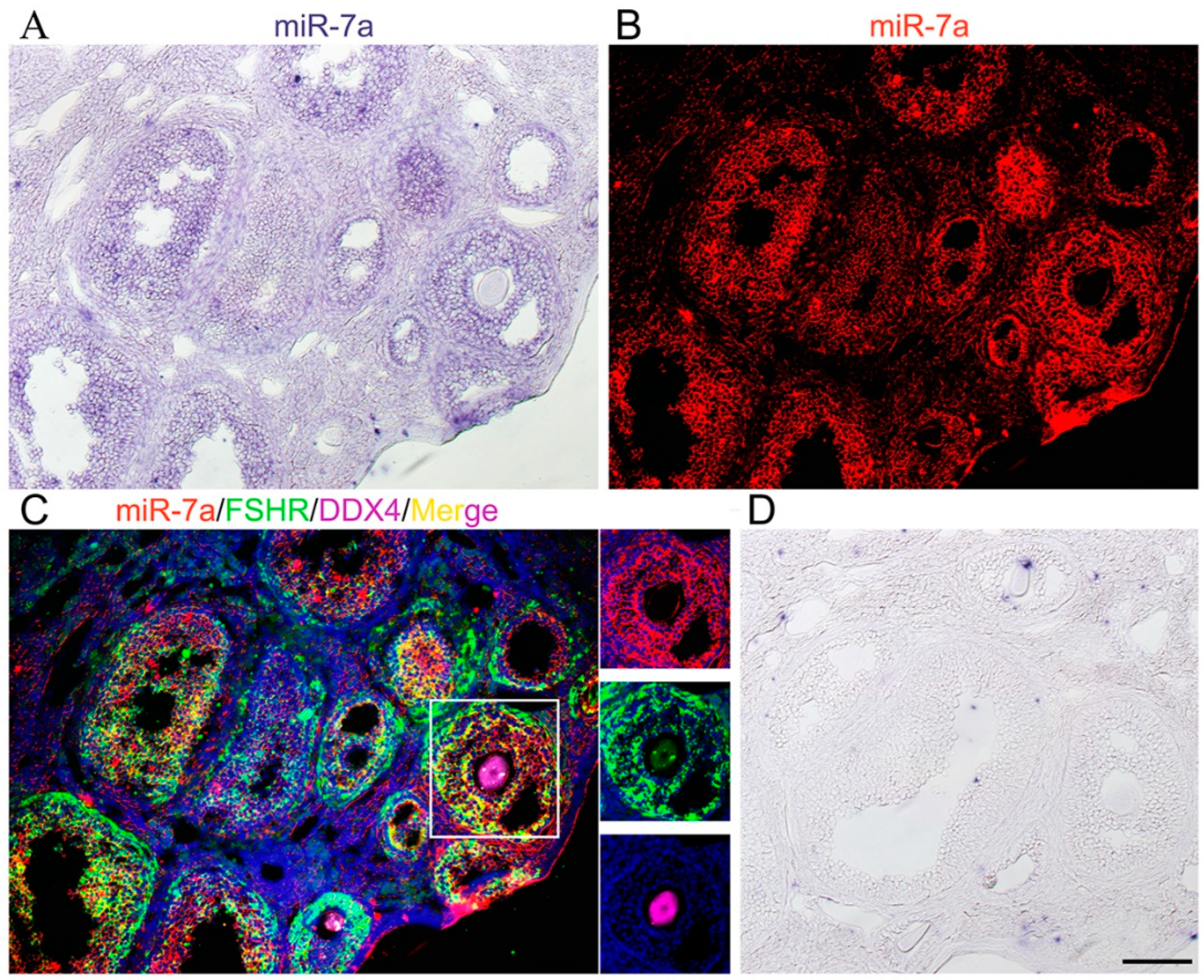
2.1. miR-7a Is Expressed in Mouse Ovaries
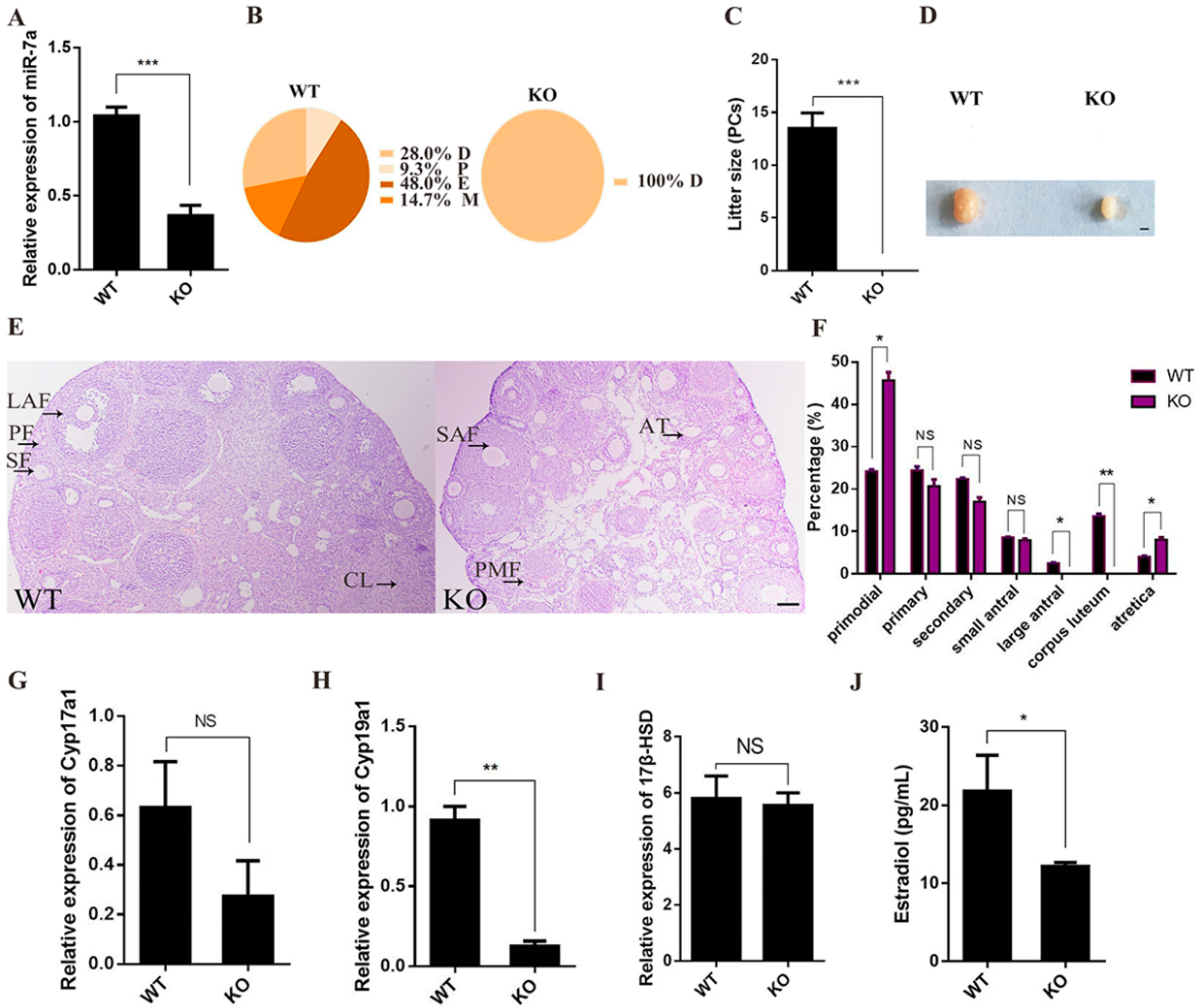
2.2. miR-7a2 KO Impairs Ovarian Function
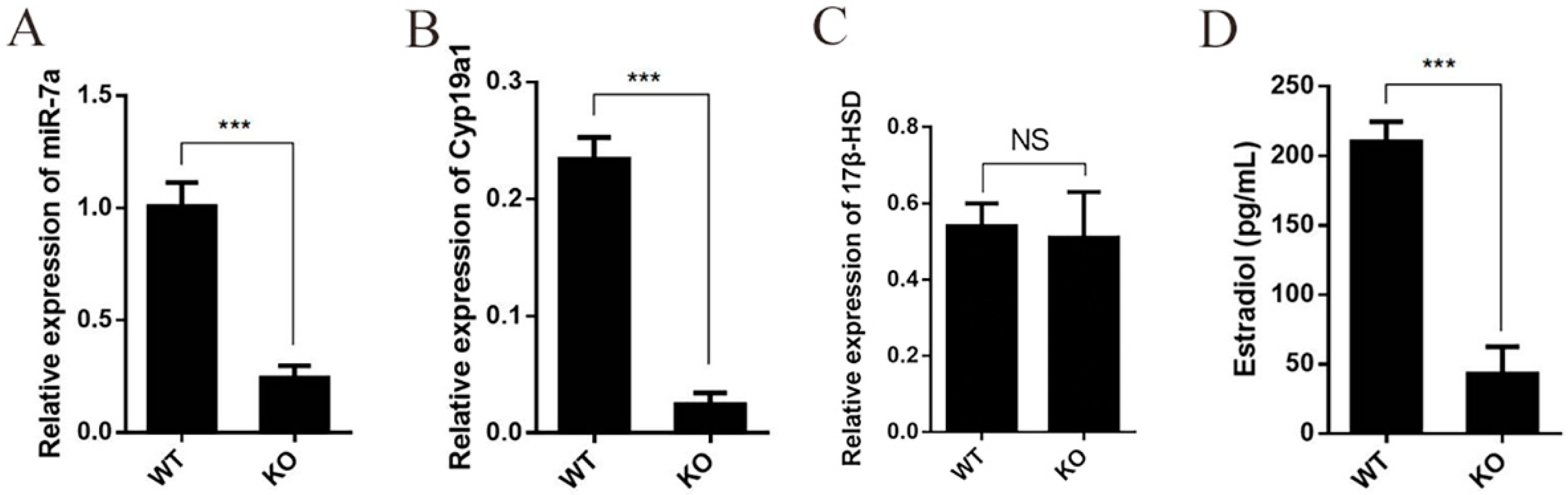
2.3. Deficiency of miR-7a2 Impairs the Function of Granulosa Cells in Estrogen Synthesis
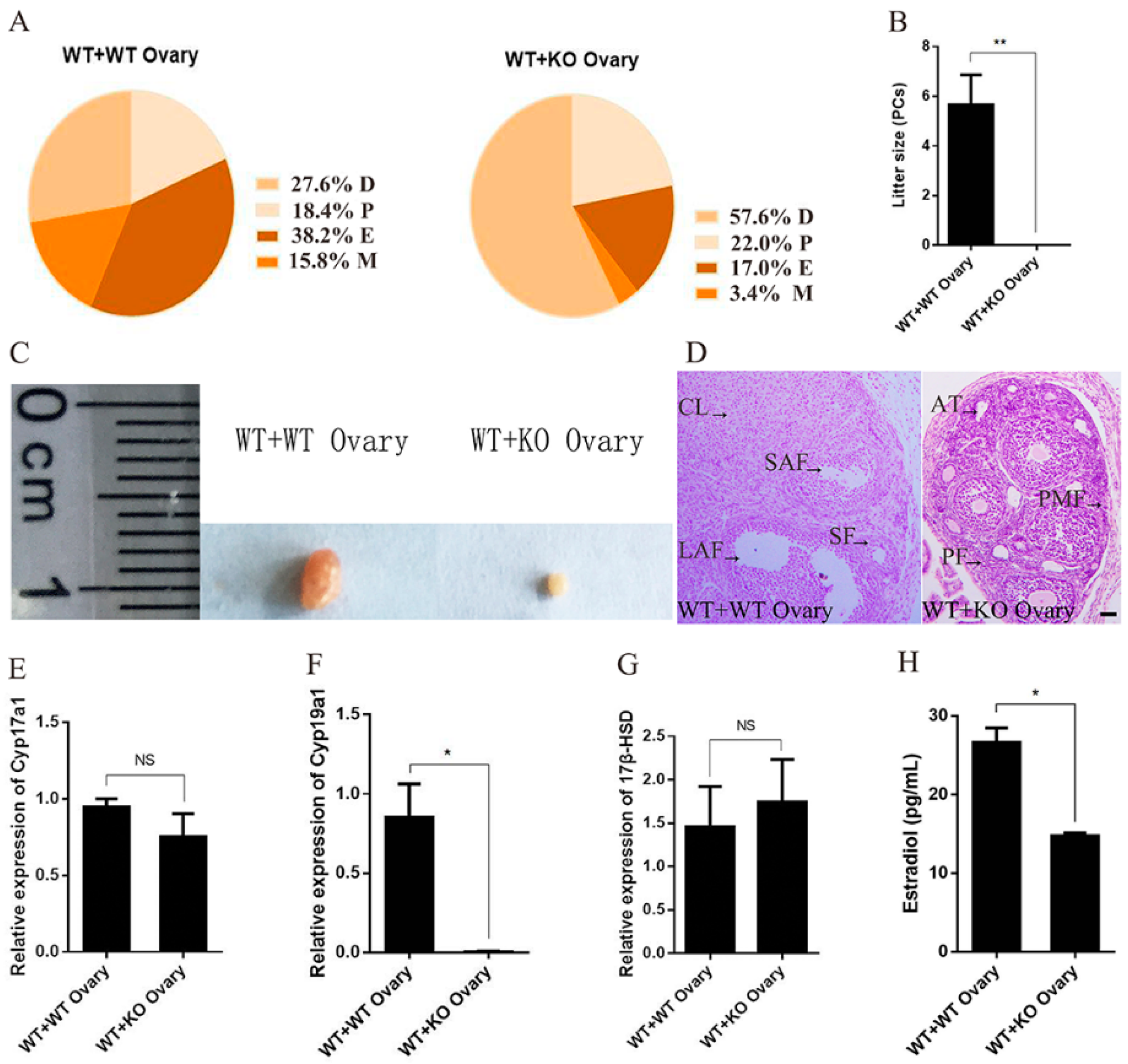
2.4. Specific Deficiency of Ovarian miR-7a2 Impairs the Ovarian Function
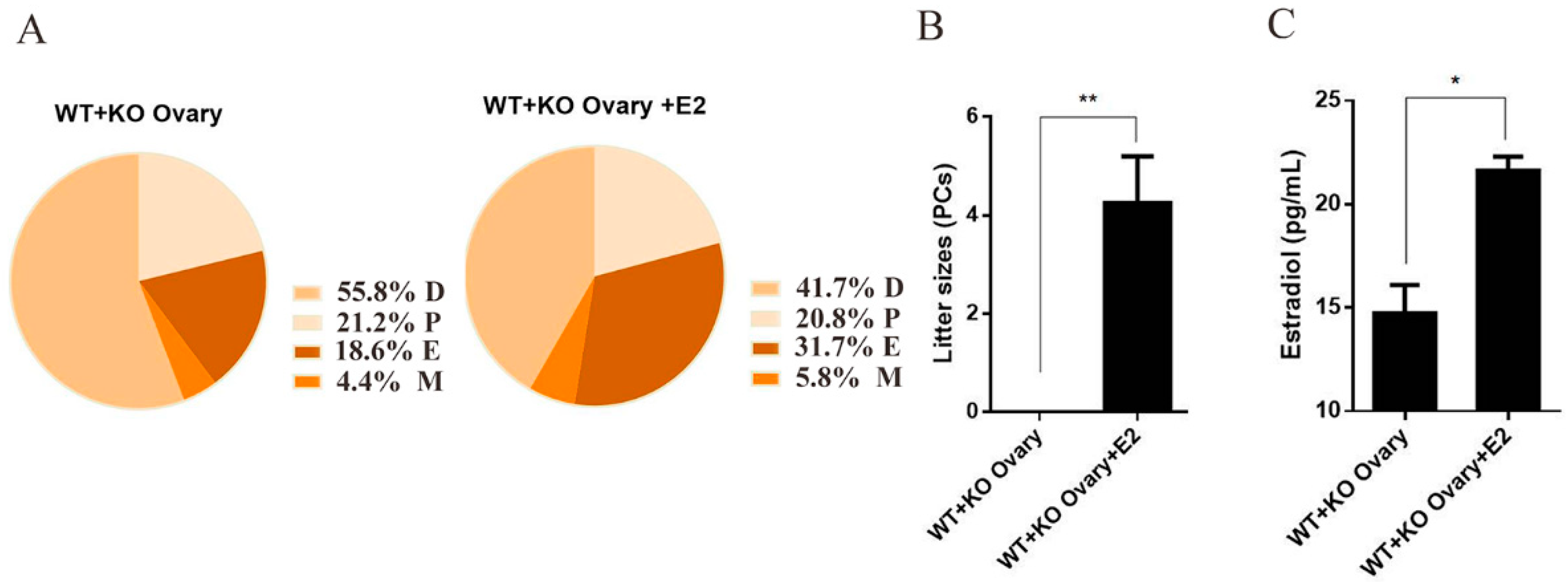
2.5. Estrogen Reverses the Infertility of Female Mice Caused by Ovarian miR-7a2 Deletion
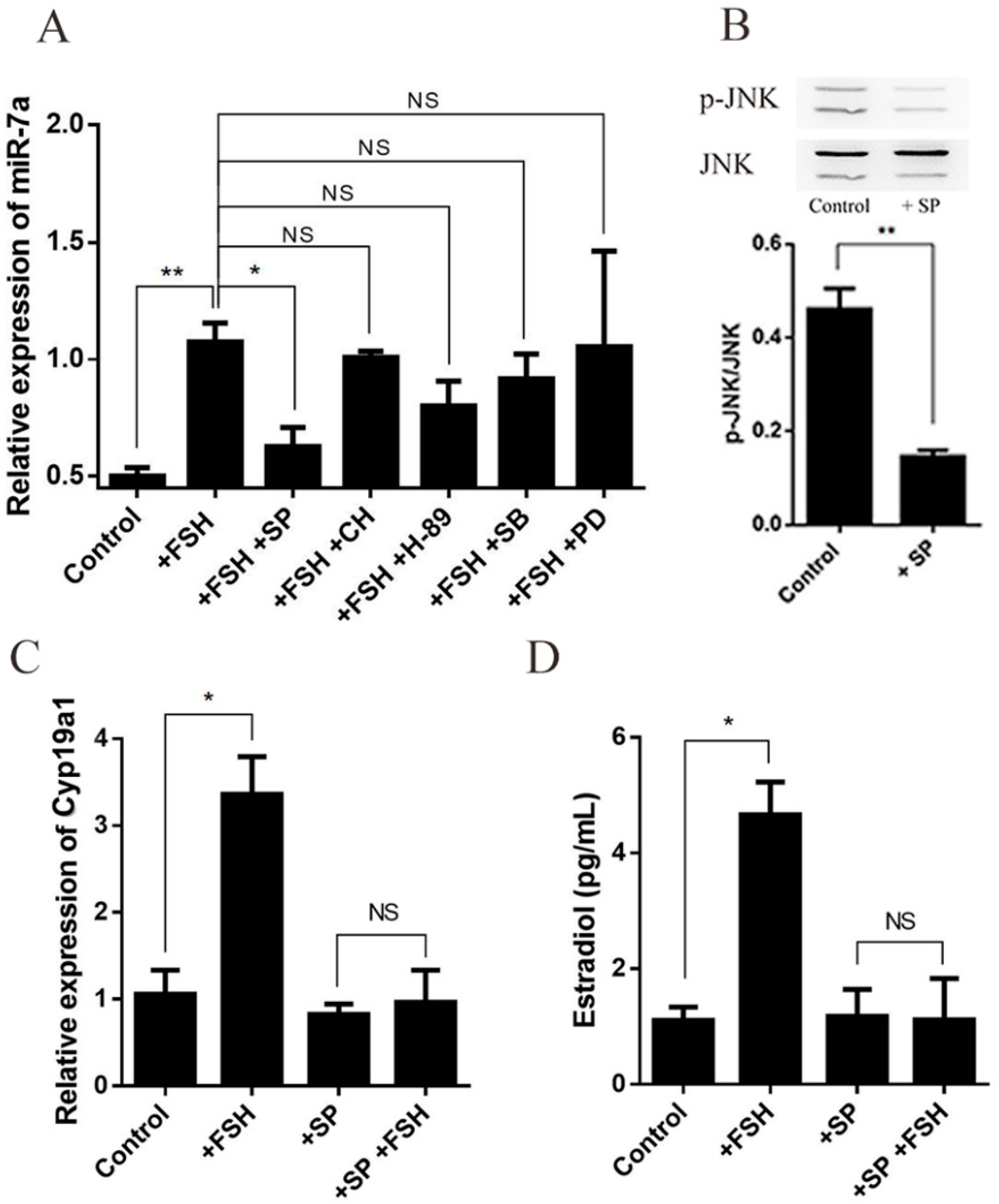
2.6. FSH Regulates Estrogen Synthesis by Modulating miR-7a in Granulosa Cells
2.7. FSH Enhances the Expression of miR-7a via JNK Signaling Pathway in Granulosa Cells
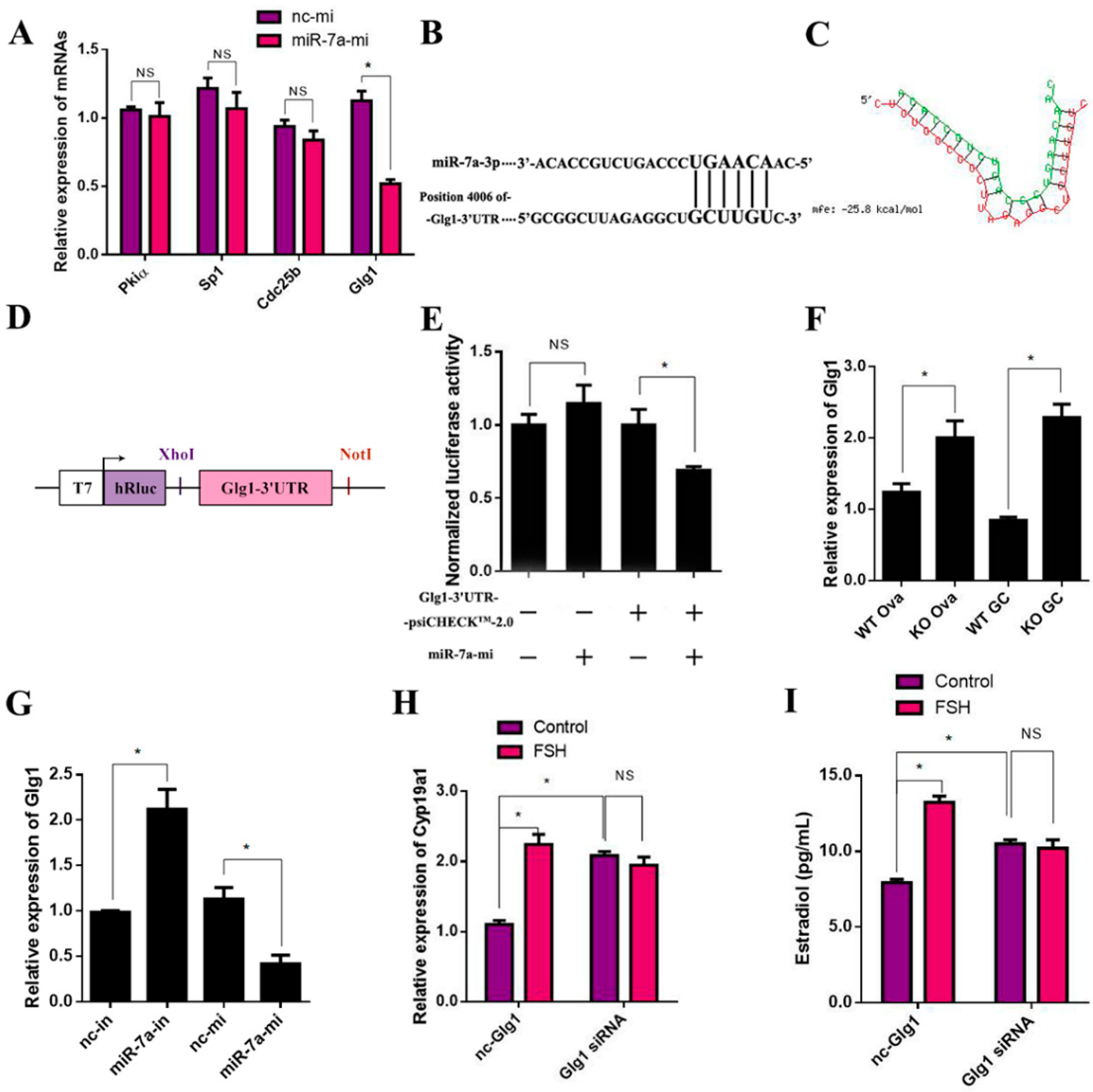
2.8. Glg1Is the Target Gene of miR-7a in Ovarian Granulosa Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. In Situhybridization (ISH) and Immunofluorescence Triple Staining
4.3. Vaginal Smear
4.4. Radioimmunoassay (RIA)
4.5. Ovarian Granulosa Cell Culture and Treatment
4.6. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.7. Western Blot
4.8. Dual-Luciferase Reporter Assay
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Plant, T.; Zeleznik, A. Knobil and Neill’s Physiology of Reproduction; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Christensen, A.; Bentley, G.E.; Cabrera, R.; Ortega, H.H.; Perfito, N.; Wu, T.J.; Micevych, P. Hormonal regulation of female reproduction. Horm. Metab. Res. 2012, 44, 587–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.Y. Effect of FSH and its different isoforms on maturation of oocytes from pre-ovulatory follicles. Reprod. Biomed. Online 2002, 5, 232–239. [Google Scholar] [CrossRef]
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 2018, 86, 3–17. [Google Scholar] [CrossRef]
- Stocco, C. Aromatase expression in the ovary: Hormonal and molecular regulation. Steroids 2008, 73, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Adashi, E.Y. Endocrinology of the ovary. Hum. Reprod. 1994, 9, 815–827. [Google Scholar] [CrossRef]
- Rojas, J.; Chavez-Castillo, M.; Olivar, L.C.; Calvo, M.; Mejias, J.; Rojas, M.; Morillo, J.; Bermudez, V. Physiologic Course of Female Reproductive Function: A Molecular Look into the Prologue of Life. J. Pregnancy 2015, 2015, 715735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britt, K.L.; Findlay, J.K. Estrogen actions in the ovary revisited. J. Endocrinol. 2002, 175, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Fisher, C.R.; Graves, K.H.; Parlow, A.F.; Simpson, E.R. Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc. Natl. Acad. Sci. USA 1998, 95, 6965–6970. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Dermatol. 2001, 45, S116–S124. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, J.; Zhang, C. Synthesis, Regulatory Factors, and Signaling Pathways of Estrogen in the Ovary. Reprod. Sci. 2022. [Google Scholar] [CrossRef]
- Xiong, S.; Tian, J.; Ge, S.; Li, Z.; Long, Z.; Guo, W.; Huang, P.; He, Y.; Xiao, T.; Gui, J.F.; et al. The microRNA-200 cluster on chromosome 23 is required for oocyte maturation and ovulation in zebrafishdagger. Biol. Reprod. 2020, 103, 769–778. [Google Scholar] [CrossRef]
- McGinnis, L.K.; Luense, L.J.; Christenson, L.K. MicroRNA in Ovarian Biology and Disease. Cold Spring Harb. Perspect. Med. 2015, 5, a22962. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Linher-Melville, K.; Yang, B.B.; Wu, D.; Li, J. Micro-RNA378 (miR-378) regulates ovarian estradiol production by targeting aromatase. Endocrinology 2011, 152, 3941–3951. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Du, X.; Liu, L.; Liu, H.; Pan, Z.; Li, Q. Upregulation of miR-146b promotes porcine ovarian granulosa cell apoptosis by attenuating CYP19A1. Domest. Anim. Endocrinol. 2021, 74, 106509. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xu, S.; Ji, Z.; Sun, Y.; Cai, B.; Zhang, S.; Wang, P. The role of miR-7 as a potential switch in the mouse hypothalamus-pituitary-ovary axis through regulation of gonadotropins. Mol. Cell. Endocrinol. 2020, 518, 110969. [Google Scholar] [CrossRef]
- Horsham, J.L.; Ganda, C.; Kalinowski, F.C.; Brown, R.A.; Epis, M.R.; Leedman, P.J. MicroRNA-7: A miRNA with expanding roles in development and disease. Int. J. Biochem. Cell Biol. 2015, 69, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E.; Plasterk, R.H. MicroRNA function in animal development. FEBS Lett. 2005, 579, 5911–5922. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.; LaPierre, M.P.; Gasser, E.; Denzler, R.; Yang, Y.; Rulicke, T.; Kero, J.; Latreille, M.; Stoffel, M. Loss of microRNA-7a2 induces hypogonadotropic hypogonadism and infertility. J. Clin. Investig. 2017, 127, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, Y.; Guo, J.; Li, L.; Liu, H.; Lu, C.; Jiang, Y.; Cui, S. MicroRNA-7a2 is required for the development of pituitary stem cells. Stem Cells Dev. 2022, 31, 357–368. [Google Scholar] [CrossRef]
- LaPierre, M.P.; Godbersen, S.; Torres, E.M.; Schad, A.N.; Treier, M.; Ghoshdastider, U.; Stoffel, M. MicroRNA-7a2 Regulates Prolactin in Developing Lactotrophs and Prolactinoma Cells. Endocrinology 2021, 162, bqaa220. [Google Scholar] [CrossRef]
- Latreille, M.; Hausser, J.; Stutzer, I.; Zhang, Q.; Hastoy, B.; Gargani, S.; Kerr-Conte, J.; Pattou, F.; Zavolan, M.; Esguerra, J.L.; et al. MicroRNA-7a regulates pancreatic beta cell function. J. Clin. Investig. 2014, 124, 2722–2735. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, H.; Zhang, D.; Xu, G.; Zhang, J.; Cui, S. miR-7 mediates the signaling pathway of NE affecting FSH and LH synthesis in pig pituitary. J. Endocrinol. 2020, 244, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhang, J.; Zhou, Y.; Li, X.; Li, H.; Liu, J.; Gou, K.; Zhao, J.; Cui, S. MicroRNA-7 inhibits melatonin synthesis by acting as a linking molecule between leptin and norepinephrine signaling pathways in pig pineal gland. J. Pineal Res. 2019, 66, e12552. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Qin, N.; Xu, X.; Sun, X.; Chen, X.; Zhao, J.; Xu, R.; Mishra, B. Synergistic inhibition of csal1 and csal3 in granulosa cell proliferation and steroidogenesis of hen ovarian prehierarchical development. Biol. Reprod. 2019, 101, 986–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zhao, F.; Lin, P.; Zhang, G.; Tang, K.; Wang, A.; Jin, Y. Knockdown of XBP1 by RNAi in Mouse Granulosa Cells Promotes Apoptosis, Inhibits Cell Cycle, and Decreases Estradiol Synthesis. Int. J. Mol. Sci. 2017, 18, 1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robker, R.L.; Richards, J.S. Hormone-induced proliferation and differentiation of granulosa cells: A coordinated balance of the cell cycle regulators cyclin D2 and p27Kip1. Mol. Endocrinol. 1998, 12, 924–940. [Google Scholar] [CrossRef]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen Hormone Biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xin, Q.; Wang, X.; Wang, S.; Wang, H.; Zhang, W.; Yang, Y.; Zhang, Y.; Zhang, Z.; Wang, C.; et al. Estrogen receptors in granulosa cells govern meiotic resumption of pre-ovulatory oocytes in mammals. Cell Death Dis. 2017, 8, e2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Srivastava, D. A developmental view of microRNA function. Trends Biochem. Sci. 2007, 32, 189–197. [Google Scholar] [CrossRef]
- Wu, P.H.; Isaji, M.; Carthew, R.W. Functionally diverse microRNA effector complexes are regulated by extracellular signaling. Mol. Cell 2013, 52, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, S.Y.; Ding, M.; Dou, X.; Zhang, F.F.; Wu, Z.Y.; Qian, S.W.; Zhang, W.; Tang, Q.Q.; Xu, C.J. The BMP4-Smad signaling pathway regulates hyperandrogenism development in a female mouse model. J. Biol. Chem. 2017, 292, 11740–11750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, K.A.; Handelsman, D.J. Role of androgens in the ovary. Mol. Cell. Endocrinol. 2018, 465, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Hunzicker-Dunn, M.; Maizels, E.T. FSH signaling pathways in immature granulosa cells that regulate target gene expression: Branching out from protein kinase A. Cell. Signal. 2006, 18, 1351–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatzirodos, N.; Irving-Rodgers, H.F.; Hummitzsch, K.; Harland, M.L.; Morris, S.E.; Rodgers, R.J. Transcriptome profiling of granulosa cells of bovine ovarian follicles during growth from small to large antral sizes. BMC Genom. 2014, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Behringer, R. Mouse Ovary Transplantation. Cold Spring Harb. Protoc. 2017, 2017, pdb.prot094458. [Google Scholar] [CrossRef]
- Jones, E.C.; Krohn, P.L. Orthotopic ovarian transplantation in mice. J. Endocrinol. 1960, 20, 135–146. [Google Scholar] [CrossRef]
- Cora, M.C.; Kooistra, L.; Travlos, G. Vaginal Cytology of the Laboratory Rat and Mouse: Review and Criteria for the Staging of the Estrous Cycle Using Stained Vaginal Smears. Toxicol. Pathol. 2015, 43, 776–793. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wen, H.; Wang, C.; Li, Q. SMAD7 antagonizes key TGFbeta superfamily signaling in mouse granulosa cells in vitro. Reproduction 2013, 146, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yao, W.; Li, C.; Wu, W.; Li, Q.; Liu, H. Administration of follicle-stimulating hormone induces autophagy via upregulation of HIF-1alpha in mouse granulosa cells. Cell Death Dis. 2017, 8, e3001. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Wen, X.; Lin, P.; Chen, H.; Wang, A.; Jin, Y. Activation of CREBZF Increases Cell Apoptosis in Mouse Ovarian Granulosa Cells by Regulating the ERK1/2 and mTOR Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 3517. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Sequence |
|---|---|
| U6 RT Primer | AACGCTTCACGAATTTGCGT |
| U6 Forward Primer | CTCGCTTCGGCAGCACA |
| U6 Reverse Primer | AACGCTTCACGAATTTGCGT |
| miR-7a RT Primer | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACAACAAAAT |
| miR-7a Forward Primer | GATGAGCTGTCCACCTGCTT |
| miR-7a Reverse Primer | CTGTCCCCTGTCCCACTCTA |
| GAPDH Forward Primer | GGTTGTCTCCTGCGACTTCA |
| GAPDH Reverse Primer | GGGTGGTCCAGGGTTTCTTA |
| Cyp19a1 Forward Primer | TTGGAAATGCTGAACCCCAT |
| Cyp19a1 Reverse Primer | CAAGAATCTGCCATGGGAAA |
| Glg1 Forward Primer | GGGCTGTACCTGACCTCT |
| Glg1 Reverse Primer | CCTTGTCACCACCTGTCT |
| 17βHSD Forward Primer | GACCGTTCCCAGAGCTTCAA |
| 17βHSD Reverse Primer | CAGCACCCACAGCGTTCAAT |
| Cyp17a1 Forward Primer | GATCTAAGAAGCGCTCAGGCA |
| Cyp17a1 Reverse Primer | GGGCACTGCATCACGATAAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhang, J.; Lu, C.; Wang, B.; Guo, J.; Zhang, H.; Cui, S. MicroRNA-7a2 Contributes to Estrogen Synthesis and Is Modulated by FSH via the JNK Signaling Pathway in Ovarian Granulosa Cells. Int. J. Mol. Sci. 2022, 23, 8565. https://doi.org/10.3390/ijms23158565
Li L, Zhang J, Lu C, Wang B, Guo J, Zhang H, Cui S. MicroRNA-7a2 Contributes to Estrogen Synthesis and Is Modulated by FSH via the JNK Signaling Pathway in Ovarian Granulosa Cells. International Journal of Molecular Sciences. 2022; 23(15):8565. https://doi.org/10.3390/ijms23158565
Chicago/Turabian StyleLi, Liuhui, Jinglin Zhang, Chenyang Lu, Bingjie Wang, Jiajia Guo, Haitong Zhang, and Sheng Cui. 2022. "MicroRNA-7a2 Contributes to Estrogen Synthesis and Is Modulated by FSH via the JNK Signaling Pathway in Ovarian Granulosa Cells" International Journal of Molecular Sciences 23, no. 15: 8565. https://doi.org/10.3390/ijms23158565