
Leafamine®, a Free Amino Acid-Rich Biostimulant, Promotes Growth Performance of Deficit-Irrigated Lettuce

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of Leafamine® on Shoot and Root Biomass
2.2. Effect of Leafamine® on Morphological Parameters
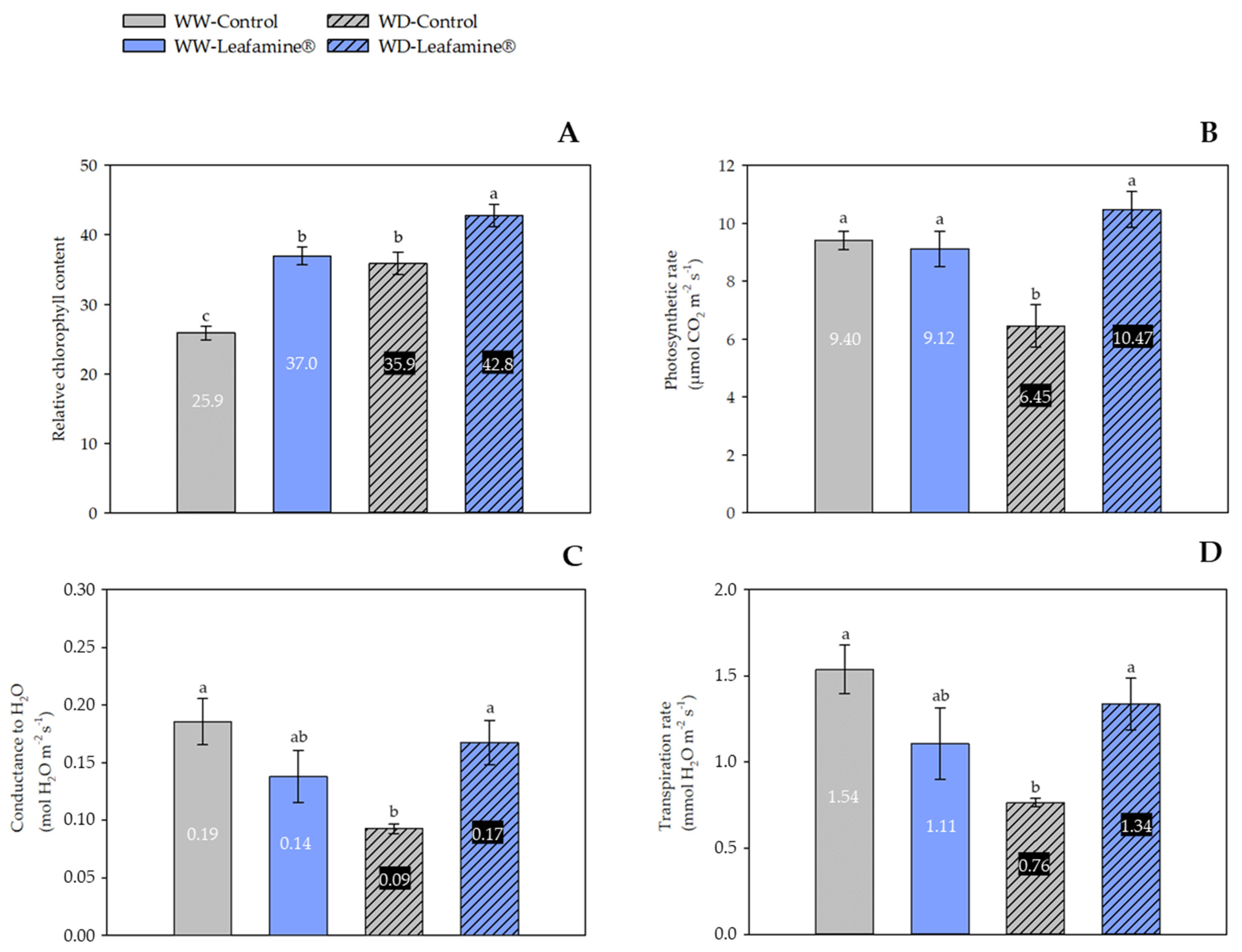
2.3. Effect of Leafamine® on Leaf Gas Exchange Parameters
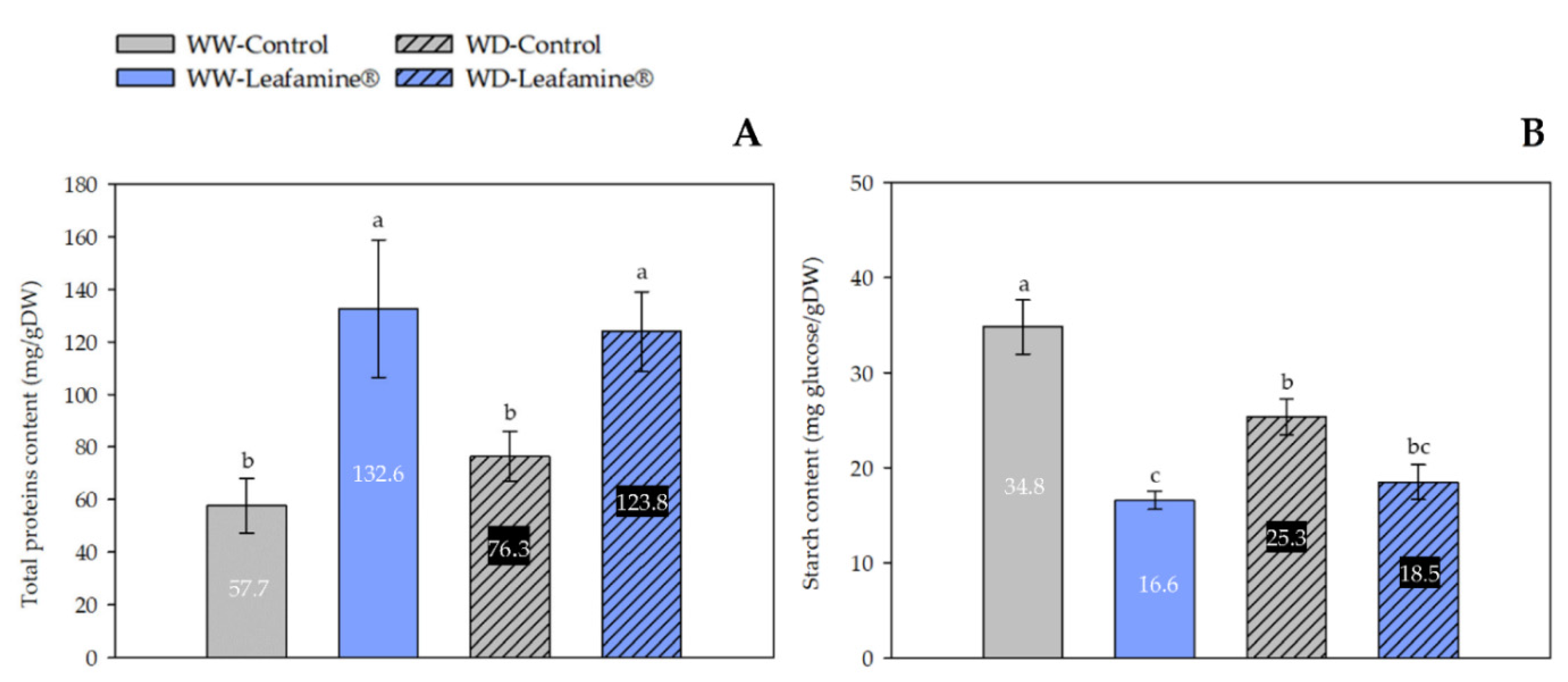
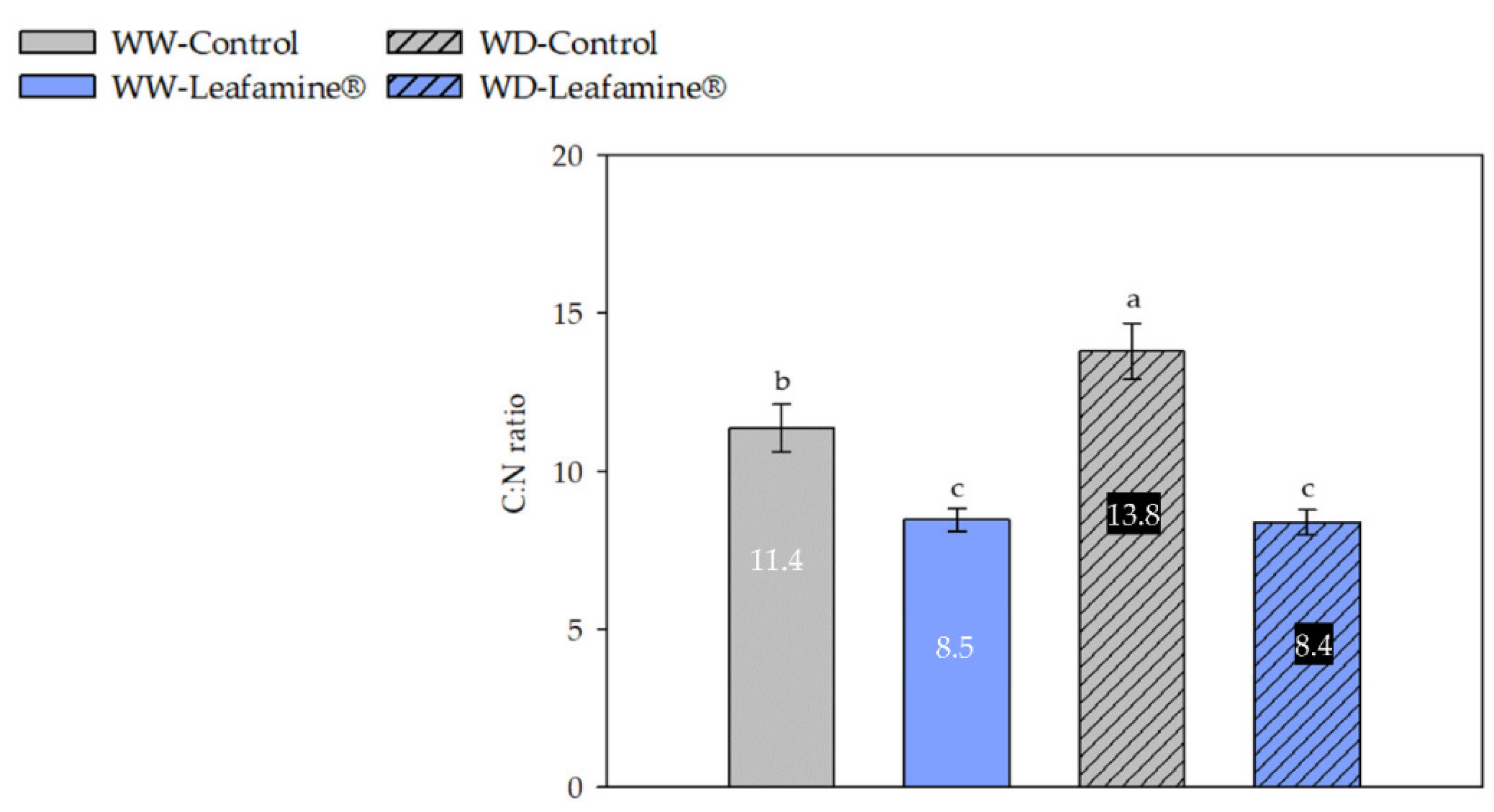
2.4. Effect of Leafamine® on the Leaf Carbon, Nitrogen and Sulfur Contents
2.5. Leafamine® on Leaf Defense and Growth Phytohormones
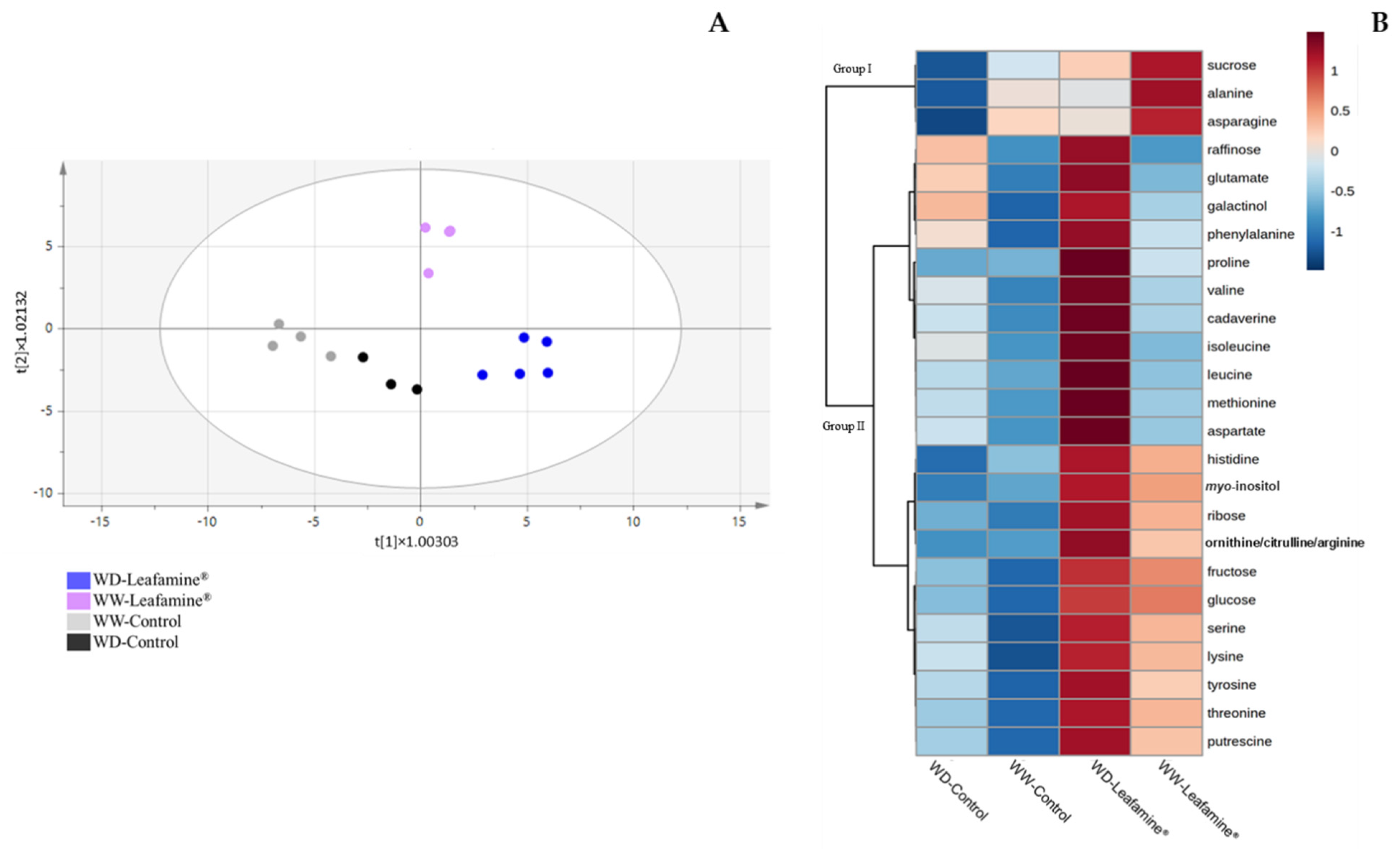
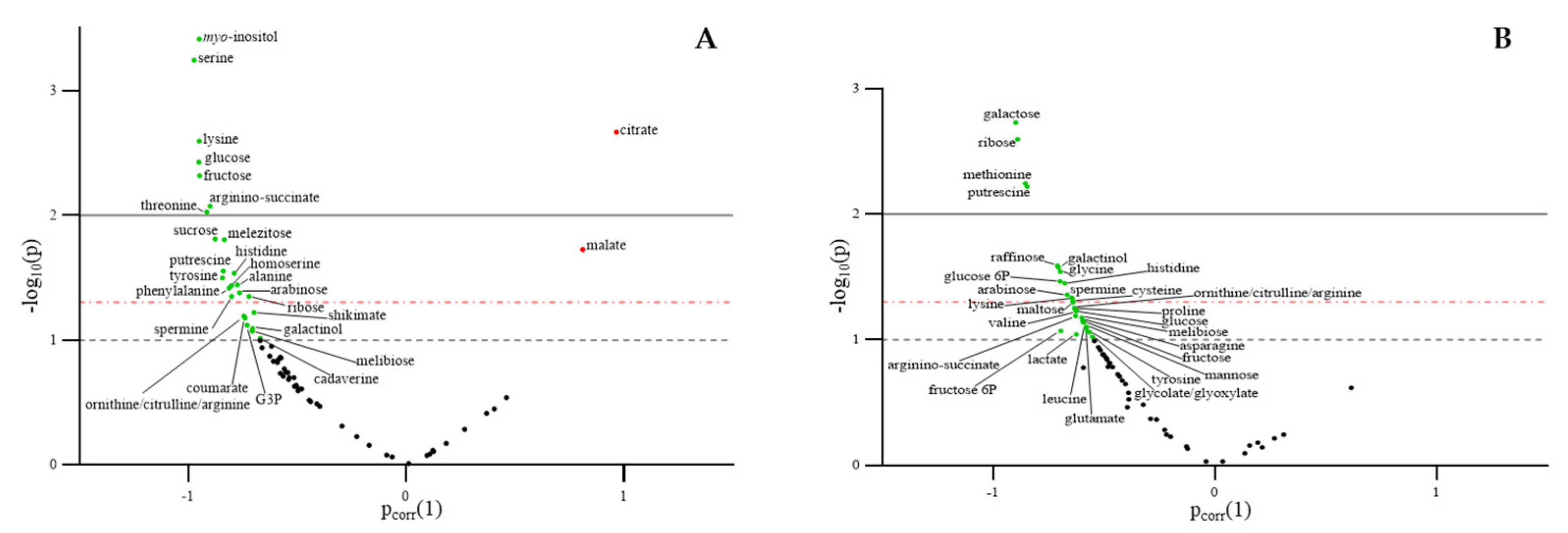
2.6. Effect of Leafamine® on Metabolomic Profiling
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions and Harvesting Procedure
4.2. Elemental C, N and S Content Measurements
4.3. Gas Exchange and Chlorophyll Content Measurements
4.4. Starch Content Measurement
4.5. Total Protein Content Measurement
4.6. Phytohormone Content Measurements
4.7. Metabolomics Analysis
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry, 1st ed.; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–16. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Kaur, H.; Kohli, S.K.; Khanna, K.; Bhardwaj, R. Scrutinizing the impact of water deficit in plants: Transcriptional regulation, signaling, photosynthetic efficacy, and management. Physiol. Plant. 2021, 172, 935–962. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Cifre, J.; Escalona, J.M.; Galmés, J.; Gulias, J.; Lefi, E.; Martinez-Canellas, S.F.; Moreno, M.T.; Ribas-Carbo, M.; et al. Understanding down-regulation of photosynthesis under water stress: Future prospects and searching for physiological tools for irrigation management. Ann. Appl. Biol. 2004, 144, 273–283. [Google Scholar] [CrossRef]
- Bradford, K.J.; Hsiao, T.C. Physiological Responses to Moderate Water Stress. In Physiological Plant Ecology II Water Relations and Carbon Assimilation, 1st ed.; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 263–324. [Google Scholar] [CrossRef]
- Salvi, P.; Manna, M.; Kaur, H.; Thakur, T.; Gandass, N.; Bhatt, D.; Muthamilarasan, M. Phytohormone signaling and crosstalk in regulating drought stress response in plants. Plant Cell Rep. 2021, 40, 1305–1329. [Google Scholar] [CrossRef]
- Wei, L.; Wang, L.; Eyang, Y.; Wang, P.; Guo, T.; Kang, G. Abscisic acid enhances tolerance of wheat seedlings to drought and regulates transcript levels of genes encoding ascorbate-glutathione biosynthesis. Front. Plant Sci. 2015, 6, 458. [Google Scholar] [CrossRef]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible Solute Engineering in Plants for Abiotic Stress Tolerance—Role of Glycine Betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, M.; Unal, B.T.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Jiménez-Arias, D.; García-Machado, F.J.; Morales-Sierra, S.; García-García, A.L.; Herrera, A.J.; Valdés, F.; Luis, J.C.; Borges, A.A. A Beginner’s Guide to Osmoprotection by Biostimulants. Plants 2021, 10, 363. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M.; Olsovska, K.; Kalaji, H.M.; Shao, H.; Hakeem, K.R. Heat Signaling and Stress Responses in Photosynthesis. In Plant Signaling: Understanding the Molecular Crosstalk, 1st ed.; Hakeem, K., Rehman, R., Tahir, I., Eds.; Springer: New Delhi, India, 2014; pp. 241–256. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic Adjustment and Plant Adaptation to Drought Stress. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry, 1st ed.; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 105–143. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. The contribution of carbohydrates including raffinose family oligosaccharides and sugar alcohols to protection of plant cells from oxidative damage. Plant Signal. Behav. 2008, 3, 1016–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and Raffinose Constitute a Novel Function to Protect Plants from Oxidative Damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [Green Version]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, S.; Mukherjee, S.; Basak, P.; Majumder, A.L. Significance of galactinol and raffinose family oligosaccharide synthesis in plants. Front. Plant Sci. 2015, 6, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef]
- Kamiab, F.; Tavassolian, I.; Hosseinifarahi, M. Biologia futura: The role of polyamine in plant science. Biol. Futur. 2020, 71, 183–194. [Google Scholar] [CrossRef]
- Nayyar, H.; Chander, S. Protective Effects of Polyamines against Oxidative Stress Induced by Water and Cold Stress in Chickpea. J. Agron. Crop Sci. 2004, 190, 355–365. [Google Scholar] [CrossRef]
- Kubiś, J. Polyamines and “scavenging system”: Influence of exogenous spermidine on catalase and guaiacol peroxidase activities, and free polyamine level in barley leaves under water deficit. Acta Physiol. Plant. 2003, 25, 337–343. [Google Scholar] [CrossRef]
- Shen, W.; Nada, K.; Tachibana, S. Involvement of Polyamines in the Chilling Tolerance of Cucumber Cultivars. Plant Physiol. 2000, 124, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galston, A.W.; Kaur-Sawhney, R. Polyamines as Endogenous Growth Regulators. In Plant Hormones, Physiology, Biochemistry and Molecular Biology, 1st ed.; Davies, P.J., Ed.; Springer: Dordrecht, Holland, 1995. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Lucini, L.; Canaguier, R.; Stefanoni, W.; Fiorillo, A.; Cardarelli, M. Protein hydrolysate-based biostimulants: Origin, biological activity and application methods. Acta Hortic. 2016, 1148, 27–34. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Baptistella, J.L.C.; Horz, D.C.; Bortolato, L.M.; Mazzafera, P. Organic Plant Biostimulants and Fruit Quality—A Review. Agronomy 2020, 10, 988. [Google Scholar] [CrossRef]
- Gurav, R.G.; Jadhav, J.P. A novel source of biofertilizer from feather biomass for banana cultivation. Environ. Sci. Pollut. Res. 2013, 20, 4532–4539. [Google Scholar] [CrossRef]
- Jiménez-Arias, D.; García-Machado, F.J.; Morales-Sierra, S.; Luis, J.C.; Suarez, E.; Hernández, M.; Valdés, F.; Borges, A.A. Lettuce plants treated with L-pyroglutamic acid increase yield under water deficit stress. Environ. Exp. Bot. 2019, 158, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein Hydrolysate or Plant Extract-based Biostimulants Enhanced Yield and Quality Performances of Greenhouse Perennial Wall Rocket Grown in Different Seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; El Nakhel, C.; Leone, V.; Mori, M. Effect of seaweed (Ecklonia maxima) extract and legume-derived protein hydrolysate biostimulants on baby leaf lettuce grown on optimal doses of nitrogen under greenhouse conditions. Aust. J. Crop Sci. 2020, 14, 1456–1464. [Google Scholar] [CrossRef]
- Kizil, U.; Genc, L.; Inalpulat, M.; Sapolyo Ucan, D.; Mirik, M. Lettuce (Lactuca sativa L.) yield prediction under water stress using artificial neural network (ANN) model and vegetation indices. Zemdirb. = Agric. 2012, 99, 409–418. [Google Scholar]
- Vinson, C.C.; Mota, A.P.Z.; Porto, B.N.; Oliveira, T.N.; Sampaio, I.; Lacerda, A.L.; Danchin, E.G.J.; Guimaraes, P.M.; Williams, T.C.R.; Brasileiro, A.C.M. Characterization of raffinose metabolism genes uncovers a wild Arachis galactinol synthase conferring tolerance to abiotic stresses. Sci. Rep. 2020, 10, 15258. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Rouphael, Y. Biostimulant Substances for Sustainable Agriculture: Origin, Operating Mechanisms and Effects on Cucurbits, Leafy Greens, and Nightshade Vegetables Species. Biomolecules 2021, 11, 1103. [Google Scholar] [CrossRef]
- Bhatnagar, P.; Minocha, R.; Minocha, S.C. Genetic Manipulation of the Metabolism of Polyamines in Poplar Cells. The Regulation of Putrescine Catabolism. Plant Physiol. 2002, 128, 1455–1469. [Google Scholar] [CrossRef] [Green Version]
- Page, A.F.; Cseke, L.J.; Minocha, R.; Turlapati, S.A.; Podila, G.K.; Ulanov, A.; Li, Z.; Minocha, S.C. Genetic manipulation of putrescine biosynthesis reprograms the cellular transcriptome and the metabolome. BMC Plant Biol. 2016, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Elbar, O.H.A.; Farag, R.; Shehata, S.A. Effect of putrescine application on some growth, biochemical and anatomical characteristics of Thymus vulgaris L. under drought stress. Ann. Agric. Sci. 2019, 64, 129–137. [Google Scholar] [CrossRef]
- Mirza, J.I.; Bagni, N. Effects of exogenous polyamines and difluoromethylornithine on seed germination and root growth of Arabidopsis thaliana. Plant Growth Regul. 1991, 10, 163–168. [Google Scholar] [CrossRef]
- Sánchez-Rangel, D.; Chávez-Martínez, A.I.; Rodríguez-Hernández, A.A.; Maruri-López, I.; Urano, K.; Shinozaki, K.; Jiménez-Bremont, J.F. Simultaneous Silencing of Two Arginine Decarboxylase Genes Alters Development in Arabidopsis. Front. Plant Sci. 2016, 7, 300. [Google Scholar] [CrossRef] [Green Version]
- Hummel, I.; Couée, I.; El Amrani, A.; Martin-Tanguy, J.; Hennion, F. Involvement of polyamines in root development at low temperature in the subantarctic cruciferous species Pringlea antiscorbutica. J. Exp. Bot. 2002, 53, 1463–1473. [Google Scholar] [CrossRef] [Green Version]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.-L.; Wang, Y.; Shu, S.; Jahan, M.S.; Zhong, M.; Wu, J.-Q.; Sun, J.; Guo, S.-R. Exogenous putrescine regulates leaf starch overaccumulation in cucumber under salt stress. Sci. Hortic. 2019, 253, 99–110. [Google Scholar] [CrossRef]
- Silveira, N.M.; Ribeiro, R.V.; de Morais, S.F.; de Souza, S.C.; da Silva, S.F.; Seabra, A.B.; Hancock, J.T.; Machado, E.C. Leaf arginine spraying improves leaf gas exchange under water deficit and root antioxidant responses during the recovery period. Plant Physiol. Biochem. 2021, 162, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the Biostimulant Action of Vegetal-Derived Protein Hydrolysates by High-Throughput Plant Phenotyping and Metabolomics: A Case Study on Tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Ye, J.; Wang, B.; Ren, J.; Yin, L.; Deng, X.; Wang, S. Melatonin Mitigates Salt Stress in Wheat Seedlings by Modulating Polyamine Metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irani, H.; ValizadehKaji, B.; Naeini, M.R. Biostimulant-induced drought tolerance in grapevine is associated with physiological and biochemical changes. Chem. Biol. Technol. Agric. 2021, 8, 5. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Huang, Y.; Xian, W.; Wang, J.; Liao, H. Identification and expression analysis of the Glycine max CYP707A gene family in response to drought and salt stresses. Ann. Bot. 2012, 110, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef] [PubMed]
- Espasandin, F.D.; Maiale, S.J.; Calzadilla, P.; Ruiz, O.A.; Sansberro, P.A. Transcriptional regulation of 9-cis-epoxycarotenoid dioxygenase (NCED) gene by putrescine accumulation positively modulates ABA synthesis and drought tolerance in Lotus tenuis plants. Plant Physiol. Biochem. 2014, 76, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.H.; Li, J.; Guo, S.R.; Tezuka, T. Effects of exogenous putrescine on gas-exchange characteristics and chlorophyll fluorescence of NaCl-stressed cucumber seedlings. Photosynth. Res. 2009, 100, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Ebeed, H.T.; Hassan, N.M.; Aljarani, A.M. Exogenous applications of Polyamines modulate drought responses in wheat through osmolytes accumulation, increasing free polyamine levels and regulation of polyamine biosynthetic genes. Plant Physiol. Biochem. 2017, 118, 438–448. [Google Scholar] [CrossRef]
- Verslues, P.E.; Bray, E.A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. J. Exp. Bot. 2005, 57, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Ozfidan, C.; Turkan, I.; Sekmen, A.H.; Seckin, B. Time course analysis of ABA and non-ionic osmotic stress-induced changes in water status, chlorophyll fluorescence and osmotic adjustment in Arabidopsis thaliana wild-type (Columbia) and ABA-deficient mutant (aba2). Environ. Exp. Bot. 2013, 86, 44–51. [Google Scholar] [CrossRef]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Liu, J.J.; Odegard, W.; De Lumen, B.O. Galactinol Synthase from Kidney Bean Cotyledon and Zucchini Leaf (Purification and N-Terminal Sequences). Plant Physiol. 1995, 109, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Stick, R.V.; Williams, S.J. Monosaccharide Metabolism. In Carbohydrates: The Essential Molecules of Life, 2nd ed.; Stick, R.V., Williams, S.J., Eds.; Elsevier: Oxford, UK, 2009; pp. 225–251. [Google Scholar]
- Damiano, C.; Arias, P.M.D.; La Starza, S.R.; Frattarelli, A. Temporary immersion system for temperate fruit trees. Acta Hortic. 2007, 748, 87–90. [Google Scholar] [CrossRef]
- Ma, W.T.; Tcherkez, G.; Wang, X.M.; Schäufele, R.; Schnyder, H.; Yang, Y.; Gong, X.Y. Accounting for mesophyll conductance substantially improves 13 C-based estimates of intrinsic water-use efficiency. New Phytol. 2021, 229, 1326–1338. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic Composition of Plant Carbon Correlates with Water-Use Efficiency of Wheat Genotypes. Funct. Plant Biol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Ubierna, N.; Gandin, A.; Cousins, A.B. The response of mesophyll conductance to short-term variation in CO2 in the C4 plants Setaria viridis and Zea mays. J. Exp. Bot. 2018, 69, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Corot, A.; Roman, H.; Douillet, O.; Autret, H.; Perez-Garcia, M.-D.; Citerne, S.; Bertheloot, J.; Sakr, S.; LeDuc, N.; Demotes-Mainard, S. Cytokinins and Abscisic Acid Act Antagonistically in the Regulation of the Bud Outgrowth Pattern by Light Intensity. Front. Plant Sci. 2017, 8, 1724. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, C.; Del Prete, S.; Boutet-Mercey, S.; Perreau, F.; Balagué, C.; Roby, D.; Fagard, M.; Gaudin, V. The hnRNP-Q Protein LIF2 Participates in the Plant Immune Response. PLoS ONE 2014, 9, e99343. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Shoot Fresh Biomass (g Plant−1) | Root Fresh Biomass (g Plant−1) | Shoot: Root (Fresh biomass) | Shoot Dry Biomass (g Plant−1) | Root Dry Biomass (g Plant−1) | Shoot: Root (Dry biomass) | Water Content (%) |
|---|---|---|---|---|---|---|---|
| Treatment | |||||||
| Control | 57.7 ± 1.4 | 12.1 ± 0.6 | 6.4 ± 0.4 | 5.0 ± 0.2 | 1.2 ± 0.1 | 4.0 ± 0.2 | 92.3 ± 0.2 |
| [100] | [100] | [100] | [100] | [100] | [100] | [100] | |
| Leafamine® | 75.5 ± 2.4 | 9.6 ± 0.5 | 10.1 ± 0.4 | 5.6 ± 0.3 | 0.9 ± 0.1 | 6.4 ± 0.3 | 93.3 ± 0.2 |
| [131] | [79] | [158] | [112] | [75] | [160] | [101] | |
| Significance | *** | *** | *** | * | *** | *** | * |
| n | n = 88 | n = 48 | n = 48 | n = 48 | n = 48 | n = 48 | n = 48 |
| Water condition | |||||||
| Well-watered (WW) | 79.3 ± 2.1 | 12.6 ± 0.6 | 8.1 ± 0.4 | 6.3 ± 0.2 | 1.2 ± 0.1 | 5.4 ± 0.3 | 93.1 ± 0.2 |
| [100] | [100] | [100] | [100] | [100] | [100] | [100] | |
| Water deficit (WD) | 53.9 ± 1.1 | 9.0 ± 0.4 | 8.4 ± 0.6 | 4.3 ± 0.2 | 0.9 ± 0.0 | 5.1 ± 0.2 | 92.5 ± 0.2 |
| [68] | [71] | [104] | [68] | [75] | [94] | [99] | |
| Significance | *** | *** | NS | *** | *** | NS | * |
| n | n = 88 | n = 48 | n = 48 | n = 48 | n = 48 | n = 48 | n = 48 |
| Treatment × Water condition | |||||||
| Control × WW | 66.2 ± 1.6 b | 14.0 ± 1.0 | 6.3 ± 0.5 | 5.8 ± 0.2 | 1.5 ± 0.1 | 3.8 ± 0.2 b | 92.5 ± 0.3 |
| [100] | [100] | [100] | [100] | [100] | [100] | [100] | |
| Leafamine® × WW | 92.4 ± 2.7 a | 11.3 ± 0.6 | 10.0 ± 0.5 | 6.8 ± 0.3 | 1.0 ± 0.1 | 6.8 ± 0.4 a | 93.8 ± 0.2 |
| [140] | [81] | [159] | [117] | [67] | [179] | [101] | |
| Control × WD | 49.0 ± 1.2 d | 10.1 ± 0.4 | 6.6 ± 0.7 | 4.2 ± 0.2 | 1.0 ± 0.0 | 4.2 ± 0.2 b | 92.2 ± 0.2 |
| [74] | [72] | [105] | [72] | [67] | [111] | [100] | |
| Leafamine® × WD | 58.6 ± 1.5 c | 7.8 ± 0.5 | 10.2 ± 0.7 | 4.5 ± 0.2 | 0.8 ± 0.1 | 6.0 ± 0.3 a | 92.9 ± 0.2 |
| [89] | [56] | [162] | [78] | [53] | [158] | [100] | |
| Significance | *** | NS | NS | NS | NS | * | NS |
| n | n = 44 | n = 24 | n = 24 | n = 24 | n = 24 | n = 24 | n = 24 |
| Factor | Projected Leaf Area (cm2) 21 DAT | Projected Leaf Area (cm2) 34 DAT |
|---|---|---|
| Treatment | ||
| Control | 755.4 ± 46.7 | 787.3 ± 51.3 |
| [100] | [100] | |
| Leafamine® | 790.5 ± 41.3 | 910.3 ± 70.5 |
| [105] | [116] | |
| Significance | NS | ** |
| n | n = 14 | n = 14 |
| Water condition | ||
| Well-watered (WW) | 897.0 ± 26.2 | 1021.6 ± 48.7 |
| [100] | [100] | |
| Water deficit (WD) | 640.8 ± 18.8 | 667.5 ± 22.6 |
| [71] | [65] | |
| Significance | *** | *** |
| n | n = 14 | n = 14 |
| Treatment × Water condition | ||
| Control × WW | 883.3 ± 33.4 | 930.5 ± 38.4 |
| [100] | [100] | |
| Leafamine® × WW | 910.7 ± 45.1 | 1112.7 ± 80.3 |
| [103] | [120] | |
| Control × WD | 606.3 ± 32.0 | 620.2 ± 23.7 |
| [69] | [67] | |
| Leafamine® × WD | 670.3 ± 18.3 | 708.0 ± 30.9 |
| [76] | [76] | |
| Significance | NS | NS |
| n | n = 7 | n = 7 |
| Factor | Carbon Content (% Dry Weight) | Nitrogen Content (% Dry Weight) | Sulphur Content (% Dry Weight) | Intrinsic Water-Use Efficiency (µmol mol−1) |
|---|---|---|---|---|
| Treatment | ||||
| Control | 40.18 ± 0.16 | 3.27 ± 0.19 | 0.29 ± 0.02 | 145.77 ± 6.96 |
| [100] | [100] | [100] | [100] | |
| Leafamine® | 39.71 ± 0.24 | 4.75 ± 0.13 | 0.38 ± 0.03 | 153.85 ± 9.38 |
| [99] | [145] | [131] | [106] | |
| Significance | NS | *** | * | NS |
| n | n = 10 | n = 10 | n = 10 | n = 10 |
| Water condition | ||||
| Well-watered (WW) | 40.10 ± 0.21 | 4.17 ± 0.25 | 0.37 ± 0.03 | 134.67 ± 5.36 |
| [100] | [100] | [100] | [100] | |
| Water deficit (WD) | 39.80 ± 0.22 | 3.85 ± 0.34 | 0.30 ± 0.02 | 164.95 ± 7.43 |
| [99] | [92] | [81] | [122] | |
| Significance | NS | NS | * | ** |
| n | n = 10 | n = 10 | n = 10 | n = 10 |
| Treatment × Water condition | ||||
| Control × WW | 40.45 ± 0.16 | 3.62 ± 0.25 | 0.31 ± 0.05 | 135.72 ± 9.27 |
| [100] | [100] | [100] | [100] | |
| Leafamine® × WW | 39.74 ± 0.35 | 4.73 ± 0.21 | 0.43 ± 0.05 | 133.62 ± 7.65 |
| [98] | [131] | [139] | [98] | |
| Control × WD | 39.92 ± 0.25 | 2.93 ± 0.19 | 0.27 ± 0.02 | 155.82 ± 9.80 |
| [99] | [81] | [87] | [115] | |
| Leafamine® × WD | 39.69 ± 0.41 | 4.77 ± 0.20 | 0.33 ± 0.02 | 174.08 ± 11.46 |
| [98] | [132] | [106] | [128] | |
| Significance | NS | NS | NS | NS |
| n | n = 5 | n = 5 | n = 5 | n = 5 |
| Factor | Endogenous Salicylic Acid (ng/gDW) | Endogenous Jasmonic Acid (ng/gDW) | Endogenous Abscisic Acid (ng/gDW) | Zeatin-o-Glucoside Ribose (ng/gDW) |
|---|---|---|---|---|
| Treatment | ||||
| Control | 417.4 ± 42.8 | 87.2 ± 19.8 | 977.4 ± 502.4 | 93.2 ± 2.8 |
| [100] | [100] | [100] | [100] | |
| Leafamine® | 776.4 ± 152.1 | 113.0 ± 49.3 | 2760.8 ± 798.1 | 143.4 ± 8.8 |
| [186] | [130] | [282] | [154] | |
| Significance | * | NS | *** | *** |
| n | n = 8 | n = 8 | n = 8 | n = 8 |
| Water condition | ||||
| Well-watered (WW) | 492.9 ± 61.5 | 95.2 ± 20.9 | 106.3 ± 51.0 | 101.6 ± 7.4 |
| [100] | [100] | [100] | [100] | |
| Water deficit (WD) | 701.0 ± 167.8 | 104.9 ± 49.3 | 3631.9 ± 362.8 | 135.0 ± 12.1 |
| [142] | [110] | [3417] | [133] | |
| Significance | NS | NS | *** | *** |
| n | n = 8 | n = 8 | n = 8 | n = 8 |
| Treatment × Water condition | ||||
| Control × WW | 421.1 ± 61.0 | 94.9 ± 28.6 | 20.4 ± 3.2 c | 90.5 ± 1.5 c |
| [100] | [100] | [100] | [100] | |
| Leafamine® × WW | 612.5 ± 119.6 | 95.8 ± 45.8 | 249.5 ± 85.2 c | 120.2 ± 15.8 b |
| [145] | [101] | [1223] | [133] | |
| Control × WD | 411.3 ± 87.0 | 74.3 ± 36.7 | 2572.4 ± 196.9 b | 97.7 ± 8.0 bc |
| [98] | [78] | [12610] | [108] | |
| Leafamine® × WD | 874.8 ± 243.3 | 123.3 ± 82.7 | 4267.6 ± 238.1 a | 157.4 ± 3.6 a |
| [208] | [130] | [20920] | [174] | |
| Significance | NS | NS | *** | * |
| n | n = 4 | n = 4 | n = 4 | n = 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malécange, M.; Pérez-Garcia, M.-D.; Citerne, S.; Sergheraert, R.; Lalande, J.; Teulat, B.; Mounier, E.; Sakr, S.; Lothier, J. Leafamine®, a Free Amino Acid-Rich Biostimulant, Promotes Growth Performance of Deficit-Irrigated Lettuce. Int. J. Mol. Sci. 2022, 23, 7338. https://doi.org/10.3390/ijms23137338
Malécange M, Pérez-Garcia M-D, Citerne S, Sergheraert R, Lalande J, Teulat B, Mounier E, Sakr S, Lothier J. Leafamine®, a Free Amino Acid-Rich Biostimulant, Promotes Growth Performance of Deficit-Irrigated Lettuce. International Journal of Molecular Sciences. 2022; 23(13):7338. https://doi.org/10.3390/ijms23137338
Chicago/Turabian StyleMalécange, Marthe, Maria-Dolores Pérez-Garcia, Sylvie Citerne, Renaud Sergheraert, Julie Lalande, Béatrice Teulat, Emmanuelle Mounier, Soulaiman Sakr, and Jérémy Lothier. 2022. "Leafamine®, a Free Amino Acid-Rich Biostimulant, Promotes Growth Performance of Deficit-Irrigated Lettuce" International Journal of Molecular Sciences 23, no. 13: 7338. https://doi.org/10.3390/ijms23137338