
Transcriptome Analysis Reveals Key Gene Expression Changes in Blue Catfish Sperm in Response to Cryopreservation

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cryostorage Affects Sperm Motility, Increases the Level of Oxidative Stress, and Causes DNA Damage and Apoptosis
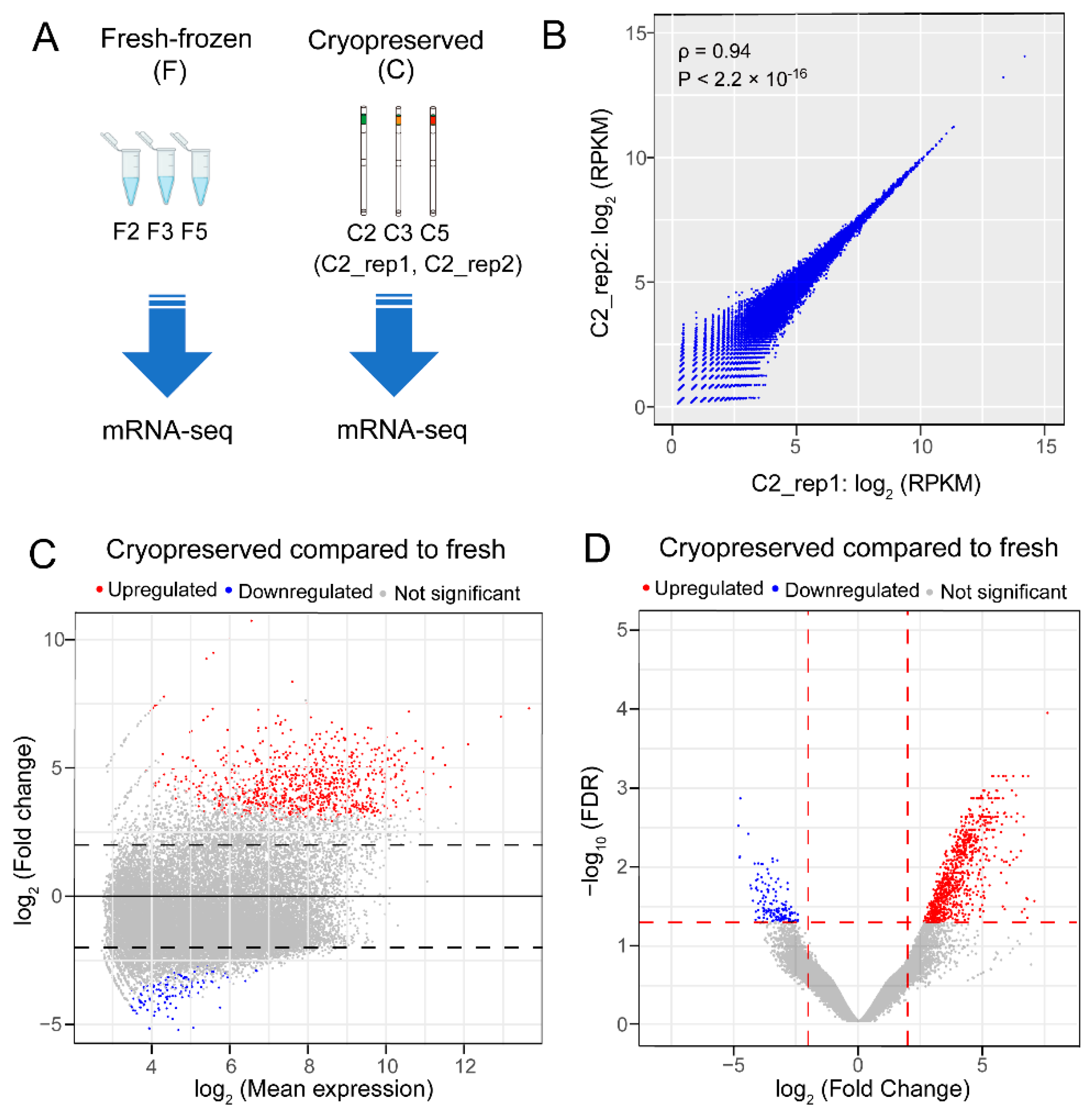
2.2. RNA Sequencing Revealed 849 Upregulated and 143 Downregulated Genes in the Blue Catfish Sperm Transcriptome after Cryopreservation
2.3. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Validation of Differentially Expressed Genes
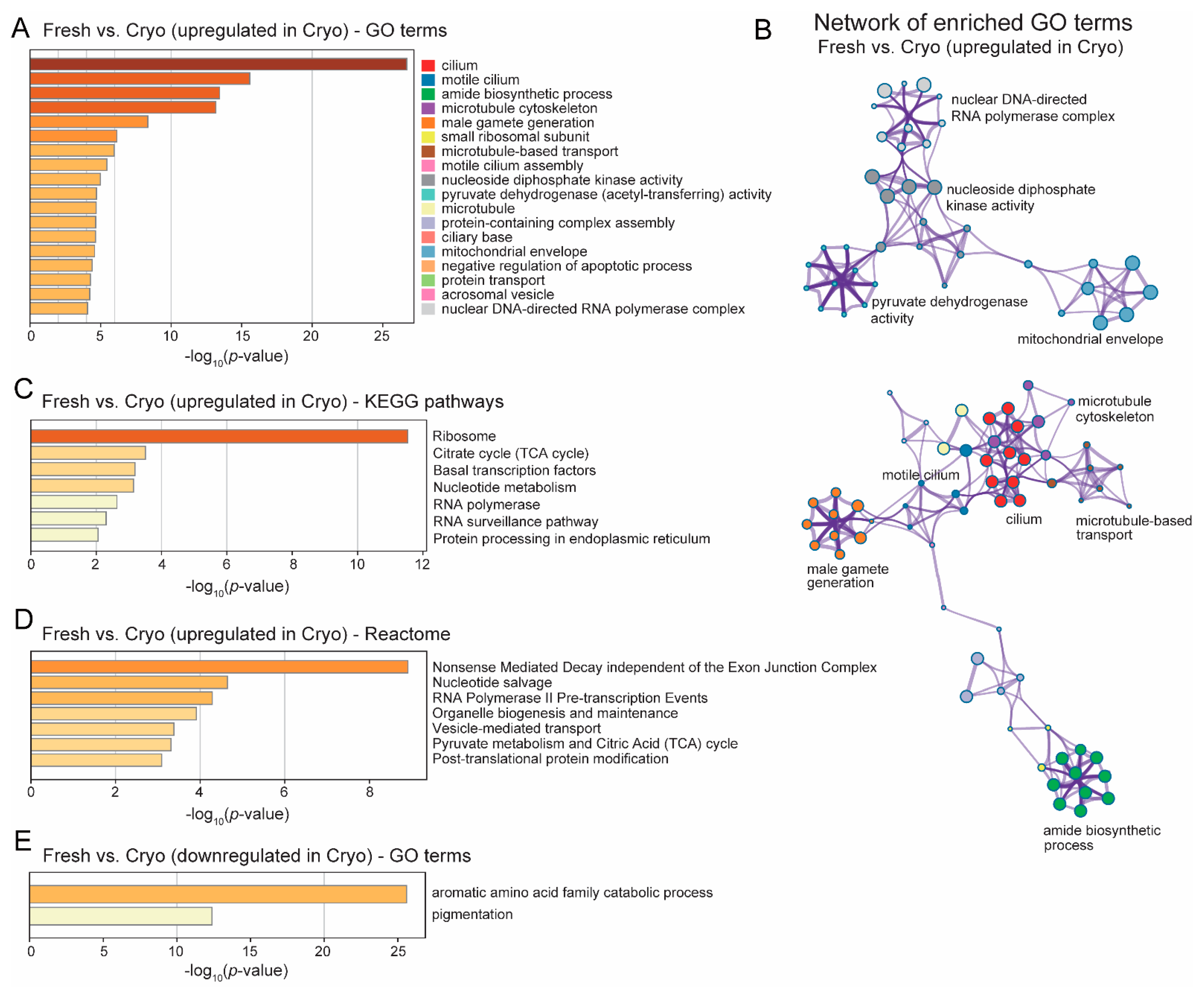
2.4. GO Analyses Revealed Upregulated Functional Clusters after Cryopreservation
2.5. The KEGG Pathway Enriched for Upregulated DEGs after Cryopreservation—mRNA Expression, Oxidative Phosphoralation and Mitochondrial ROS Regulation
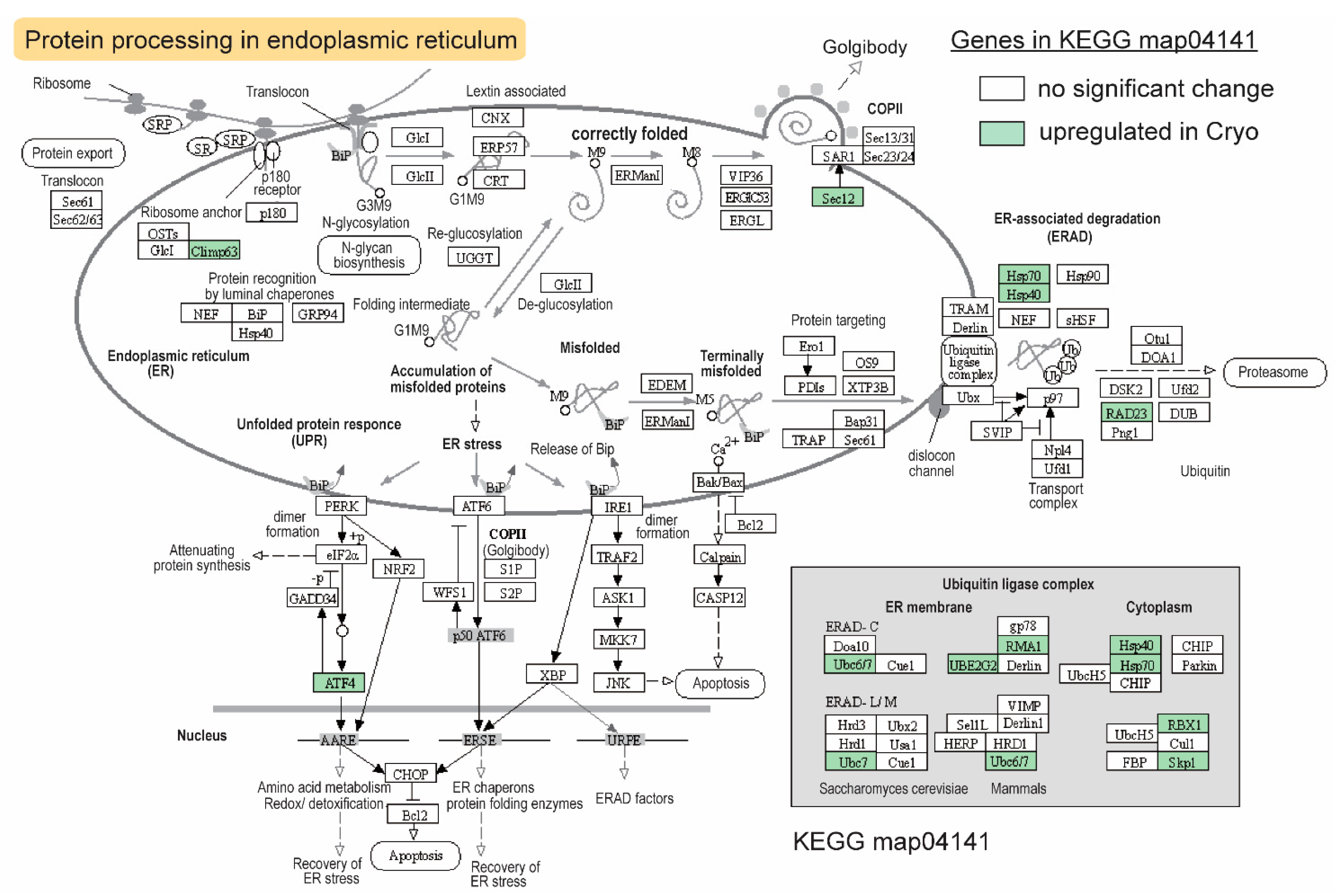
2.6. The KEGG Pathway Enriched for Upregulated DEGs after Cryopreservation—Heat Shock Proteins and Protein Ubiquitination/Degradation
2.7. The KEGG Pathway Enriched for Upregulated DEGs after Cryopreservation—Apoptosis and Necroptosis Pathways
3. Discussion
3.1. Sperm Quality Measurement Revealed Potential Impairment of Cryopreservation in Motility, DNA Fragmentation, and Level of Oxidative Stress
3.2. Cryopreservation on Sperm Transcriptome—Global Upregulation of Gene Expression, Increased Oxidative Phosphorylation, and Glycolysis/TCA Activities in the Mitochondria
3.3. Compensatory Changes after Cryopreservation in Sperm Transcriptome to Improve Cilia Structure and Sperm Motility
3.4. Compensatory Changes after Cryopreservation in Sperm Transcriptome—Negative Regulation of Apoptosis
3.5. Compensatory Changes after Cryopreservation in Sperm Transcriptome—Amide Biosynthesis
3.6. Compensatory Changes after Cryopreservation in Sperm Transcriptome—Managing ROS by Upregulation of MnSOD
3.7. Compensatory Changes after Cryopreservation in Sperm Transcriptome—ER-Stress-Related HSPs and Ubiquitin-Dependent Protein Degradation
3.8. Compensatory Changes after Cryopreservation in Sperm Transcriptome—Potential Alterations in Immune Functions?
3.9. Aromatic Amino Acid Catabolic Process-Related Genes Were Downregulated in Blue Catfish Sperm after Cryopreservation
3.10. Toward a Better Understanding of in Impact of Cryopreservation on Sperm, and Identification of Potential Biomarkers of Sperm Quality
4. Materials and Methods
4.1. Fish Maintenance, Sperm Sample Collection, and Cryopreservation
4.2. Sperm Cryopreservation
4.3. Sperm Quality Measurements
4.4. Total RNA Extraction, RNA-seq Library Construction, Quality Control, and Sequencing
4.5. RNA-seq QC, Genome Alignments, and Differential Gene Expression Analysis
4.6. Functional Enrichment and Pathway Analysis of Differentially Expressed Genes in Blue Catfish Sperm before and after Cryopreservation
4.7. Quantitative Reverse Transcription PCR Validation of Differentially Expressed Genes
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2013 Revision, Key Findings and Advance Tables; United Nations: New York, NY, USA, 2013. [Google Scholar]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Castel, V.; Rosales, M.; de Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food and Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Tomlinson, I. Doubling food production to feed the 9 billion: A critical perspective on a key discourse of food security in the UK. J. Rural. Stud. 2013, 29, 81–90. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Tacon, A.G.J. Global trends in aquaculture and compound aquafeed production. Mag. World Aquac. Soc. 2018, 49, 33–46. [Google Scholar]
- Food and Agriculture Organization. The State of World Fisheries and Aquaculture (SOFIA); Food and Agriculture Organization: Rome, Italy, 2020. [Google Scholar]
- Waite, R.; Beveridge, M.; Brummett, R.; Castine, S.; Chaiyawannakarn, N.; Kaushik, S.; Mungkung, R.; Nawapakpilai, S.; Phillips, M. Improving Productivity and Environmental Performance of Aquaculture; WorldFish: Penang, Malaysia, 2014. [Google Scholar]
- Dunham, R.A.; Ramboux, A.C.R.; Perera, D.A. Effect of strain on the growth, survival and sexual dimorphism of channel × blue catfish hybrids grown in earthen ponds. Aquaculture 2014, 420–421, S20–S24. [Google Scholar] [CrossRef]
- Su, B.; Perera, D.A.; Zohar, Y.; Abraham, E.; Stubblefield, J.; Fobes, M.; Beam, R.; Argue, B.; Ligeon, C.; Padi, J.; et al. Relative effectiveness of carp pituitary extract, luteininzing hormone releasing hormone analog (LHRHa) injections and LHRHa implants for producing hybrid catfish fry. Aquaculture 2013, 372–375, 133–136. [Google Scholar] [CrossRef]
- Bart, A.N.; Dunham, R.A. Effects of sperm concentration and egg number on fertilization efficiency with channel catfish (Ictalurus punctatus) eggs and blue catfish (I. furcatus) spermatozoa. Theriogenology 1996, 45, 673–682. [Google Scholar] [CrossRef]
- Dunham, R.A.; Brummett, R.E.; Ella, M.O.; Smitherman, R.O. Genotype-environment interactions for growth of blue, channel and hybrid catfish in ponds and cages at varying densities. Aquaculture 1990, 85, 143–151. [Google Scholar] [CrossRef]
- Dunham, R.A.; Smitherman, R.O.; Brooks, M.J.; Benchakan, M.; Chappell, J.A. Paternal predominance in reciprocal channel-blue hybrid catfish. Aquaculture 1982, 29, 389–396. [Google Scholar] [CrossRef]
- Yant, D.R.; Smitherman, R.O.; Green, O.L. Production of Hybrid (Blue × Channel) Catfish and Channel Catfish in Ponds. Proc. Annu. Conf. Southeast. Assoc. 1975, 82–86. [Google Scholar]
- Torrans, L.; Ott, B. Effect of grading fingerling hybrid catfish (♀ channel catfish × ♂ blue catfish) on growth, production, feed conversion, and food fish size distribution. N. Am. J. Aquac. 2018, 80, 187–192. [Google Scholar] [CrossRef]
- Dunham, R.A.; Umali, G.M.; Beam, R.; Kristanto, A.H.; Trask, M. Comparison of production traits of NWAC103 channel catfish, NWAC103 channel catfish× blue catfish hybrids, Kansas Select 21 channel catfish, and blue catfish grown at commercial densities and exposed to natural bacterial epizootics. N. Am. J. Aquac. 2008, 70, 98–106. [Google Scholar] [CrossRef]
- Brown, T.W.; Chappell, J.A.; Boyd, C.E. A commercial-scale, in-pond raceway system for Ictalurid catfish production. Aquac. Eng. 2011, 44, 72–79. [Google Scholar] [CrossRef]
- Bosworth, B.G. Effects of winter feeding on growth, body composition, and processing traits of co-cultured Blue Catfish, Channel Catfish, and Channel Catfish × Blue Catfish hybrids. N. Am. J. Aquac. 2012, 74, 553–559. [Google Scholar] [CrossRef]
- Dunham, R.A.; Smitherman, R.O.; Webber, C. Relative tolerance of channel x blue hybrid and channel catfish to low oxygen concentrations. Progress. Fish-Cult. 1983, 45, 55–57. [Google Scholar] [CrossRef]
- Arias, C.R.; Cai, W.; Peatman, E.; Bullard, S.A. Catfish hybrid Ictalurus punctatus × I. furcatus exhibits higher resistance to columnaris disease than the parental species. Dis. Aquat. Org. 2012, 100, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunham, R.A.; Masser, M.P. Production of Hybrid Catfish; Southern Regional Aquaculture Center: Stoneville, MI, USA, 2012; Volume 436. [Google Scholar]
- Bott, L.B.; Roy, L.A.; Hanson, T.R.; Chappell, J.; Whitis, G.N. Research verification of production practices using intensive aeration at a hybrid catfish operation. N. Am. J. Aquac. 2015, 77, 460–470. [Google Scholar] [CrossRef]
- Kumar, G.; Engle, C.R.; Hanson, T.R.; Tucker, C.S.; Brown, T.W.; Bott, L.B.; Roy, L.A.; Boyd, C.E.; Recsetar, M.S.; Park, J.; et al. Economics of alternative catfish production technologies. J. World Aquac. Soc. 2018, 49, 1039–1057. [Google Scholar] [CrossRef]
- Graham, K. A review of the biology and management of blue catfish. Am. Fish. Soc. Symp. 1999, 24, 37–49. [Google Scholar]
- Bart, A.N.; Dunham, R.A. Factors affecting survival of channel catfish after surgical removal of testes. Progress. Fish-Cult. 1990, 52, 241–246. [Google Scholar] [CrossRef]
- Dunham, R.A.; Bart, A.N.; Kucuktas, H. Effects of fertilization method and of selection for body weight and species on fertilization efficiency of channel catfish eggs with blue or channel catfish sperm. N. Am. J. Aquac. 1999, 61, 156–161. [Google Scholar] [CrossRef]
- Myers, J.N.; Chatakondi, N.G.; Dunham, R.A.; Butts, I.A.E. Genetic architecture of early life history traits for channel catfish, Ictalurus punctatus ♀ × blue catfish, I. furcatus ♂ hybrid production. Aquaculture 2020, 514, 734436. [Google Scholar] [CrossRef]
- Bart, A.N.; Wolfe, D.F.; Dunham, R.A. Cryopreservation of blue catfish spermatozoa and subsequent fertilization of channel catfish eggs. Trans. Am. Fish. Soc. 1998, 127, 819–824. [Google Scholar] [CrossRef]
- Hu, E.; Tiersch, T.R. Development of high-throughput cryopreservation for aquatic species. Cryopreserv. Aquat. Species 2011, 2, 995–1003. [Google Scholar]
- Hu, E.; Bosworth, B.; Baxter, J.; Tiersch, T.R. On-site evaluation of commercial-scale hybrid catfish production using cryopreserved blue catfish sperm. Aquaculture 2014, 426, 88–95. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Litvak, M.; Trippel, E. Seasonal variations in seminal plasma and sperm characteristics of wild-caught and cultivated Atlantic cod, Gadus morhua. Theriogenology 2010, 73, 873–885. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Babiak, I.; Ciereszko, A.; Litvak, M.; Słowińska, M.; Soler, C.; Trippel, E. Semen characteristics and their ability to predict sperm cryopreservation potential of Atlantic cod, Gadus morhua L. Theriogenology 2011, 75, 1290–1300. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Alavi, S.M.H.; Mokdad, A.; Pitcher, T.E. Physiological functions of osmolality and calcium ions on the initiation of sperm motility and swimming performance in redside dace, Clinostomus elongatus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 147–157. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Prokopchuk, G.; Kašpar, V.; Cosson, J.; Pitcher, T.E. Ovarian fluid impacts flagellar beating and biomechanical metrics of sperm between alternative reproductive tactics. J. Exp. Biol. 2017, 220, 2210–2217. [Google Scholar] [CrossRef] [Green Version]
- Butts, I.A.E.; Hilmarsdóttir, G.S.; Zadmajid, V.; Gallego, V.; Støttrup, J.G.; Jacobsen, C.; Krüger-Johnsen, M.; Politis, S.N.; Asturiano, J.F.; Holst, L.K. Dietary amino acids impact sperm performance traits for a catadromous fish, Anguilla anguilla reared in captivity. Aquaculture 2020, 518, 734602. [Google Scholar] [CrossRef]
- Gage, M.J.; Macfarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal traits and sperm competition in Atlantic salmon: Relative sperm velocity is the primary determinant of fertilization success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef]
- Babiak, I.; Ottesen, O.; Rudolfsen, G.; Johnsen, S. Quantitative characteristics of Atlantic halibut, Hippoglossus hippoglossus L., semen throughout the reproductive season. Theriogenology 2006, 65, 1587–1604. [Google Scholar] [CrossRef]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.G.; Sarasquete, C.; Herráez, M.P.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Acosta, I.B.; Corcini, C.D.; Gheller, S.M.M.; Brito, C.R.; Goulart, T.L.S.; Varela Junior, A.S. Effect of Amide on Semen Cryopreservation of Curimba (Prochilodus lineatus). Cryoletters 2020, 41, 1–5. [Google Scholar]
- Madrigal-Valverde, M.; Bittencourt, R.F.; de Lisboa Ribeiro Filho, A.; Araujo, G.R.; Lents, M.P.; Santos, E.S.; Lima, A.S.; Mattos, P. Can amides be alternative cryoprotectors for the preservation of feline semen? Cryobiology 2020, 97, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Pukazhenthi, B.S.; Johnson, A.; Guthrie, H.D.; Songsasen, N.; Padilla, L.R.; Wolfe, B.A.; Coutinho da Silva, M.; Alvarenga, M.A.; Wildt, D.E. Improved sperm cryosurvival in diluents containing amides versus glycerol in the Przewalski’s horse (Equus ferus przewalskii). Cryobiology 2014, 68, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, I.; Calderam, K.; Maschio, É.F.; Madeira, E.M.; da Rosa Ulguim, R.; Corcini, C.D.; Bongalhardo, D.C.; Corrêa, É.K.; Lucia, T.; Deschamps, J.C.; et al. Evaluation of amides and centrifugation temperature in boar semen cryopreservation. Theriogenology 2008, 69, 632–638. [Google Scholar] [CrossRef]
- Fridovich, I. The biology of oxygen radicals. Science 1978, 201, 875–880. [Google Scholar] [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Holley, A.K.; Bakthavatchalu, V.; Velez-Roman, J.M.; St Clair, D.K. Manganese superoxide dismutase: Guardian of the powerhouse. Int. J. Mol. Sci. 2011, 12, 7114–7162. [Google Scholar] [CrossRef]
- Kinnally, K.W.; Peixoto, P.M.; Ryu, S.Y.; Dejean, L.M. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochim. Biophys. Acta 2011, 1813, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Faust, O.; Abayev-Avraham, M.; Wentink, A.S.; Maurer, M.; Nillegoda, N.B.; London, N.; Bukau, B.; Rosenzweig, R. HSP40 proteins use class-specific regulation to drive HSP70 functional diversity. Nature 2020, 587, 489–494. [Google Scholar] [CrossRef]
- Watson, P.; Holt, W. Principles of Cryopreservation. Cryobanking the Genetic Resource: Wildlife Conservation for the Future; CRC Press: Boca Raton, FL, USA, 2001; pp. 23–46. [Google Scholar]
- Watson, P.F.; Morris, G.J. Cold shock injury in animal cells. Symp. Soc. Exp. Biol. 1987, 41, 311–340. [Google Scholar] [PubMed]
- Miki, K. Energy metabolism and sperm function. Soc. Reprod. Fertil. Suppl. 2007, 65, 309–325. [Google Scholar] [PubMed]
- Raad, G.; Lteif, L.; Lahoud, R.; Azoury, J.; Azoury, J.; Tanios, J.; Hazzouri, M.; Azoury, J. Cryopreservation media differentially affect sperm motility, morphology and DNA integrity. Andrology 2018, 6, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Lahnsteiner, F.; Berger, B.; Weismann, T.; Patzner, R. Changes in morphology, physiology, metabolism, and fertilization capacity of rainbow trout semen following cryopreservation. Progress. Fish-Cult. 1996, 58, 149–159. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Mohanty, G.; Van der Linde, M. Oxidative phosphorylation versus glycolysis: What fuel do spermatozoa use? Asian J. Androl. 2015, 17, 230. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cano, F.E.; Gaitskell-Phillips, G.; Ortiz-Rodríguez, J.M.; Silva-Rodríguez, A.; Román, Á.; Rojo-Domínguez, P.; Alonso-Rodríguez, E.; Tapia, J.A.; Gil, M.C.; Ortega-Ferrusola, C.; et al. Proteomic profiling of stallion spermatozoa suggests changes in sperm metabolism and compromised redox regulation after cryopreservation. J. Proteom. 2020, 221, 103765. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, F.; Malama, E.; Siuda, M.; Leiding, C.; Bollwein, H. Effects of sodium pyruvate on viability, synthesis of reactive oxygen species, lipid peroxidation and DNA integrity of cryopreserved bovine sperm. Anim. Reprod. Sci. 2017, 185, 18–27. [Google Scholar] [CrossRef]
- Dacheux, D.; Roger, B.; Bosc, C.; Landrein, N.; Roche, E.; Chansel, L.; Trian, T.; Andrieux, A.; Papaxanthos-Roche, A.; Marthan, R.; et al. Human FAM154A (SAXO1) is a microtubule-stabilizing protein specific to cilia and related structures. J. Cell Sci. 2015, 128, 1294–1307. [Google Scholar] [CrossRef] [Green Version]
- Chawan, V.; Yevate, S.; Gajbhiye, R.; Kulkarni, V.; Parte, P. Acetylation/deacetylation and microtubule associated proteins influence flagellar axonemal stability and sperm motility. Biosci. Rep. 2020, 40, BSR20202442. [Google Scholar] [CrossRef]
- Tang, S.; Wang, X.; Li, W.; Yang, X.; Li, Z.; Liu, W.; Li, C.; Zhu, Z.; Wang, L.; Wang, J.; et al. Biallelic mutations in CFAP43 and CFAP44 cause male infertility with multiple morphological abnormalities of the sperm flagella. Am. J. Hum. Genet. 2017, 100, 854–864. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wu, H.; Li, F.; Tian, S.; Kherraf, Z.-E.; Zhang, J.; Ni, X.; Lv, M.; Liu, C.; Tan, Q.; et al. Biallelic mutations in CFAP65 cause male infertility with multiple morphological abnormalities of the sperm flagella in humans and mice. J. Med. Genet. 2020, 57, 89–95. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, X.; Lian, S.; Li, Y.; Hu, N.; Hu, X.; Bao, Z.; Wang, S. Functional Characterization of Cfap206 for Bivalve Ciliogenesis by RNAi and CRISPR/Cas9 Technologies. Front. Mar. Sci. 2022, 392. [Google Scholar] [CrossRef]
- Aitken, R.J.; Findlay, J.K.; Hutt, K.J.; Kerr, J.B. Apoptosis in the germ line. Reproduction 2011, 141, 139. [Google Scholar] [CrossRef] [Green Version]
- Paasch, U.; Sharma, R.K.; Gupta, A.K.; Grunewald, S.; Mascha, E.J.; Thomas Jr, A.J.; Glander, H.-J.; Agarwal, A. Cryopreservation and thawing is associated with varying extent of activation of apoptotic machinery in subsets of ejaculated human spermatozoa. Biol. Reprod. 2004, 71, 1828–1837. [Google Scholar] [CrossRef] [Green Version]
- Cisternas, P.; Moreno, R.D. Comparative analysis of apoptotic pathways in rat, mouse, and hamster spermatozoa. Mol. Reprod. Dev. 2006, 73, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Sabido, O.; Durand, P.; Levy, R. Cryopreservation induces an apoptosis-like mechanism in bull sperm. Biol. Reprod. 2004, 71, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesco, M.F.; Oliveira, C.; Soares, F.; Gavaia, P.J.; Dinis, M.T.; Cabrita, E. Solea senegalensis sperm cryopreservation: New insights on sperm quality. PLoS ONE 2017, 12, e0186542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef]
- Kluck, R.M.; Bossy-Wetzel, E.; Green, D.R.; Newmeyer, D.D. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, J.C.; Arnoult, D.; Youle, R.J. Control of mitochondrial permeability by Bcl-2 family members. Biochim. Biophys. Acta 2004, 1644, 107–113. [Google Scholar] [CrossRef]
- Starheim, K.K.; Gromyko, D.; Evjenth, R.; Ryningen, A.; Varhaug, J.E.; Lillehaug, J.R.; Arnesen, T. Knockdown of human Nα-terminal acetyltransferase complex C leads to p53-dependent apoptosis and aberrant human Arl8b localization. Mol. Cell. Biol. 2009, 29, 3569–3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, M.A.; Papa, F.O.; Landim-Alvarenga, F.C.; Medeiros, A.S.L. Amides as cryoprotectants for freezing stallion semen: A review. Anim. Reprod. Sci. 2005, 89, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ma, J.; Wan, Z.; Wang, Q.; Wang, Z.; Zhao, J.; Wang, F.; Zhang, Y. Characterization of sheep spermatogenesis through single-cell RNA sequencing. FASEB J. 2021, 35, e21187. [Google Scholar] [CrossRef]
- Malafa, M.; Margenthaler, J.; Webb, B.; Neitzel, L.; Christophersen, M. MnSOD expression is increased in metastatic gastric cancer. J. Surg. Res. 2000, 88, 130–134. [Google Scholar] [CrossRef]
- Wispe, J.; Warner, B.B.; Clark, J.C.; Dey, C.R.; Neuman, J.; Glasser, S.W.; Crapo, J.; Chang, L.; Whitsett, J.A. Human Mn-superoxide dismutase in pulmonary epithelial cells of transgenic mice confers protection from oxygen injury. J. Biol. Chem. 1992, 267, 23937–23941. [Google Scholar] [CrossRef]
- Lebovitz, R.M.; Zhang, H.; Vogel, H.; Cartwright, J.; Dionne, L.; Lu, N.; Huang, S.; Matzuk, M.M. Neurodegeneration, myocardial injury, and perinatal death in mitochondrial superoxide dismutase-deficient mice. Proc. Natl. Acad. Sci. USA 1996, 93, 9782–9787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murawski, M.; Saczko, J.; Marcinkowska, A.; Chwiłkowska, A.; Gryboś, M.; Banaś, T. Evaluation of superoxide dismutase activity and its impact on semen quality parameters of infertile men. Folia Histochem. Cytobiol. 2007, 45, 123–126. [Google Scholar]
- Westfalewicz, B.; Dietrich, M.; Ciereszko, A. Impact of cryopreservation on bull (Bos taurus) semen proteome. J. Anim. Sci. 2015, 93, 5240–5253. [Google Scholar] [CrossRef]
- Kaul, S.C.; Obuchi, K.; Iwahashi, H.; Komatsu, Y. Cryoprotection provided by heat shock treatment in Saccharomyces cerevisiae. Cell Mol. Biol. 1992, 38, 135–143. [Google Scholar]
- Feng, L.; Chen, J. The E3 ligase RNF8 regulates KU80 removal and NHEJ repair. Nat. Struct. Mol. Biol. 2012, 19, 201–206. [Google Scholar] [CrossRef]
- Guo, Y.; Song, Y.; Guo, Z.; Hu, M.; Liu, B.; Duan, H.; Wang, L.; Yuan, T.; Wang, D. Function of RAD6B and RNF8 in spermatogenesis. Cell Cycle 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Azenabor, A.; Ekun, A.O.; Akinloye, O. Impact of Inflammation on Male Reproductive Tract. J. Reprod. Infertil. 2015, 16, 123–129. [Google Scholar] [PubMed]
- Wang, F.; Chen, R.; Han, D. Innate immune defense in the male reproductive system and male fertility. In Innate Immunity in Health and Disease; IntechOpen: London, UK, 2019. [Google Scholar]
- Bai, H.; Sun, F.; Yang, G.; Wang, L.; Zhang, Q.; Zhang, Q.; Zhan, Y.; Chen, J.; Yu, M.; Li, C.; et al. CBLB502, a Toll-like receptor 5 agonist, offers protection against radiation-induced male reproductive system damage in mice. Biol. Reprod. 2019, 100, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, J.; Fu, W.; Xu, W.; Zhang, H.; Chen, S.; Liu, W.; Peng, L.; Xiao, Y. Comparative Transcriptome and DNA methylation analyses of the molecular mechanisms underlying skin color variations in Crucian carp (Carassius carassius L.). BMC Genet. 2017, 18, 95. [Google Scholar] [CrossRef]
- Ohnishi, K.; Saito, S.; Miura, T.; Ohta, A.; Tominaga, M.; Sokabe, T.; Kuhara, A. OSM-9 and OCR-2 TRPV channels are accessorial warm receptors in Caenorhabditis elegans temperature acclimatisation. Sci. Rep. 2020, 10, 18566. [Google Scholar]
- Sonoda, S.; Ohta, A.; Maruo, A.; Ujisawa, T.; Kuhara, A. Sperm affects head sensory neuron in temperature tolerance of Caenorhabditis elegans. Cell Rep. 2016, 16, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Del Signore, S.J.; Biber, S.A.; Lehmann, K.S.; Heimler, S.R.; Rosenfeld, B.H.; Eskin, T.L.; Sweeney, S.T.; Rodal, A.A. dOCRL maintains immune cell quiescence by regulating endosomal traffic. PLoS Genet. 2017, 13, e1007052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kang, S.A.; Mukherjee, T.; Bale, S.; Crane, B.R.; Begley, T.P.; Ealick, S.E. Crystal structure and mechanism of tryptophan 2, 3-dioxygenase, a heme enzyme involved in tryptophan catabolism and in quinolinate biosynthesis. Biochemistry 2007, 46, 145–155. [Google Scholar] [CrossRef]
- Zhai, L.; Ladomersky, E.; Lenzen, A.; Nguyen, B.; Patel, R.; Lauing, K.L.; Wu, M.; Wainwright, D.A. IDO1 in cancer: A Gemini of immune checkpoints. Cell Mol. Immunol. 2018, 15, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Jrad-Lamine, A.; Henry-Berger, J.; Gourbeyre, P.; Damon-Soubeyrand, C.; Lenoir, A.; Combaret, L.; Saez, F.; Kocer, A.; Tone, S.; Fuchs, D.; et al. Deficient tryptophan catabolism along the kynurenine pathway reveals that the epididymis is in a unique tolerogenic state. J. Biol. Chem. 2011, 286, 8030–8042. [Google Scholar] [CrossRef] [Green Version]
- Routy, J.-P.; Routy, B.; Graziani, G.M.; Mehraj, V. The kynurenine pathway is a double-edged sword in immune-privileged sites and in cancer: Implications for immunotherapy. Int. J. Tryptophan Res. 2016, 9, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Wang, X.; Liu, P.; Liu, H.; Li, R.; Li, Z.; He, Y.; Qi, J. Effects of Cryopreservation on Sperm with Cryodiluent in Viviparous Black Rockfish (Sebastes schlegelii). Int. J. Mol. Sci. 2022, 23, 3392. [Google Scholar] [CrossRef] [PubMed]
- Bizuayehu, T.T.; Babiak, I. MicroRNA in teleost fish. Genome Biol. Evol. 2014, 6, 1911–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles, V.; Valcarce, D.G.; Riesco, M.F. Non-coding RNA regulation in reproduction: Their potential use as biomarkers. Non-Coding RNA Res. 2019, 4, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Tiersch, T.R.; Goudie, C.A.; Carmichael, G.J. Cryopreservation of channel catfish sperm: Storage in cryoprotectants, fertilization trials, and growth of channel catfish produced with cryopreserved sperm. Trans. Am. Fish. Soc. 1994, 123, 580–586. [Google Scholar] [CrossRef]
- Hu, E.; Yang, H.; Tiersch, T.R. High-throughput cryopreservation of spermatozoa of blue catfish (Ictalurus furcatus): Establishment of an approach for commercial-scale processing. Cryobiology 2011, 62, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Gosálvez, J.; López-Fernández, C.; Hermoso, A.; Fernández, J.L.; Kjelland, M.E. Sperm DNA fragmentation in zebrafish (Danio rerio) and its impact on fertility and embryo viability—Implications for fisheries and aquaculture. Aquaculture 2014, 433, 173–182. [Google Scholar] [CrossRef]
- Wang, H.; Bruce, T.J.; Su, B.; Li, S.; Dunham, R.A.; Wang, X. Environment-Dependent Heterosis and Transgressive Gene Expression in Reciprocal Hybrids between the Channel Catfish Ictalurus punctatus and the Blue Catfish Ictalurus furcatus. Biology 2022, 11, 117. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 7 May 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Su, B.; Butts, I.A.E.; Dunham, R.; Wang, X. Chromosome-level assembly and annotation of the blue catfish Ictalurus furcatus, an aquaculture species for hybrid catfish reproduction, epigenetics, and heterosis studies. Gigascience 2022, 11, giac070. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, S.; Yao, J.; Bao, L.; Zhang, J.; Li, Y.; Jiang, C.; Sun, L.; Wang, R.; Zhang, Y.; et al. The channel catfish genome sequence provides insights into the evolution of scale formation in teleosts. Nat. Commun. 2016, 7, 11757. [Google Scholar] [CrossRef] [Green Version]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Törönen, P.; Medlar, A.; Holm, L. PANNZER2: A rapid functional annotation web server. Nucleic Acids Res. 2018, 46, W84–W88. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Montague, H.R.; Hess, H.N.; Zhang, Y.; Aguilar, G.L.; Dunham, R.A.; Butts, I.A.E.; Wang, X. Transcriptome Analysis Reveals Key Gene Expression Changes in Blue Catfish Sperm in Response to Cryopreservation. Int. J. Mol. Sci. 2022, 23, 7618. https://doi.org/10.3390/ijms23147618
Wang H, Montague HR, Hess HN, Zhang Y, Aguilar GL, Dunham RA, Butts IAE, Wang X. Transcriptome Analysis Reveals Key Gene Expression Changes in Blue Catfish Sperm in Response to Cryopreservation. International Journal of Molecular Sciences. 2022; 23(14):7618. https://doi.org/10.3390/ijms23147618
Chicago/Turabian StyleWang, Haolong, Helen R. Montague, Hana N. Hess, Ying Zhang, Gavin L. Aguilar, Rex A. Dunham, Ian A. E. Butts, and Xu Wang. 2022. "Transcriptome Analysis Reveals Key Gene Expression Changes in Blue Catfish Sperm in Response to Cryopreservation" International Journal of Molecular Sciences 23, no. 14: 7618. https://doi.org/10.3390/ijms23147618