
Loss of TRIM67 Attenuates the Progress of Obesity-Induced Non-Alcoholic Fatty Liver Disease

 , , and
, , and
Abstract
:1. Introduction
2. Results
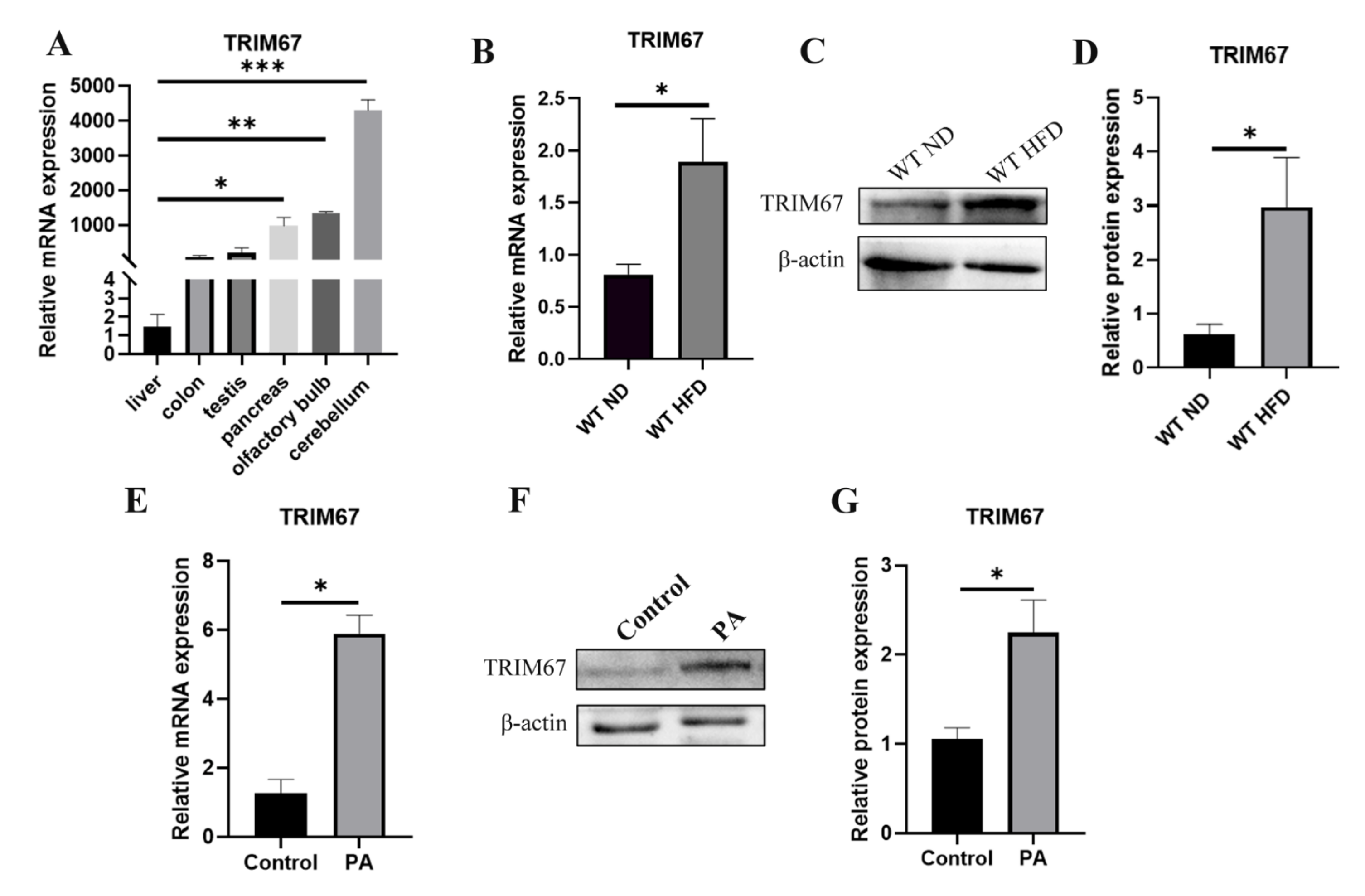
2.1. High-Fat Diet Induces the Expression of TRIM67
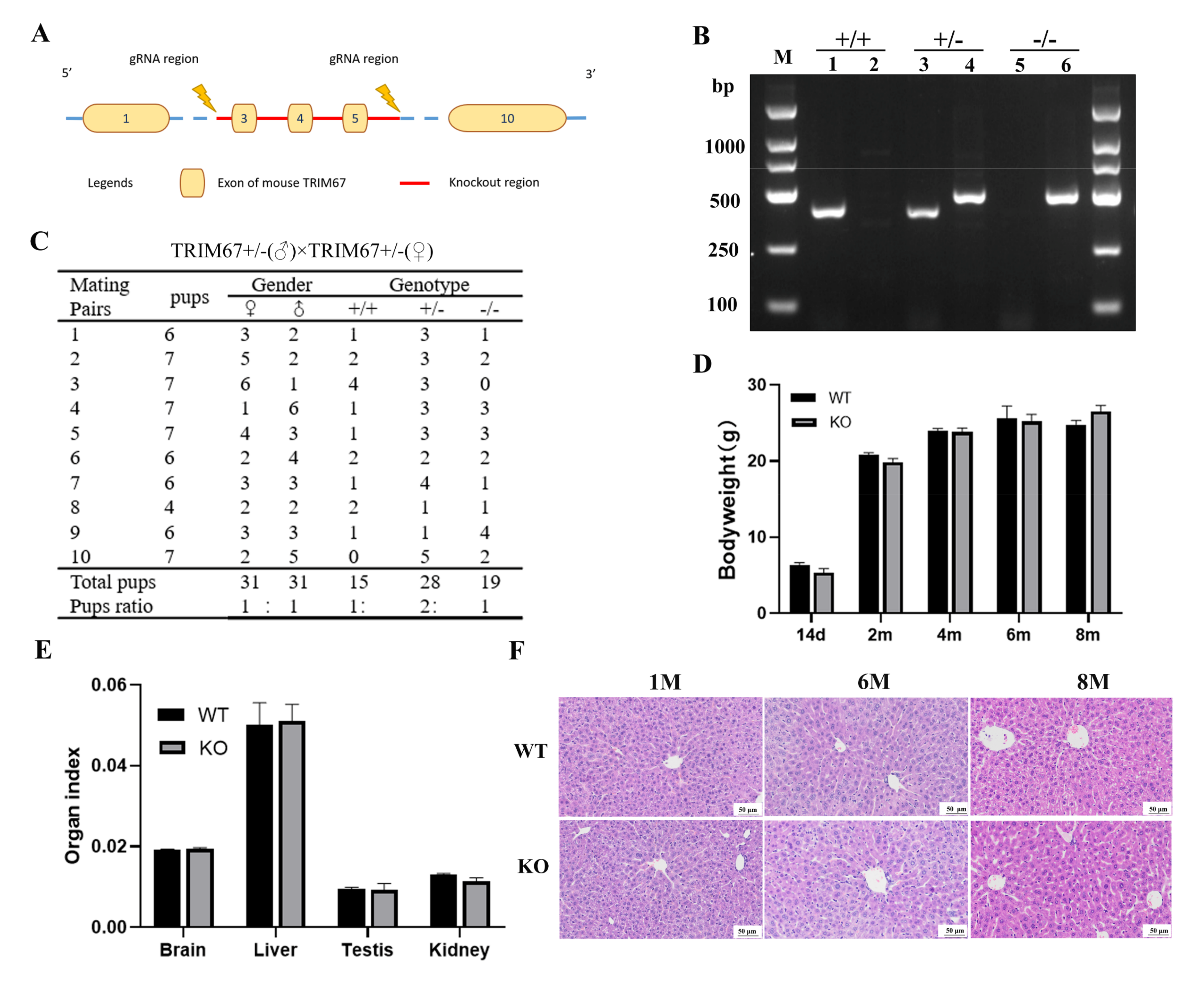
2.2. TRIM67 Is Not Essential for Normal Development of Mouse Liver
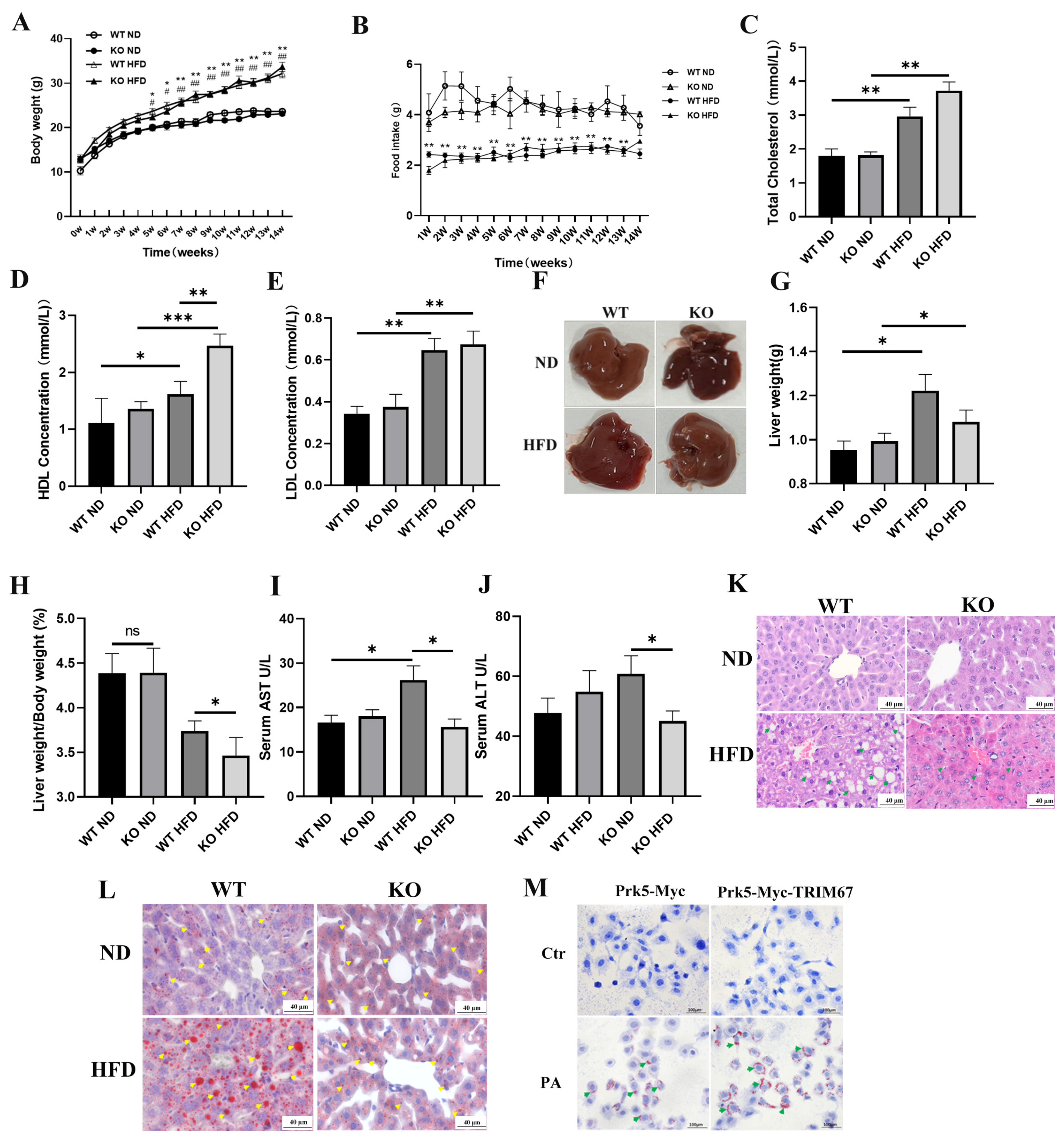
2.3. Deficiency of TRIM67 Is Protective against Hepatic Lipid Accumulation in Obese Mice
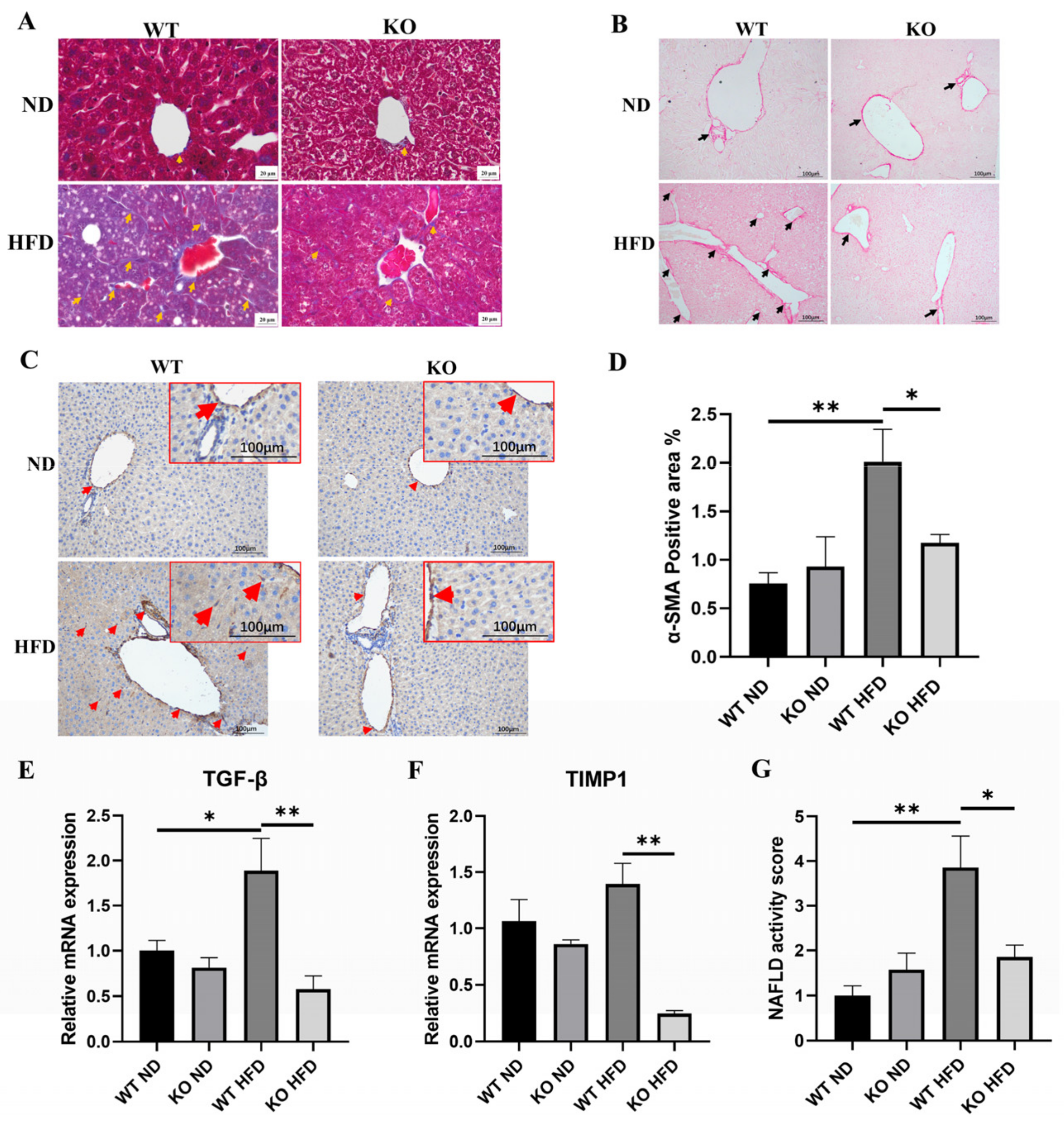
2.4. TRIM67 KO Improves Hepatic Fibrosis Induced by High-Fat-Diet
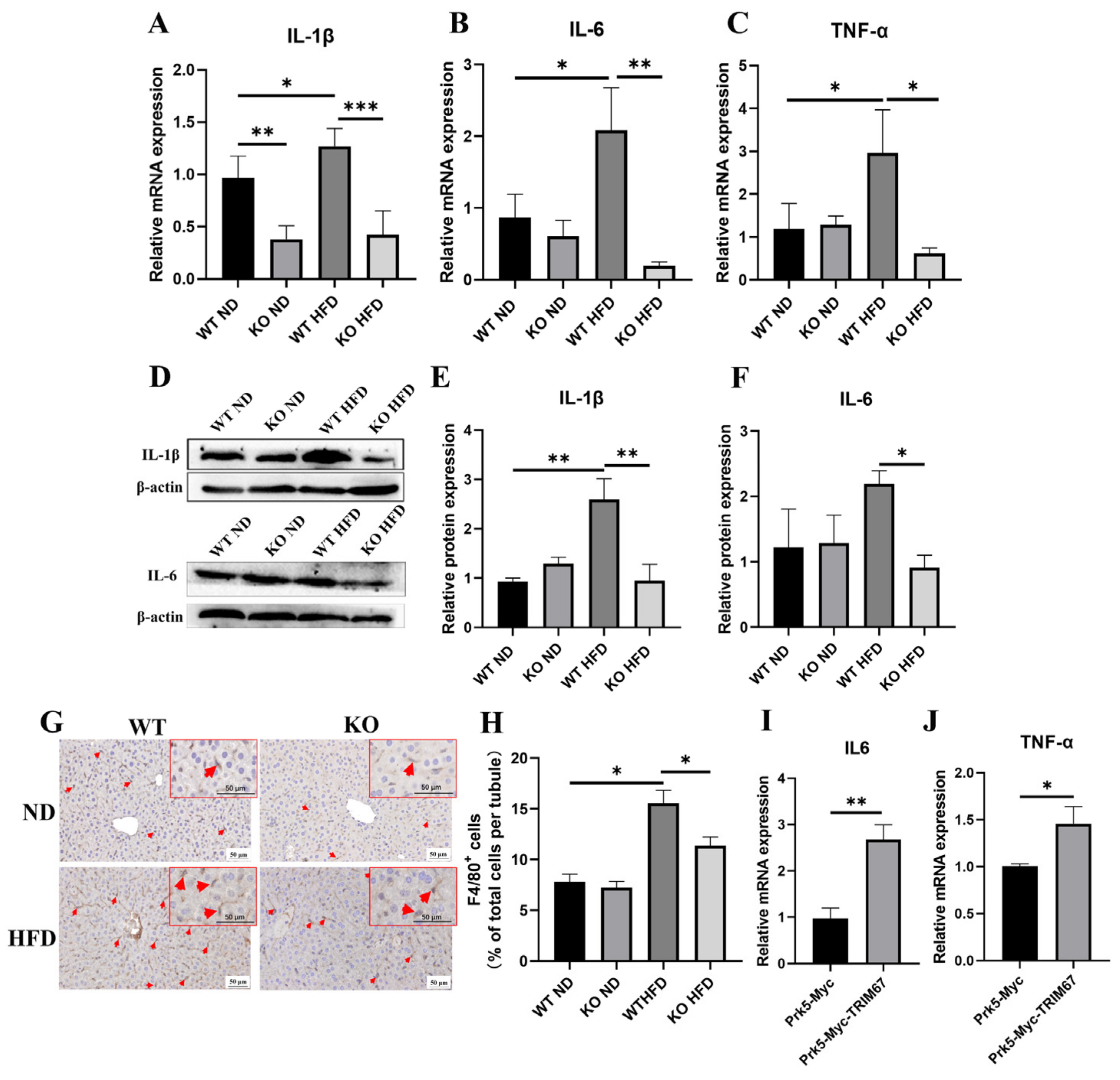
2.5. TRIM67 Implicates in the Regulation of Hepatic Inflammation and Lipid Homeostasis
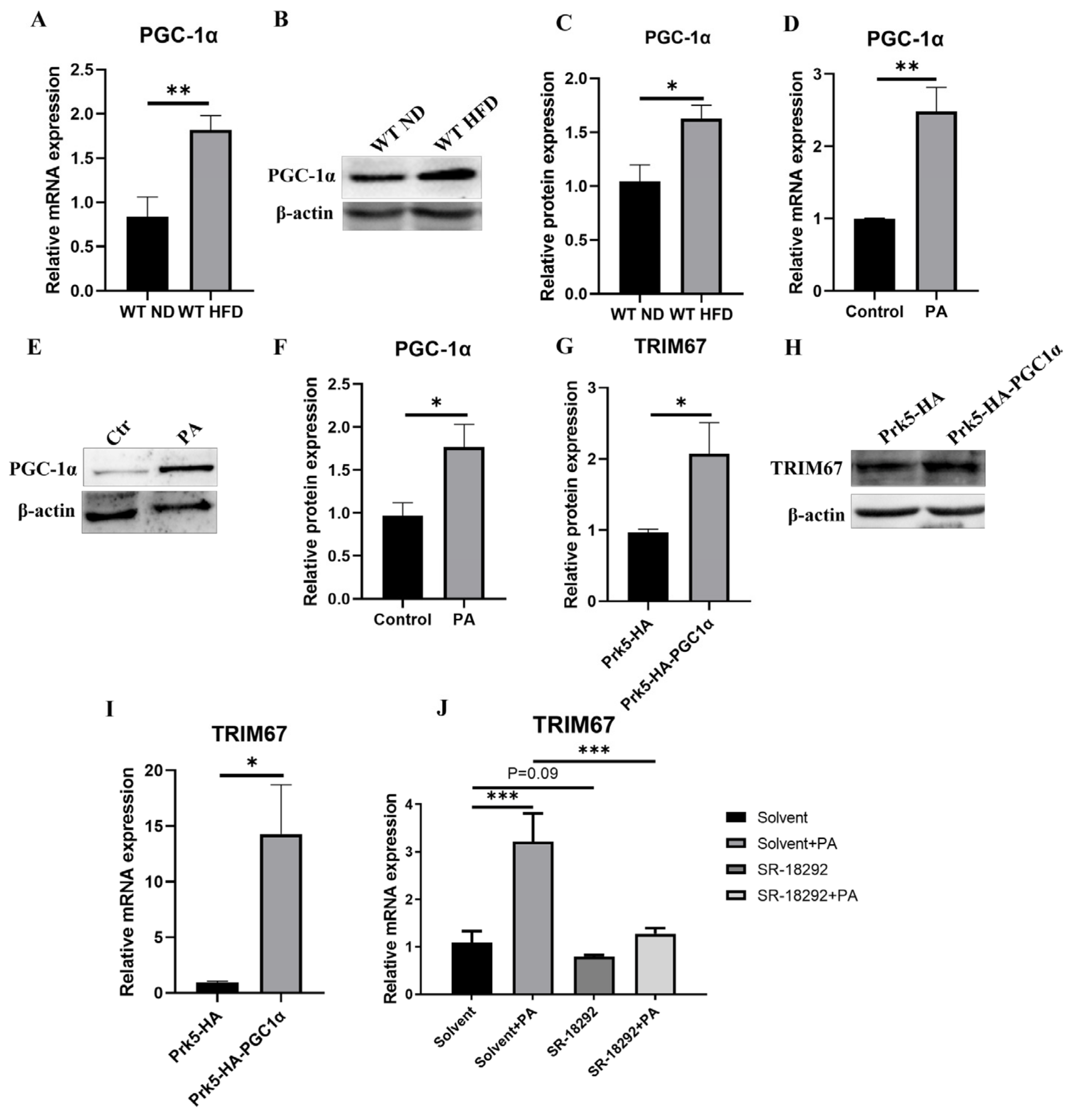
2.6. PGC-1α Induces Hepatic TRIM67 in Obese Mice
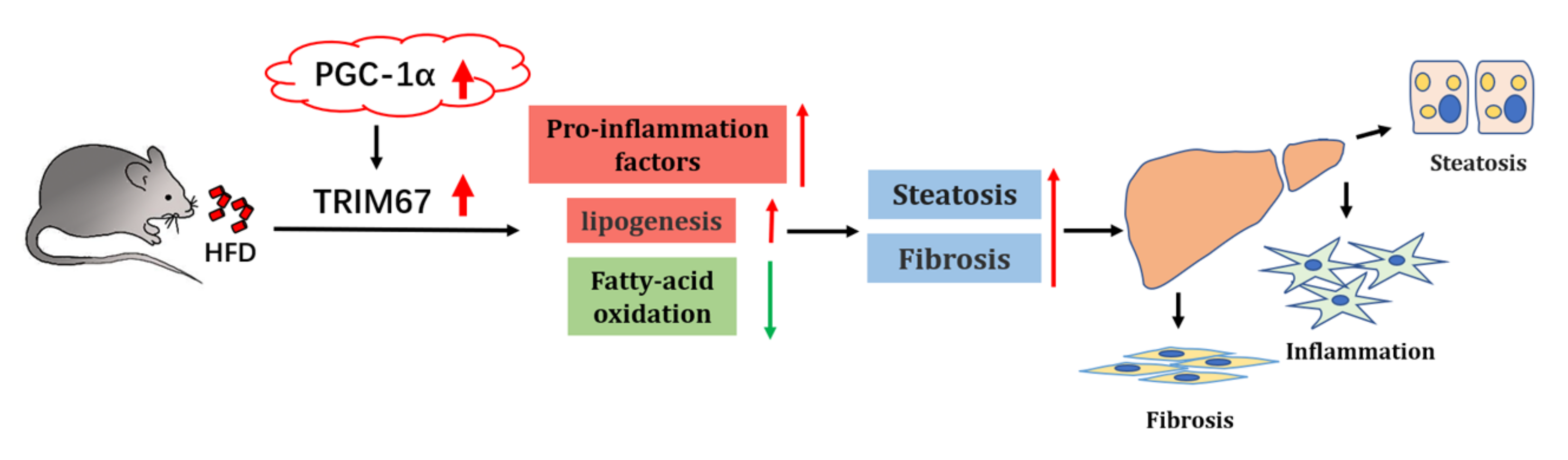
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. Induction of Obesity/NAFLD with High-Fat Diet
4.3. Histological Staining
4.4. Immunohistochemical Staining
4.5. Quantitative Realtime PCR
4.6. Western Blots
4.7. Cell Culture
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janssen, F.; Bardoutsos, A.; Vidra, N. Obesity Prevalence in the Long-Term Future in 18 European Countries and in the USA. Obes. Facts 2020, 13, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.M.; Danaei, G.; Farzadfar, F.; Stevens, G.A.; Woodward, M.; Wormser, D.; Kaptoge, S.; Whitlock, G.; Qiao, Q.; Lewington, S.; et al. The age-specific quantitative effects of metabolic risk factors on cardiovascular diseases and diabetes: A pooled analysis. PLoS ONE 2013, 8, e65174. [Google Scholar] [CrossRef] [PubMed]
- Czernichow, S.; Kengne, A.P.; Stamatakis, E.; Hamer, M.; Batty, G.D. Body mass index, waist circumference and waist-hip ratio: Which is the better discriminator of cardiovascular disease mortality risk? Evidence from an individual-participant meta-analysis of 82,864 participants from nine cohort studies. Obes. Rev. 2011, 12, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K.; International Agency for Research on Cancer Handbook Working, G. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [Green Version]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metab. Clin. Exp. 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Qureshi, K.; Abrams, G.A. Metabolic liver disease of obesity and role of adipose tissue in the pathogenesis of nonalcoholic fatty liver disease. World J. Gastroenterol. 2007, 13, 3540–3553. [Google Scholar] [CrossRef] [Green Version]
- Porro, S.; Genchi, V.A.; Cignarelli, A.; Natalicchio, A.; Laviola, L.; Giorgino, F.; Perrini, S. Dysmetabolic adipose tissue in obesity: Morphological and functional characteristics of adipose stem cells and mature adipocytes in healthy and unhealthy obese subjects. J. Endocrinol. Investig. 2021, 44, 921–941. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef]
- Hardy, T.; Oakley, F.; Anstee, Q.M.; Day, C.P. Nonalcoholic fatty liver disease: Pathogenesis and disease spectrum. Annu. Rev. Pathol. Mech. Dis. 2016, 11, 451–496. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metab. Clin. Exp. 2019, 92, 82–97. [Google Scholar] [CrossRef]
- Otgonsuren, M.; Stepanova, M.; Gerber, L.; Younossi, Z.M. Anthropometric and clinical factors associated with mortality in subjects with nonalcoholic fatty liver disease. Dig. Dis. Sci. 2013, 58, 1132–1140. [Google Scholar] [CrossRef]
- Bugianesi, E. Nonalcoholic fatty liver disease (NAFLD) and cardiac lipotoxicity: Another piece of the puzzle. Hepatology 2008, 47, 2–4. [Google Scholar] [CrossRef]
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev. Gastroenterol. Hepatol. 2009, 3, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Gushchina, L.V.; Kwiatkowski, T.A.; Bhattacharya, S.; Weisleder, N.L. Conserved structural and functional aspects of the tripartite motif gene family point towards therapeutic applications in multiple diseases. Pharmacol. Ther. 2018, 185, 12–25. [Google Scholar] [CrossRef]
- Hatakeyama, S. TRIM Family Proteins: Roles in Autophagy, Immunity, and Carcinogenesis. Trends Biochem. Sci. 2017, 42, 297–311. [Google Scholar] [CrossRef]
- Reymond, A.; Meroni, G.; Fantozzi, A.; Merla, G.; Cairo, S.; Luzi, L.; Riganelli, D.; Zanaria, E.; Messali, S.; Cainarca, S.; et al. The tripartite motif family identifies cell compartments. EMBO J. 2001, 20, 2140–2151. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, S. TRIM proteins and cancer. Nat. Rev. Cancer 2011, 11, 792–804. [Google Scholar] [CrossRef]
- Boyer, N.P.; Monkiewicz, C.; Menon, S.; Moy, S.S.; Gupton, S.L. Mammalian TRIM67 Functions in Brain Development and Behavior. eNeuro 2018, 5, ENEURO.0186-18.2018. [Google Scholar] [CrossRef] [Green Version]
- Menon, S.; Goldfarb, D.; Ho, C.T.; Cloer, E.W.; Boyer, N.P.; Hardie, C.; Bock, A.J.; Johnson, E.C.; Anil, J.; Major, M.B.; et al. The TRIM9/TRIM67 neuronal interactome reveals novel activators of morphogenesis. Mol. Biol. Cell 2021, 32, 314–330. [Google Scholar] [CrossRef]
- Jiang, J.; Ren, H.; Xu, Y.; Wudu, M.; Wang, Q.; Liu, Z.; Su, H.; Jiang, X.; Zhang, Y.; Zhang, B.; et al. TRIM67 Promotes the Proliferation, Migration, and Invasion of Non-Small-Cell Lung Cancer by Positively Regulating the Notch Pathway. J. Cancer 2020, 11, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Chen, Y.; Shou, T.; Hu, J.; Chen, J.; Qing, C. TRIM67 promotes NFkappaB pathway and cell apoptosis in GA13315treated lung cancer cells. Mol. Med. Rep. 2019, 20, 2936–2944. [Google Scholar]
- Liu, Y.; Wang, G.; Jiang, X.; Li, W.; Zhai, C.; Shang, F.; Chen, S.; Zhao, Z.; Yu, W. TRIM67 inhibits tumor proliferation and metastasis by mediating MAPK11 in Colorectal Cancer. J. Cancer 2020, 11, 6025–6037. [Google Scholar] [CrossRef]
- Fan, W.; Liu, X.; Zhang, J.; Qin, L.; Du, J.; Li, X.; Qian, S.; Chen, H.; Qian, P. TRIM67 Suppresses TNFalpha-Triggered NF-kB Activation by Competitively Binding Beta-TrCP to IkBa. Front. Immunol. 2022, 13, 793147. [Google Scholar] [CrossRef]
- Vunjak, M.; Versteeg, G.A. TRIM proteins. Curr. Biol. 2019, 29, R42–R44. [Google Scholar] [CrossRef] [Green Version]
- Gowda, S.; Desai, P.B.; Hull, V.V.; Avinash, A.K.; Vernekar, S.N.; Kulkarni, S.S. A review on laboratory liver function tests. Pan Afr. Med. J. 2009, 3, 17. [Google Scholar]
- Mehlem, A.; Hagberg, C.E.; Muhl, L.; Eriksson, U.; Falkevall, A. Imaging of neutral lipids by oil red O for analyzing the metabolic status in health and disease. Nat. Protoc. 2013, 8, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Stal, P. Liver fibrosis in non-alcoholic fatty liver disease—Diagnostic challenge with prognostic significance. World J. Gastroenterol. 2015, 21, 11077–11087. [Google Scholar] [CrossRef]
- Ma, L.; Yang, X.; Wei, R.; Ye, T.; Zhou, J.-K.; Wen, M.; Men, R.; Li, P.; Dong, B.; Liu, L. MicroRNA-214 promotes hepatic stellate cell activation and liver fibrosis by suppressing Sufu expression. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Zaldivar, M.M.; Pauels, K.; von Hundelshausen, P.; Berres, M.L.; Schmitz, P.; Bornemann, J.; Kowalska, M.A.; Gassler, N.; Streetz, K.L.; Weiskirchen, R. CXC chemokine ligand 4 (Cxcl4) is a platelet-derived mediator of experimental liver fibrosis. Hepatology 2010, 51, 1345–1353. [Google Scholar] [CrossRef]
- Brunt, E.M.; Kleiner, D.E.; Wilson, L.A.; Belt, P.; Neuschwander-Tetri, B.A.; Network, N.C.R. Nonalcoholic fatty liver disease (NAFLD) activity score and the histopathologic diagnosis in NAFLD: Distinct clinicopathologic meanings. Hepatology 2011, 53, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Peverill, W.; Powell, L.W.; Skoien, R. Evolving concepts in the pathogenesis of NASH: Beyond steatosis and inflammation. Int. J. Mol. Sci. 2014, 15, 8591–8638. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Varghese, Z.; Ruan, X.Z. The molecular pathogenic role of inflammatory stress in dysregulation of lipid homeostasis and hepatic steatosis. Genes Dis. 2014, 1, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Xia, H. TRIM Proteins in Inflammation: From Expression to Emerging Regulatory Mechanisms. Inflammation 2021, 44, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Tacke, F. Liver macrophages: Old dogmas and new insights. Hepatol. Commun. 2019, 3, 730–743. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tang, T.; Wang, G.-D.; Liu, B. LncRNA-H19 promotes hepatic lipogenesis by directly regulating miR-130a/PPARγ axis in non-alcoholic fatty liver disease. Biosci. Rep. 2019, 39, BSR20181722. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Carracedo, A.; Weiss, D.; Arai, F.; Ala, U.; Avigan, D.E.; Schafer, Z.T.; Evans, R.M.; Suda, T.; Lee, C.-H. A PML–PPAR-δ pathway for fatty acid oxidation regulates hematopoietic stem cell maintenance. Nat. Med. 2012, 18, 1350–1358. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-F.; Ku, H.-C.; Lin, H. PGC-1α as a pivotal factor in lipid and metabolic regulation. Int. J. Mol. Sci. 2018, 19, 3447. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.-H.; Satoh, H.; Herzig, S.; Lee, C.-H.; Hedrick, S.; Kulkarni, R.; Evans, R.M.; Olefsky, J.; Montminy, M. PGC-1 promotes insulin resistance in liver through PPAR-α-dependent induction of TRB-3. Nat. Med. 2004, 10, 530–534. [Google Scholar] [CrossRef]
- Lin, H.; Sharabi, K.; Lin, L.; Ruiz, C.; Zhu, D.; Cameron, M.D.; Novick, S.J.; Griffin, P.R.; Puigserver, P.; Kamenecka, T.M. Structure–Activity Relationship and Biological Investigation of SR18292 a Suppressor of Glucagon-Induced Glucose Production. J. Med. Chem. 2021, 64, 980–990. [Google Scholar] [CrossRef]
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metab. Clin. Exp. 2019, 92, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, D.W.; Yan, H.Y.; Wang, Z.Y.; Zhao, S.H.; Wang, B. Obesity is an independent risk factor for non-alcoholic fatty liver disease: Evidence from a meta-analysis of 21 cohort studies. Obes. Rev. 2016, 17, 510–519. [Google Scholar] [CrossRef]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metab. Clin. Exp. 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.; Tianyu, W.; Jiangqiao, Z.; Zhongbao, C.; Xiaoxiong, M.; Long, Z.; Jilin, Z. Tripartite motif 8 deficiency relieves hepatic ischaemia/reperfusion injury via TAK1-dependent signalling pathways. Int. J. Biol. Sci. 2019, 15, 1618. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Minter, L.C.; Stratton, S.A.; Yang, P.; Abbas, H.A.; Akdemir, Z.C.; Pant, V.; Post, S.; Gagea, M.; Lee, R.G.; et al. TRIM24 suppresses development of spontaneous hepatic lipid accumulation and hepatocellular carcinoma in mice. J. Hepatol. 2015, 62, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Piccinin, E.; Villani, G.; Moschetta, A. Metabolic aspects in NAFLD, NASH and hepatocellular carcinoma: The role of PGC1 coactivators. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 160–174. [Google Scholar] [CrossRef]
- Ropelle, E.R.; Pauli, J.R.; Cintra, D.E.; Frederico, M.J.; De Pinho, R.A.; Velloso, L.A.; De Souza, C.T. Acute exercise modulates the Foxo1/PGC-1α pathway in the liver of diet-induced obesity rats. J. Physiol. 2009, 587, 2069–2076. [Google Scholar] [CrossRef]
- Barbuio, R.; Milanski, M.; Bertolo, M.B.; Saad, M.J.; Velloso, L.A. Infliximab reverses steatosis and improves insulin signal transduction in liver of rats fed a high-fat diet. J. Endocrinol. 2007, 194, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, H.; Kiyono, S.; Kondo, T.; Sekimoto, T.; Yokosuka, O. Palmitate-induced regulation of PPARγ via PGC1α: A mechanism for lipid accumulation in the liver in nonalcoholic fatty liver disease. Int. J. Med. Sci. 2016, 13, 169. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.; Joe, Y.; Zheng, M.; Kim, S.; Lee, H.; Kwon, T.-O.; Chung, H.T. Inhibitory effects of chung hun wha dam tang (CHWDT) on high-fat diet-induced obesity via AMP-activated protein kinase activation. Evid. Based Complement. Altern. Med. 2012, 2012, 652473. [Google Scholar] [CrossRef]
- Kristensen, C.M.; Dethlefsen, M.M.; Tøndering, A.S.; Lassen, S.B.; Meldgaard, J.N.; Ringholm, S.; Pilegaard, H. PGC-1α in hepatic UPR during high-fat high-fructose diet and exercise training in mice. Physiol. Rep. 2018, 6, e13819. [Google Scholar] [CrossRef]
- Piao, L.; Choi, J.; Kwon, G.; Ha, H. Endogenous catalase delays high-fat diet-induced liver injury in mice. Korean J. Physiol. Pharmacol. 2017, 21, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb-prot5439. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| Mouse β-actin | F | 5′-AGAGGGAAATCGTGCGTGAC-3′ |
| R | 5′-CAATAGTGATGACCTGGCCGT-3′ | |
| Mouse TRIM67 | F | 5′-GGCGAAGGAGTTTCTGGTTC-3′ |
| R | 5′-TAGCTTCAGGGTGCAGTGATT-3′ | |
| Mouse IL-1β | F | 5′-CCCCAGGGCATGTTAAGGAG-3′ |
| R | 5′-TCTTGGCCGAGGACTAAGGA-3′ | |
| Mouse IL-6 | F | 5′-CTTCCATCCAGTTGCCTTCTTG-3′ |
| R | 5′-AATTAAGCCTCCGACTTGTGAAG-3′ | |
| Mouse TNF-α | F | 5′-ACGGCATGGATCTCAAAGAC-3′ |
| R | 5′-GTGGGTGAGGAGCACGTAG-3′ | |
| Mouse ACC1 | F | 5′-ATTGTGGCTCAAACTGCAGGT-3′ |
| R | 5′-GCCAATCCACTCGAAGACCA-3′ | |
| Mouse SCD1 | F | 5′-CACACCTTCCCCTTCGACTAC-3′ |
| R | 5′-GAAACAGGAACTCAGAAGCCCA-3′ | |
| Mouse PPAR-α | F | 5′-AACATCGAGTGTCGAATATGTGG-3′ |
| R | 5′-CCGAATAGTTCGCCGAAAGAA-3′ | |
| Mouse Cpt-1α | F | 5′-TGGCATCATCACTGGTGTGTT-3′ |
| R | 5′-GTCTAGGGTCCGATTGATCTTTG-3′ | |
| Mouse PGC-1α | F | 5′-TATGGAGTGACATAGAGTGTGCT-3′ |
| R | 5′-CCACTTCAATCCACCCAGAAAG-3′ | |
| Mouse α-SMA | F | 5′-CGCTGCTCCAGCTATGTGTGA-3′ |
| R | 5′-TTTGGCCCATTCCAACCATTAC-3′ | |
| Human ACC1 | F | 5′-AGTGAGGATGGCAGCTCTGTCTC-3′ |
| R | 5′-TGAGATGTGGGCAGCATGAAC-3′ | |
| Human SCD1 | F | 5′-GCAGGACGATATCTCTAGCT-3′ |
| R | 5′-GTCTCCAACTTATCTCCTCCATTC-3′ | |
| Human PPAR-α | F | 5′-CAATGCACTGGAACTGGATGA-3′ |
| R | 5′-GTTGCTCTGCAGGTGGAGTCT-3′ | |
| Human CPT-1α | F | 5′-CGTCTTTTGGGATCCACGATT-3′ |
| R | 5′-TGTGCTGGATGGTGTCTGTCTC-3′ | |
| Human PGC-1α | F | 5′-TGAAGGGTACTTTTCTGCCCC-3′ |
| R | 5′-TCACTGCACCACTTGAGTCC-3′ | |
| Human TRIM67 | F | 5′-AAACGGACTGGACTACGAA-3′ |
| R | 5′-ATCTGGTCCCAAACCATCTT-3′ |
| Antibodies | Source | Catalog No./Dilution |
|---|---|---|
| IL-1β Rabbit pAb | Bioss | bs-6319R/WB 1:1000 |
| IL-6 Rabbit pAb | ABclonal | A0286/WB 1:1000 |
| PGC1α mouse mAb | SANTA | sc-518025/WB 1:1000 |
| F4/80 Rabbit pAb | Bioss | bs11182R/IHC 1:500 |
| α-SMA Rabbit mAb | HuaBio | ET1607-53/WB 1:1000 IHC 1:200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Wei, X.; Luo, Q.; Xia, Y.; Pan, T.; He, J.; Jahangir, A.; Jia, L.; Liu, W.; Zou, Y.; et al. Loss of TRIM67 Attenuates the Progress of Obesity-Induced Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 7475. https://doi.org/10.3390/ijms23137475
Huang C, Wei X, Luo Q, Xia Y, Pan T, He J, Jahangir A, Jia L, Liu W, Zou Y, et al. Loss of TRIM67 Attenuates the Progress of Obesity-Induced Non-Alcoholic Fatty Liver Disease. International Journal of Molecular Sciences. 2022; 23(13):7475. https://doi.org/10.3390/ijms23137475
Chicago/Turabian StyleHuang, Chao, Xiaoli Wei, Qihui Luo, Yu Xia, Ting Pan, Junbo He, Asad Jahangir, Lanlan Jia, Wentao Liu, Yuanfeng Zou, and et al. 2022. "Loss of TRIM67 Attenuates the Progress of Obesity-Induced Non-Alcoholic Fatty Liver Disease" International Journal of Molecular Sciences 23, no. 13: 7475. https://doi.org/10.3390/ijms23137475