
Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media

 ,
,  , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
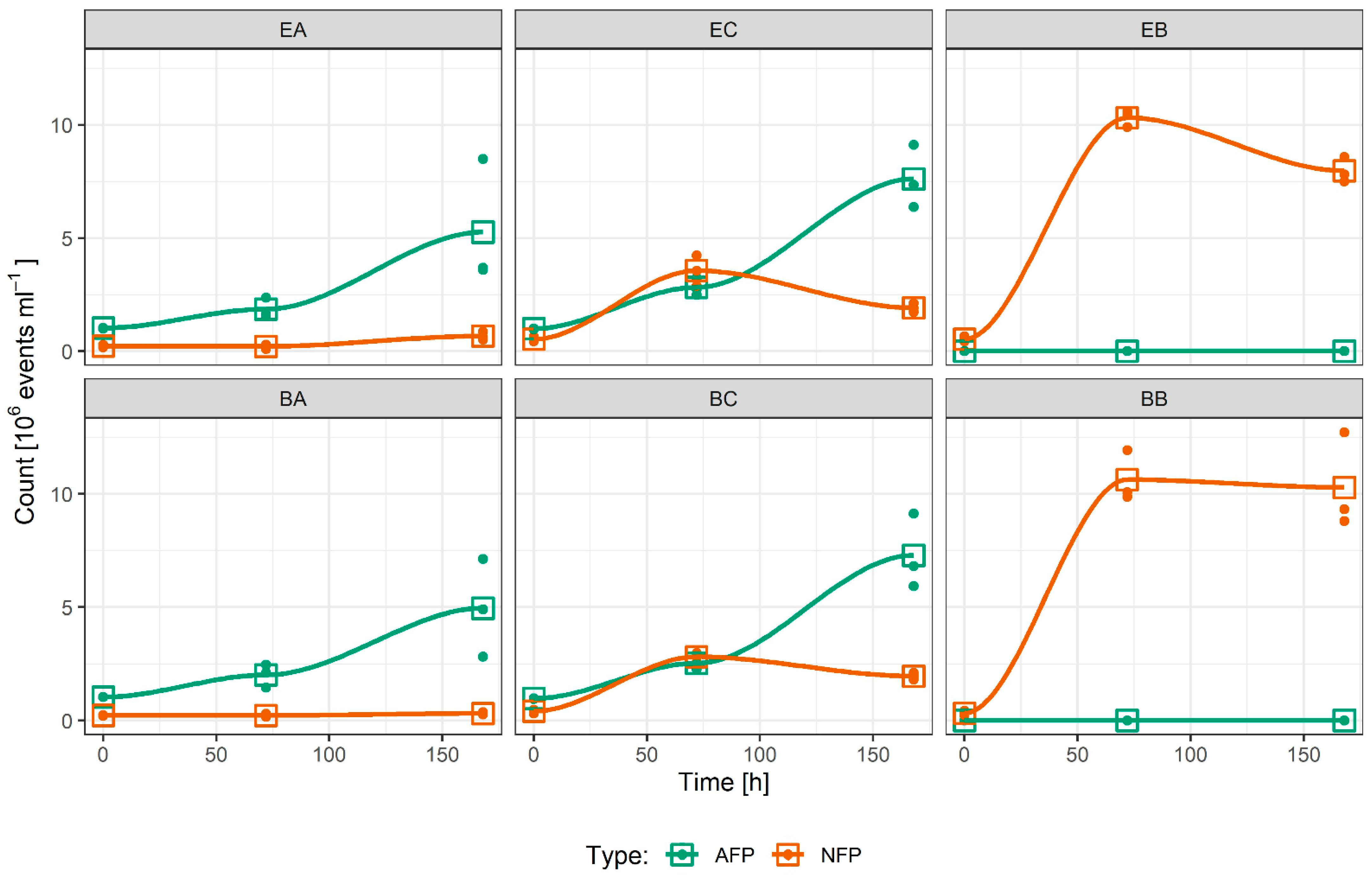
2.1. Culture Growth
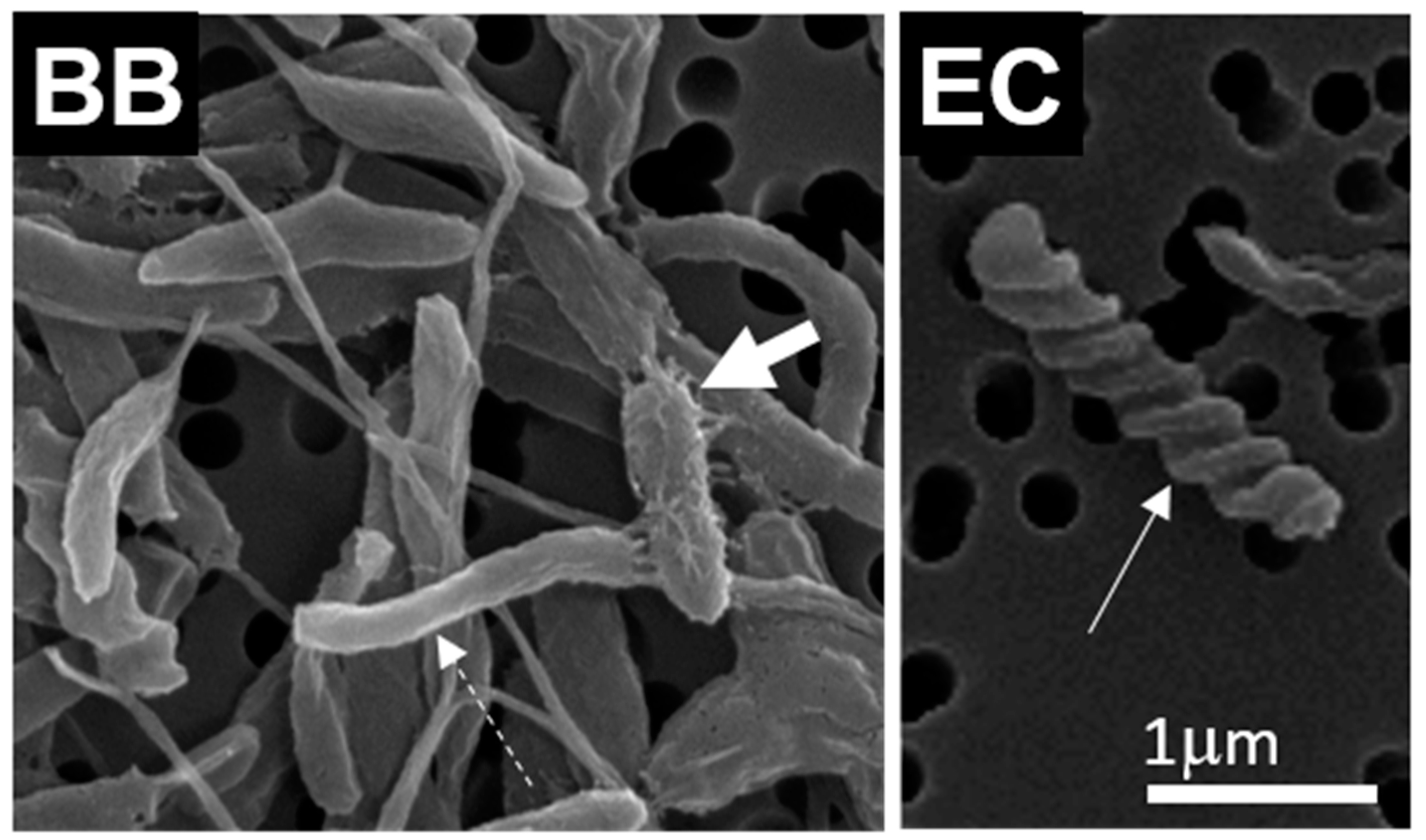
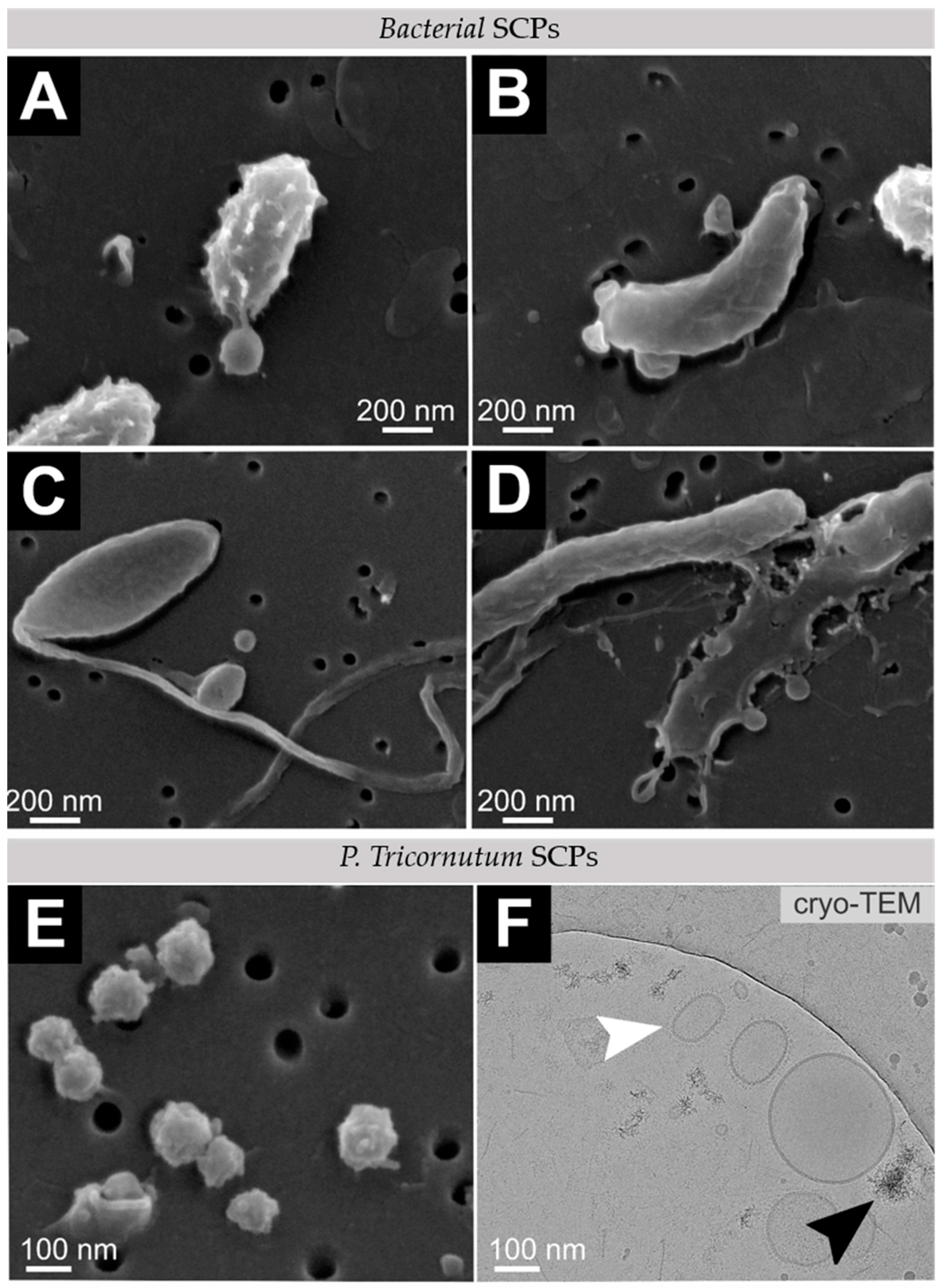
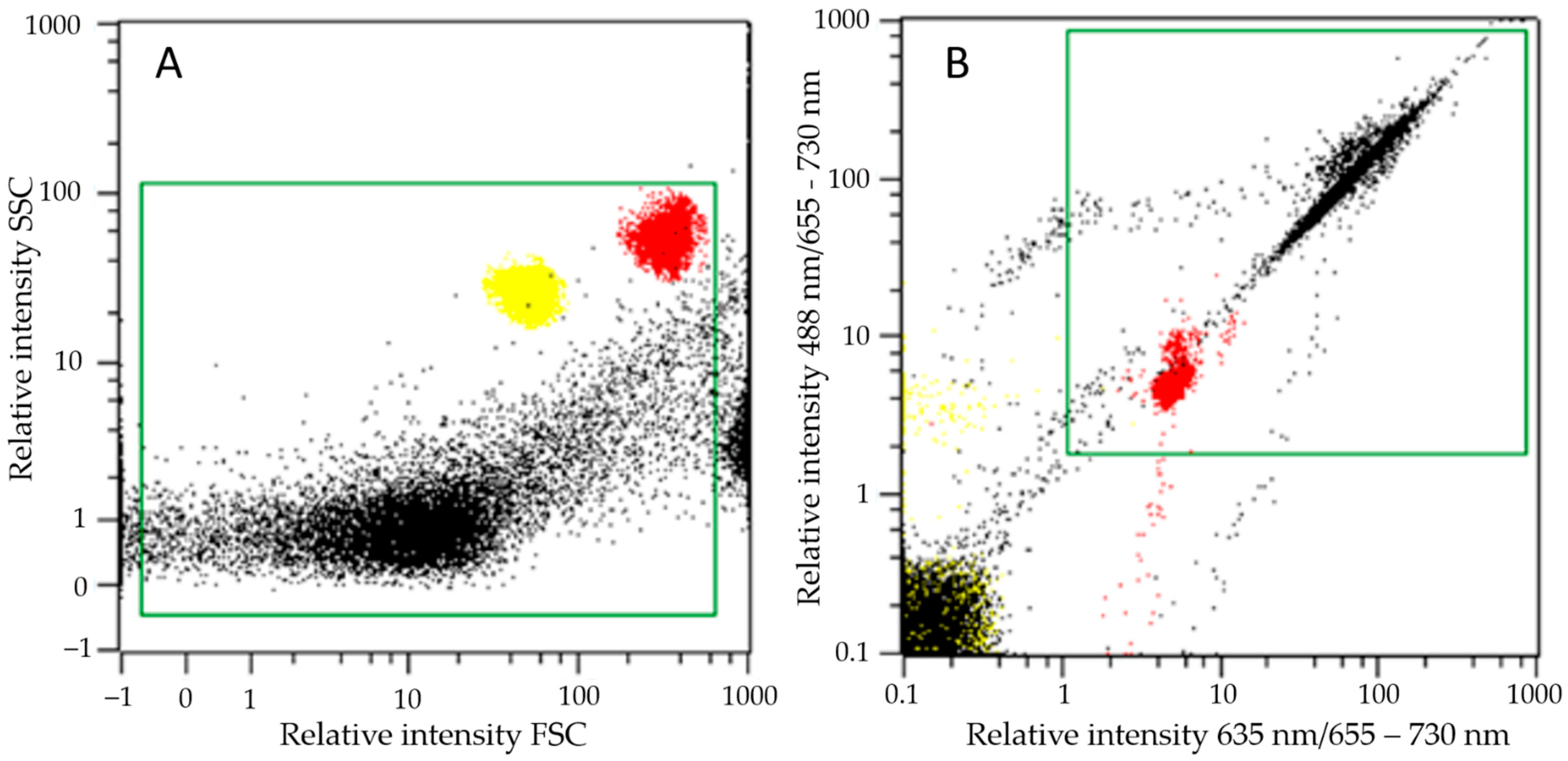
2.2. Cell Morphology and SCP Characterization
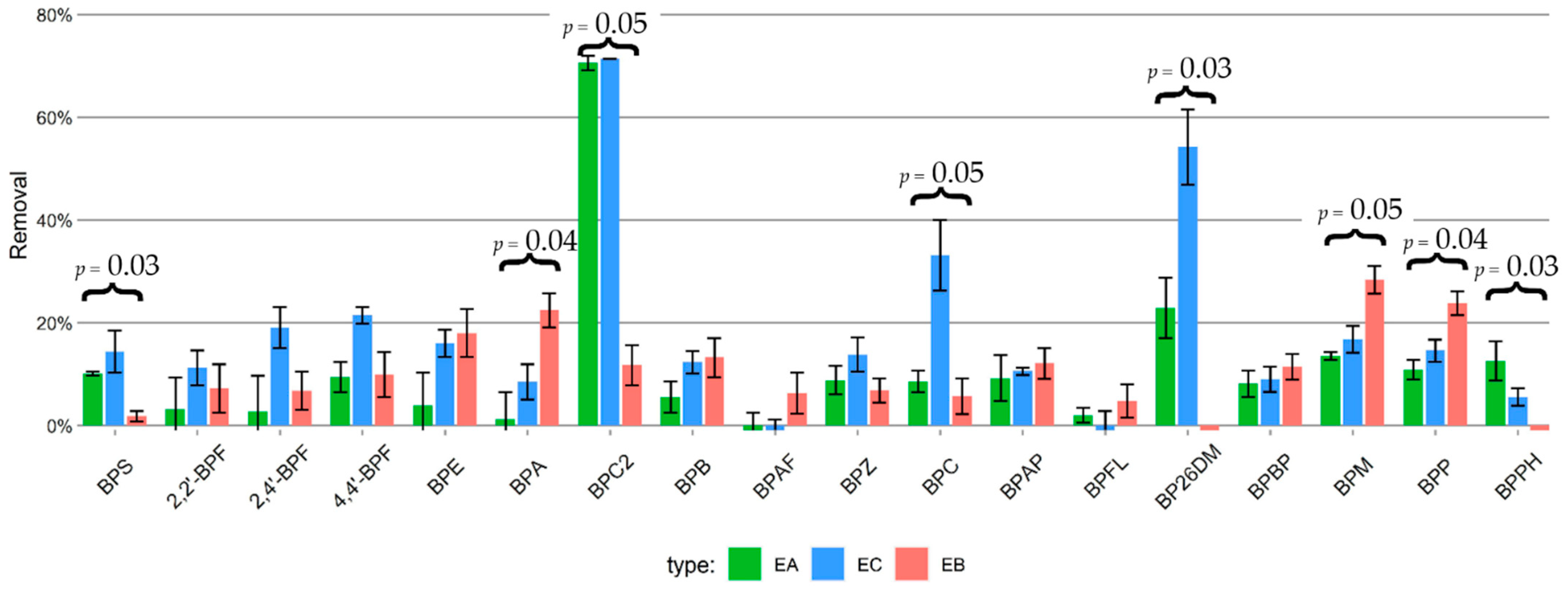
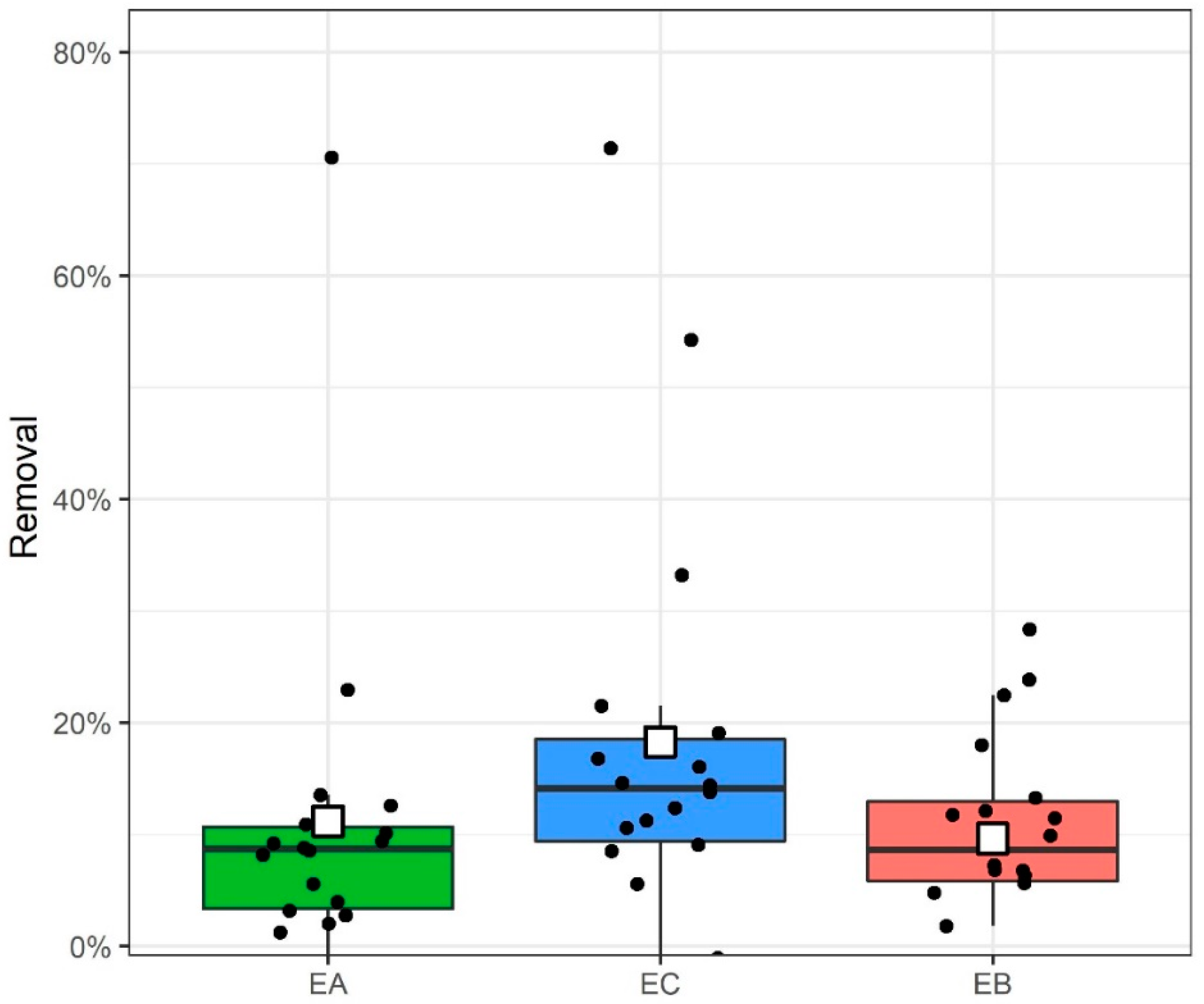
2.3. Bisphenol Residue Mass Balance and Removal
3. Discussion
3.1. Culture Growth
3.2. Cell and SCP Morphology
3.3. Bisphenol Residue Mass Balance and Removal
4. Materials and Methods
4.1. Cultures and Medium Composition
4.2. Cell Concentration Measurement by Flow Cytometry
4.3. SCP Isolation
4.4. SEM Imaging
4.5. Cryo-TEM Imaging
4.6. Bisphenol Quantification
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Langergraber, G.; Castellar, J.A.C.; Andersen, T.R.; Andreucci, M.-B.; Baganz, G.F.M.; Buttiglieri, G.; Canet-Martí, A.; Carvalho, P.N.; Finger, D.C.; Griessler Bulc, T.; et al. Towards a cross-sectoral view of nature-based solutions for enabling circular cities. Water 2021, 13, 2352. [Google Scholar] [CrossRef]
- Norvill, Z.N.; Shilton, A.; Guieysse, B. Emerging contaminant degradation and removal in algal wastewater treatment ponds: Identifying the research gaps. J. Hazard. Mater. 2016, 313, 291–309. [Google Scholar] [CrossRef]
- Craggs, R.; Sutherland, D.; Campbell, H. Hectare-scale demonstration of high rate algal ponds for enhanced wastewater treatment and biofuel production. J. Appl. Phycol. 2012, 24, 329–337. [Google Scholar] [CrossRef]
- Sutherland, D.L.; Park, J.; Heubeck, S.; Ralph, P.J.; Craggs, R.J. Size matters—Microalgae production and nutrient removal in wastewater treatment high rate algal ponds of three different sizes. Algal. Res. 2020, 45, 101734. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Park, J.B.K.; Craggs, R.J.; Shilton, A.N. Wastewater treatment high rate algal ponds for biofuel production. Bioresour. Technol. 2011, 102, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Aziz, M.A.; Al-khulaidi, R.A.; Sakib, N.; Islam, M. Biodiesel production from microalgae Spirulina maxima by two step process: Optimization of process variable. J. Radiat. Res. Appl. Sci. 2017, 10, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Putman, R.J.; Inan, K.; Sal, F.A.; Sathish, A.; Smith, T.; Nielsen, C.; Sims, R.C.; Miller, C.D. Polyhydroxybutyrate production using a wastewater microalgae based media. Algal Res. 2015, 8, 95–98. [Google Scholar] [CrossRef]
- Armingol, E.; Officer, A.; Harismendy, O.; Lewis, N.E. Deciphering cell–cell interactions and communication from gene expression. Nat. Rev. Genet. 2021, 22, 71–88. [Google Scholar] [CrossRef]
- Combarnous, Y.; Nguyen, T.M.D. Cell Communications among microorganisms, plants, and animals: Origin, evolution, and interplays. Int. J. Mol. Sci. 2020, 21, 8052. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.-H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2019, 43, 273–303. [Google Scholar] [CrossRef]
- Aaronson, S. The synthesis of extracellular macromolecules and membranes by a population of the phytoflagellate Ochromonas danica. Limnol. Oceanogr. 1971, 16, 1–9. [Google Scholar] [CrossRef]
- McLean, R.J.; Laurendi, C.J.; Brown, R.M. The Relationship of Gamone to the Mating Reaction in Chlamydomonas moewusii. Proc. Natl. Acad. Sci. USA 1974, 71, 2610–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozic, D.; Hočevar, M.; Jeran, M.; Kisovec, M.; Bedina Zavec, A.; Romolo, A.; Škufca, D.; Podobnik, M.; Kogej, K.; Iglič, A.; et al. Ultrastructure and stability of cellular nanoparticles isolated from Phaeodactylum tricornutum and Dunaliella tertiolecta conditioned media. Open Res. Eur. 2022. submit. [Google Scholar]
- Picciotto, S.; Barone, M.E.; Fierli, D.; Aranyos, A.; Adamo, G.; Božič, D.; Romancino, D.P.; Stanly, C.; Parkes, R.; Morsbach, S.; et al. Isolation of extracellular vesicles from microalgae: Towards the production of sustainable and natural nanocarriers of bioactive compounds. Biomater. Sci. 2021, 9, 2917–2930. [Google Scholar] [CrossRef]
- Adamo, G.; Fierli, D.; Romancino, D.P.; Picciotto, S.; Barone, M.E.; Aranyos, A.; Božič, D.; Morsbach, S.; Raccosta, S.; Stanly, C.; et al. Nanoalgosomes: Introducing extracellular vesicles produced by microalgae. J. Extracell. Vesicles 2021, 10, e12081. [Google Scholar] [CrossRef] [PubMed]
- Noguera-Oviedo, K.; Aga, D.S. Lessons learned from more than two decades of research on emerging contaminants in the environment. J. Hazard. Mater. 2016, 316, 242–251. [Google Scholar] [CrossRef]
- Tolboom, S.N.; Carrillo-Nieves, D.; de Jesús Rostro-Alanis, M.; de la Cruz Quiroz, R.; Barceló, D.; Iqbal, H.M.N.; Parra-Saldivar, R. Algal-based removal strategies for hazardous contaminants from the environment—A review. Sci. Total Environ. 2019, 665, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Rangabhashiyam, S.; Verma, P.; Singh, P.; Devi, P.; Kumar, P.; Hussain, C.M.; Gaurav, G.K.; Kumar, K.S. Environmental and health impacts of contaminants of emerging concerns: Recent treatment challenges and approaches. Chemosphere 2021, 272, 129492. [Google Scholar] [CrossRef] [PubMed]
- Geens, T.; Goeyens, L.; Covaci, A. Are potential sources for human exposure to bisphenol-A overlooked? Int. J. Hyg. Environ. Health 2011, 214, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Vehar, A.; Kovačič, A.; Hvala, N.; Škufca, D.; Levstek, M.; Stražar, M.; Žgajnar Gotvajn, A.; Heath, E. Fate of bisphenols during conventional wastewater treatment. Proc. Socrat. Lectures. 2021, 6, 57–62. [Google Scholar] [CrossRef]
- Chen, D.; Kannan, K.; Tan, H.; Zheng, Z.; Feng, Y.-L.; Wu, Y.; Widelka, M. Bisphenol analogues other than BPA: Environmental occurrence, human exposure, and toxicity—A review. Environ. Sci. Technol. 2016, 50, 5438–5453. [Google Scholar] [CrossRef]
- Muñoz, I.; López-Doval, J.C.; Ricart, M.; Villagrasa, M.; Brix, R.; Geiszinger, A.; Ginebreda, A.; Guasch, H.; de Alda, M.J.L.; Romaní, A.M.; et al. Bridging levels of pharmaceuticals in river water with biological community structure in the Llobregat river basin (northeast Spain). Environ. Toxicol. Chem. 2009, 28, 2706. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, D.L.; Ralph, P.J. Microalgal bioremediation of emerging contaminants—Opportunities and challenges. Water Res. 2019, 164, 114921. [Google Scholar] [CrossRef]
- Škufca, D.; Prosenc, F.; Kovačič, A.; Buttiglieri, G.; Heath, D.; Griessler Bulc, T.; Heath, E. Removal of contaminants of emerging concern in algal photobioreactors: From lab-scale to pilot-scale. Proc. Socrat. Lect. 2021, 6, 207. [Google Scholar]
- Maryjoseph, S.; Ketheesan, B. Microalgae based wastewater treatment for the removal of emerging contaminants: A review of challenges and opportunities. Case Stud. Chem. Environ. Eng. 2020, 2, 100046. [Google Scholar] [CrossRef]
- Liu, R.; Li, S.; Tu, Y.; Hao, X. Capabilities and mechanisms of microalgae on removing micropollutants from wastewater: A review. J. Environ. Man. 2021, 285, 112149. [Google Scholar] [CrossRef]
- Božič, D.; Hočevar, M.; Jeran, M.; Matos, T.; Tomazin, R.; Pocsfalvi, G.; Iglič, A.; Kralj-Iglič, V. Scanning electron microscopy of microorganisms growing in co-cultures with microalgae. Proc. Socrat. Lect. 2021, 6, 206. [Google Scholar]
- Ji, X.; Jiang, M.; Zhang, J.; Jiang, X.; Zheng, Z. Bioresource Technology The interactions of algae-bacteria symbiotic system and its effects on nutrients removal from synthetic wastewater. Bioresour. Technol. 2018, 247, 44–50. [Google Scholar] [CrossRef]
- Kumari, M.; Ghosh, P.; Thakur, I.S. Landfill leachate treatment using bacto-algal co-culture: An integrated approach using chemical analyses and toxicological assessment. Ecotoxicol. Environ. Saf. 2016, 128, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Škufca, D.; Prosenc, F.; Bulc, T.G.; Heath, E. Removal and fate of 18 bisphenols in lab-scale algal bioreactors. Sci. Total Environ. 2021, 804, 149878. [Google Scholar] [CrossRef]
- Watabe, Y.; Kondo, T.; Imai, H.; Morita, M.; Tanaka, N.; Hosoya, K. Reducing Bisphenol A Contamination from analytical procedures to determine ultralow levels in environmental samples using automated HPLC microanalysis. Anal. Chem. 2004, 76, 105–109. [Google Scholar] [CrossRef]
- Prosenc, F.; Piechocka, J.; Škufca, D.; Heath, E.; Griessler Bulc, T.; Istenič, D.; Buttiglieri, G. Microalgae-based removal of contaminants of emerging concern: Mechanisms in Chlorella vulgaris and mixed algal-bacterial cultures. J. Hazard. Mater. 2021, 418, 126284. [Google Scholar] [CrossRef]
- Emam, S.E.; Ando, H.; Abu Lila, A.S.; Shimizu, T.; Ukawa, M.; Okuhira, K.; Ishima, Y.; Mahdy, M.A.; Ghazy, F.S.; Ishida, T. A novel strategy to increase the yield of exosomes (extracellular vesicles) for an expansion of basic research. Biol. Pharm. Bull. 2018, 41, 733. [Google Scholar] [CrossRef] [Green Version]
- Paganini, C.; Capasso Palmiero, U.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable production and isolation of extracellular vesicles: Available sources and lessons from current industrial bioprocesses. Biotechnol. J. 2019, 14, 1800528. [Google Scholar] [CrossRef] [PubMed]
- Maadane, A.; Merghoub, N.; El Mernissi, N.; Ainane, T.; Amzazi, S. Antimicrobial activity of marine microalgae isolated from Moroccan coastlines. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1257–1260. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Said, I.H.; Thorstenson, C.; Thomsen, C.; Ullrich, M.S.; Kuhnert, N.; Thomsen, L. Pilot-scale production of antibacterial substances by the marine diatom Phaeodactylum tricornutum Bohlin. Algal Res. 2018, 32, 113–120. [Google Scholar] [CrossRef]
- Desbois, A.P.; Walton, M.; Smith, V.J. Differential antibacterial activities of fusiform and oval morphotypes of Phaeodactylum tricornutum (Bacillariophyceae). J. Mar. Biol. Assoc. UK 2010, 90, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-H.; Ramanan, R.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Role of Rhizobium, a plant growth promoting bacterium, in enhancing algal biomass through mutualistic interaction. Biomass Bioenergy 2014, 69, 95–105. [Google Scholar] [CrossRef]
- Mujtaba, G.; Lee, K. Treatment of real wastewater using co-culture of immobilized Chlorella vulgaris and suspended activated sludge. Water Res. 2017, 120, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.H.; Ramanan, R.; Heo, J.; Lee, J.; Kim, B.H.; Oh, H.M.; Kim, H.S. Enhancing microalgal biomass productivity by engineering a microalgal-bacterial community. Bioresour. Technol. 2015, 175, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Bister, N.; Pistono, C.; Huremagic, B.; Jolkkonen, J.; Giugno, R.; Malm, T. Hypoxia and extracellular vesicles: A review on methods, vesicular cargo and functions. J. Extracell. Vesicles 2020, 10, e12002. [Google Scholar] [CrossRef] [PubMed]
- Yarana, C.; St. Clair, D. Chemotherapy-Induced Tissue Injury: An insight into the role of extracellular vesicles-mediated oxidative stress responses. Antioxidants 2017, 6, 75. [Google Scholar] [CrossRef] [PubMed]
- Abramowicz, A.; Widłak, P.; Pietrowska, M. Different Types of cellular stress affect the proteome composition of small extracellular vesicles: A mini review. Proteomes 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. European Commission A new Circular Economy Action Plan for a Cleaner and More Competitive Europe; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- García-Galán, M.J.; Arashiro, L.; Santos, L.H.M.L.M.; Insa, S.; Rodríguez-Mozaz, S.; Barceló, D.; Ferrer, I.; Garfí, M. Fate of priority pharmaceuticals and their main metabolites and transformation products in microalgae-based wastewater treatment systems. J. Hazard. Mater. 2020, 390, 121771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škufca, D.; Kovačič, A.; Prosenc, F.; Griessler Bulc, T.; Heath, D.; Heath, E. Phycoremediation of municipal wastewater: Removal of nutrients and contaminants of emerging concern. Sci. Total Environ. 2021, 782, 146949. [Google Scholar] [CrossRef]
- Cooper, M.B.; Smith, A.G.; Paszkowski, U.; Scott, B. Exploring mutualistic interactions between microalgae and bacteria in the omics age This review comes from a themed issue on Biotic interactions Edited by. Curr. Opin. Plant Biol. 2015, 26, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Kouzuma, A.; Watanabe, K. Exploring the potential of algae/bacteria interactions. Curr. Opin. Biotechnol. 2015, 33, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Woith, E.; Fuhrmann, G.; Melzig, M.F. Extracellular vesicles—Connecting kingdoms. Int. J. Mol. Sci. 2019, 20, 5695. [Google Scholar] [CrossRef] [Green Version]
- Dominique, M.; Simon, N.; Vaulot, D. Phytoplankton Cell Counting by Flow Cytometry. In Algal Culturing Techniqes, 1st ed.; Andersen, R.A., Ed.; Elsevier Academic Press: London, UK, 2005; pp. 253–267. [Google Scholar]
- Božič, D.; Hočevar, M.; Kisovec, M.; Pajnič, M.; Pađen, L.; Jeran, M.; Bedina Zavec, A.; Podobnik, M.; Kogej, K.; Iglič, A.; et al. Stability of Erythrocyte-derived nanovesicles assessed by light scattering and electron microscopy. Int. J. Mol. Sci. 2021, 22, 12772. [Google Scholar] [CrossRef] [PubMed]
- Lešer, V.; Drobne, D.; Pipan, Ž.; Milani, M.; Tatti, F. Comparison of different preparation methods of biological samples for FIB milling and SEM investigation. J. Microsc. 2009, 233, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Mammadova, R.; Fiume, I.; Bokka, R.; Kralj-Iglič, V.; Božič, D.; Kisovec, M.; Podobnik, M.; Zavec, A.B.; Hočevar, M.; Gellén, G.; et al. Identification of tomato infecting viruses that co-isolate with nanovesicles using a combined proteomics and electron-microscopic approach. Nanomaterials 2021, 11, 1922. [Google Scholar] [CrossRef]
- Škufca, D.; Kovačič, A.; Bulc, T.G.; Heath, E. Determination of 18 bisphenols in aqueous and biomass phase of high rate algal ponds: Development, validation and application. Chemosphere 2021, 271, 129786. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Dragulescu, A.; Arendt, C. Xlsx: Read, Write, Format Excel 2007 and Excel 97/2000/XP/2003 Files 2020. Available online: https://rdrr.io/cran/xlsx/ (accessed on 20 April 2022).
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests 2020. Available online: https://rdrr.io/github/kassambara/rstatix/ (accessed on 20 April 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Series | r * | F/2 [mL] | LB [mL] | Microalgae Inoculum [mL] | Bacterial Inoculum [mL] | Bisphenol Standard [µL] | Blank MeOH [µL] | Ill ** | |
|---|---|---|---|---|---|---|---|---|---|
| control series | abiotic dark (ABD) | 3 | 200 | 0 | 0 | 0 | 40 | 0 | NO |
| abiotic light (ABL) | 3 | 200 | 0 | 0 | 0 | 40 | 0 | YES | |
| blank axenic microalgae (BA) | 3 | 150 | 0 | 50 | 0 | 0 | 40 | YES | |
| blank co-culture (BC) | 3 | 149.5 | 0 | 50 | 0.5 | 0 | 40 | YES | |
| blank bacteria (BB) | 3 | 197.5 | 2 | 0 | 0.5 | 0 | 40 | YES | |
| experimental series | axenic microalgae (EA) | 3 | 150 | 0 | 50 | 0 | 40 | 0 | YES |
| co-culture (EC) | 3 | 149.5 | 0 | 50 | 0.5 | 40 | 0 | YES | |
| bacteria (EB) | 3 | 197.5 | 2 | 0 | 0.5 | 40 | 0 | YES |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Škufca, D.; Božič, D.; Hočevar, M.; Jeran, M.; Bedina Zavec, A.; Kisovec, M.; Podobnik, M.; Matos, T.; Tomazin, R.; Iglič, A.; et al. Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media. Int. J. Mol. Sci. 2022, 23, 8447. https://doi.org/10.3390/ijms23158447
Škufca D, Božič D, Hočevar M, Jeran M, Bedina Zavec A, Kisovec M, Podobnik M, Matos T, Tomazin R, Iglič A, et al. Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media. International Journal of Molecular Sciences. 2022; 23(15):8447. https://doi.org/10.3390/ijms23158447
Chicago/Turabian StyleŠkufca, David, Darja Božič, Matej Hočevar, Marko Jeran, Apolonija Bedina Zavec, Matic Kisovec, Marjetka Podobnik, Tadeja Matos, Rok Tomazin, Aleš Iglič, and et al. 2022. "Interaction between Microalgae P. tricornutum and Bacteria Thalassospira sp. for Removal of Bisphenols from Conditioned Media" International Journal of Molecular Sciences 23, no. 15: 8447. https://doi.org/10.3390/ijms23158447